Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence


 ,
,  , and
, and 
Abstract
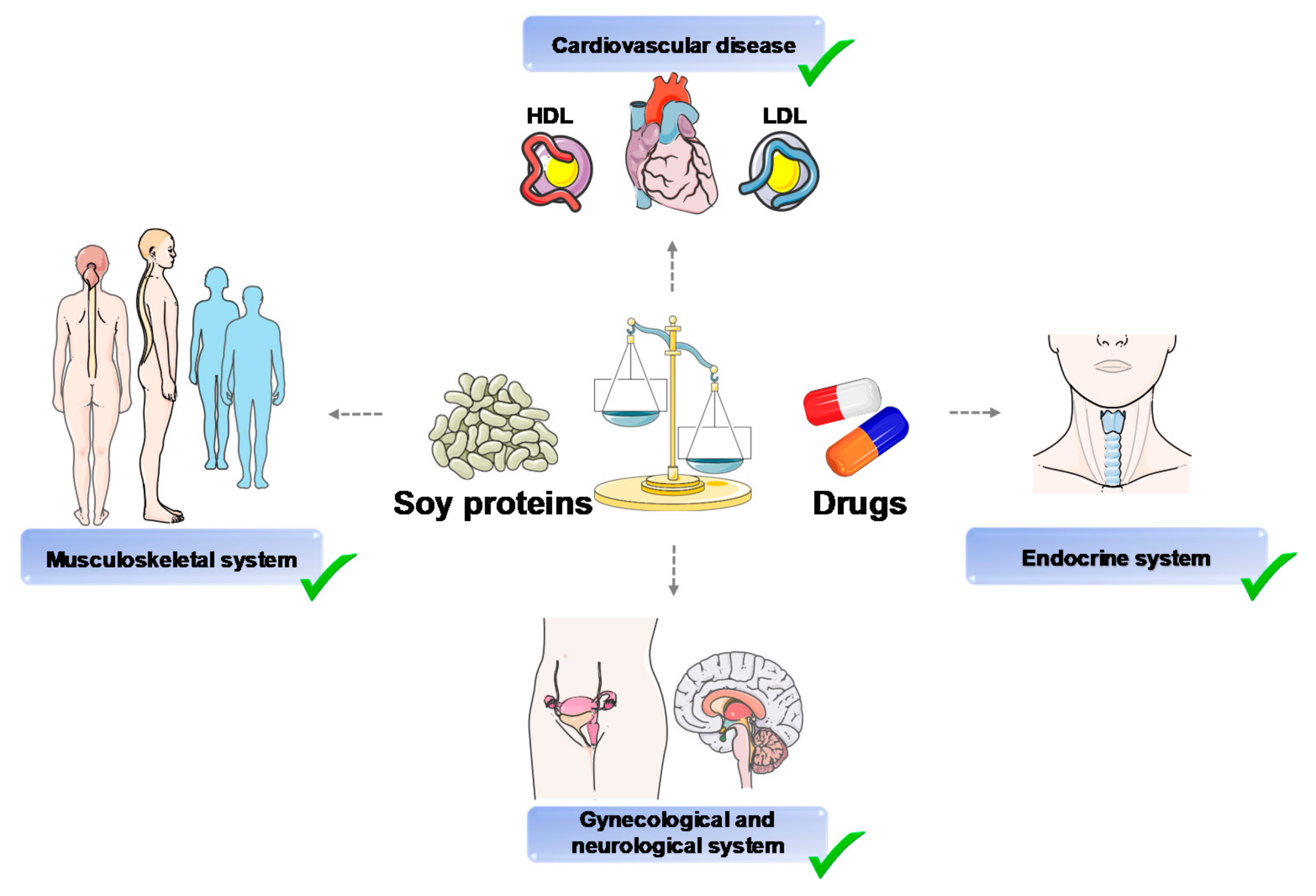
:1. Introduction
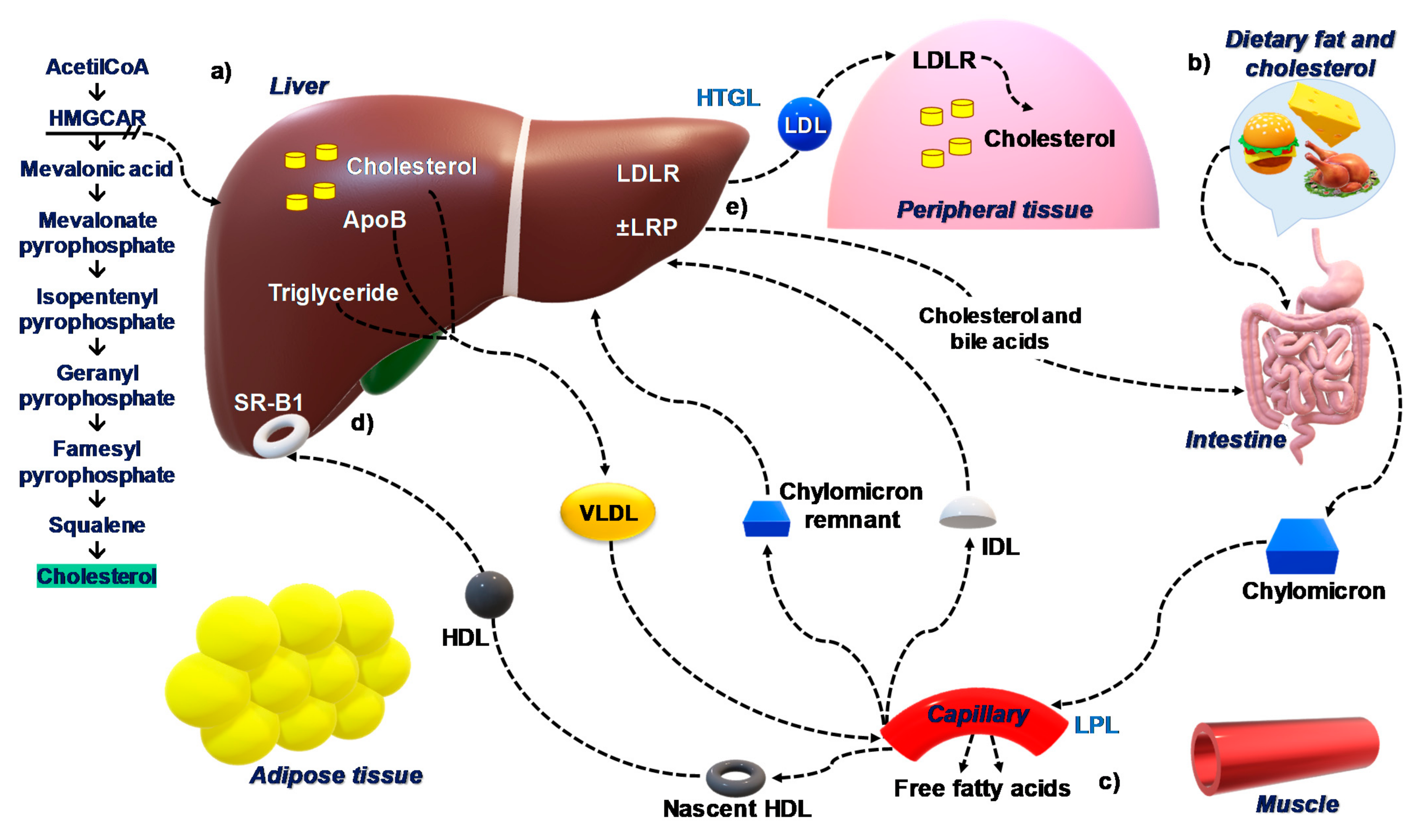
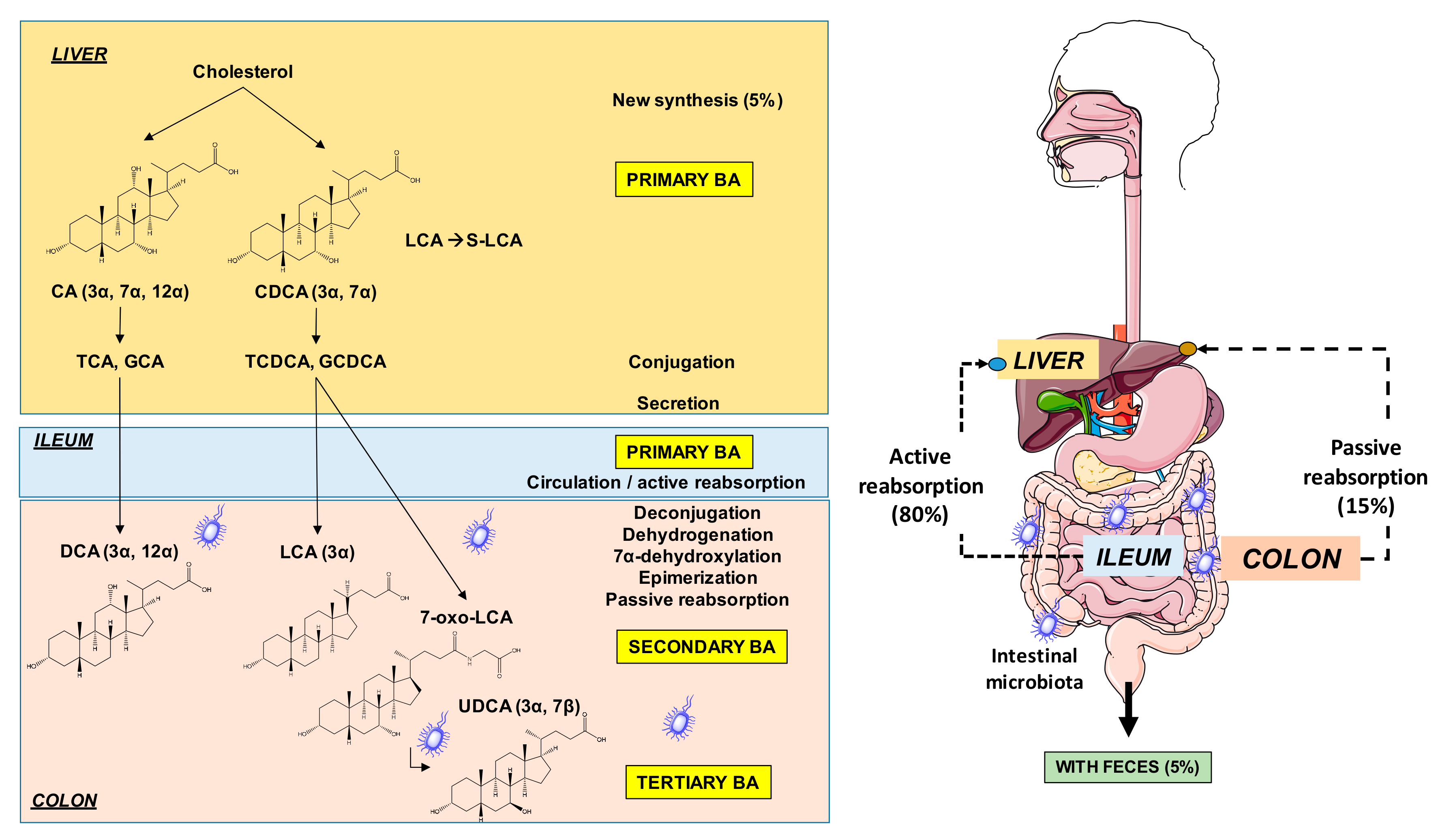
2. The Burden of Cardiovascular Diseases (CVD) as Components of the Metabolic Syndrome
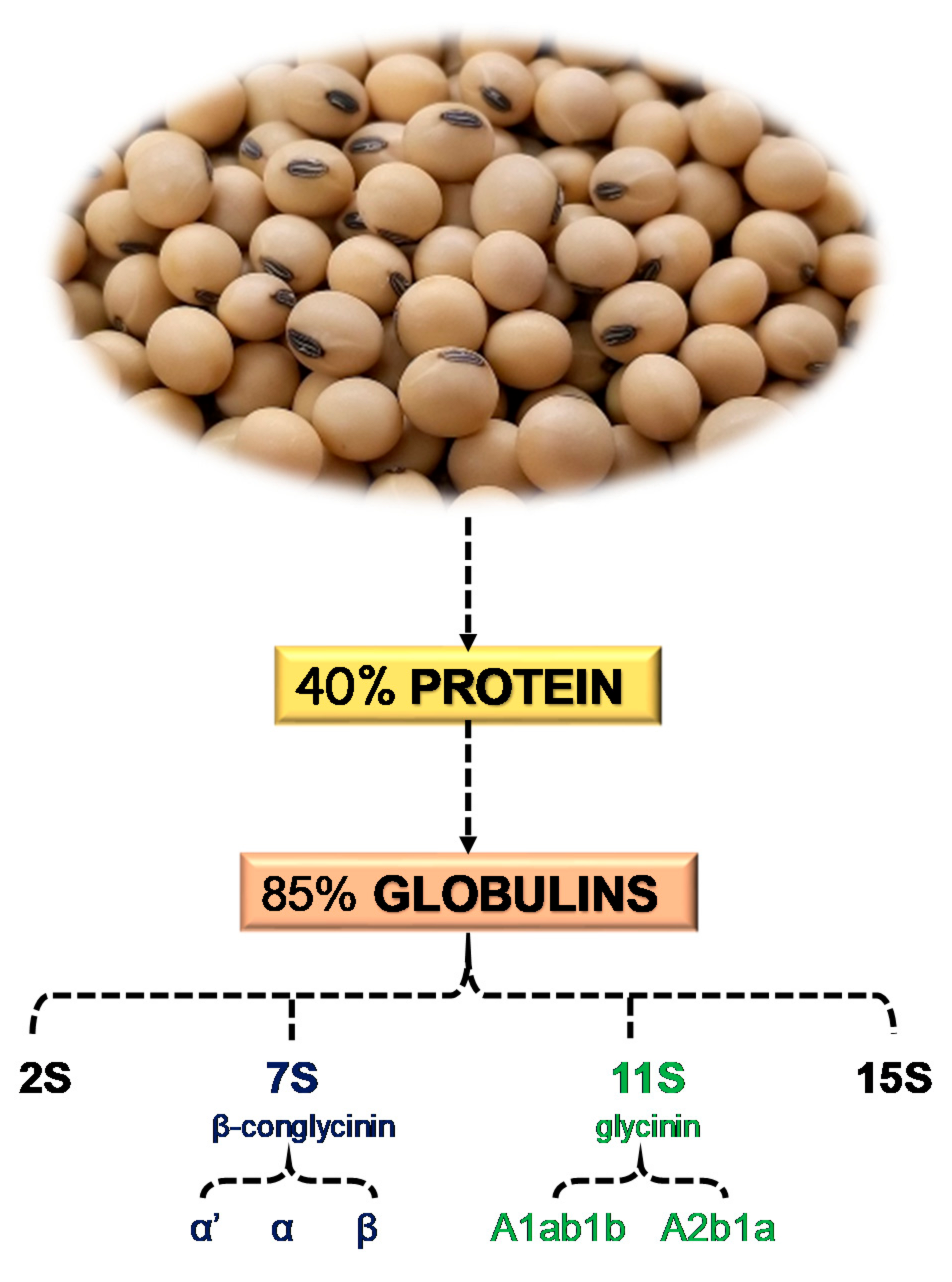
3. Soy Proteins
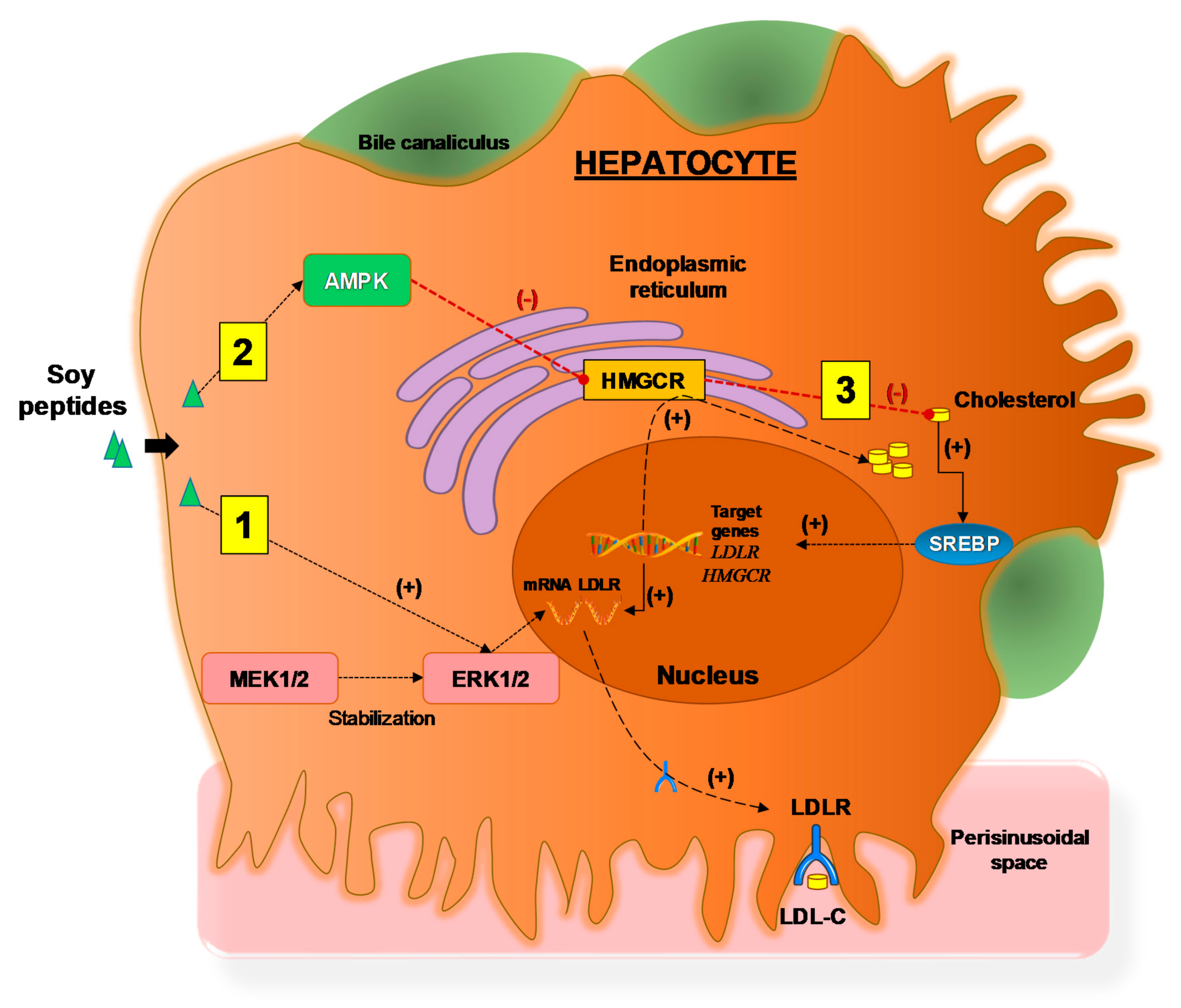
4. Effects of Soy Proteins and Peptides on Lipid Metabolism
5. Effect of Soy Proteins on Lipid Metabolism—Clinical Evidence
6. Discussion
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shimelis, E.A.; Rakshit, S.K. Antinutritional factors andin vitroprotein digestibility of improved haricot bean (Phaseolus vulgaris L.) varieties grown in Ethiopia. Int. J. Food Sci. Nutr. 2005, 56, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Polak, R.; Phillips, E.M.; Campbell, A. Legumes: Health Benefits and Culinary Approaches to Increase Intake. Clin. Diabetes 2015, 33, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hymowitz, T.; Harlan, J.R. Introduction of soybean to North America by Samuel Bowen in 1765. Econ. Bot. 1983, 37, 371–379. [Google Scholar] [CrossRef]
- Abubakar, I.I.; Tillmann, T.; Banerjee, A. Causes of Death, Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death, 1990-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 385, 117–171. [Google Scholar]
- Roth, G.A.; Huffman, M.D.; Moran, A.E.; Feigin, V.L.; Mensah, G.A.; Naghavi, M.; Murray, C.J. Global and Regional Patterns in Cardiovascular Mortality From 1990 to 2013. Circulation 2015, 132, 1667–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Heart Disease and Stroke Statistics-2014 Update: A Report From the American Heart Association. Circulation 2013, 129, e28–e292. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart disease and stroke statistics—2019 update: A report from the American heart association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- Averna, M.R.; Cefalù, A.B.; Casula, M.; Noto, D.; Arca, M.; Bertolini, S.; Calandra, S.; Catapano, A.L.; Tarugi, P.; Pellegatta, F.; et al. Familial hypercholesterolemia: The Italian Atherosclerosis Society Network (LIPIGEN). Atheroscler. Suppl. 2017, 29, 11–16. [Google Scholar] [CrossRef]
- Wang, D.Q.-H.; Neuschwander-Tetri, B.A.; Portincasa, P. The Biliary System. Colloq. Ser. Integr. Syst. Physiol. Mol. Funct. 2012, 4, 1–148. [Google Scholar] [CrossRef]
- Spady, D.K.; Dietschy, J.M. Rates of cholesterol synthesis and low-density lipoprotein uptake in the adrenal glands of the rat, hamster and rabbit in vivo. Biochim. Biophys. Acta 1985, 836, 167–175. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D. Control of Cholesterol Turnover in the Mouse. J. Biol. Chem. 2001, 277, 3801–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spady, D.K.; Turley, S.D.; Dietschy, J.M. Rates of low density lipoprotein uptake and cholesterol synthesis are regulated inde-pendently in the liver. J. Lipid Res. 1985, 26, 465–472. [Google Scholar] [PubMed]
- Spady, D.K.; Turley, S.D.; Dietschy, J.M. Receptor-independent low density lipoprotein transport in the rat in vivo. Quantitation, characterization, and metabolic consequences. J. Clin. Investig. 1985, 76, 1113–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tso, P.; Fujimoto, K. The absorption and transport of lipids by the small intestine. Brain Res. Bull. 1991, 27, 477–482. [Google Scholar] [CrossRef]
- Wang, D.Q.; Lee, S.P. Physical chemistry of intestinal absorption of biliary cholesterol in mice. Hepatology 2008, 48, 177–185. [Google Scholar] [CrossRef]
- Bhattacharyya, A.K.; Eggen, D.A. Relationships between dietary cholesterol, cholesterol absorption, cholesterol synthesis, and plasma cholesterol in rhesus monkeys. Atherosclerosis 1987, 67, 33–39. [Google Scholar] [CrossRef]
- Trautwein, E.A.; Forgbert, K.; Rieckhoff, D.; Erbersdobler, H.F. Impact of β-cyclodextrin and resistant starch on bile acid metabolism and fecal steroid excretion in regard to their hypolipidemic action in hamsters. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1999, 1437, 1–12. [Google Scholar] [CrossRef]
- Turley, S.D.; Daggy, B.P.; Dietschy, J.M. Effect of Feeding Psyllium and Cholestyramine in Combination on Low Density Lipoprotein Metabolism and Fecal Bile Acid Excretion in Hamsters with Dietary-Induced Hypercholesterolemia. J. Cardiovasc. Pharmacol. 1996, 27, 71–79. [Google Scholar] [CrossRef]
- Turley, S.D.; Daggy, B.P.; Dietschy, J.M. Cholesterol-lowering action of psyllium mucilloid in the hamster: Sites and possible mechanisms of action. Metabolism 1991, 40, 1063–1073. [Google Scholar] [CrossRef]
- Wang, D.Q.; Paigen, B.; Carey, M.C. Genetic factors at the enterocyte level account for variations in intestinal cholesterol ab-sorption efficiency among inbred strains of mice. J. Lipid. Res. 2001, 42, 1820–1830. [Google Scholar]
- Turley, S.D.; Dietschy, J.M. Sterol absorption by the small intestine. Curr. Opin. Lipidol. 2003, 14, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Q.-H. Regulation of Intestinal Cholesterol Absorption. Annu. Rev. Physiol. 2007, 69, 221–248. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.W.; Davis, H.R., Jr.; Zhu, L.-J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.N.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-Pick C1 Like 1 Protein Is Critical for Intestinal Cholesterol Absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, D.D. Development and physiological regulation of intestinal lipid absorption. I. Development of intestinal lipid absorption: Cel-lular events in chylomicron assembly and secretion. Am. J. Physiol. Gastrointest Liver Physiol. 2007, 293, G519–G524. [Google Scholar] [CrossRef]
- Nakajima, K.; Nagamine, T.; Fujita, M.Q.; Ai, M.; Tanaka, A.; Schaefer, E. Apolipoprotein B-48. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 117–177. [Google Scholar]
- Wang, D.Q.-H.; Portincasa, P.; Wang, H.H. Bile Formation and Pathophysiology of Gallstones. In Encyclopedia of Gastroenterology; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 287–306. [Google Scholar]
- Wang, D.Q.H.; Neuschwander-Tetri, B.A.; Portincasa, P. The Biliary System, 2nd ed.; Colloquium Series on Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2017; Volume 8, p. i-178. [Google Scholar]
- Wang, H.H.; Liu, M.; Portincasa, P.; Wang, D.Q.-H. Recent Advances in the Critical Role of the Sterol Efflux Transporters ABCG5/G8 in Health and Disease, in Lipid Transfer in Lipoprotein Metabolism and Cardiovascular Disease. In Advances in Experimental Medicine and Biology; Jiang, X.-C., Ed.; Springer: Singapore, 2020; pp. 105–136. [Google Scholar]
- Admirand, W.H.; Small, D.M. The physicochemical basis of cholesterol gallstone formation in man. J. Clin. Investig. 2011, 47, 1043–1052. [Google Scholar] [CrossRef]
- Vítek, L.; Carey, M.C. New pathophysiological concepts underlying pathogenesis of pigment gallstones. Clin. Res. Hepatol. Gastroenterol. 2012, 36, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.Q.-H.; Carey, M.C. Characterization of crystallization pathways during cholesterol precipitation from human gallbladder biles: Identical pathways to corresponding model biles with three predominating sequences. J. Lipid Res. 1996, 37, 2539–2549. [Google Scholar]
- Wang, D.Q.-H.; Cohen, D.E.; Lammert, F.; Carey, M.C. No pathophysiologic relationship of soluble biliary proteins to cholesterol crystallization in human bile. J. Lipid Res. 1999, 40, 415–425. [Google Scholar]
- Brown, M.S.; Goldstein, J.L. How LDL Receptors Influence Cholesterol and Atherosclerosis. Sci. Am. 1984, 251, 58–66. [Google Scholar] [CrossRef]
- Brown, M.S.; Goldstein, J.L. Lipoprotein Metabolism in the Macrophage: Implications for Cholesterol Deposition in Atherosclerosis. Annu. Rev. Biochem. 1983, 52, 223–261. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Kovanen, P.T.; Goldstein, J.L. Regulation of plasma cholesterol by lipoprotein receptors. Science 1981, 212, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Bleich, H.L.; Boro, E.S.; Small, D.M. Cellular Mechanisms for Lipid Deposition in Atherosclerosis. N. Engl. J. Med. 1977, 297, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M. George Lyman Duff memorial lecture. Progression and regression of atherosclerotic lesions. Insights from lipid physical biochemistry. Arter. Off. J. Am. Hear. Assoc. Inc. 1988, 8, 103–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, D.M.; Shipley, G.G. Physical-Chemical Basis of Lipid Deposition in Atherosclerosis. Science 1974, 185, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.L.; Brown, M.S. Regulation of low-density lipoprotein receptors: Implications for pathogenesis and therapy of hyper-cholesterolemia and atherosclerosis. Circulation 1987, 76, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Ciaula, A.; Garruti, G.; Frühbeck, G.; de Angelis, M.; de Bari, O.; Wang, D.Q.-H.; Lammert, F.; Portincasa, P. The Role of Diet in the Pathogenesis of Cholesterol Gallstones. Curr. Med. Chem. 2019, 26, 3620–3638. [Google Scholar] [CrossRef]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced Dietary Intake of Carbohydrates by Obese Subjects Results in Decreased Concentrations of Butyrate and Butyrate-Producing Bacteria in Feces. Appl. Environ. Microbiol. 2006, 73, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Clandinin, M.T.; Foxwell, A.; Goh, Y.K.; Layne, K.; Jumpsen, J.A. Omega-3 fatty acid intake results in a relationship between the fatty acid composition of LDL cholesterol ester and LDL cholesterol content in humans. Biochim. Biophys. Acta Lipids Lipid Metab. 1997, 1346, 247–252. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Buitrago-Lopez, A.; Sanderson, J.; Johnson, L.; Warnakula, S.; Wood, A.; di Angelantonio, E.; Franco, O.H. Chocolate consumption and cardiometabolic disorders: Systematic review and meta-analysis. BMJ 2011, 343, d4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagna, M.T.; Diella, G.; Triggiano, F.; Caponio, G.R.; Giglio, O.D.; Caggiano, G.; Ciaula, A.D.; Portincasa, P. Chocolate, “Food of the Gods”: History, Science, and Human Health. Int. J. Environ. Res. Public Health 2019, 16, 4960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, K.; Takeda, A.; Martin, N.; Ellis, L.; Wijesekara, D.; Vepa, A.; Das, A.; Hartley, L.; Stranges, S. Mediterranean-style diet for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2019, 3, CD009825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lorgeril, M.; Salen, P. The Mediterranean-style diet for the prevention of cardiovascular diseases. Public Health Nutr. 2007, 9, 118–123. [Google Scholar] [CrossRef]
- Becerra-Tomas, N.; Mejía, S.B.; Viguiliouk, E.; Khan, T.; Kendall, W.C.C.; Kahleova, H. Mediterranean diet, cardiovascular disease and mortality in diabetes: A systematic review and me-ta-analysis of prospective cohort studies and randomized clinical trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 1207–1227. [Google Scholar] [CrossRef]
- Widmer, R.J.; Flammer, J.; Lerman, L.O.; Lerman, A. The Mediterranean Diet, its Components, and Cardiovascular Disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.W.; Johnstone, B.M.; Cook-Newell, M.E. Meta-Analysis of the Effects of Soy Protein Intake on Serum Lipids. N. Engl. J. Med. 1995, 333, 276–282. [Google Scholar] [CrossRef]
- Sacks, F.M.; Lichtenstein, A.; van Horn, L.; Harris, W.; Kris-Etherton, P.; Winston, M. Soy protein, isoflavones, and cardiovascular health: An American Heart Association Science Advisory for profes-sionals from the Nutrition Committee. Circulation 2006, 113, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Velasquez, M.T.; Bhathena, S.J. Role of Dietary Soy Protein in Obesity. Int. J. Med. Sci. 2007, 4, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Food labeling: Health claims; soy protein and coronary heart disease. Food and Drug Administration, HHS. Final rule. Fed. Regist. 1999, 64, 57700–57733.
- Nishinari, K.; Fang, Y.; Guo, S.; Phillips, G.O. Soy proteins: A review on composition, aggregation and emulsification. Food Hydrocoll. 2014, 39, 301–318. [Google Scholar] [CrossRef]
- Gianazza, E.; Eberini, I.; Arnoldi, A.; Wait, R.; Sirtori, C.R. A Proteomic Investigation of Isolated Soy Proteins with Variable Effects in Experimental and Clinical Studies. J. Nutr. 2003, 133, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, L.; Dia, V.P.; de Mejía, E.G. Soy Proteins. In Applied Food Protein Chemistry; Ustunol, Z., Ed.; Wiley-Blackwell: Hoboken, NJ, USA.
- Xiao, C.W.; Mei, J.; Wood, C. Effect of soy proteins and isoflavones on lipid metabolism and involved gene expression. Front Biosci. 2008, 13, 2660–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovati, M.R.; Manzoni, C.; Gianazza, E.; Arnoldi, A.; Kurowska, E.; Carroll, K.K.; Sirtori, C.R. Soy Protein Peptides Regulate Cholesterol Homeostasis in Hep G2 Cells. J. Nutr. 2000, 130, 2543–2549. [Google Scholar] [CrossRef] [Green Version]
- Matoba, N.; Doyama, N.; Yamada, Y.; Maruyama, N.; Utsumi, S.; Yoshikawa, M. Design and production of genetically modified soybean protein with anti-hypertensive activity by incorporating potent analogue of ovokinin(2-7). FEBS Lett. 2001, 497, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-K.; Adachi, M.; Utsumi, S. Identification of the Bile Acid-binding Region in the Soy Glycinin A1aB1b Subunit. Biosci. Biotechnol. Biochem. 2002, 66, 2395–2401. [Google Scholar] [CrossRef]
- Friedman, M.; Brandon, D.L. Nutritional and Health Benefits of Soy Proteins†. J. Agric. Food Chem. 2001, 49, 1069–1086. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A. IAVPGEVA, IAVPTGVA, and LPYP, three peptides from soy glycinin, modulate cholesterol metabolism in HepG2 cells through the activation of the LDLR-SREBP2 pathway. J. Funct. Foods 2015, 14, 469–478. [Google Scholar] [CrossRef]
- Pak, V.V.; Koo, M.S.; Kasymova, T.D.; Kwon, D.Y. Isolation and Identification of Peptides from Soy 11S-Globulin with Hypocholesterolemic Activity. Chem. Nat. Compd. 2005, 41, 710–714. [Google Scholar] [CrossRef]
- di Ciaula, A.; Garruti, G.; Baccetto, R.L.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.-H.; Portincasa, P. Bile Acid Physiology. Ann. Hepatol. 2017, 16, S4–S14. [Google Scholar] [CrossRef]
- di Ciaula, A.; Baj, J.; Garruti, G.; Celano, G.; de Angelis, M.; Wang, H.H.; di Palo, D.M.; Bonfrate, L.; Wang, D.Q.-H.; Portincasa, P. Liver Steatosis, Gut-Liver Axis, Microbiome and Environmental Factors. A Never-Ending Bidirectional Cross-Talk. J. Clin. Med. 2020, 9, 2648. [Google Scholar] [CrossRef] [PubMed]
- Balmer, J.; Zilversmit, D.B. Effects of Dietary Roughage on Cholesterol Absorption, Cholesterol Turnover and Steroid Excretion in the Rat. J. Nutr. 1974, 104, 1319–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Two Peptides from Soy beta-Conglycinin Induce a Hypocholesterolemic Effect in HepG2 Cells by a Statin-Like Mechanism: Comparative in Vitro and in Silico Modeling Studies. J. Agric. Food Chem. 2015, 63, 7945–7951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovati, M.R.; Manzoni, C.; Gianazza, E.; Sirtori, C.R. Soybean Protein Products as Regulators of Liver Low-Density Lipoprotein Receptors. I. Identification of Active β-Conglycinin Subunits. J. Agric. Food Chem. 1998, 46, 2474–2480. [Google Scholar] [CrossRef]
- Manzoni, C.; Lovati, M.R.; Gianazza, E.; Morita, Y.; Sirtori, C.R. Soybean Protein Products as Regulators of Liver Low-Density Lipoprotein Receptors. II. α−α‘ Rich Commercial Soy Concentrate and α‘ Deficient Mutant Differently Affect Low-Density Lipoprotein Receptor Activation. J. Agric. Food Chem. 1998, 46, 2481–2484. [Google Scholar] [CrossRef]
- Cho, S.-J.; Juillerat, M.A.; Lee, C.-H. Identification of LDL-Receptor Transcription Stimulating Peptides from Soybean Hydrolysate in Human Hepatocytes. J. Agric. Food Chem. 2008, 56, 4372–4376. [Google Scholar] [CrossRef]
- Lule, V.K.; Garg, S.; Pophaly, S.D.; Hitesh-Tomar, S.K. Potential Health Benefits of Lunasin: A Multifaceted Soy-Derived Bioactive Peptide. J. Food Sci. 2015, 80, R485–R494. [Google Scholar] [CrossRef]
- Hodges, R.E.; Krehl, W.A.; Stone, D.B.; Lopez, A. Dietary Carbohydrates and Low Cholesterol Diets: Effects on Serum Lipids of Man. Am. J. Clin. Nutr. 1967, 20, 198–208. [Google Scholar] [CrossRef]
- Lovati, M.R.; Manzoni, C.; Canavesi, A.; Sirtori, M.; Vaccarino, V.; Marchi, M.; Gaddi, G.; Sirtori, C.R. Soybean protein diet increases low density lipoprotein receptor activity in mononuclear cells from hypercholester-olemic patients. J. Clin. Investig. 1987, 80, 1498–1502. [Google Scholar] [CrossRef] [Green Version]
- Laurin, D.; Jacques, H.; Moorjani, S.; Steinke, F.H.; Gagné, C.; Brun, D.; Lupien, P.J. Effects of a soy-protein beverage on plasma lipoproteins in children with familial hypercholesterolemia. Am. J. Clin. Nutr. 1991, 54, 98–103. [Google Scholar] [CrossRef]
- Wang, M.-F.; Yamamoto, S.; Chung, H.-M.; Chung, S.-Y.; Miyatani, S.; Mori, M.; Okita, T.; Sugano, M. Antihypercholesterolemic Effect of Undigested Fraction of Soybean Protein in Young Female Volunteers. J. Nutr. Sci. Vitaminol. 1995, 41, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Sagara, M.; Kanda, T.; Njelekera, M.; Teramoto, T.; Armitage, L.; Birt, N.; Birt, C.; Yamori, Y. Effects of Dietary Intake of Soy Protein and Isoflavones on Cardiovascular Disease Risk Factors in High Risk, Middle-Aged Men in Scotland. J. Am. Coll. Nutr. 2004, 23, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jones, P.J.H.; Ausman, L.M.; Lichtenstein, A.H. Soy protein reduces triglyceride levels and triglyceride fatty acid fractional synthesis rate in hypercholesterolemic subjects. Atherosclerosis 2004, 173, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-T.; Chen, J.-R.; Yang, C.-S.; Peng, S.-J.; Ferng, S.-H. Effect of soya protein on serum lipid profile and lipoprotein concentrations in patients undergoing hypercholesterol-aemic haemodialysis. Br. J. Nutr. 2007, 95, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, K.C.; Butteiger, D.N.; Rains, T.M.; Lawless, A.; Reeves, M.S.; Schasteen, C.; Krul, E.S. Effects of soy protein on lipoprotein lipids and fecal bile acid excretion in men and women with moderate hypercho-lesterolemia. J. Clin. Lipidol. 2010, 4, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.W.; Smith, E.O.; Stuff, J.E.; Hachey, D.L.; Heird, W.C.; Pownell, H.J. Cholesterol-lowering effect of soy protein in normocholesterolemic and hypercholesterolemic men. Am. J. Clin. Nutr. 1998, 68, 1385S–1389S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, S.R.; Potter, S.M.; Weigel, R.; Hannum, S.; Erdman, J.W., Jr.; Hasler, C.M. Effects of feeding 4 levels of soy protein for 3 and 6 wk on blood lipids and apolipoproteins in moderately hyper-cholesterolemic men. Am. J. Clin. Nutr. 2000, 71, 1077–1084. [Google Scholar] [CrossRef]
- Potter, S.M.; Baum, J.A.; Teng, H.; Stillman, R.J.; Shay, N.F.; Erdman, J.W. Soy protein and isoflavones: Their effects on blood lipids and bone density in postmenopausal women. Am. J. Clin. Nutr. 1998, 68, 1375S–1379S. [Google Scholar] [CrossRef]
- Baum, J.A.; Teng, H.; Erdman, J.W.; Weigel, R.M.; Klein, B.P.; Persky, V.W.; Freels, S.; Surya, P.; Bakhit, R.M.; Ramos, E.; et al. Long-term intake of soy protein improves blood lipid profiles and increases mononuclear cell low-density-lipoprotein receptor messenger RNA in hypercholesterolemic, postmenopausal women. Am. J. Clin. Nutr. 1998, 68, 545–551. [Google Scholar] [CrossRef]
- Mackey, R.; Ekangaki, A.; Eden, J.A. The effects of soy protein in women and men with elevated plasma lipids. BioFactors 2000, 12, 251–257. [Google Scholar] [CrossRef]
- Mejia, S.B.; Messina, M.; Li, S.S.; Viguiliouk, E.; Chiavaroli, L.; Khan, T.A.; Srichaikul, K.; Mirrahimi, A.; Sievenpiper, J.L.; Kris-Etherton, P.M.; et al. A Meta-Analysis of 46 Studies Identified by the FDA Demonstrates that Soy Protein Decreases Circulating LDL and Total Cholesterol Concentrations in Adults. J. Nutr. 2019, 149, 968–981. [Google Scholar] [CrossRef]
- Hermansen, K.; Søndergaard, M.; Høie, L.; Carstensen, M.; Brock, B. Beneficial effects of a soy-based dietary supplement on lipid levels and cardiovascular risk markers in type 2 diabetic subjects. Diabetes Care 2001, 24, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.W.; Hoie, L.H. Weight Loss and Lipid Changes with Low-Energy Diets: Comparator Study of Milk-Based versus Soy-Based Liquid Meal Replacement Interventions. J. Am. Coll. Nutr. 2005, 24, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Crouse, J.R., 3rd; Morgan, T.; Terry, J.G.; Ellis, J.; Vitolins, M.; Burke, G.L. A randomized trial comparing the effect of casein with that of soy protein containing varying amounts of iso-flavones on plasma concentrations of lipids and lipoproteins. Arch. Intern. Med. 1999, 159, 2070–2076. [Google Scholar] [CrossRef] [Green Version]
- Washburn, S.; Burke, G.L.; Morgan, T.; Anthony, M. Effect of Soy Protein Supplementation on Serum Lipoproteins, Blood Pressure, and Menopausal Symptoms in Perimenopausal Women. Menopause 1999, 6. [Google Scholar] [CrossRef]
- Merz-Demlow, B.E.; Duncan, A.M.; Wangen, K.E.; Xu, X.; Carr, T.P.; Phipps, W.R.; Kurzer, M.S. Soy isoflavones improve plasma lipids in normocholesterolemic, premenopausal women. Am. J. Clin. Nutr. 2000, 71, 1462–1469. [Google Scholar] [CrossRef] [Green Version]
- Takatsuka, N.; Nagata, C.; Kurisu, B.P.Y.; Inaba, S.; Kawakami, N.; Shimizu, H. Hypocholesterolemic effect of soymilk supplementation with usual diet in premenopausal normolipidemic Japa-nese women. Prev. Med. 2000, 31, 308–314. [Google Scholar] [CrossRef]
- Chiechi, L.; Secreto, G.; Vimercati, A.; Greco, P.; Venturelli, E.; Pansini, F.; Fanelli, M.; Loizzi, P.; Selvaggi, L. The effects of a soy rich diet on serum lipids: The Menfis randomized trial. Maturitas 2002, 41, 97–104. [Google Scholar] [CrossRef]
- Teede, H.J.; Dalais, F.S.; Kotsopoulos, D.; Liang, Y.L.; Davis, S.; McGrath, B.P. Dietary soy has both beneficial and potentially adverse cardiovascular effects: A placebo-controlled study in men and postmenopausal women. J. Clin. Endocrinol. Metab. 2001, 86, 3053–3060. [Google Scholar] [CrossRef]
- Rosell, M.S.; Appleby, P.N.; Spencer, E.A.; Key, T.J. Soy intake and blood cholesterol concentrations: A cross-sectional study of 1033 pre- and postmenopausal women in the Oxford arm of the European Prospective Investigation into Cancer and Nutrition. Am. J. Clin. Nutr. 2004, 80, 1391–1396. [Google Scholar] [CrossRef] [Green Version]
- Nagata, C.; Takatsuka, N.; Kurisu, Y.; Shimizu, H. Decreased Serum Total Cholesterol Concentration Is Associated with High Intake of Soy Products in Japanese Men and Women. J. Nutr. 1998, 128, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; D’Angelo, A.; Romano, D.; Maffioli, P. Response to an oral fat load and effects on lipid profile, glycemia and high-sensitivity C-reactive protein after soy-bean extract consumption. Arch. Med. Sci. 2018, 14, 760–765. [Google Scholar] [PubMed] [Green Version]
- George, K.S.; Muñoz, J.; Akhavan, N.S.; Foley, E.M.; Siebert, S.C.; Tenenbaum, G.; Khalil, D.A.; Chai, S.C.; Arjmandi, B.H. Is soy protein effective in reducing cholesterol and improving bone health? Food Funct. 2020, 11, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, S.H.; Haddad, E.; Rajaram, S.; Oda, K.; Kaur, A.; Sabaté, J. The Effect of Soybean Lunasin on Cardiometabolic Risk Factors: A Randomized Clinical Trial. J. Diet. Suppl. 2019, 17, 286–299. [Google Scholar] [CrossRef]
- Willett, W. Nutritional Epidemiology, 3rd ed.; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Diet, nutrition and the prevention of chronic diseases. In World Health Organization Technical Report Series; WHO: Geneva, Switzerland, 2003; Volume 916, pp. i–viii, 1–149, backcover.
- Keller, I.; Lang, T. Food-based dietary guidelines and implementation: Lessons from four countries—Chile, Germany, New Zealand and South Africa. Public Health Nutr. 2008, 11, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wu, X.; Zhuang, W.; Xia, L.; Chen, Y.; Zhao, R.; Yi, M.; Wan, Q.; Du, L.; Zhou, Y. Soy and Isoflavone Consumption and Multiple Health Outcomes: Umbrella Review of Systematic Reviews and Me-ta-Analyses of Observational Studies and Randomized Trials in Humans. Mol. Nutr. Food Res. 2020, 64, e1900751. [Google Scholar] [CrossRef]
- Wilunda, C.; Sawada, N.; Goto, A.; Yamaji, T.; Iwasaki, M.; Tsugane, S.; Noda, M. Soy food and isoflavones are not associated with changes in serum lipids and glycohemoglobin concentrations among Japanese adults: A cohort study. Eur. J. Nutr. 2020, 59, 2075–2087. [Google Scholar] [CrossRef]
- Portincasa, P.; di Ciaula, A.; Garruti, G.; Vacca, M.; de Angelis, M.; Wang, D.Q. Bile Acids and GPBAR-1: Dynamic Interaction Involving Genes, Environment and Gut Microbiome. Nutrients 2020, 12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | Recommended Doses | Components |
|---|---|---|
| Extra-virgin olive oil | 50 g/day | Polyphenols |
| Tree nuts and peanuts | 30 g/day | Essential omega 6 and omega 3 fatty acids |
| Fresh fruits/vegetables | ≥3 servings/day | Flavonols, vitamins |
| Fish, seafood | ≥3 servings/week | Essential omega 6 and omega 3 fatty acids |
| Legumes | ≥3 servings/week | Fibers, proteins |
| Soy Protein | Peptides | Doses | Experimental Model | Effects/Bioactivity | References |
|---|---|---|---|---|---|
| Glycinin (11S) | IAVPGEVA, IAVPTGVA, LPYP | 25, 50, 100, 150, 200, 250, 300, 400, 500, 650, 1000 µM | HepG2 cell line | Increased LDLR activity Inhibition of HMGCR Activation of the AMPK pathway and ERK1/2 | [63] |
| IAVPGEVA | - | Microsomes from rat liver | Inhibition of HMGCR Seizure of bile acids | [64] | |
| β-Conglycinin (7S) | YVVNPDNDEN, YVVNPDNNEN | 350 and 500 μM | HepG2 cell line | Inhibition of HMGCR Upregulation of the mature SREBP-2 protein level Increased the level of LDLR protein | [68] |
| α, α′ subunits | 0.25, 0.50, 0.75 g/L | HepG2 cells line | Upregulation of the LDLR | [70] | |
| α, α′ and β subunits | 10−4 mol/L | HepG2 cells line | Upregulation of the LDLR | [59] | |
| FVVNATSN | 100 µM | Human hepatocytes (Hep T9A4) | Influenced LDLR transcription | [71] | |
| 2S albumin | Lunasin | - | HepG2 cells line | Reduced expression of HMGCR | [72] |
| Authors | Sample Size | Gender (Age) | Format, Dose | Duration of Study | Main Findings |
|---|---|---|---|---|---|
| Hypercholesterolemia | |||||
| Hodges et al., 1967 [73] | N = 6 | Men (33–46 years) | n.a. | 4 weeks | Reduction in the average plasma cholesterol levels by more than 100 mg/mL. |
| Lovati et al., 1987 [74] | N = 12 | 5 men and 7 women (26–64 years) | Soy proteins (20% calories) | 4 weeks | Reduction in total cholesterol by 15.9% and LDL-C by 16.4%. Degradation of LDL-C by mononuclear cells after soybean diet (increased 16-fold vs. the basal activity and 8-fold compared with the standard low lipid diet with animal proteins). |
| Laurin et al., 1991 [75] | N = 55 | Children (6–12 years) | Soy beverage (250 mL) | 4 weeks | No changes in either plasma total cholesterol, LDL-C, or apo concentrations. Reduction in triglyceride and VLDL concentrations. Increased HDL-C and HDL3-C concentrations. |
| Wang et al., 1995 [76] | n.a. | Females (median age 19 years) | Soy proteins, purified (4 or 8% of total energy) | 14 days | Decreased LDL-C, increased HDL-C and increased fecal steroid excretion. |
| Baum et al., 1998 [84] | N = 66 | Postmenopausal women (39–83 years) | Soy proteins (40 g/day containing 1.39 or 2.25 mg isoflavones/g protein) | 6 months | Decreased non-HDL-C levels. Unchanged plasma total cholesterol levels. Increased HDL-C levels and decreased ratio total cholesterol and HDL-C in both groups compared with control. Increased LDLR messenger RNA concentrations in mononuclear cells. |
| Potter et al., 1998 [83] | N = 66 | Postmenopausal women (39–83 years) | Soy proteins at 40 g/day (containing 1.39 or 2.25 mg isoflavones/g protein) | 6 months | Reduction in non-HDL-C levels. Increased HDL-C levels. Increased mononuclear cell LDLR mRNA. |
| Wong et al., 1998 [81] | N = 13 | Men (20–50 years) | Soy beverage (≥75% of the total protein content of diet) | 5 weeks | Decreased LDL-C and LDL-C/HDL-C ratio. |
| Mackey et al., 2000 [85] | N = 54 | Postmenopausal women (median age 56 years) | Soy proteins (28 g with 65 mg or 4 mg isoflavones) | 12 weeks | Reduction in TC, LDL-C, sex hormone binding globulin (SHBG), and luteinizing hormone (LH). No significant differences between treatment groups. Cholesterol-lowering effect in both women and men independent of isoflavones. |
| Teixeira et al., 2000 [82] | N = 81 | Men (23–74 years) | Soy proteins (20 g/day) | 6 weeks | Reduced non-HDL, total cholesterol and apoB. No change in HDL-C, apoA-I, lipoprotein(a), and triglyceride concentrations. |
| Maki et al., 2000 [80] | N = 30 | 13 men and 17 women (18-79 years) | Soy proteins (25 g/day) | 4 weeks | Reduced atherogenic lipoproteins, as indicated by changes in total cholesterol (27.4 and 23.6%), LDL-C (210.9 and 25.9%), non-HDL-C (210.8 and 23.9%), and apoB (29.7 and 22.4%), respectively. |
| Blanco Mejia, 2019 [86] | N = 2607 | 37% men and 63% women (median age 54.9 years) | Soy proteins (25 g/day) | 6 weeks | Decreased LDL-C by 4.76 mg/dL and total cholesterol by 6.41 mg/dL. |
| Type 2 diabetes | |||||
| Hermansen et al., 2001 [87] | N = 20 | 14 men and 6 women (median age 63.6 years) | Soy proteins (50 g/day) plus high levels (minimum 165 mg/day) of isoflavones | 6 weeks | Reduction in LDL-C, LDL/HDL ratio, apoB100, triglyceride, and homocysteine concentrations. |
| Overweight/obesity | |||||
| Anderson et al., 2005 [88] | N = 51 | 4 men and 47 women (18–65 years) | Five soy-based meal replacements (Soy MR)/day | 12 weeks | Decreased plasma total cholesterol, LDL-C, and triglyceride concentrations. |
| Healthy subjects | |||||
| Nagata et al., 1998 [96] | N = 4838 | 1242 men and 3596 women (median age 58.38 years) | Soy proteins (<8 g/day) | 1 year | Decreased total cholesterol concentrations with an increasing intake of soy products in men and women after controlling for age, smoking status and intake of total energy, total proteins and total fat. |
| Crouse et al., 1999 [89] | N = 156 | 94 men and 62 pre and postmenopausal women (20–70 years) | Isolated soy proteins (25 g) plus isoflavones at 3, 27, 37, or 62 mg | 9 weeks | Reduction in total and LDL-C levels by 4 and 6% with isolated soy protein with 62 mg of IFS vs casein. No change in plasma triglyceride and HDL-C concentrations. |
| Washburn et al., 1999 [90] | N = 51 | Perimenopausal women (45–55 years) | Soy proteins (20 g) plus 34 mg of phytoestrogens (once daily or twice daily) | 18 weeks | Significant declines in total cholesterol (6% lower) and LDL-C (7% lower). Significant decline in diastolic blood pressure (5 mm Hg lower) in the twice-daily soy diet, compared with the placebo diet. No significant effects for triglycerides, HDC-C or frequency of menopausal symptoms. |
| Merz.Demlow, 2000 [91] | N = 33 | Premenopausal women (18–35 years) | Soy isoflavones (low: 64.7 ± 9.4 and high: 128.7 ± 15.7 mg/day) | Three menstrual cycles | Change in total cholesterol, HDL-C, and LDL-C concentrations across menstrual cycle phases (P < 0.005). |
| Takatsuka et al., 2000 [92] | N = 52 | Premenopausal women (median age 26 years) | Soymilk (400 mL = 408 g) /day | Two menstrual cycles | Decrease of 10.9 mg/dL, or 5.3%, in serum total cholesterol concentrations in the soymilk-supplemented group. |
| Teede et al., 2001 [94] | N = 213 | 108 men and 105 post-menopausal women (50–75 years) | Soy proteins (40 g) and isoflavones at 118 mg/day | 3 months | Reduction in the LDL-C/HDL-C ratio and triglyceride levels No change in total cholesterol, LDL-C and HDL-C concentrations. |
| Chiechi et, 2002 [93] | N = 187 | Postmenopausal women (39–60 years) | Soy food every day with isoflavones amounting to 40–60 mg/day | 6 months | Improved in lipid profile |
| Rosell et al., 2004 [95] | N = 1033 | Pre-and postmenopausal women (≥20 years) | Soy proteins <0.5, 0.5–2.9, 3.0–5.9, and ≥6.0 g/day | n.a. | Inversely associated relationship between soy protein intake and total cholesterol and LDL-C concentrations and the ratio of total cholesterol to HDL-C, but not to HDL-C concentrations. |
| Derosa et al., 2018 [97] | N = 124 | Men and women (≥18 years) | Fermented soybean extract | n.a. | Decreased of total cholesterol, TG and LDL-C levels. No change in HDL-C levels. |
| George et al., 2020 [98] | N = 135 | 65 men and 70 women (27–87 years) | Soy proteins 40 g/day | 3 months | No effect on total cholesterol, HDL-C, or LDL-C levels. Reduction in bone alkaline phosphatase and body fat percentages. |
| Haddad Tabrizi, 2020 [99] | N = 31 | 12 men and 19 women (median age 61 years) | Lunasin enriched soybean extract 335 mg/day | 8 weeks | No significant changes in serum lipids, glucose, insulin resistance, blood pressure, BMI, or waist circumference. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caponio, G.R.; Wang, D.Q.-H.; Di Ciaula, A.; De Angelis, M.; Portincasa, P. Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence. Int. J. Mol. Sci. 2021, 22, 227. https://doi.org/10.3390/ijms22010227
Caponio GR, Wang DQ-H, Di Ciaula A, De Angelis M, Portincasa P. Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence. International Journal of Molecular Sciences. 2021; 22(1):227. https://doi.org/10.3390/ijms22010227
Chicago/Turabian StyleCaponio, Giusy Rita, David Q.-H. Wang, Agostino Di Ciaula, Maria De Angelis, and Piero Portincasa. 2021. "Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence" International Journal of Molecular Sciences 22, no. 1: 227. https://doi.org/10.3390/ijms22010227
APA StyleCaponio, G. R., Wang, D. Q.-H., Di Ciaula, A., De Angelis, M., & Portincasa, P. (2021). Regulation of Cholesterol Metabolism by Bioactive Components of Soy Proteins: Novel Translational Evidence. International Journal of Molecular Sciences, 22(1), 227. https://doi.org/10.3390/ijms22010227