Abstract
Modified mRNA (modRNA)-based somatic reprogramming is an effective and safe approach that overcomes the genomic mutation risk caused by viral integrative methods. It has improved the disadvantages of conventional mRNA and has better stability and immunogenicity. The modRNA molecules encoding multiple pluripotent factors have been applied successfully in reprogramming somatic cells such as fibroblasts, mesenchymal stem cells, and amniotic fluid stem cells to generate pluripotent stem cells (iPSCs). Moreover, it also can be directly used in the terminal differentiation of stem cells and fibroblasts into functional therapeutic cells, which exhibit great promise in disease modeling, drug screening, cell transplantation therapy, and regenerative medicine. In this review, we summarized the reprogramming applications of modified mRNA in iPSC generation and therapeutic applications of functionally differentiated cells.
1. Introduction
Embryonic stem cells (ESCs) are derived from the inner cell mass (ICM) of embryos at the early stage of blastocyst development [1]. ESCs are pluripotent stem cells and are able to differentiate into any cell type of the body. Thus, pluripotent stem cells hold promise in regenerative medicine and disease modeling. However, harvesting the embryoblast results in the destruction of the blastocyst and therefore faces ethical issues. In 2006, Yamanaka et al. first discovered and generated induced pluripotent stem cells (iPSCs) from mouse fibroblasts transduced with four transcription factors such as Oct3/4, Sox2, c-Myc, and Klf4 (OSKM), under ES cell culture conditions [2]. These cells exhibit characteristics similar to ESCs such as morphology, growth, pluripotency, and marker expression [3,4]. ESCs are derived from the ICM of the blastocyst, whereas iPSCs are derived from a variety of somatic cell types using various reprogramming techniques. Unlike ESCs, iPSCs are more readily obtainable for therapy and research, and their harvest does not face ethical concerns [5]. For example, the mutated genes of iPSCs derived from diseased patients can be repaired using the homologous recombination method and gene editing using CRISPR–Cas9 technology. The gene-corrected terminally differentiated cells derived from diseased iPSCs can be transferred into diseased patients for cell therapy. iPSCs generated from healthy or diseased cells can also be used for the in vitro screening of drug candidates [6]. Therefore, autologous iPSCs-derived therapeutic cells are preferred for use in diseased patients. This shows great promise for precision and personalized medicine.
There are four major reprogramming methods for the generation of iPSCs derived from somatic cells through the overexpression of pluripotent facts. (1) Virus-based integrative reprogramming method: virus transduction of reprogramming factors such as retrovirus and lentivirus were used for overexpression of Oct3/4, Sox2, c-Myc, and Klf4 in somatic cells, which lead to somatic cell transformation into pluripotent stem cells. Although virus-based methods lead to highly efficient reprogramming cells (0.01~0.1%) [7,8,9], virus-carrying transgenes would be randomly integrated into chromosomes of somatic cells. This causes iPSCs to be at risk of chromosome gene mutation. The use of proto-oncogene c-Myc in gene reactivation could increase the risk of transgene-derived tumor formation. Although cure-excisable lentiviral systems offer a solution to genome integration, they require lengthy sub-cloning procedures and screening to ensure excision of the reprogramming factors. (2) RNA virus-based non-integrative reprogramming method: Sendai virus is used for the overexpression of pluripotent transgenes and subsequently iPSC generation. The Sendai virus method is easy to use and leads to highly efficient reprogramming cells (0.01~1%) [10,11,12], whereas residual Sendai virus is difficult to clear from somatic cells. This needs several rounds of clonal expansion and analysis. (3) Plasmid-based non-integrative reprogramming method: Episomal DNA plasmid-carried pluripotent genes are used for iPSC generation, which are transgene free [13,14]. The technique was gradually improved, and the reprogramming efficiency ranged between 0.04% and 0.3% [13]. The elimination of residual episomal plasmids in somatic cells would need several rounds of cell culture. (4) mRNA-based non-integrative method: mRNA-carrying pluripotent genes are used for iPSC generation. mRNA reprogramming technology is the most unambiguously footprint free and genomic integration free for iPSC generation. However, conventional mRNA transcripts exhibit some disadvantages such as instability, immune activation, and difficult delivery, which limit their applications and reduce reprogramming efficiency. In recent years, researchers have gradually developed modified mRNA transcripts, which enhance their stability, reduce immunogenicity, and improve their delivery. Therefore, in this review, we summarized the reprogramming applications of modified mRNA (modRNA) in iPSC generation and therapeutic applications of these iPSCs and functionally differentiated cells.
2. Messenger RNA
2.1. Natural mRNA
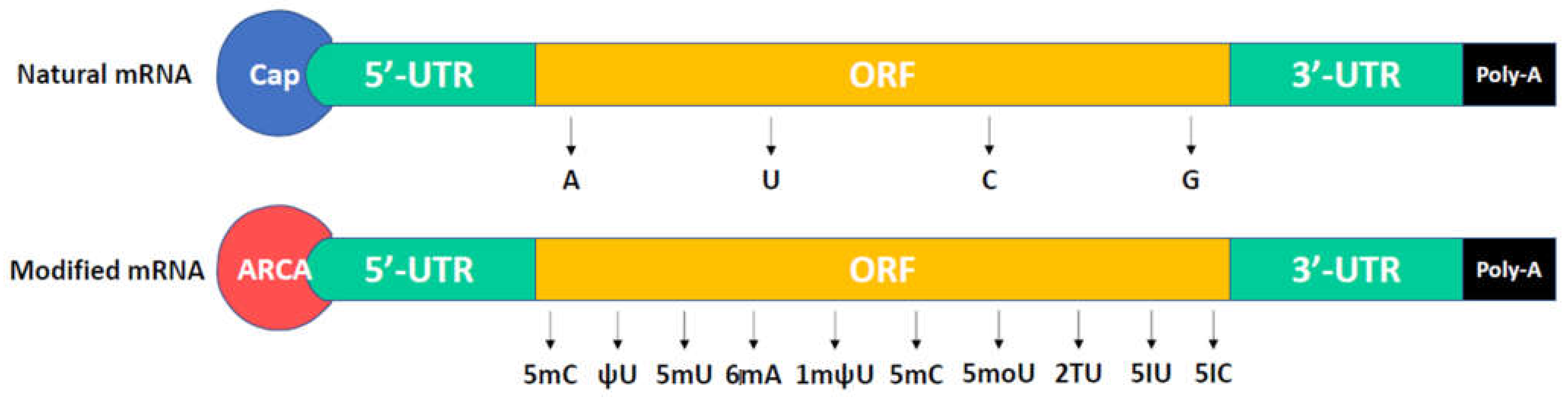
Transcription is a process that makes an RNA copy from a DNA template through RNA polymerase. The RNA copy or transcript carries information and can be translated as a polypeptide [15]. In eukaryotes, an RNA transcript needs some processing including splicing and incorporation of 5′ cap and 3′ ploy-A tail on their ends [16]. The structure of a mature eukaryotic mRNA is shown in Figure 1. A mature mRNA contains 5′ cap, poly-A tail, 5′UTR (untranslated region), 3′ UTR, and ORF (open reading frame). The 5′ cap is a modified guanine nucleotide and is added to the 5′ end of mRNA. It contains a 7-methylguanosine residue (m7G), which is linked to the first nucleotide of mRNA through a 5′-5′-triphosphate. 5′cap exhibits protection from RNases and stabilizes the mRNA, in addition to the recognition by the ribosome for protein translation [17,18]. The poly-A tail with an optimal length of 100–200 nucleotides is added to the 3′ end of mRNA through polyadenylate polymerase, which the process calls polyadenylation [19]. An abnormal length makes the mRNA structure unstable. Polyadenylation is the covalent linkage of a polyadenylyl moiety to mRNA. A poly-U tail is also found in some mRNA [20]. The poly-A tail facilitates mRNA protection from exonucleases-caused degradation [21,22]. Poly-A tail also contributes to the export of mRNA from nucleus into cytosol for protein translation. 5′ UTR and 3′ UTR are located before the start codon of mRNA and after the stop codon of mRNA, respectively. UTRs can facilitate mRNA stability, translational efficiency, and mRNA localization depends on their sequence [23,24,25]. UTRs aid the stability of mRNA because of various affinities for RNA-degrading enzymes (ribonucleases). UTRs may influence translation efficiency due to the competition of other proteins with ribosomes for binding to mRNA [26]. 3′ UTR functions as the cytoplasmic localization of mRNA, which contains sequences that allow RNA transcript to go to the translated region [27,28]. The open reading frame is a coding region, which is decoded and translated into proteins by ribosomes. The stability of mRNA is also associated with internal base pairs of coding regions [29,30].
Figure 1.
The structure of a mature eukaryotic mRNA and a modified mRNA. The anti-reverse cap analogs (ARCAs) and modified nucleotides such as 5-methyluridine (5mU), 5-methylcytidine (5mC), pseudouridine (ψU), N6-methyladenosine (6mA), N1-methylpseudouridine (1mψU), 5-methoxyuridine (5moU), 2-thiouridine (2TU), 5-iodouridine (5IU), and 5-iodocytidine (5IC) are usually used in modified mRNA.
2.2. Modified mRNA
In recent years, substantial modifications of mRNA have been investigated for their extensive applications. The structure of a modified mRNA (modRNA) is shown in Figure 1, including the 5′ cap, poly-A tail, 5′ UTR, 3′ UTR, and ORF. In in vitro transcription (IVT), the natural 5′ cap (m7G) has the risk of uncapping or becoming inactive mRNA because of competitive incorporation between m7G and GTP. Pasquinelli et al. found that m7G was bound to the first nucleotide of the reverse orientated mRNA by 3′-5′ phosphodiester interaction, and one-third of the natural 5′ cap was incorporated in the reverse orientation [31]. Such reverse caps are unlikely to be recognized by translation initiation factor 4E (eIF4E). In 2001, Stepinski et al. designed a novel cap analog (P(1)-3′-O,7-dimethylguanosine-5′ P3-guanosine-5′ triphosphate), called anti-reverse cap analogs (ARCAs), which are not able to incorporate in the reverse orientation and skip degeneration by Dcp2 (mRNA-decaying enzymes) [32]. These significantly enhance translation efficiency and modRNA stability [32,33,34,35,36]. The natural poly-A tail can aid mRNA stability, thus, the optimal length of 120–150 nucleotides can be added to modRNA in in vitro transcription. There are two methods for poly-A tail addition: encoding the poly-A tail from the template vector or enzymatically adding adenine nucleotides using recombinant poly(A) polymerase [35,37]. The natural UTRs can enhance the stability and translation efficiency of modRNA depending on their sequences. Therefore, highly stabilizing UTRs derived from α/β-globin genes are used for desired modRNA [38].
Unmodified mRNAs such as ssRNA or dsRNA can induce interferon production through the activation of Toll-like receptor 7 (TLR7) and TLR8, thus limiting their extensive applications due to highly immunogenic properties [39,40,41,42]. Substantial modification in mRNA was reported [43], whereas only a small subset of modified nucleotides such as 5-methyluridine (5mU), 5-methylcytidine (5mC), pseudouridine (ψU), N6-methyladenosine (6mA), N1-methylpseudouridine (1mψU), and 5-methoxyuridine (5moU) were demonstrated to reduce innate immune responses and enhance translation efficiency and stability [34,35,38,44,45,46]. In 2010, 5mC and ψU were first used in the IVT modRNA-based reprogramming by inducing transgene expression of Oct4, SOX2, KLF4, c-Myc, and Lin-28 in fibroblasts for iPSC generation. The modRNA showed high transfection efficiency and high protein expression, whereas immune responses and cytotoxicity did not occur in the host [47]. In 2015, 1mψU was first incorporated into modRNA, which can reduce innate immune responses through inhibiting TLR3 activation [48]. In 2016, 5moU-modified mRNA was first investigated and exhibited high protein expression and extensive half-life [49]. In 2017, the 5mC was reported to increase mRNA-binding affinity and mRNA export through recognition by the mRNA export adaptor ALYREF [50]. Taken together, modRNA improves the disadvantages of original IVT mRNA and displays some advantages such as highly transient protein expression, very low immunogenicity, stability, improved delivery, and no genomic integration [51,52,53]. It is not yet suitable for the long-term expression of proteins [53].
Based on the understanding of modified mRNA, it can be designed to be synthesized in vitro by the robust T7 RNA polymerase-mediated transcription from a linearized DNA template, which incorporates UTRs such as 5′ UTR containing a Kozak sequence and α-Globin 3′ UTR. ARCA, modified nucleotides (5-Methylcytidine-5′-Triphosphate and Pseudouridine (ψ)-5′-Triphosphate) and poly-A tail could be incorporated in the mRNA to enhance the stability and reduce the immune response of host cells. DNase I could be added to digest the DNA template after modRNA synthesis. Phosphatase could be added to remove the 5′ triphosphates at the end of the modRNA to further reduce innate immune response in mammalian cells. After purification, the modRNA could be diluted in the buffer to the desired concentration for further application.
3. modRNA Delivery for Reprogramming
Some researchers used the modRNA strategy to overexpress pluripotent factors in somatic cells and subsequently reprogram cells into iPSCs. The modRNA-based reprogramming method contains the in vitro transfection of modRNA in somatic cells [47,52,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75]. However, the cell membrane is a natural barrier for modRNA delivery. It is composed of a lipid bilayer including zwitterionic and negatively charged phospholipids [76]. There are two major delivery methods such as cationic lipid and electroporation used in modRNA-based reprogramming, listed in Table 1. The cationic lipids contain a positively charged head group and one or two hydrocarbon chains [77,78,79,80]. The positively charged head group controls the association between the lipid and phosphate backbone of the RNA and assists RNA condensation [81,82]. Cationic lipids are usually formulated with a helper lipid or a neutral co-lipid and result in liposomes with a positive surface charge when in water. The positively charged liposomes spontaneously interact with the negatively charged RNA and result in an RNA–liposome complex. This complex can fuse with cell membrane and enter the cytoplasm through endocytosis [83]. The modRNA of pluripotent factors immediately translate their target proteins and induce somatic cell reprogramming. The commercial RNAiMAX transfection reagent was most used in somatic cell reprogramming with modRNA, and other reagents such as Lipofectamine and Stemfect reagents were also used. Therefore, the cationic lipid-mediated delivery is a simple and fast method for transferring modRNA into somatic cells [47,54,56,57,58,59,60,61,62,63,64,65,66,68,69,70,71]. It was applied in somatic cell reprogramming such as adherent fibroblasts and mesenchymal stem cells [47,54,55,56,57,58,59,60,61,62,63,64,65,66,67,69,70,71].

Table 1.
Summary of reported iPSC generation using modRNA-based reprogramming.
Another modRNA delivery method used in reprogramming is electroporation [55,56]. This is a physical transfection method in which an electrical pulse is used to disturb the phospholipid bilayer of the cell membrane and results in transient pores in the membrane. The charged modRNA can be simultaneously driven across the cell membrane through the pores in a manner similar to electrophoresis when the electric potential moves across the membrane [84]. Electroporation is a highly efficient method for transferring exogenous RNA into many cell types and even blood cells that are difficult to transfect [85]. However, a high voltage pulse causes substantial cell death and only a small part of cells can survive through successful cell membrane repair. Thus, the electroporation method needs the use of greater quantities of cells and instruments compared with cationic lipid methods.
The switch of somatic cell reprogramming needs a substantial number of pluripotent factor expressions, which drive cell transformation into pluripotent states [2,7,86]. Therefore, modRNA-based reprogramming needs repeated transfections to induce overexpression of pluripotent proteins and further promote reprogramming due to the transient expression of modRNA transfections. In 2010, Warren et al. first used modRNAs including Oct4, SOX2, KLF4, c-Myc, and Lin-28 to reprogram various fibroblasts into iPSCs after 17 consecutive transfections of modRNA [47]. Although the modRNA-based reprogramming has a higher safety level with no genome insertion compared with integrative methods, the protocol is relatively complicated because of daily transfections over 2 weeks. Researchers gradually modified the protocol including a reduction in transfection numbers (1~5) and an increase in RNA dosages (43~5 μg) and successfully generated iPSCs using the modRNA transfection strategy [54,55,56,57,67,69]. The modRNA was also applied in cell differentiation; modified MYOD1 was able to induce fibroblasts to differentiate into footprint-free myoblasts. The transformation efficiency increases correlated with higher amounts of transfected modRNA [66].
In addition, another RNA delivery system such as graphene oxide-polyethylenimine (GO-PEI) was used for the reprogramming of somatic cells into iPSCs. Choi et al. used the delivery vehicle (GO-PEI) to avoid repetitive daily transfection of mRNA [67]. GO-PEI complexes were demonstrated to be very effective for transferring mRNA of pluripotent factors and protecting from mRNA degradation by RNase. GO-PEI/mRNA complexes-treated fibroblasts significantly enhanced the reprogramming efficiency and successfully generated human and rat iPSCs without daily transfection.
4. Modified mRNA-Based Reprogramming
The reprogramming efficiency can be as high as up to 4%, and colony formation begins around 14–18 days using modRNA-based technology. The modified mRNA was improved by highly transient protein expression, very low immunogenicity, stability, and no genomic integration. There are four combinations of various modifications that were used in the modRNA-based reprogramming, listed in Table 1. The modifications of mRNA (5mC, ψU, 5′ UTR containing a Kozak sequence, α-Globin 3′ UTR, Poly-A tail, ARCA) were frequently applied in nine studies [47,58,60,61,64,66,68,70,71]. Secondly, RNA modifications including 5mC, ψU, and ARCA were used in four studies [59,62,63,65].
The combinations of pluripotent factors such as OSKM and LIN28 were the most chosen for reprogramming modRNAs, and were used in seven studies. These modRNAs were usually mixed in a molar ratio of 3:1:1:1:1 (O:S:K:M:LIN28) [47,60,62,63,70,71]. NANOG was included in these five factors in four studies as well. Interestingly, microRNA also was combined with these modRNAs of pluripotent factors for enhancing the efficiency and kinetics of somatic cell reprogramming [65,70,71]. Lee et al. found that the numbers of daily modRNA transfections required for reprogramming were reduced from 17 [47] to 11 because of the combination with the microRNA cocktail (microRNA (miR)302a-d and miR367) [65]. The miRNAs-367/302s family of miRNAs has been demonstrated to induce pluripotency in fibroblasts [87]. The miR302/367 cluster can rapidly and efficiently reprogram mouse and human fibroblasts to a pluripotent state without exogenous transcription factors. The miR302/367-mediated somatic reprogramming occurs through the activation of Oct4 gene expression and suppression of Hdac2.
The modRNA-based reprogramming technology is utilized in various somatic cells such as fibroblasts [47,54,55,56,57,58,60,61,63,64,65,66,67,69,70,71], adipose-derived stem cells (ADSCs) [59], bone marrow-derived mesenchymal stem cells (BMSCs) [62], and amniotic fluid stem cells [68] for iPSC generation. Thirteen studies reported fibroblast reprogramming using modRNAs encoding pluripotent factors and generated disease iPSCs from patients with diseases such as cystic fibrosis [47], Huntington’s disease [56], DiGeorge syndrome [63], low-density lipoprotein receptor (LDLR) deficiency [64], and Down syndrome [70]. The β-thalassemia iPSCs were also generated from BMSCs using the modRNA platform [62].
The literature listed in Table 1 reported that modRNA-induced iPSCs differentiate into three germ layers in addition to five studies without evidence [54,55,57,60,71]. The modRNA-derived iPSCs (RiPSCs) also can further differentiate into other cell types in vitro such as myogenic cells [47], cardiomyocytes [58,63,68], hepatocytes [61,64], MSCs [64], and hematopoietic progenitors [62]. Taken together, RiPSCs have great promise in cardiac repair [58,63,68] and a new drug discovery [61,64], especially genetic correction of specific defects such as β-thalassemia [62,88,89] and low-density lipoprotein receptor (LDLR) deficiency familial hypercholesterolemia [64].
To date, modRNA-based reprogramming has been performed mostly on fibroblasts. However, blood cells have acted as a popular source for iPSC generation due to easy harvesting. The application of modRNA technology in the reprogramming of blood cells into iPSCs seems to be limited. Although the reprogramming efficiency of episomal plasmid-induced iPSC generation is lower than that of RNA systems, much literature has shown that various blood cells such as progenitors, lymphoblasts, monocytes, B, and T cells can be successfully reprogrammed using the episomal system combined with an electroporation delivery method [13]. In transfection numbers, the RNA system needs consecutive daily transfections for more than two weeks and lots of hands-on time, whereas the episomal system only needs a one-time plasmid transfection. Moreover, blood cells are well known to be difficult to transfect because of resistance with cationic reagents. Although the electroporation method can facilitate RNA delivery into somatic cells, repeat transfections can cause substantial death of somatic cells. Therefore, reduction in transfection numbers is required for modRNA-induced blood cell reprogramming to pluripotency. The increase in transfection efficiency can reduce transfection numbers by using an improved delivery method such as GO-PEI [67]. Additional reprogramming factors such as engineered chimeric pluripotent factors with extra transactivating domains can be incorporated into the modRNA system for enhancing reprogramming efficiency and then reducing transfection numbers. The co-transfection of microRNA also can synergize with pluripotent proteins to promote reprogramming and pluripotency [65,70,71]. In addition, the episomal system only prepares one plasmid including defined pluripotent factors such as OSKM, whereas the RNA system needs four separate OSKM plasmids to be prepared for in vitro transcription. Although the RNA system is more complicated and has a higher cost than the episomal system, RNA-based reprogramming holds great promise in the unambiguous footprint for future clinical-grade iPSC production. It is valuable for its use in overcoming the difficulty of reprogramming blood cell lineages with modRNA.
5. modRNA Applications in Cell Differentiation
iPSCs possess pluripotent properties and differentiate into any cell type in the body, which makes them potential therapeutic drugs in regenerative medicine. In addition to RiPSCs-differentiated therapeutic cells, modRNA can directly reprogram somatic cells into therapeutic cells bypassing iPSC generation. The modRNA-induced cell differentiation such as myoblasts, vascular endothelial cells, cardiomyocytes, endothelium, insulin-producing cells, neurons, and bone regeneration used in the therapeutic investigation are summarized in Table 2. Although modRNA-based reprogramming has advantages such as higher efficiency and safety than the viral methods, it usually requires repeat transfections of modRNA. In the application of pluripotent stem cell-derived therapeutic cells in vitro, the number of modRNA transfections required in the cell differentiation are reduced to 1~3 times. The MYOD modRNA can differentiate human RiPSCs into myogenic cells in vitro [47]. The vascular endothelial growth factor (VEGF) modRNA induces not only the vascular endothelial specification from ISL1+ heart progenitors in vitro but also promotes engraftment, proliferation, and survival of the human ISL1+ progenitors in vivo [90]. The modRNA encoding transcription factor ETV2 drove transient protein expression and was indeed sufficient to reprogram human pluripotent stem cells such as ESCs and iPSCs into hemogenic endothelial cells compared with lentivirus-mediated continuous ETV2 protein overexpression [91,92]. The ETV2 modRNA-induced hematoendothelial progenitors from human iPSCs were further cultured with GM-CSF, FGF-2, and UM171 to amplify myelomonocytic progenitors, followed by treatment with G-CSF and retinoic acid agonist Am580 to induce neutrophil differentiation [93,94]. The pancreatic-duodenal (PDX1) modRNA was also used in the driven differentiation of insulin-producing cells from human ESCs in vitro [95] and mouse pancreas-derived MSCs [96], which displays a promising approach for cell-based diabetic therapy. A cocktail of five transcription factors (NEUROG1, NEUROG2, NEUROG3, NEUROD1, and NEUROD2) as modRNAs can differentiate hPSCs into motor neurons in 7 days, analyzed by calcium imaging and electrophysiology [97]. In bone regeneration, the bone morphogenetic proteins (BMP-2) play a critical role in the development of bone and cartilage and induce osteoblast differentiation in a variety of cell types [98,99]. The modRNA encoding BMP-2 protein was transferred into BMSCs combined with biomaterials such as micro-macro biphasic calcium phosphate (MBCP) and resulted in cellular osteogenesis [100].
Table 2.
Summary of modRNA-induced transdifferentiation used in therapeutic investigations.
The modRNA-related cell differentiation has been applied in animal models including myocardial infarction and bone defect, which are summarized in Table 2. A myocardial infarction (MI), commonly known as a heart attack, occurs when blood flow decreases or stops to a part of the heart, resulting in injury to the heart muscle. This is usually caused by a blockage of one or more of the coronary arteries [101]. Intramyocardial injection of VEGF modRNA (100 μg/heart) resulted in the expansion and directed differentiation of endogenous heart progenitors and further improved heart function in a mouse myocardial infarction model [102]. In addition, the differentiation of heart WT1+ epicardial cells requires the activation of insulin-like growth factor 1 receptor (IGF1R). The intramyocardial injection of IGF modRNA drives epicardial adipose tissue formation after myocardial injury [103]. The intramyocardial delivery of Brachyury modRNA-induced cardiopoietic stem cells from adipose-derived stem cells (ADSCs) can improve cardiac performance and protected against decompensated heart failure [104]. A microencapsulated modified messenger RNA (M3RNA) platform was used for Brachyury modRNA delivery to achieve a nonintegrating and viral-free transfection [105]. In application for bone defect, the modRNA combined with biomaterial was coated on the titanium implants, which was designed to produce BMP-2 protein for bone regeneration. In the bone defect models such as rat femur and calvarial defects, BMP-2 modRNA combined with different biomaterials was implanted into defects and resulted in bone regeneration and the acceleration of bone healing [106,107,108,109].
In the application of transdifferentiation in vitro, the number of modRNA transfections required in the specification of cell fate ranged 1~14 times. There are two studies that used MYOD modRNA to generate myoblasts directly differentiated from fibroblasts or MSCs in vitro, suggesting a potentially clinically relevant source of autologous cells for cardiac repair [66,110]. The delivery technology C-Lipo including the polyarginine-fused heart-targeting peptide and lipofectamine complex was applied in the encapsulation of three modRNAs (Gata4, Mef2c, and Tbx5), which dramatically enhanced modRNA transfection and resulted in the direct reprogramming of cardiac fibroblasts toward cardiomyocytes [111]. Insulin-producing cells can be directly reprogrammed from pancreatic exocrine cells AR42J [112] by the transfection of PDX1 modRNA. In the application of neural transdifferentiation in vitro, a set of modRNA cocktails hold great promise in nerve regeneration and diseases. A cocktail of two transcription factors (SOX2 and PAX6) as modRNAs can transdifferentiate adult fibroblasts into neural precursor cells, which generate immature GABAergic or glutamatergic neuronal phenotypes in conjunction with astrocytes [113].
The modRNA-related transdifferentiation has been applied in animal models including ischemia and diabetes, which are summarized in Table 2. In applications for ischemia, a study showed that ETV2 modRNA combined with hypoxia (5% oxygen) for 14 days can produce functional endothelial progenitor cells (EPCs) that can form capillary-like structures from skin fibroblasts. The transfer of functional EPCs could improve hindlimb ischemia in a mouse model [114]. In application for diabetes, the transcription factor MafA (musculoaponeurotic fibrosarcoma oncogene homolog A) was applied for modRNA and was sufficient to drive β-cell transdifferentiation from human pancreatic duct-derived cells. The functional β cells can secret insulin and C-peptide in response to glucose. Transplantation of the β cells into diabetic SCID-beige mice mitigated hyperglycemia through their functional glucose-responsive insulin secretion [115].
6. Safety
To facilitate the translation of iPSC technology to clinical medicine, an unambiguous footprint reprogramming method is required. The mRNA-based reprogramming technology toward iPSC generation and transdifferentiation is the most unambiguously footprint-free, effective, and safe method. It eliminates bio-containment concerns associated with viral integrative vectors, which need to screen cells for several weeks to confirm that viral material has been completely removed during cell passaging. However, exogenous mRNA was demonstrated to induce innate immune responses through the activation of type I interferons when fibroblasts were transfected with mRNA in vitro [116]. The mRNA was also reported to elicit adaptive immune responses against cancer progress by intramuscular injection of carcinoembryonic antigen mRNA [117]. The exogenous mRNA causes inflammatory immune responses and suppresses mRNA replication due to triphosphorylated mRNA and double-stranded RNA, which can be recognized by TLR 3, 7, and 8. These TLRs initiate the expression of inflammatory cytokines such as type I interferons, IL-6, IL-12, and TNF-α [45]. The mRNA delivery into cells is through endocytosis and results in the RNA-containing endosomes in cytoplasm. These TLRs are expressed on endosomal membranes and recognize ssRNA [118].
In current years, some modifications of mRNA were found to reduce immune responses compared with unmodified mRNA. The activation of TLR 3, 7, 8, and retinoic acid-inducible gene I can be inhibited by 5mC, 6mA, 5mU, ψU, and 2TU [34,44,45,46]. RNA-dependent protein kinase-mediated the immune responses and translation suppression by phosphorylating α subunit of translation initiation factor 2 can be limited when mRNA contains modified nucleotides such as ψU and 5mC [119]. The activation of innate immunity-associated components such as interferon-induced enzymes 2′-5′-oligoadenylate synthetase and RNase L can be reduced by ψU [120]. When modified mRNAs were used combined with the B18R interferon inhibitor for somatic reprogramming, it was possible to keep daily modRNA transfections for two weeks without eliciting significant immune responses [47,121]. Although a very low level of inflammation may occur when modRNA and B18R were applied, this might facilitate reprogramming. Reports showed that deliberate control of TLR signaling-mediated inflammation is critical in the success of mRNA-induced somatic reprogramming [122,123,124,125].
On the other hand, modRNA delivery methods such as cationic lipid and electroporation were the most used for somatic reprogramming in vitro, shown in Table 1. However, electroporation utilizes high voltage pulses for modRNA delivery into cells and causes substantial cell death, and only a small number of cells can survive. Cationic lipids are also associated with cell toxicity, which disrupts the integrity of a membrane structure due to the detergent property [126]. In high concentrations, a lipoplex consisting of cationic lipid molecules can trigger cell lysis and necrosis [127]. Cationic groups may interact with cellular protein kinase C to induce cell toxicity [128]. Moreover, the modRNA-based reprogramming for iPSC generation needs repeat RNA transfections for more than two weeks, which may cause a lot of cell death. Therefore, an improved modRNA delivery method is required for somatic reprogramming. The GO-PEI was used to avoid repetitive daily transfection of modRNA and transfection was reduced to three times [67]. In addition, microRNA such as miR302/367 was also applied to facilitate efficient reprogramming and to reduce transfection numbers [65,70,71].
The modRNA technology has also been applied in myocardial infarction animal models, which can improve cardiac dysfunction and long-term survival when VEGF modRNA are delivered with a lipid carrier [102]. However, a lipid-based delivery system causes infusion-associated hypersensitivity reactions, tissue injury, and local innate immune responses [129,130,131]. Recently, reports demonstrated that modRNA can be expressed in the heart without using lipid carriers [132] and can even improve cardiac function after intracardiac injection of VEGF modRNA [133]. In addition, although modRNA technology is a rapid and effective method to produce target proteins in vitro and in vivo, the protein levels encoded by modRNA cannot be precisely predicted. This weak point may be improved through the repeated administration of different dosages of modRNA for predicting protein concentrations.
7. Conclusions
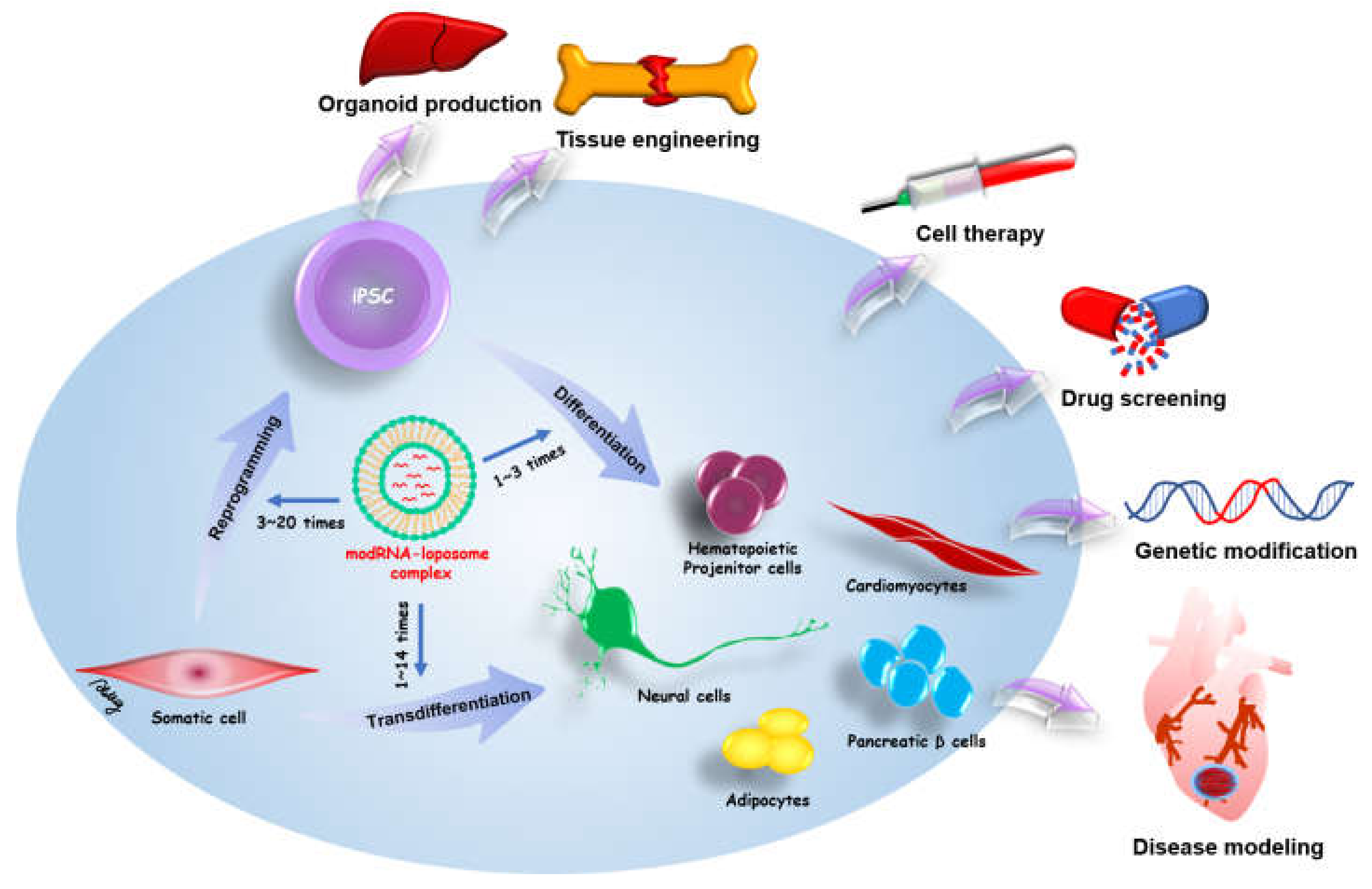
Exogenous proteins can drive somatic cell reprogramming and transdifferentiation for iPSC-related medicine. Therefore, a safe and effective method is required for use in research and medicine. Initially, the most common method for exogenous protein expression depended on viral integrative methods, which have the risk of genomic mutation through the integration of transgenes into the host genome. The modified mRNA of in vitro transcription provides non-integrative and footprint technology for target protein expression. modRNA can directly drive specific cell fate and cell reprogramming from various somatic cells. Moreover, a modRNA cocktail can be designed to simultaneously induce multiple different proteins in somatic cells. The in vitro transcription has benefits for greater flexibility, control over stoichiometric ratios, and dose titrations. The modRNA-based reprogramming and transdifferentiation hold great promise in iPSC-related regenerative medicine and disease modeling. Schematic overview of modRNA applications in cell fate conversion and potential therapies of modRNA-induced cells is shown in Figure 2.
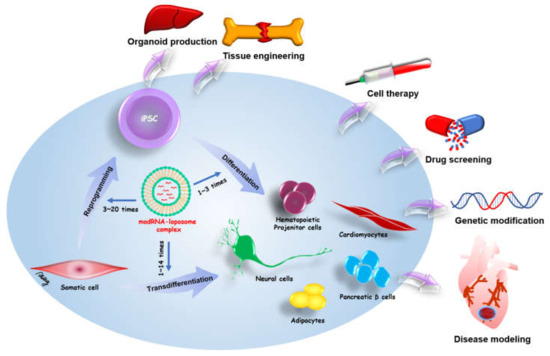
Figure 2.
Schematic overview of modRNA applications in cell fate conversion and potential therapies of modRNA-induced cells. modRNA can be applied for somatic reprogramming to pluripotency (transfection numbers: 3~20 times) and stem cell differentiation to therapeutic cells (transfection numbers: 1~3 times). modRNA also can induce the transdifferentiation of somatic cells to distinct therapeutic cells (transfection numbers: 1~14 times). In addition, one direct injection of therapeutic modRNAs to defective organs may promote tissue regeneration and repair.
Funding
This work was supported by grants from Chang Gung Medical Foundation, Chang Gung Memorial Hospital, Taiwan (CMRPG1H0083) and Ministry of Science and Technology, Taiwan (109-2314-B-182A-056-MY3 and MOST 110-2314-B-182A-137-MY3).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares that there is no conflict of interest.
References
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amabile, G.; Meissner, A. Induced pluripotent stem cells: Current progress and potential for regenerative medicine. Trends Mol. Med. 2009, 15, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Narsinh, K.H.; Plews, J.; Wu, J.C. Comparison of human induced pluripotent and embryonic stem cells: Fraternal or identical twins? Mol. Ther. 2011, 19, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.L. Human induced pluripotent stem cell-derived exosomes as a new therapeutic strategy for various diseases. Int. J. Mol. Sci. 2021, 22, 1769. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.G.; Daley, G.Q. Induced pluripotent stem cells in disease modelling and drug discovery. Nat. Rev. Genet. 2019, 20, 377–388. [Google Scholar] [CrossRef]
- Park, I.H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Fusaki, N.; Ban, H.; Nishiyama, A.; Saeki, K.; Hasegawa, M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 348–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, H.; Nishishita, N.; Fusaki, N.; Tabata, T.; Saeki, K.; Shikamura, M.; Takada, N.; Inoue, M.; Hasegawa, M.; Kawamata, S.; et al. Efficient generation of transgene-free human induced pluripotent stem cells (iPSCs) by temperature-sensitive Sendai virus vectors. Proc. Natl. Acad. Sci. USA 2011, 108, 14234–14239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Muench, M.O.; Fusaki, N.; Beyer, A.I.; Wang, J.; Qi, Z.; Yu, J.; Kan, Y.W. Blood cell-derived induced pluripotent stem cells free of reprogramming factors generated by Sendai viral vectors. Stem Cells Transl. Med. 2013, 2, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.L.; Loh, C.Y.Y. Episomal induced pluripotent stem cells: Functional and potential therapeutic applications. Cell Transplant. 2019, 28, 112S–131S. [Google Scholar] [CrossRef]
- Loh, C.Y.Y.; Wang, A.Y.L.; Kao, H.K.; Cardona, E.; Chuang, S.H.; Wei, F.C. Episomal induced pluripotent stem cells promote functional recovery of transected murine peripheral nerve. PLoS ONE 2016, 11, e0164696. [Google Scholar] [CrossRef] [Green Version]
- Cobb, M. Who discovered messenger RNA? Curr. Biol. 2015, 25, R526–R532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, M. Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles. Microbiol. Rev. 1983, 47, 1–45. [Google Scholar] [CrossRef]
- Lopez-Lastra, M.; Rivas, A.; Barria, M.I. Protein synthesis in eukaryotes: The growing biological relevance of cap-independent translation initiation. Biol. Res. 2005, 38, 121–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, S.E.; Hillner, P.E.; Vale, R.D.; Sachs, A.B. Circularization of mRNA by eukaryotic translation initiation factors. Mol. Cell 1998, 2, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Wahle, E.; Keller, W. The biochemistry of polyadenylation. Trends Biochem. Sci. 1996, 21, 247–250. [Google Scholar] [CrossRef]
- Choi, Y.S.; Patena, W.; Leavitt, A.D.; McManus, M.T. Widespread RNA 3′-end oligouridylation in mammals. RNA 2012, 18, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Korner, C.G.; Wahle, E. Poly(A) tail shortening by a mammalian poly(A)-specific 3′-exoribonuclease. J. Biol. Chem. 1997, 272, 10448–10456. [Google Scholar] [CrossRef] [Green Version]
- Sachs, A.; Wahle, E. Poly(A) tail metabolism and function in eucaryotes. J. Biol. Chem. 1993, 268, 22955–22958. [Google Scholar] [CrossRef]
- Barrett, L.W.; Fletcher, S.; Wilton, S.D. Regulation of eukaryotic gene expression by the untranslated gene regions and other non-coding elements. Cell Mol. Life Sci. 2012, 69, 3613–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.; Pal, J.K. Role of 5′- and 3′-untranslated regions of mRNAs in human diseases. Biol. Cell 2009, 101, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, P.; Mebrahtu, A.; Barzadd, M.M.; Thalén, N.; Klanert, G.; Weinguny, M.; Sandegren, A.; Su, C.; Hatton, D.; Borth, N.; et al. Systematic use of synthetic 5′-UTR RNA structures to tune protein translation improves yield and quality of complex proteins in mammalian cell factories. Nucleic Acids Res. 2020, 48, e119. [Google Scholar] [CrossRef]
- Yergert, K.M.; Doll, C.A.; O’Rouke, R.; Hines, J.H.; Appel, B. Identification of 3′ UTR motifs required for mRNA localization to myelin sheaths in vivo. PLoS Biol. 2021, 19, e3001053. [Google Scholar] [CrossRef]
- Rasekhian, M.; Roohvand, F.; Habtemariam, S.; Marzbany, M.; Kazemimanesh, M. The role of 3′UTR of RNA viruses on mRNA stability and translation enhancement. Mini. Rev. Med. Chem. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Shabalina, S.A.; Ogurtsov, A.Y.; Spiridonov, N.A. A periodic pattern of mRNA secondary structure created by the genetic code. Nucleic Acids Res. 2006, 34, 2428–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, L.; Burge, C.B. Widespread selection for local RNA secondary structure in coding regions of bacterial genes. Genome Res. 2003, 13, 2042–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquinelli, A.E.; Dahlberg, J.E.; Lund, E. Reverse 5′ caps in RNAs made in vitro by phage RNA polymerases. RNA 1995, 1, 957–967. [Google Scholar]
- Stepinski, J.; Waddell, C.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Synthesis and properties of mRNAs containing the novel “anti-reverse” cap analogs 7-methyl(3′-O-methyl)GpppG and 7-methyl (3′-deoxy)GpppG. RNA 2001, 7, 1486–1495. [Google Scholar] [PubMed]
- Mockey, M.; Gonçalves, C.; Dupuy, F.P.; Lemoine, F.M.; Pichon, C.; Midoux, P. mRNA transfection of dendritic cells: Synergistic effect of ARCA mRNA capping with Poly(A) chains in cis and in trans for a high protein expression level. Biochem. Biophys. Res. Commun. 2006, 340, 1062–1068. [Google Scholar] [CrossRef]
- Kormann, M.S.D.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A.; et al. Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef]
- Sahin, U.; Kariko, K.; Tureci, O. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef] [PubMed]
- Hadas, Y.; Katz, M.G.; Bridges, C.R.; Zangi, L. Modified mRNA as a therapeutic tool to induce cardiac regeneration in ischemic heart disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Quabius, E.S.; Krupp, G. Synthetic mRNAs for manipulating cellular phenotypes: An overview. New Biotechnol. 2015, 32, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Athirasala, A.; Menezes, P.P.; Ashwanikumar, N.; Zou, T.; Sahay, G.; Bertassoni, L.E. Messenger RNA delivery for tissue engineering and regenerative medicine applications. Tissue Eng. Part A 2019, 25, 91–112. [Google Scholar] [CrossRef]
- Isaacs, A.; Cox, R.A.; Rotem, Z. Foreign nucleic acids as the stimulus to make interferon. Lancet 1963, 2, 113–116. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Sousa, C.R. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- Machnicka, M.A.; Milanowska, K.; Oglou, O.O.; Purta, E.; Kurkowska, M.; Olchowik, A.; Januszewski, W.; Kalinowski, S.; Dunin-Horkawicz, S.; Rother, K.M.; et al. MODOMICS: A database of RNA modification pathways—2013 update. Nucleic Acids Res. 2013, 41, D262–D267. [Google Scholar] [CrossRef]
- Haque, A.K.M.A.; Dewerth, A.; Antony, J.S.; Riethmüller, J.; Schweizer, G.R.; Weinmann, P.; Latifi, N.; Yasar, H.; Pedemonte, N.; Sondo, E.; et al. Chemically modified hCFTR mRNAs recuperate lung function in a mouse model of cystic fibrosis. Sci. Rep. 2018, 8, 16776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA recognition by Toll-like receptors: The impact of nucleoside modification and the evolutionary origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Durbin, A.F.; Wang, C.; Marcotrigiano, J.; Gehrke, L. RNAs Containing Modified Nucleotides Fail To Trigger RIG-I Conformational Changes for Innate Immune Signaling. mBio 2016, 7, e00833. [Google Scholar] [CrossRef] [Green Version]
- Warren, L.; Manos, P.D.; Ahfeldt, T.; Loh, Y.H.; Li, H.; Lau, F.; Ebina, W.; Mandal, P.K.; Smith, Z.D.; Meissner, A.; et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 2010, 7, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Andries, O.; Cafferty, S.M.; Smedt, S.C.D.; Weiss, R.; Sanders, N.N.; Kitada, T. N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J. Control. Release 2015, 217, 337–344. [Google Scholar] [CrossRef]
- Li, B.; Luo, X.; Dong, Y. Effects of Chemically Modified Messenger RNA on Protein Expression. Bioconjug. Chem. 2016, 27, 849–853. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-methylcytosine promotes mRNA export—NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef] [Green Version]
- Kariko, K. In vitro-Transcribed mRNA Therapeutics: Out of the Shadows and Into the Spotlight. Mol. Ther. 2019, 27, 691–692. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magadum, A.; Kaur, K.; Zangi, L. mRNA-based protein replacement therapy for the heart. Mol. Ther. 2019, 27, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Yakubov, E.; Rechavi, G.; Rozenblatt, S.; Givol, D. Reprogramming of human fibroblasts to pluripotent stem cells using mRNA of four transcription factors. Biochem. Biophys. Res. Commun. 2010, 394, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Plews, J.R.; Li, J.L.; Jones, M.; Moore, H.D.; Mason, C.; Andrews, P.W.; Na, J. Activation of pluripotency genes in human fibroblast cells by a novel mRNA based approach. PLoS ONE 2010, 5, e14397. [Google Scholar] [CrossRef]
- Antje Arnold, Y.M.N.; Fabian, C.; Wirth, H.; Hans Binder, G.N.; Armstrong, L.; Stolzing, A. Reprogramming of human Huntington fibroblasts using mRNA. Int. Sch. Res. Netw. 2011, 2012, 124878. [Google Scholar] [CrossRef] [Green Version]
- Tavernier, G.; Wolfrum, K.; Demeester, J.; Smedt, S.C.D.; Adjaye, J.; Rejman, J. Activation of pluripotency-associated genes in mouse embryonic fibroblasts by non-viral transfection with in vitro-derived mRNAs encoding Oct4, Sox2, Klf4 and cMyc. Biomaterials 2012, 33, 412–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, L.; Ni, Y.; Wang, J.; Guo, X. Feeder-free derivation of human induced pluripotent stem cells with messenger RNA. Sci. Rep. 2012, 2, 657. [Google Scholar] [CrossRef]
- Heng, B.C.; Heinimann, K.; Miny, P.; Iezzi, G.; Glatz, K.; Scherberich, A.; Zulewski, H.; Fussenegger, M. mRNA transfection-based, feeder-free, induced pluripotent stem cells derived from adipose tissue of a 50-year-old patient. Metab. Eng. 2013, 18, 9–24. [Google Scholar] [CrossRef]
- Mandal, P.K.; Rossi, D.J. Reprogramming human fibroblasts to pluripotency using modified mRNA. Nat. Protoc. 2013, 8, 568–582. [Google Scholar] [CrossRef]
- Sjogren, A.K.M.; Liljevald, M.; Glinghammar, B.; Sagemark, J.; Li, X.Q.; Jonebring, A.; Cotgreave, I.; Brolén, G.; Andersson, T.B. Critical differences in toxicity mechanisms in induced pluripotent stem cell-derived hepatocytes, hepatic cell lines and primary hepatocytes. Arch. Toxicol. 2014, 88, 1427–1437. [Google Scholar] [CrossRef]
- Varela, I.; Karagiannidou, A.; Oikonomakis, V.; Tzetis, M.; Tzanoudaki, M.; Siapati, E.K.; Vassilopoulos, G.; Graphakos, S.; Kanavakis, E.; Goussetis, E. Generation of human beta-thalassemia induced pluripotent cell lines by reprogramming of bone marrow-derived mesenchymal stromal cells using modified mRNA. Cell. Reprogram. 2014, 16, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Durruthy-Durruthy, J.; Briggs, S.F.; Awe, J.; Ramathal, C.Y.; Karumbayaram, S.; Lee, P.C.; Heidmann, J.D.; Clark, A.; Karakikes, I.; Loh, K.M.; et al. Rapid and efficient conversion of integration-free human induced pluripotent stem cells to GMP-grade culture conditions. PLoS ONE 2014, 9, e94231. [Google Scholar] [CrossRef]
- Ramakrishnan, V.M.; Yang, J.Y.; Tien, K.T.; McKinley, T.R.; Bocard, B.R.; Maijub, J.G.; Burchell, P.O.; Williams, S.K.; Morris, M.E.; Hoying, J.B.; et al. Restoration of physiologically responsive low-density lipoprotein receptor-mediated endocytosis in genetically deficient induced pluripotent stem cells. Sci. Rep. 2015, 5, 13231. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.I.; Lee, S.Y.; Hwang, S.Y. Extracellular matrix-dependent generation of integration- and xeno-free iPS cells using a modified mRNA transfection method. Stem Cells Int. 2016, 2016, 6853081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preskey, D.; Allison, T.F.; Jones, M.; Mamchaoui, K.; Unger, C. Synthetically modified mRNA for efficient and fast human iPS cell generation and direct transdifferentiation to myoblasts. Biochem. Biophys. Res. Commun. 2016, 473, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Lee, T.J.; Yang, G.M.; Oh, J.; Won, J.; Han, J.; Jeong, G.J.; Kim, J.; Kim, J.H.; Kim, B.S.; et al. Efficient mRNA delivery with graphene oxide-polyethylenimine for generation of footprint-free human induced pluripotent stem cells. J. Control. Release 2016, 235, 222–235. [Google Scholar] [CrossRef]
- Velasquez-Mao, A.J.; Tsao, C.J.M.; Monroe, M.N.; Legras, X.; Bissig-Choisat, B.; Bissig, K.D.; Ruano, R.; Jacot, J.G. Differentiation of spontaneously contracting cardiomyocytes from non-virally reprogrammed human amniotic fluid stem cells. PLoS ONE 2017, 12, e0177824. [Google Scholar]
- Chen, H.; Zuo, Q.; Wang, Y.; Song, J.; Yang, H.; Zhang, Y.; Li, B. Inducing goat pluripotent stem cells with four transcription factor mRNAs that activate endogenous promoters. BMC Biotechnol. 2017, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Kogut, I.; McCarthy, S.M.; Pavlova, M.; Astling, D.P.; Chen, X.; Jakimenko, A.; Jones, K.L.; Getahun, A.; Cambier, J.C.C.; Pasmooij, A.M.G.; et al. High-efficiency RNA-based reprogramming of human primary fibroblasts. Nat. Commun. 2018, 9, 745. [Google Scholar] [CrossRef] [Green Version]
- McGrath, P.S.; McGarvey, S.S.; Kogut, I.; Bilousova, G. Efficient RNA-based reprogramming of disease-associated primary human fibroblasts into induced pluripotent stem cells. Methods Mol. Biol. 2020, 2117, 271–284. [Google Scholar]
- Kaczmarek, J.C.; Kowalski, P.S.; Anderson, D.G. Advances in the delivery of RNA therapeutics: From concept to clinical reality. Genome Med. 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and challenges in the delivery of mRNA-based vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Guevara, M.L.; Persano, F.; Persano, S. Advances in Lipid Nanoparticles for mRNA-based cancer immunotherapy. Front. Chem. 2020, 8, 589959. [Google Scholar] [CrossRef]
- Trepotec, Z.; Lichtenegger, E.; Plank, C.; Aneja, M.K.; Rudolph, C. Delivery of mRNA therapeutics for the treatment of hepatic diseases. Mol. Ther. 2019, 27, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Balazs, D.A.; Godbey, W. Liposomes for use in gene delivery. J. Drug Deliv. 2011, 2011, 326497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.Y.; Guo, P.; Wen, W.C.; Wong, H.L. Lipid-based nanocarriers for RNA delivery. Curr. Pharm. Des. 2015, 21, 3140–3147. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.H.; Yu, B.; Wang, X.; Lu, Y.; Schmidt, C.R.; Lee, R.J.; Lee, L.J.; Jacob, S.T.; Ghoshal, K. Cationic lipid nanoparticles for therapeutic delivery of siRNA and miRNA to murine liver tumor. Nanomedicine 2013, 9, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- Simberg, D.; Weisman, S.; Talmon, Y.; Barenholz, Y. DOTAP (and other cationic lipids): Chemistry, biophysics, and transfection. Crit. Rev. Ther. Drug. Carrier. Syst. 2004, 21, 257–317. [Google Scholar] [CrossRef]
- Semple, S.C.; Akinc, A.; Chen, J.; Sandhu, A.P.; Mui, B.L.; Cho, C.K.; Sah, D.W.Y.; Stebbing, D.; Crosley, E.J.; Yaworski, E.; et al. Rational design of cationic lipids for siRNA delivery. Nat. Biotechnol. 2010, 28, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Heyes, J.; Palmer, L.; Bremner, K.; MacLachlan, I. Cationic lipid saturation influences intracellular delivery of encapsulated nucleic acids. J. Control. Release 2005, 107, 276–287. [Google Scholar] [CrossRef]
- Kauffman, K.J.; Webber, M.J.; Anderson, D.G. Materials for non-viral intracellular delivery of messenger RNA therapeutics. J. Control. Release 2016, 240, 227–234. [Google Scholar] [CrossRef]
- Shigekawa, K.; Dower, W.J. Electroporation of eukaryotes and prokaryotes: A general approach to the introduction of macromolecules into cells. Biotechniques 1988, 6, 742–751. [Google Scholar]
- Zhao, Y.; Zheng, Z.; Cohen, C.J.; Gattinoni, L.; Palmer, D.C.; Restifo, N.P.; Rosenberg, S.A.; Morgan, R.A. High-efficiency transfection of primary human and mouse T lymphocytes using RNA electroporation. Mol. Ther. 2006, 13, 151–159. [Google Scholar] [CrossRef]
- Buganim, Y.; Faddah, D.A.; Jaenisch, R. Mechanisms and models of somatic cell reprogramming. Nat. Rev. Genet. 2013, 14, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Anokye-Danso, F.; Trivedi, C.M.; Juhr, D.; Gupta, M.; Cui, Z.; Tian, Y.; Zhang, Y.; Yang, W.; Gruber, P.J.; Epstein, J.A.; et al. Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency. Cell Stem Cell 2011, 8, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Papapetrou, E.P.; Lee, G.; Malani, N.; Setty, M.; Riviere, I.; Tirunagari, L.M.S.; Kadota, K.; Roth, S.L.; Giardina, P.; Viale, A.; et al. Genomic safe harbors permit high beta-globin transgene expression in thalassemia induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zheng, C.G.; Jiang, Y.; Zhang, J.; Chen, J.; Yao, C.; Zhao, Q.; Liu, S.; Chen, K.; Du, J.; et al. Genetic correction of beta-thalassemia patient-specific iPS cells and its use in improving hemoglobin production in irradiated SCID mice. Cell Res. 2012, 22, 637–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, K.O.; Zangi, L.; Silva, E.A.; Bu, L.; Sahara, M.; Li, R.A.; Mooney, D.J.; Chien, K.R. Driving vascular endothelial cell fate of human multipotent Isl1+ heart progenitors with VEGF modified mRNA. Cell Res. 2013, 23, 1172–1186. [Google Scholar] [CrossRef] [Green Version]
- Elcheva, I.; Brok-Volchanskaya, V.; Kumar, A.; Liu, P.; Lee, J.H.; Tong, L.; Vodyanik, M.; Swanson, S.; Stewart, R.; Kyba, M.; et al. Direct induction of haematoendothelial programs in human pluripotent stem cells by transcriptional regulators. Nat. Commun. 2014, 5, 4372. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Lin, R.Z.; Hong, X.; Ng, A.H.; Lee, C.N.; Neumeyer, J.; Wang, G.; Wang, X.; Ma, M.; Pu, W.T.; et al. Robust differentiation of human pluripotent stem cells into endothelial cells via temporal modulation of ETV2 with modified mRNA. Sci. Adv. 2020, 6, eaba7606. [Google Scholar] [CrossRef]
- Brok-Volchanskaya, V.S.; Bennin, D.A.; Suknuntha, K.; Klemm, L.C.; Huttenlocher, A.; Slukvin, I. Effective and rapid generation of functional neutrophils from induced pluripotent stem cells using ETV2-modified mRNA. Stem Cell Rep. 2019, 13, 1099–1110. [Google Scholar] [CrossRef] [Green Version]
- Majumder, A.; Suknuntha, K.; Bennin, D.; Klemm, L.; Brok-Volchanskaya, V.S.; Huttenlocher, A.; Slukvin, I. Generation of human neutrophils from induced pluripotent stem cells in chemically defined conditions using ETV2 modified mRNA. STAR Protoc. 2020, 1, 100075. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Yu, L.; Ding, Y.; Guo, X.R.; Yuan, Y.H.; Li, D.S. Gene manipulation of human embryonic stem cells by in vitro-synthesized mRNA for gene therapy. Curr. Gene Ther. 2015, 15, 428–435. [Google Scholar] [CrossRef]
- Guo, X.R.; Wang, X.L.; Li, M.C.; Yuan, Y.H.; Chen, Y.; Zou, D.D.; Bian, L.J.; Li, D.S. PDX-1 mRNA-induced reprogramming of mouse pancreas-derived mesenchymal stem cells into insulin-producing cells in vitro. Clin. Exp. Med. 2015, 15, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Goparaju, S.K.; Kohda, K.; Ibata, K.; Soma, A.; Nakatake, Y.; Akiyama, T.; Wakabayashi, S.; Matsushita, M.; Sakota, M.; Kimura, H.; et al. Rapid differentiation of human pluripotent stem cells into functional neurons by mRNAs encoding transcription factors. Sci. Rep. 2017, 7, 42367. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J.; Debiais, F.; Hay, E. Regulation of human cranial osteoblast phenotype by FGF-2, FGFR-2 and BMP-2 signaling. Histol. Histopathol. 2002, 17, 877–885. [Google Scholar] [PubMed]
- Balmayor, E.R.; Geiger, J.P.; Koch, C.; Aneja, M.K.; Griensven, M.V.; Rudolph, C.; Plank, C. Modified mRNA for BMP-2 in combination with biomaterials serves as a transcript-activated matrix for effectively inducing osteogenic pathways in stem cells. Stem Cells Dev. 2017, 26, 25–34. [Google Scholar] [CrossRef]
- White, H.D.; Chew, D.P. Acute myocardial infarction. Lancet 2008, 372, 570–584. [Google Scholar] [CrossRef] [Green Version]
- Zangi, L.; Lui, K.O.; Gise, A.V.; Ma, Q.; Ebina, W.; Ptaszek, L.M.; Später, D.; Xu, H.; Tabebordbar, M.; Gorbatov, R.; et al. Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nat. Biotechnol. 2013, 31, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zangi, L.; Oliveira, M.S.; Ye, L.Y.; Ma, Q.; Sultana, N.; Hadas, Y.; Chepurko, E.; Später, D.; Zhou, B.; Chew, W.L.; et al. Insulin-like growth factor 1 receptor-dependent pathway drives epicardial adipose tissue formation after myocardial injury. Circulation 2017, 135, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yamada, S.; Shi, A.; Singh, R.D.; Rolland, T.J.; Jeon, R.; Lopez, N.; Shelerud, L.; Terzic, A.; Behfar, A. Brachyury engineers cardiac repair competent stem cells. Stem Cells Transl. Med. 2021, 10, 385–397. [Google Scholar] [CrossRef]
- Singh, R.D.; Hillestad, M.L.; Livia, C.; Li, M.; Alekseev, A.E.; Witt, T.A.; Stalboerger, P.G.; Yamada, S.; Terzic, A.; Behfar, A. M(3)RNA drives targeted gene delivery in acute myocardial infarction. Tissue Eng. Part A 2019, 25, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, S.; Khorsand, B.; Do, A.V.; Hong, L.; Dewerth, A.; Kormann, M.; Ross, R.D.; Sumner, D.R.; Allamargot, C.; Salem, A.K. Chemically modified RNA activated matrices enhance bone regeneration. J. Control. Release 2015, 218, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Balmayor, E.R.; Geiger, J.P.; Aneja, M.K.; Berezhanskyy, T.; Utzinger, M.; Mykhaylyk, O.; Rudolph, C.; Plank, C. Chemically modified RNA induces osteogenesis of stem cells and human tissue explants as well as accelerates bone healing in rats. Biomaterials 2016, 87, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Badieyan, Z.S.; Berezhanskyy, T.; Utzinger, M.; Aneja, M.K.; Emrich, D.; Erben, R.; Schüler, C.; Altpeter, P.; Ferizi, M.; Hasenpusch, G.; et al. Transcript-activated collagen matrix as sustained mRNA delivery system for bone regeneration. J. Control. Release 2016, 239, 137–148. [Google Scholar] [CrossRef]
- Khorsand, B.; Elangovan, S.; Hong, L.; Dewerth, A.; Kormann, M.S.D.; Salem, A.K. A comparative study of the bone regenerative effect of chemically modified RNA encoding BMP-2 or BMP-9. AAPS J. 2017, 19, 438–446. [Google Scholar] [CrossRef]
- Hausburg, F.; Na, S.; Voronina, N.; Skorska, A.; Müller, P.; Steinhoff, G.; David, R. Defining optimized properties of modified mRNA to enhance virus- and DNA- independent protein expression in adult stem cells and fibroblasts. Cell Physiol. Biochem. 2015, 35, 1360–1371. [Google Scholar] [CrossRef]
- Lee, K.; Yu, P.; Lingampalli, N.; Kim, H.J.; Tang, R.; Murthy, N. Peptide-enhanced mRNA transfection in cultured mouse cardiac fibroblasts and direct reprogramming towards cardiomyocyte-like cells. Int. J. Nanomed. 2015, 10, 1841–1854. [Google Scholar]
- Koblas, T.; Leontovyc, I.; Loukotova, S.; Kosinova, L.; Saudek, F. Reprogramming of pancreatic exocrine cells AR42J into insulin-producing cells using mRNAs for Pdx1, Ngn3, and MafA transcription factors. Mol. Ther. Nucleic Acids 2016, 5, e320. [Google Scholar] [CrossRef] [Green Version]
- Connor, B.; Firmin, E.; McCaughey-Chapman, A.; Monk, R.; Lee, K.; Liot, S.; Geiger, J.; Rudolph, C.; Jones, K. Conversion of adult human fibroblasts into neural precursor cells using chemically modified mRNA. Heliyon 2018, 4, e00918. [Google Scholar] [CrossRef] [Green Version]
- Pham, P.V.; Vu, N.B.; Dao, T.T.T.; Le, H.T.N.; Phi, L.T.; Phan, N.K. Production of endothelial progenitor cells from skin fibroblasts by direct reprogramming for clinical usages. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 207–216. [Google Scholar] [CrossRef]
- Corritore, E.; Lee, Y.S.; Pasquale, V.; Liberati, D.; Hsu, M.J.; Lombard, C.A.; Smissen, P.V.D.; Vetere, A.; Bonner-Weir, S.; Piemonti, L.; et al. V-Maf musculoaponeurotic fibrosarcoma oncogene homolog A synthetic modified mRNA drives reprogramming of human pancreatic duct-derived cells into insulin-secreting cells. Stem Cells Transl. Med. 2016, 5, 1525–1537. [Google Scholar] [CrossRef] [Green Version]
- Angel, M.; Yanik, M.F. Innate immune suppression enables frequent transfection with RNA encoding reprogramming proteins. PLoS ONE 2010, 5, e11756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conry, R.M.; LoBuglio, A.F.; Wright, M.; Sumerel, L.; Pike, M.J.; Johanning, F.; Benjamin, R.; Lu, D.; Curiel, D.T. Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 1995, 55, 1397–1400. [Google Scholar] [PubMed]
- Crozat, K.; Beutler, B. TLR7: A new sensor of viral infection. Proc. Natl. Acad. Sci. USA 2004, 101, 6835–6836. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.R.; Muramatsu, H.; Nallagatla, S.R.; Bevilacqua, P.C.; Sansing, L.H.; Weissman, D.; Karikó, K. Incorporation of pseudouridine into mRNA enhances translation by diminishing PKR activation. Nucleic Acids Res. 2010, 38, 5884–5892. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.R.; Muramatsu, H.; Jha, B.K.; Silverman, R.H.; Weissman, D.; Karikó, K. Nucleoside modifications in RNA limit activation of 2′-5′-oligoadenylate synthetase and increase resistance to cleavage by RNase L. Nucleic Acids Res. 2011, 39, 9329–9338. [Google Scholar] [CrossRef] [Green Version]
- Warren, L.; Lin, C. mRNA-Based Genetic Reprogramming. Mol. Ther. 2019, 27, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Sayed, N.; Hunter, A.; Au, K.F.; Wong, W.H.; Mocarski, E.S.; Pera, R.R.; Yakubov, E.; Cooke, J.P. Activation of innate immunity is required for efficient nuclear reprogramming. Cell 2012, 151, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Xu, L.; Gibson, T.M.; Gersbach, C.A.; Sullenger, B.A. Differential effects of toll-like receptor stimulation on mRNA-driven myogenic conversion of human and mouse fibroblasts. Biochem. Biophys. Res. Commun. 2016, 478, 1484–1490. [Google Scholar] [CrossRef]
- Zhou, G.; Meng, S.; Li, Y.; Ghebre, Y.T.; Cooke, J.P. Optimal ROS signaling is critical for nuclear reprogramming. Cell Rep. 2016, 15, 919–925. [Google Scholar] [CrossRef] [Green Version]
- Brady, J.J.; Li, M.; Suthram, S.; Jiang, H.; Wong, W.H.; Blau, H.M. Early role for IL-6 signalling during generation of induced pluripotent stem cells revealed by heterokaryon RNA-Seq. Nat. Cell Biol. 2013, 15, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.Y.; Liu, S.; Wong, H.L. Nanotoxicity: A key obstacle to clinical translation of siRNA-based nanomedicine. Nanomedicine 2014, 9, 295–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Lizarzaburu, M.E.; Kurth, M.J.; Liu, L.; Wege, H.; Zern, M.A.; Nantz, M.H. Cationic lipid polymerization as a novel approach for constructing new DNA delivery agents. Bioconjug. Chem. 2001, 12, 251–257. [Google Scholar] [CrossRef]
- Aberle, A.M.; Tablin, F.; Zhu, J.; Walker, N.J.; Gruenert, D.C.; Nantz, M.H. A novel tetraester construct that reduces cationic lipid-associated cytotoxicity. Implications for the onset of cytotoxicity. Biochemistry 1998, 37, 6533–6540. [Google Scholar] [CrossRef]
- Rudin, C.M.; Marshall, J.L.; Huang, C.H.; Kindler, H.L.; Zhang, C.; Kumar, D.; Gokhale, P.C.; Steinberg, J.; Wanaski, S.; Kasid, U.N.; et al. Delivery of a liposomal c-raf-1 antisense oligonucleotide by weekly bolus dosing in patients with advanced solid tumors: A phase I study. Clin. Cancer Res. 2004, 10, 7244–7251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, T.; Adams, D.; Silva, A.; Lozeron, P.; Hawkins, P.N.; Mant, T.; Perez, J.; Chiesa, J.; Warrington, S.; Tranter, E.; et al. Safety and efficacy of RNAi therapy for transthyretin amyloidosis. N. Engl. J. Med. 2013, 369, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Pollard, C.; Rejman, J.; Haes, W.D.; Verrier, B.; Gulck, E.V.; Naessens, T.; Smedt, S.D.; Bogaert, P.; Grooten, J.; Vanham, G.; et al. Type I IFN counteracts the induction of antigen-specific immune responses by lipid-based delivery of mRNA vaccines. Mol. Ther. 2013, 21, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, N.; Magadum, A.; Hadas, Y.; Kondrat, J.; Singh, N.; Youssef, E.; Calderon, D.; Chepurko, E.; Dubois, N.; Hajjar, R.J.; et al. Optimizing cardiac delivery of modified mRNA. Mol. Ther. 2017, 25, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsson, L.; Clarke, J.C.; Yen, C.; Gregoire, F.; Albery, T.; Billger, M.; Egnell, A.C.; Gan, L.M.; Jennbacken, K.; Johansson, E.; et al. Biocompatible, purified VEGF-A mRNA improves cardiac function after intracardiac injection 1 week post-myocardial infarction in swine. Mol. Ther. Methods Clin. Dev. 2018, 9, 330–346. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).