
Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats

 , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Results
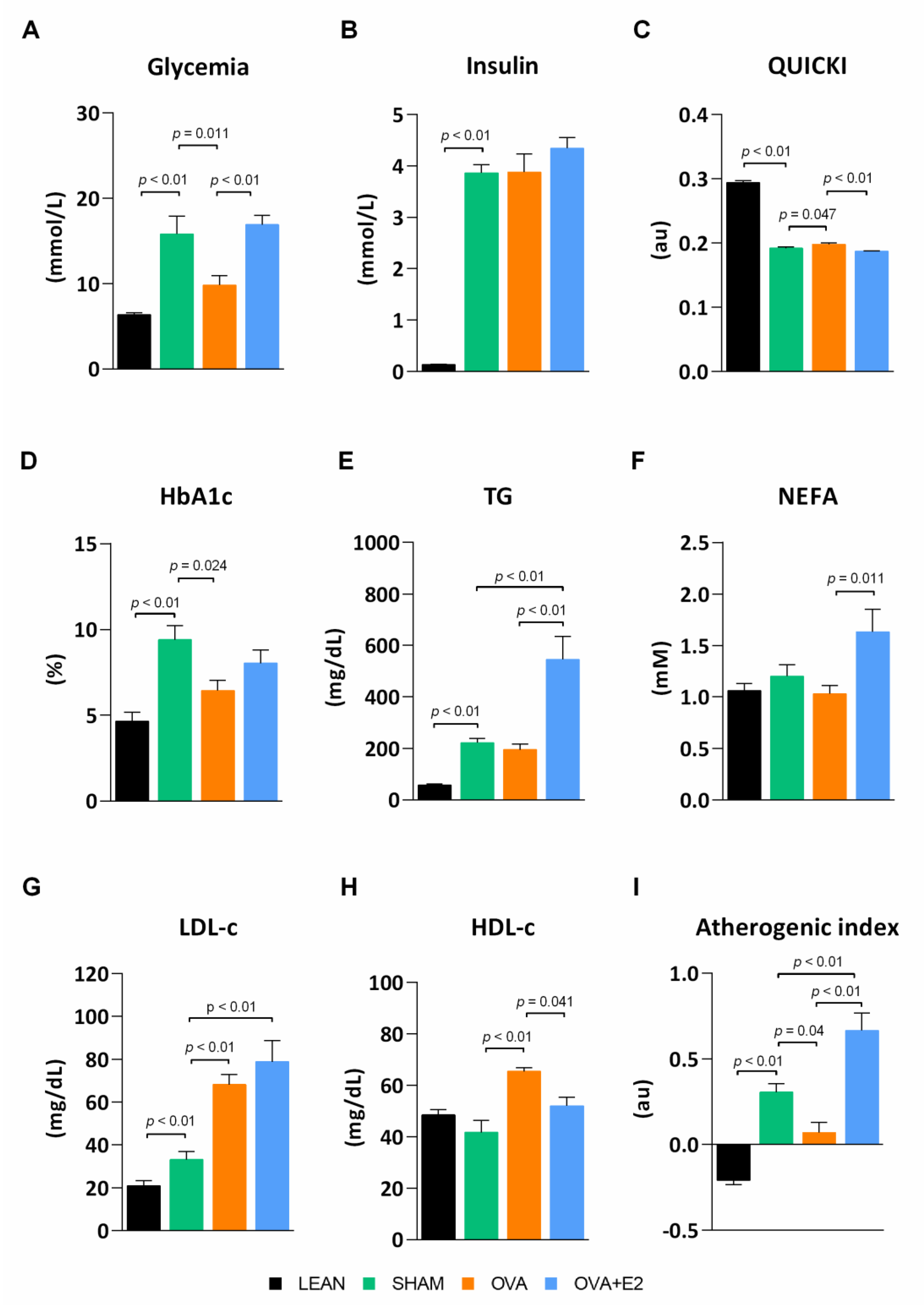
2.1. Effects of Ovariectomy and E2 Supplementation on Biometric Parameters and Blood Metabolic Profile
2.2. Effects of Ovariectomy and E2 Supplementation on WAT Insulin Pathway and Lipid Transporters
2.3. Effects of Ovariectomy and E2 Supplementation on WAT Remodeling
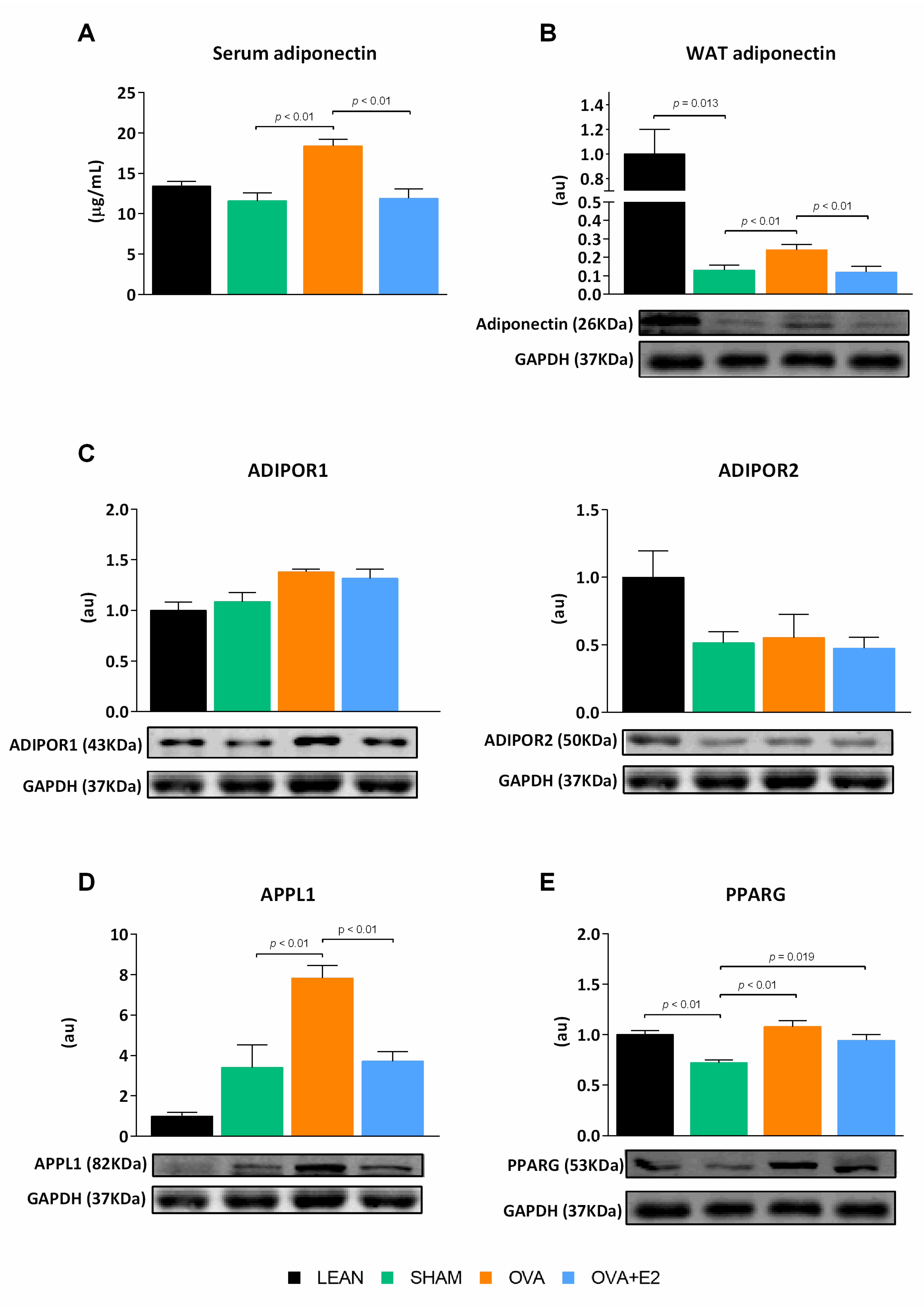
2.4. Effects of Ovariectomy and E2 Supplementation on Expression Levels of Adipokines and Markers of Apoptosis in WAT
2.5. Effects of Ovariectomy and E2 Supplementation on the Expression of PPARG and Elements of Adiponectin Signaling Pathway in WAT
2.6. Effects of Ovariectomy and E2 Supplementation on Cardiac Insulin Pathway and Fibrosis
3. Discussion
4. Materials and Methods
4.1. Animal, Diets and Treatments
4.2. Glucose and Insulin Tolerance Tests
4.3. Serum Parameters
4.4. Tissue Sample Preparation and Determinations
4.5. Morphological Analysis of WAT
4.6. RNA Isolation and Real-Time PCR
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Sexual dimorphism in cardiometabolic health: The role of adipose tissue, muscle and liver. Nat. Rev. Endocrinol. 2021, 17, 47–66. [Google Scholar] [CrossRef] [PubMed]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; De La Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-Induced Changes in Adipose Tissue Microenvironment and Their Impact on Cardiovascular Disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Hearth J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [Green Version]
- Norhammar, A. Diabetes and cardiovascular mortality: The impact of sex. Lancet Diabetes Endocrinol. 2018, 6, 517–519. [Google Scholar] [CrossRef]
- Gerdts, E.; Regitz-Zagrosek, V. Sex differences in cardiometabolic disorders. Nat. Med. 2019, 25, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Resnick, E.M.; Simon, V.R.; Iskikian, S.O.; Marts, S.A. Future research in sex differences in obesity and cardiovascular disease: Report by the society for women’s health research. J. Investig. Med. 2007, 55, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Tramunt, B.; Smati, S.; Grandgeorge, N.; Lenfant, F.; Arnal, J.-F.; Montagner, A.; Gourdy, P. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia 2020, 63, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Leeners, B.; Geary, N.; Tobler, P.; Asarian, L. Ovarian hormones and obesity. Hum. Reprod. Update 2017, 23, 300–321. [Google Scholar] [CrossRef] [PubMed]
- Ribas, V.; Nguyen, M.T.A.; Henstridge, D.C.; Nguyen, A.-K.; Beaven, S.W.; Watt, M.J.; Hevener, A.L. Impaired oxidative metabolism and inflammation are associated with insulin resistance in ERα-deficient mice. Am. J. Physiol. Metab. 2010, 298, E304–E319. [Google Scholar] [CrossRef] [Green Version]
- D’Eon, T.M.; Souza, S.C.; Aronovitz, M.; Obin, M.S.; Fried, S.K.; Greenberg, A.S. Estrogen Regulation of Adiposity and Fuel Partitioning. J. Biol. Chem. 2005, 280, 35983–35991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevaskis, J.L.; Turek, V.F.; Wittmer, C.; Griffin, P.S.; Wilson, J.K.; Reynolds, J.M.; Zhao, Y.; Mack, C.M.; Parkes, D.G.; Roth, J.D. Enhanced Amylin-Mediated Body Weight Loss in Estradiol-Deficient Diet-Induced Obese Rats. Endocrinology 2010, 151, 5657–5668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keteepe-Arachi, T.; Sharma, S. Cardiovascular Disease in Women: Understanding Symptoms and Risk Factors. Eur. Cardiol. Rev. 2017, 12, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Nakhjavani, M.; Imani, M.; Larry, M.; Aghajani-Nargesi, A.; Morteza, A.; Esteghamati, A. Metabolic syndrome in premenopausal and postmenopausal women with type 2 diabetes: Loss of protective effects of premenopausal status. J. Diabetes Metab. Disord. 2014, 13, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schianca, G.P.C.; Fra, G.P.; Colli, E.; Bigliocca, M.; Mella, R.; Scaglia, E.; Bartoli, E. Sex differences in lipid profiles in relation to the progression of glucose abnormalities. J. Diabetes 2012, 4, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Masding, M.G.; Stears, A.J.; Burdge, G.C.; Wootton, S.A.; Sandeman, D.D. Premenopausal advantages in postprandial lipid metabolism are lost in women with type 2 diabetes. Diabetes Care 2003, 26, 3243–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergeron, R.; Mentor, J.S.; Côté, I.; Sock, E.N.; Rabasa-Lhoret, R.; Lavoie, J.M. Loss of Ovarian Estrogens Causes Only Mild Deterioration of Glucose Homeostasis in Female ZDF Rats Preventable by Voluntary Running Exercise. Horm. Metab. Res. 2014, 46, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Dobiásová, M. AIP—Atherogenic index of plasma as a significant predictor of cardiovascular risk: From research to practice. Vnitrni Lek. 2006, 52, 64–71. [Google Scholar]
- Nicholls, H.T.; Kowalski, G.; Kennedy, D.J.; Risis, S.; Zaffino, L.A.; Watson, N.; Kanellakis, P.; Watt, M.J.; Bobik, A.; Bonen, A.; et al. Hematopoietic Cell-Restricted Deletion of CD36 Reduces High-Fat Diet-Induced Macrophage Infiltration and Improves Insulin Signaling in Adipose Tissue. Diabetes 2011, 60, 1100–1110. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M.; Fucho, R.; Görgün, C.Z.; Tuncman, G.; Cao, H.; Hotamisligil, G.S. Adipocyte/macrophage fatty acid–binding proteins contribute to metabolic deterioration through actions in both macrophages and adipocytes in mice. J. Clin. Investig. 2008, 118, 2640–2650. [Google Scholar] [CrossRef] [Green Version]
- Itoh, M.; Suganami, T.; Hachiya, R.; Ogawa, Y. Adipose Tissue Remodeling as Homeostatic Inflammation. Int. J. Inflamm. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Podolsky, M.J.; Atabai, K. Fat fibrosis: Friend or foe? JCI Insight 2018, 3, 1–16. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular matrix remodeling of adipose tissue in obesity and metabolic diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chait, A.; Hartigh, L.J.D. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almon, R.R.; Wang, X.; Dubois, D.C.; Sukumaran, S.; Ayyar, V.; Jusko, W.J. Variability in Zucker diabetic fatty rats: Differences in disease progression in hyperglycemic and normoglycemic animals. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Faghfouri, A.H.; Khajebishak, Y.; Payahoo, L.; Faghfuri, E.; Alivand, M. PPAR-gamma agonists: Potential modulators of autophagy in obesity. Eur. J. Pharmacol. 2021, 912, 174562. [Google Scholar] [CrossRef] [PubMed]
- Al-Salameh, A.; Chanson, P.; Bucher, S.; Ringa, V.; Becquemont, L. Cardiovascular Disease in Type 2 Diabetes: A Review of Sex-Related Differences in Predisposition and Prevention. Mayo Clin. Proc. Elsevier 2019, 94, 287–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmborg, M.; Schmiegelow, M.D.S.; Noergaard, C.H.; Munch, A.; Gerds, T.; Schou, M.; Kistorp, C.; Torp-Pedersen, C.; Hlatky, M.A.; Gislason, G. Does type 2 diabetes confer higher relative rates of cardiovascular events in women compared with men? Eur. Heart J. 2020, 41, 1346–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korićanac, G.; Tepavcevic, S.; Romic, S.; Zivkovic, M.; Stojiljkovic, M.; Milosavljevic, T.; Stanković, A.; Petković, M.; Kamčeva, T.; Zakula, Z. Estradiol enhances effects of fructose rich diet on cardiac fatty acid transporter CD36 and triglycerides accumulation. Eur. J. Pharmacol. 2012, 694, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.; Brinton, E.A.; Grunfeld, C. The Effect of Endocrine Disorders on Lipids and Lipoproteins; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Gormsen, L.C.; Høst, C.; Hjerrild, B.E.; Gravholt, C.H.; Nielsen, S. Acute estrogen exposure does not affect basal very low-density lipoprotein–triglyceride production or oxidation in postmenopausal women. Eur. J. Endocrinol. 2010, 163, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Tawfik, S.H.; Mahmoud, B.F.; Saad, M.I.; Shehata, M.; Kamel, M.; Helmy, M.H. Similar and Additive Effects of Ovariectomy and Diabetes on Insulin Resistance and Lipid Metabolism. Biochem. Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmailidehaj, M.; Kuchakzade, F.; Rezvani, M.E.; Farhadi, Z.; Esmaeili, H.; Azizian, H. 17β-Estradiol improves insulin signalling and insulin resistance in the aged female hearts: Role of inflammatory and anti-inflammatory cytokines. Life Sci. 2020, 253, 117673. [Google Scholar] [CrossRef] [PubMed]
- Leffler, K.; Abdel-Rahman, A.A. Estrogen-Dependent Disruption of Adiponectin-Connexin43 Signaling Underlies Exacerbated Myocardial Dysfunction in Diabetic Female Rats. J. Pharmacol. Exp. Ther. 2019, 368, 208–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigt, C.; Hertrampf, T.; Flenker, U.; Hülsemann, F.; Kurnaz, P.; Fritzemeier, K.H.; Diel, P. Effects of estradiol, estrogen receptor subtype-selective agonists and genistein on glucose metabolism in leptin resistant female Zucker diabetic fatty (ZDF) rats. J. Steroid Biochem. Mol. Biol. 2015, 154, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Francisco, V.L.G.; Pino, J.; Gonzalez-Gay, M.A.; Mera, A.; Lago, F.; Gómez, R.; Mobasheri, A.; Gualillo, O. Adipokines and inflammation: Is it a question of weight? Br. J. Pharmacol. 2018, 175, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.; Church, C.; Baker, D.J.; Jones, S.W. The role of adipokines in skeletal muscle inflammation and insulin sensitivity. J. Inflamm. 2018, 15, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Apovian, C. Macrophage functions in lean and obese adipose tissue. Metabolism 2017, 72, 120–143. [Google Scholar] [CrossRef] [PubMed]
- Galic, S.; Oakhill, J.; Steinberg, G.R. Adipose tissue as an endocrine organ. Mol. Cell. Endocrinol. 2010, 316, 129–139. [Google Scholar] [CrossRef]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New Insight into Adiponectin Role in Obesity and Obesity-Related Diseases. BioMed Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- N, O.; K, W.; Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar]
- Diggins, N.; Webb, D.J. APPL1 is a multifunctional endosomal signaling adaptor protein. Biochem. Soc. Trans. 2017, 45, 771–779. [Google Scholar] [CrossRef]
- Deepa, M.; Farooq, S.; Deepa, R.; Manjula, D.; Mohan, V. Prevalence and significance of generalized and central body obesity in an urban Asian Indian population in Chennai, India (CURES: 47). Eur. J. Clin. Nutr. 2007, 63, 259–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, K.-C.; Xu, A.; Craig, M.E.; Martin, A.; Lam, K.S.; O’Sullivan, A.J. Adiponectin isoform distribution in women—relationship to female sex steroids and insulin sensitivity. Metabolism 2009, 58, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Merki-Feld, G.S.; Imthurn, B.; Rosselli, M.; Spanaus, K. Serum concentrations of high–molecular weight adiponectin and their association with sex steroids in premenopausal women. Metabolism 2011, 60, 180–185. [Google Scholar] [CrossRef]
- Eldridge, R.C.; Wentzensen, N.; Pfeiffer, R.M.; Brinton, L.A.; Hartge, P.; Guillemette, C.; Kemp, T.J.; Pinto, L.A.; Trabert, B. Endogenous estradiol and inflammation biomarkers: Potential interacting mechanisms of obesity-related disease. Cancer Causes Control 2020, 31, 309–320. [Google Scholar] [CrossRef]
- Love-Gregory, L.; Abumrad, N.A. CD36 genetics and the metabolic complications of obesity. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Bonen, A.; Tandon, N.N.; Glatz, J.F.C.; Luiken, J.J.F.P.; Heigenhauser, G.J.F. The fatty acid transporter FAT/CD36 is upregulated in subcutaneous and visceral adipose tissues in human obesity and type 2 diabetes. Int. J. Obes. 2006, 30, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Luiken, J.J.F.P.; Arumugam, Y.; Dyck, D.J.; Bell, R.C.; Pelsers, M.M.L.; Turcotte, L.P.; Tandon, N.N.; Glatz, J.F.C.; Bonen, A. Increased Rates of Fatty Acid Uptake and Plasmalemmal Fatty Acid Transporters in Obese Zucker Rats. J. Biol. Chem. 2001, 276, 40567–40573. [Google Scholar] [CrossRef] [Green Version]
- Lawler, H.M.; Underkofler, C.M.; Kern, P.A.; Erickson, C.; Bredbeck, B.; Rasouli, N. Adipose Tissue Hypoxia, Inflammation, and Fibrosis in Obese Insulin-Sensitive and Obese Insulin-Resistant Subjects. J. Clin. Endocrinol. Metab. 2016, 101, 1422–1428. [Google Scholar] [CrossRef] [Green Version]
- Schoettl, T.; Fischer, I.P.; Ussar, S. Heterogeneity of adipose tissue in development and metabolic function. J. Exp. Biol. 2018, 221, jeb162958. [Google Scholar] [CrossRef] [Green Version]
- Muir, L.; Neeley, C.K.; Meyer, K.A.; Baker, N.A.; Brosius, A.M.; Washabaugh, A.R.; Varban, O.A.; Finks, J.F.; Zamarron, B.F.; Flesher, C.G.; et al. Adipose tissue fibrosis, hypertrophy, and hyperplasia: Correlations with diabetes in human obesity. Obesity 2016, 24, 597–605. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clément, K.; Scherer, P.E. Fibrosis and Adipose Tissue Dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, C.; Liu, T.; Ling, H.; Cheng, M.; Zhou, X.; Wang, S.; Mao, Y.; Chen, L.; Zhang, R.; Li, W. Glucose variability aggravates cardiac fibrosis by altering AKT signalling path. Diabetes Vasc. Dis. Res. 2017, 14, 327–335. [Google Scholar] [CrossRef]
- Lou, P.-H.; Lucchinetti, E.; Scott, K.Y.; Huang, Y.; Gandhi, M.; Hersberger, M.; Clanachan, A.S.; Lemieux, H.; Zaugg, M. Alterations in fatty acid metabolism and sirtuin signaling characterize early type-2 diabetic hearts of fructose-fed rats. Physiol. Rep. 2017, 5, e13388. [Google Scholar] [CrossRef] [PubMed]
- Bayeva, M.; Sawicki, K.T.; Ardehali, H. Taking Diabetes to Heart—Deregulation of Myocardial Lipid Metabolism in Diabetic Cardiomyopathy. J. Am. Heart Assoc. 2013, 2, e000433. [Google Scholar] [CrossRef] [Green Version]
- Abumrad, N.A.; Goldberg, I.J. CD36 actions in the heart: Lipids, calcium, inflammation, repair and more? Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1442–1449. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.-R.; Clarke, K.; Levelt, E. Metabolic remodelling in diabetic cardiomyopathy. Cardiovasc. Res. 2017, 113, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Sobel, B.E.; Schneider, D.J.; Lee, Y.-H.; Pratley, R.E. Insulin resistance increases PAI-1 in the heart. Biochem. Biophys. Res. Commun. 2006, 346, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Nambi, S.S.; Mather, K.; Baron, A.D.; Follmann, D.A.; Sullivan, G.; Quon, M.J. Quantitative Insulin Sensitivity Check Index: A Simple, Accurate Method for Assessing Insulin Sensitivity In Humans. J. Clin. Endocrinol. Metab. 2000, 85, 2402–2410. [Google Scholar] [CrossRef]
- Cacho, J.; Sevillano, J.; De Castro, J.; Herrera, E.; Ramos-Alvarez, M.P. Validation of simple indexes to assess insulin sensitivity during pregnancy in Wistar and Sprague-Dawley rats. Am. J. Physiol. Metab. 2008, 295, E1269–E1276. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward Primer (5′->3′) | Reverse Primer (5′->3′) | Product Length (bp) |
|---|---|---|---|---|
| Ap2 | XM_039095687.1 | CCGATCCACTCCTTACCT | GCCACCGTGACCTTGTAC | 254 |
| Bad | NM_022698.1 | AGAGTTTGAGCCGAGTGAGCACT | CCGGGTCTCCATAGTCC | 186 |
| Bcl2 | NM_016993.1 | CTTCTTTGAGTTCGGTGGGGTGGA | GAAATCAAACAGAGGTCGC | 151 |
| Ccl3 | NM_013025.2 | TGCCCTTGCTGTTCTTCTCT | AAAGGCTGCTGGTCTCAAAA | 152 |
| Cd36 | NM_031561 | CTCACACAACTCAGATACTGCTG | TCCAAACACAGCCAGGACAG | 200 |
| Cd68 | NM_001031638.1 | CCCGAACAAAACCAAGGTCC | CTGCGCTGAGAATGTCCACT | 196 |
| Col1a1 | NM_053304.1 | GGAGAGAGCATGACCGATGG | GGGACTTCTTGAGGTTGCCA | 184 |
| Col3a1 | NM_032085.1 | TCCCCTGGAATCTGTGAATC | TGAGTCGAATTGGGGAGAAT | 63 |
| Col4a1 | NM_001135009.1 | CTCTGGGGACAACATCCG | TCTTCTCATGCACACTTGGC | 397 |
| Col6a1 | XM_008767345.2 | GGGACACACGTCTTCAGGTT | CCATGACTGATTGTTGTTGGG | 150 |
| Fn1 | NM_019143.2 | CAGCCCCTGATTGGAGTC | TGGGTGACACCTGAGTGAAC | 73 |
| Gapdh | NM_002046 | CTGGTGGTCCAGGGGTCTTA | CCACTCCTCCACCTTTGACG | 156 |
| Il1b | NM_031512.2 | CTGTGACTCGTGGGATGATG | GGGATTTTGTCGTTGCTTGT | 210 |
| Il6 | NM_012589.2 | TGTTCTCAGGGAGATCTTGG | TCCCAGGTAGAAACGGAACTC | 485 |
| Mmp12 | NM_053963.2 | GGCTGCTCCCATGAACGAG | GAGGTGTCCAGTTGCCCAG | 177 |
| Mmp14 | NM_031056.1 | AGGCCAATGTTCGGAGGAAG | GTGGCACTCTCCCATACTCG | 154 |
| Mmp2 | NM_031054.2 | ATGGTCGGGAATACAGCAGC | AGCTGTTGTAAGAGGTGCCC | 195 |
| Serpine1 | NM_012620.1 | TCTCTCCCTATGGCGTGTCC | GGCATCCGCAGTACTGATCT | 188 |
| Slc2a4 | XM_041665189.1 | TATTTGGCTTTGTGGCCTTC | CGGCAAATAGAAGGAAGACG | 201 |
| Tgfb1 | NM_021578.2 | GCAACAACGCAATCTATGAC | CCTGTATTCCGTCTCCTT | 300 |
| Tnf | NM_012675.3 | CTGAACTTCGGGGTGATCGG | CTTGGTGGTTTGCTACGACG | 151 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Cignoni, M.R.; González-Vicens, A.; Morán-Costoya, A.; Proenza, A.M.; Gianotti, M.; Valle, A.; Lladó, I. Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats. Int. J. Mol. Sci. 2021, 22, 13573. https://doi.org/10.3390/ijms222413573
Martínez-Cignoni MR, González-Vicens A, Morán-Costoya A, Proenza AM, Gianotti M, Valle A, Lladó I. Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats. International Journal of Molecular Sciences. 2021; 22(24):13573. https://doi.org/10.3390/ijms222413573
Chicago/Turabian StyleMartínez-Cignoni, Melanie Raquel, Agustí González-Vicens, Andrea Morán-Costoya, Ana María Proenza, Magdalena Gianotti, Adamo Valle, and Isabel Lladó. 2021. "Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats" International Journal of Molecular Sciences 22, no. 24: 13573. https://doi.org/10.3390/ijms222413573
APA StyleMartínez-Cignoni, M. R., González-Vicens, A., Morán-Costoya, A., Proenza, A. M., Gianotti, M., Valle, A., & Lladó, I. (2021). Estrogen Impairs Adipose Tissue Expansion and Cardiometabolic Profile in Obese-Diabetic Female Rats. International Journal of Molecular Sciences, 22(24), 13573. https://doi.org/10.3390/ijms222413573