
Probiotic Activity of Staphylococcus epidermidis Induces Collagen Type I Production through FFaR2/p-ERK Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
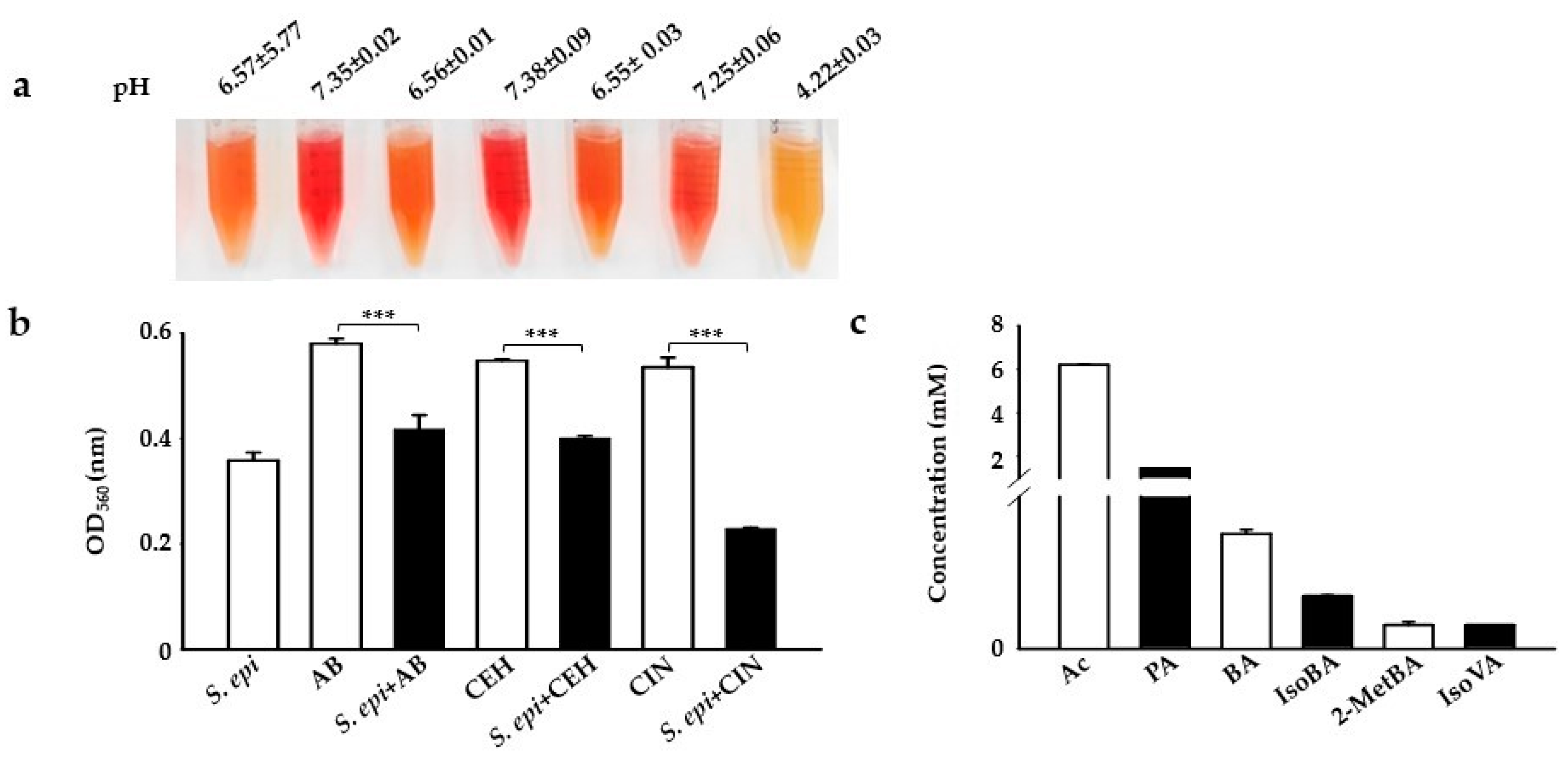
2.1. CIN as a Selective Fermentation Initiators (SFIs) for S. epidermidis
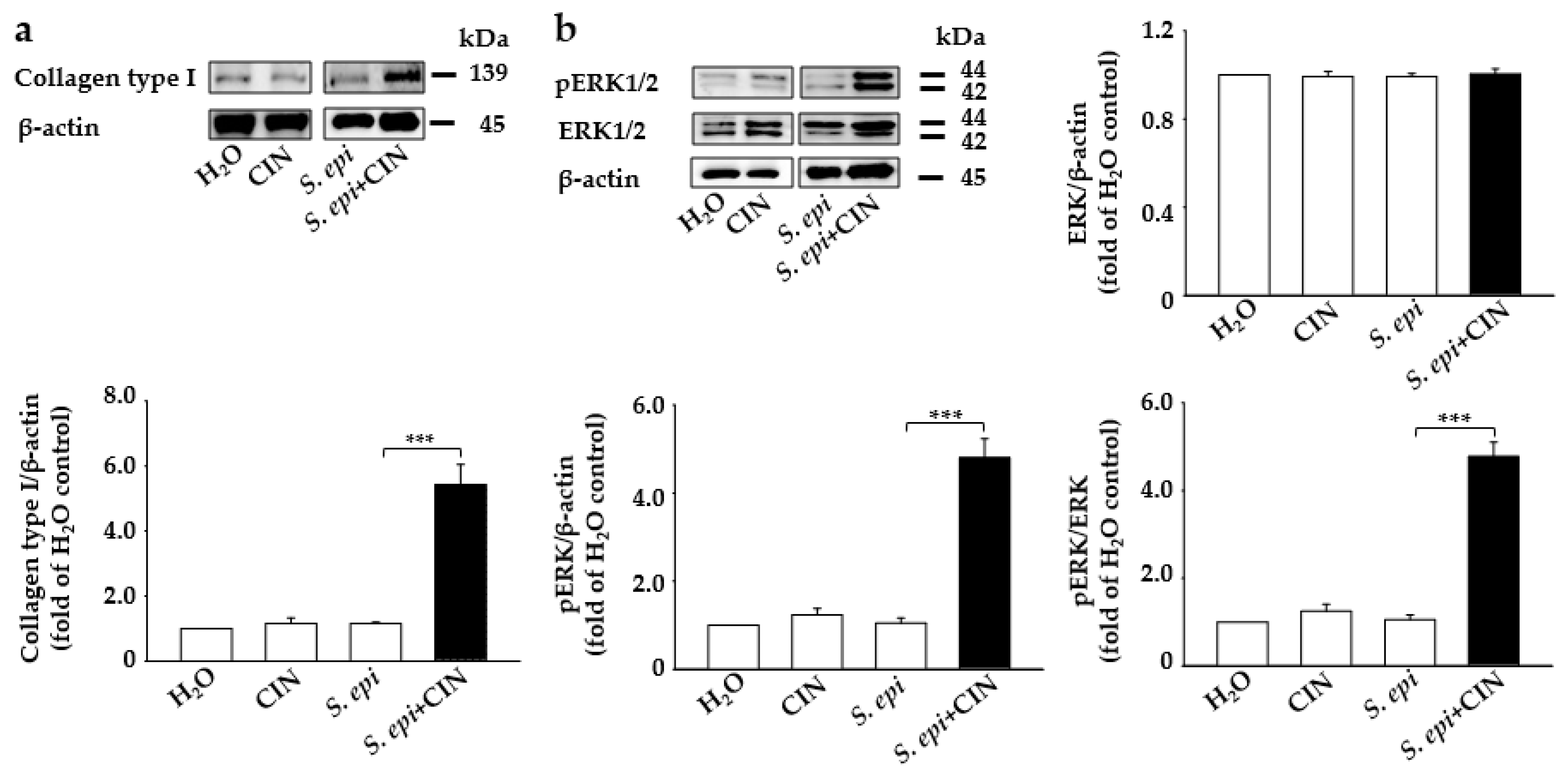
2.2. Mixture of CIN and S. epidermidis Induces the Expression of Collagen Type I and p-ERK Production in Mouse Skin
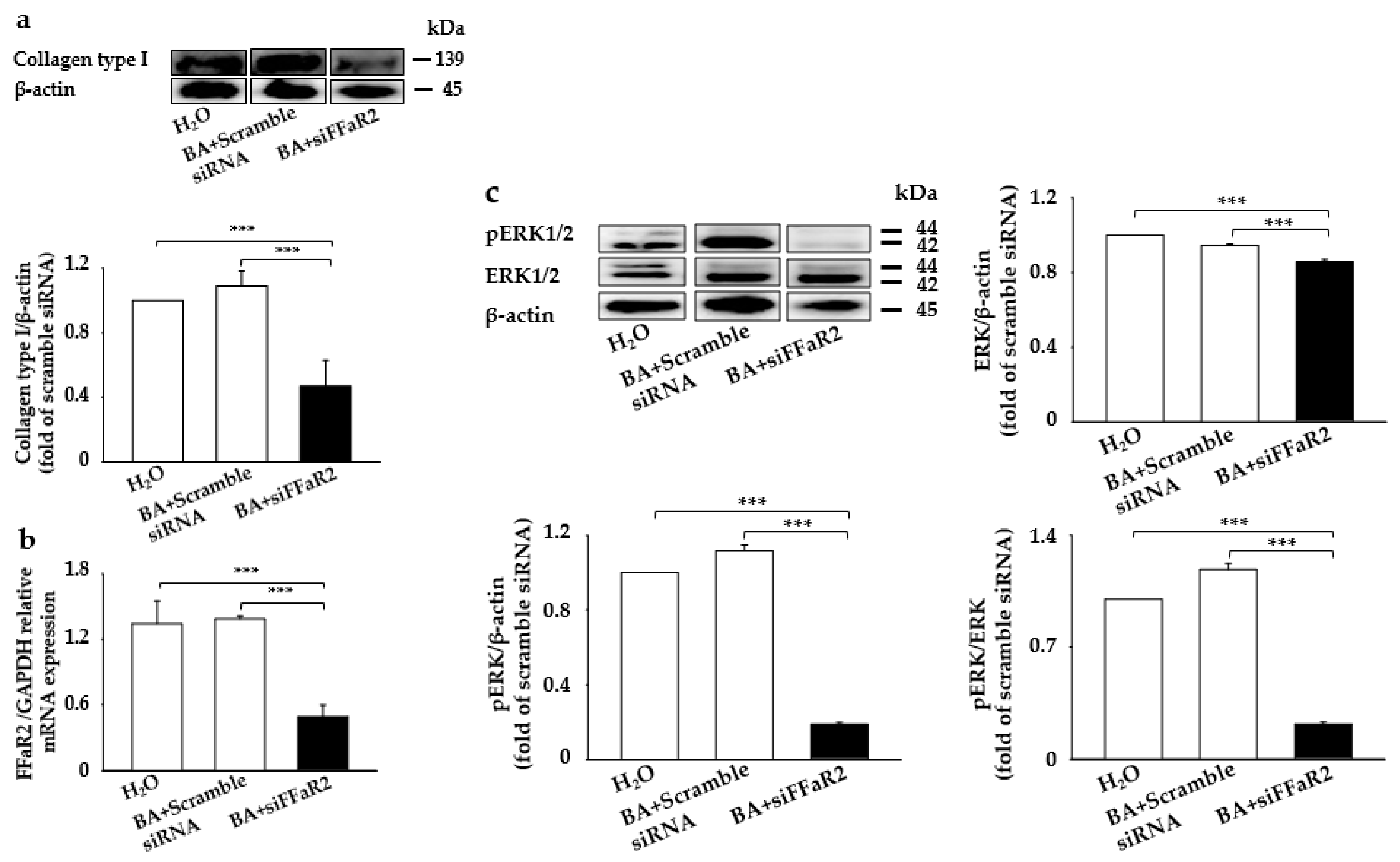
2.3. Knocking Down FFaR2-Inhibited BA-Mediated Induction of Collagen Type I and p-ERK in Mouse Fibroblasts
2.4. Knocking Down FFaR2-Inhibited Fermentation Metabolite-Mediated Induction of Collagen Type I and p-ERK in Mouse Skin
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacterial Culture
4.3. Fermentation of Bacteria
4.4. GC-MS Analysis
4.5. Cell Culture
4.6. Western-Blotting
4.7. siRNA-Mediated Gene Silencing of GPR43/FFaR2
4.8. Drug Treatment
4.9. RT-PCR
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AB | C12-14 alkyl benzoate |
| Ac | Acetic acid |
| ALS | Acetolactate synthease |
| ATCC | American type culture collection |
| BA | Butyric acid |
| BSA | Bovine serum albumin |
| cDNA | Complementary deoxyribonucleic acid |
| CEH | Cetyl ethylhexanoate |
| CFU | Colony forming unit |
| CIN | Cetearyl isononanoate |
| CIR | Cosmetic ingredient review |
| CO2 | Carbon dioxide |
| H2O | Dihydrogen monoxide |
| DMEM | Dulbecco’s modified eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| ECM | Extracellular matrix |
| EDTA | Ethylenediamine tetraacetic acid |
| ERK | Extracellular signal regulated kinase |
| FBS | Fetal bovine serum |
| FDA | Food and drug administration |
| FFaR2 | Free fatty acid receptor 2 |
| FFAs | Free fatty acids |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GC-MS | Gas chromatography-mass spectrometry |
| GPCRs | G-protein coupled receptors |
| GPR43 | G-protein coupled receptor 43 |
| H | Hour |
| IACUC | Institutional animal care and use committee |
| IgG | Immunoglobulin G |
| INCI | International nomenclature of cosmetic ingredients |
| IsoBA | Isobutyric acid |
| IsoVA | Isovaleric acid |
| kDa | Kilodalton |
| MAP | Mitogen-activated protein |
| MEK | Mitogen-extracellular signal regulated kinase |
| MMP | Matrix metalloproteinase |
| 2MetBA | 2-Methyl butyric acid |
| Min | Minute |
| mRNA | Messenger ribonucleic acid |
| μM | Micromolar |
| NaB | Sodium butyrate |
| NCBI | National center for biotechnology information |
| NCU | National Central University |
| NM | Nanometer |
| OD | Optical density |
| PA | Propionic acid |
| PBS | Phosphate buffer saline |
| PEG | Polyethylene glycol |
| p-ERK | Phosphorylated extracellular signal regulated kinase |
| pH | Power of hydrogen |
| PVDF | Polyvinylidene difluoride |
| ROS | Reactive oxygen species |
| RNA | Ribonucleic acid |
| RT-PCR | Real time polymerase chain reaction |
| S. epidermidis | Staphylococcus epidermidis |
| SCFAs | Short chain fatty acids |
| SD | Standard deviation |
| SDS | Sodium dodecyl sulfate |
| SFIs | Selective fermentation initiators |
| siFFaR2 | Small interfering free fatty acid receptor 2 |
| siRNA | Small interfering ribonucleic acid |
| TAME | Target aging with metformin |
| TBST | Tris-buffered saline tween 20 |
| T-PER | Tissue protein extraction reagent |
| TSB | Tryptic soy broth |
| UVB | Ultraviolet B |
References
- Quan, C.; Cho, M.K.; Perry, D.; Quan, T. Age-associated reduction of cell spreading induces mitochondrial DNA common deletion by oxidative stress in human skin dermal fibroblasts: Implication for human skin connective tissue aging. J. Biomed. Sci. 2015, 22, 62. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Fisher, G.J. Role of Age-Associated Alterations of the Dermal Extracellular Matrix Microenvironment in Human Skin Aging: A Mini-Review. Gerontology 2015, 61, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Cho, D.H.; Kim, H.J.; Lee, J.Y.; Cho, B.K.; Bang, S.I.; Song, S.Y.; Yamasaki, K.; Di Nardo, A.; Gallo, R.L. Collagen synthesis is suppressed in dermal fibroblasts by the human antimicrobial peptide LL-37. J. Investig. Dermatol. 2009, 129, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Egbert, M.; Ruetze, M.; Sattler, M.; Wenck, H.; Gallinat, S.; Lucius, R.; Weise, J.M. The matricellular protein periostin contributes to proper collagen function and is downregulated during skin aging. J. Dermatol. Sci. 2014, 73, 40–48. [Google Scholar] [CrossRef]
- Lee, D.J.; Rosenfeldt, H.; Grinnell, F. Activation of ERK and p38 MAP Kinases in Human Fibroblasts during Collagen Matrix Contraction. Exp. Cell Res. 2000, 257, 190–197. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2018, 4, eaao4502. [Google Scholar] [CrossRef]
- Keshari, S.; Balasubramaniam, A.; Myagmardoloonjin, B.; Herr, D.R.; Negari, I.P.; Huang, C.M. Butyric Acid from Probiotic Staphylococcus epidermidis in the Skin Microbiome Down-Regulates the Ultraviolet-Induced Pro-Inflammatory IL-6 Cytokine via Short-Chain Fatty Acid Receptor. Int. J. Mol. Sci. 2019, 20, 4477. [Google Scholar] [CrossRef]
- Tagliari, E.; Campos, L.F.; Campos, A.C.; Costa-Casagrande, T.A.; Noronha, L. Effect of Probiotic Oral Administration on Skin Wound Healing in Rats. Arq. Bras. Cir. Dig. 2019, 32, e1457. [Google Scholar] [CrossRef]
- Bowden, M.G.; Visai, L.; Longshaw, C.M.; Holland, K.T.; Speziale, P.; Hook, M. Is the GehD lipase from Staphylococcus epidermidis a collagen binding adhesin? J. Biol. Chem. 2002, 277, 43017–43023. [Google Scholar] [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Mohammad, S. Role of Free Fatty Acid Receptor 2 (FFAR2) in the Regulation of Metabolic Homeostasis. Curr. Drug. Targets 2015, 16, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Shimpukade, B.; Ulven, T.; Hudson, B.D. Complex Pharmacology of Free Fatty Acid Receptors. Chem. Rev. 2017, 117, 67–110. [Google Scholar] [CrossRef] [PubMed]
- Offermanns, S. Free fatty acid (FFA) and hydroxy carboxylic acid (HCA) receptors. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 407–434. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Layden, B.T.; Angueira, A.R.; Brodsky, M.; Durai, V.; Lowe, W.L., Jr. Short chain fatty acids and their receptors: New metabolic targets. Transl. Res. 2013, 161, 131–140. [Google Scholar] [CrossRef]
- Kao, M.S.; Huang, S.; Chang, W.L.; Hsieh, M.F.; Huang, C.J.; Gallo, R.L.; Huang, C.M. Microbiome precision editing: Using PEG as a selective fermentation initiator against methicillin-resistant Staphylococcus aureus. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef]
- Marito, S.; Keshari, S.; Huang, C.M. PEG-8 Laurate Fermentation of Staphylococcus epidermidis Reduces the Required Dose of Clindamycin Against Cutibacterium acnes. Int. J. Mol. Sci. 2020, 21, 5103. [Google Scholar] [CrossRef]
- Yang, A.J.; Marito, S.; Yang, J.J.; Keshari, S.; Chew, C.H.; Chen, C.C.; Huang, C.M. A Microtube Array Membrane (MTAM) Encapsulated Live Fermenting Staphylococcus epidermidis as a Skin Probiotic Patch against Cutibacterium acnes. Int. J. Mol. Sci. 2018, 20, 14. [Google Scholar] [CrossRef]
- Kumar, M.; Myagmardoloonjin, B.; Keshari, S.; Negari, I.P.; Huang, C.M. 5-methyl Furfural Reduces the Production of Malodors by Inhibiting Sodium l-lactate Fermentation of Staphylococcus epidermidis: Implication for Deodorants Targeting the Fermenting Skin Microbiome. Microorganisms 2019, 7, 239. [Google Scholar] [CrossRef]
- Kao, M.S.; Wang, Y.; Marito, S.; Huang, S.; Lin, W.Z.; Gangoiti, J.A.; Barshop, B.A.; Hyun, C.; Lee, W.R.; Sanford, J.A.; et al. The mPEG-PCL Copolymer for Selective Fermentation of Staphylococcus lugdunensis Against Candida parapsilosis in the Human Microbiome. J. Microb. Biochem. Technol. 2016, 8, 259–265. [Google Scholar] [CrossRef]
- Stipcevic, T.; Piljac, J.; Vanden Berghe, D. Effect of different flavonoids on collagen synthesis in human fibroblasts. Plant Foods Hum. Nutr. 2006, 61, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kim, H.J.; Woo, K.J.; Cho, D.; Bang, S.I. Lipo-PGE1 suppresses collagen production in human dermal fibroblasts via the ERK/Ets-1 signaling pathway. PLoS ONE 2017, 12, e0179614. [Google Scholar] [CrossRef]
- Karna, E.; Miltyk, W.; Pałka, J.A. Butyrate-induced collagen biosynthesis in cultured fibroblasts is independent on α2β1 integrin signalling and undergoes through IGF-I receptor cascade. Mol. Cell. Biochem. 2006, 286, 147. [Google Scholar] [CrossRef]
- Suto, M.; Masutomi, H.; Ishihara, K.; Masaki, H. A Potato Peel Extract Stimulates Type I Collagen Synthesis via Akt and ERK Signaling in Normal Human Dermal Fibroblasts. Biol. Pharm. Bull. 2019, 42, 1510–1516. [Google Scholar] [CrossRef]
- Bindels, L.B.; Dewulf, E.M.; Delzenne, N.M. GPR43/FFA2: Physiopathological relevance and therapeutic prospects. Trends Pharmacol. Sci. 2013, 34, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hasegawa, S.; Kasubuchi, M.; Kimura, I. Free fatty acid receptors as therapeutic targets for the treatment of diabetes. Front. Pharmacol. 2014, 5, 236. [Google Scholar] [CrossRef]
- Bjorkman, L.; Martensson, J.; Winther, M.; Gabl, M.; Holdfeldt, A.; Uhrbom, M.; Bylund, J.; Hojgaard Hansen, A.; Pandey, S.K.; Ulven, T.; et al. The Neutrophil Response Induced by an Agonist for Free Fatty Acid Receptor 2 (GPR43) Is Primed by Tumor Necrosis Factor Alpha and by Receptor Uncoupling from the Cytoskeleton but Attenuated by Tissue Recruitment. Mol. Cell. Biol. 2016, 36, 2583–2595. [Google Scholar] [CrossRef] [PubMed]
- Curaj, A.; Schumacher, D.; Rusu, M.; Staudt, M.; Li, X.; Simsekyilmaz, S.; Jankowski, V.; Jankowski, J.; Dumitrascu, A.R.; Hausenloy, D.J.; et al. Neutrophils Modulate Fibroblast Function and Promote Healing and Scar Formation after Murine Myocardial Infarction. Int. J. Mol. Sci. 2020, 21, 3685. [Google Scholar] [CrossRef]
- El-Domyati, M.; Attia, S.; Saleh, F.; Brown, D.; Birk, D.E.; Gasparro, F.; Ahmad, H.; Uitto, J. Intrinsic aging vs. photoaging: A comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp. Dermatol. 2002, 11, 398–405. [Google Scholar] [CrossRef]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced expression of connective tissue growth factor (CTGF/CCN2) mediates collagen loss in chronologically aged human skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Yasui, T.; Yonetsu, M.; Tanaka, R.; Tanaka, Y.; Fukushima, S.; Yamashita, T.; Ogura, Y.; Hirao, T.; Murota, H.; Araki, T. In vivo observation of age-related structural changes of dermal collagen in human facial skin using collagen-sensitive second harmonic generation microscope equipped with 1250-nm mode-locked Cr:Forsterite laser. J. Biomed. Opt. 2013, 18, 31108. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Merriam, G.; Kargi, A.Y. Growth Hormone in Aging; MDText.com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar]
- Moskalev, A. Is anti-ageing drug discovery becoming a reality? Expert Opin. Drug Dis. 2020, 15, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Dunaway, S.; Champer, J.; Kim, J.; Alikhan, A. Changing our microbiome: Probiotics in dermatology. Br. J. Dermatol. 2020, 182, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The role of topical probiotics in skin conditions: A systematic review of animal and human studies and implications for future therapies. Exp. Dermatol. 2020, 29, 15–21. [Google Scholar] [CrossRef]
- Lam, T.H.; Verzotto, D.; Brahma, P.; Ng, A.H.Q.; Hu, P.; Schnell, D.; Tiesman, J.; Kong, R.; Ton, T.M.U.; Li, J.; et al. Understanding the microbial basis of body odor in pre-pubescent children and teenagers. Microbiome 2018, 6, 213. [Google Scholar] [CrossRef]
- Bawdon, D.; Cox, D.S.; Ashford, D.; James, A.G.; Thomas, G.H. Identification of axillary Staphylococcus sp. involved in the production of the malodorous thioalcohol 3-methyl-3-sufanylhexan-1-ol. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef]
- Callewaert, C.; Kerckhof, F.M.; Granitsiotis, M.S.; Van Gele, M.; Van de Wiele, T.; Boon, N. Characterization of Staphylococcus and Corynebacterium clusters in the human axillary region. PLoS ONE 2013, 8, e70538. [Google Scholar] [CrossRef]
- Kaur, A.; Ecker, B.L.; Douglass, S.M.; Kugel, C.H., 3rd; Webster, M.R.; Almeida, F.V.; Somasundaram, R.; Hayden, J.; Ban, E.; Ahmadzadeh, H.; et al. Remodeling of the Collagen Matrix in Aging Skin Promotes Melanoma Metastasis and Affects Immune Cell Motility. Cancer Discov. 2019, 9, 64–81. [Google Scholar] [CrossRef]
- Jung, Y.R.; Kim, D.H.; Kim, S.R.; An, H.J.; Lee, E.K.; Tanaka, T.; Kim, N.D.; Yokozawa, T.; Park, J.N.; Chung, H.Y. Anti-wrinkle effect of magnesium lithospermate B from Salvia miltiorrhiza BUNGE: Inhibition of MMPs via NF-kB signaling. PLoS ONE 2014, 9, e102689. [Google Scholar] [CrossRef]
- Dupont, E.; Gomez, J.; Bilodeau, D. Beyond UV radiation: A skin under challenge. Int. J. Cosmet. Sci. 2013, 35, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Worthen, C.A.; Quan, T. Cell-size-dependent upregulation of HGF expression in dermal fibroblasts: Impact on human skin connective tissue aging. J. Dermatol. Sci. 2017, 88, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Dos Anjos Oliveira Ferreira, L.; de Paula Barros de Melo, C.; Saito, P.; Iwanaga, C.C.; Nakamura, C.V.; Casagrande, R.; da Conceicao Torrado Truiti, M. Nectandra cuspidata fraction and the isolated polyphenols protect fibroblasts and hairless mice skin from UVB-induced inflammation and oxidative stress. J. Photochem. Photobiol. B 2020, 205, 111824. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Hoshino, T.; Yamakawa, N.; Tahara, K.; Adachi, H.; Sobue, G.; Maji, D.; Ihn, H.; Mizushima, T. Suppression of UV-induced wrinkle formation by induction of HSP70 expression in mice. J. Investig. Dermatol. 2013, 133, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Vardar, G.; Altikatoglu, M.; Basaran, Y.; Isildak, I. Synthesis of glucose oxidase-PEG aldehyde conjugates and improvement of enzymatic stability. Artif. Cells Nanomed. Biotechnol. 2018, 46, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Fiume, M.M.; Heldreth, B.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G., Jr.; Shank, R.C.; Slaga, T.J.; et al. Safety Assessment of Alkyl PEG/PPG Ethers as Used in Cosmetics. Int. J. Toxicol. 2016, 35, 60S–89S. [Google Scholar] [CrossRef] [PubMed]
- Che, J.; Okeke, C.I.; Hu, Z.B.; Xu, J. DSPE-PEG: A distinctive component in drug delivery system. Curr. Pharm. Des. 2015, 21, 1598–1605. [Google Scholar] [CrossRef]
- Garland, S.H. Short chain fatty acids may elicit an innate immune response from preadipocytes: A potential link between bacterial infection and inflammatory diseases. Med. Hypotheses 2011, 76, 881–883. [Google Scholar] [CrossRef]
- Shah, V.P.; Flynn, G.L.; Yacobi, A.; Maibach, H.I.; Bon, C.; Fleischer, N.M.; Franz, T.J.; Kaplan, S.A.; Kawamoto, J.; Lesko, L.J.; et al. Bioequivalence of topical dermatological dosage forms–methods of evaluation of bioequivalence. Pharm. Res. 1998, 15, 167–171. [Google Scholar] [CrossRef]
- Raney, S.G.; Franz, T.J.; Lehman, P.A.; Lionberger, R.; Chen, M.L. Pharmacokinetics-Based Approaches for Bioequivalence Evaluation of Topical Dermatological Drug Products. Clin. Pharmacokinet. 2015, 54, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Baumann, A.; Tuerck, D.; Prabhu, S.; Dickmann, L.; Sims, J. Pharmacokinetics, metabolism and distribution of PEGs and PEGylated proteins: Quo vadis? Drug Discov. Today 2014, 19, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meng, Q.; Yang, M.; Liu, D.; Hou, X.; Tang, L.; Wang, X.; Lyu, Y.; Chen, X.; Liu, K.; et al. Current trends in drug metabolism and pharmacokinetics. Acta Pharm. Sin. B 2019, 9, 1113–1144. [Google Scholar] [CrossRef] [PubMed]
- Haines, J.R.; Alexander, M. Microbial degradation of polyethylene glycols. Appl. Microbiol. 1975, 29, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.; Josse, R.G.; Leiter, L.A.; Chiasson, J.L. Time of day and glucose tolerance status affect serum short-chain fatty acid concentrations in humans. Metabolism 1997, 46, 805–811. [Google Scholar] [CrossRef]
- Daniel, P.; Brazier, M.; Cerutti, I.; Pieri, F.; Tardivel, I.; Desmet, G.; Baillet, J.; Chany, C. Pharmacokinetic study of butyric acid administered in vivo as sodium and arginine butyrate salts. Clin. Chim. Acta 1989, 181, 255–263. [Google Scholar] [CrossRef]
- Vassilev, I.; Hernandez, P.A.; Batlle-Vilanova, P.; Freguia, S.; Krömer, J.O.; Keller, J.; Ledezma, P.; Virdis, B. Microbial Electrosynthesis of Isobutyric, Butyric, Caproic Acids, and Corresponding Alcohols from Carbon Dioxide. ACS Sust. Chem. Eng. 2018, 6, 8485–8493. [Google Scholar] [CrossRef]
- Kober, M.-M.; Bowe, W.P. The effect of probiotics on immune regulation, acne, and photoaging. Int. J. Women’s Dermatol. 2015, 1, 85–89. [Google Scholar] [CrossRef]
- Karna, E.; Trojan, S.; Palka, J.A. The mechanism of butyrate-induced collagen biosynthesis in cultured fibroblasts. Acta Pol. Pharm. 2009, 66, 129–134. [Google Scholar]
- Wang, Y.; Zhang, L.; Yu, J.; Huang, S.; Wang, Z.; Chun, K.A.; Lee, T.L.; Chen, Y.T.; Gallo, R.L.; Huang, C.M. A Co-Drug of Butyric Acid Derived from Fermentation Metabolites of the Human Skin Microbiome Stimulates Adipogenic Differentiation of Adipose-Derived Stem Cells: Implications in Tissue Augmentation. J. Investig. Dermatol. 2017, 137, 46–56. [Google Scholar] [CrossRef]
- Fluhr, J.W.; Darlenski, R.; Surber, C. Glycerol and the skin: Holistic approach to its origin and functions. Br. J. Dermatol. 2008, 159, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Lim, I.J.; Phan, T.T.; Tan, E.K.; Nguyen, T.T.; Tran, E.; Longaker, M.T.; Song, C.; Lee, S.T.; Huynh, H.T. Synchronous activation of ERK and phosphatidylinositol 3-kinase pathways is required for collagen and extracellular matrix production in keloids. J. Biol. Chem. 2003, 278, 40851–40858. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, H.; Tsukui, M.; Takada, S.; Kurashina, K.; Choi, Y.L.; Soda, M.; Yamashita, Y.; Haruta, H.; Hamada, T.; Ueno, T.; et al. Identification of transforming activity of free fatty acid receptor 2 by retroviral expression screening. Cancer Sci. 2010, 101, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef]
- Pirozzi, C.; Francisco, V.; Guida, F.D.; Gomez, R.; Lago, F.; Pino, J.; Meli, R.; Gualillo, O. Butyrate Modulates Inflammation in Chondrocytes via GPR43 Receptor. Cell Physiol. Biochem. 2018, 51, 228–243. [Google Scholar] [CrossRef]
- Boivin, G.P.; Hickman, D.L.; Creamer-Hente, M.A.; Pritchett-Corning, K.R.; Bratcher, N.A. Review of CO(2) as a Euthanasia Agent for Laboratory Rats and Mice. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 491–499. [Google Scholar]
- Keshari, S.; Wang, Y.; Herr, D.R.; Wang, S.M.; Yang, W.C.; Chuang, T.H.; Chen, C.L.; Huang, C.M. Skin Cutibacterium acnes Mediates Fermentation to Suppress the Calcium Phosphate-Induced Itching: A Butyric Acid Derivative with Potential for Uremic Pruritus. J. Clin. Med. 2020, 9, 312. [Google Scholar] [CrossRef]
- Pizzonero, M.; Dupont, S.; Babel, M.; Beaumont, S.; Bienvenu, N.; Blanque, R.; Cherel, L.; Christophe, T.; Crescenzi, B.; De Lemos, E.; et al. Discovery and optimization of an azetidine chemical series as a free fatty acid receptor 2 (FFA2) antagonist: From hit to clinic. J. Med. Chem. 2014, 57, 10044–10057. [Google Scholar] [CrossRef]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negari, I.P.; Keshari, S.; Huang, C.-M. Probiotic Activity of Staphylococcus epidermidis Induces Collagen Type I Production through FFaR2/p-ERK Signaling. Int. J. Mol. Sci. 2021, 22, 1414. https://doi.org/10.3390/ijms22031414
Negari IP, Keshari S, Huang C-M. Probiotic Activity of Staphylococcus epidermidis Induces Collagen Type I Production through FFaR2/p-ERK Signaling. International Journal of Molecular Sciences. 2021; 22(3):1414. https://doi.org/10.3390/ijms22031414
Chicago/Turabian StyleNegari, Indira Putri, Sunita Keshari, and Chun-Ming Huang. 2021. "Probiotic Activity of Staphylococcus epidermidis Induces Collagen Type I Production through FFaR2/p-ERK Signaling" International Journal of Molecular Sciences 22, no. 3: 1414. https://doi.org/10.3390/ijms22031414
APA StyleNegari, I. P., Keshari, S., & Huang, C.-M. (2021). Probiotic Activity of Staphylococcus epidermidis Induces Collagen Type I Production through FFaR2/p-ERK Signaling. International Journal of Molecular Sciences, 22(3), 1414. https://doi.org/10.3390/ijms22031414