
Gene Expression Analysis of Induced Plum pox virus (Sharka) Resistance in Peach (Prunus persica) by Almond (P. dulcis) Grafting

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
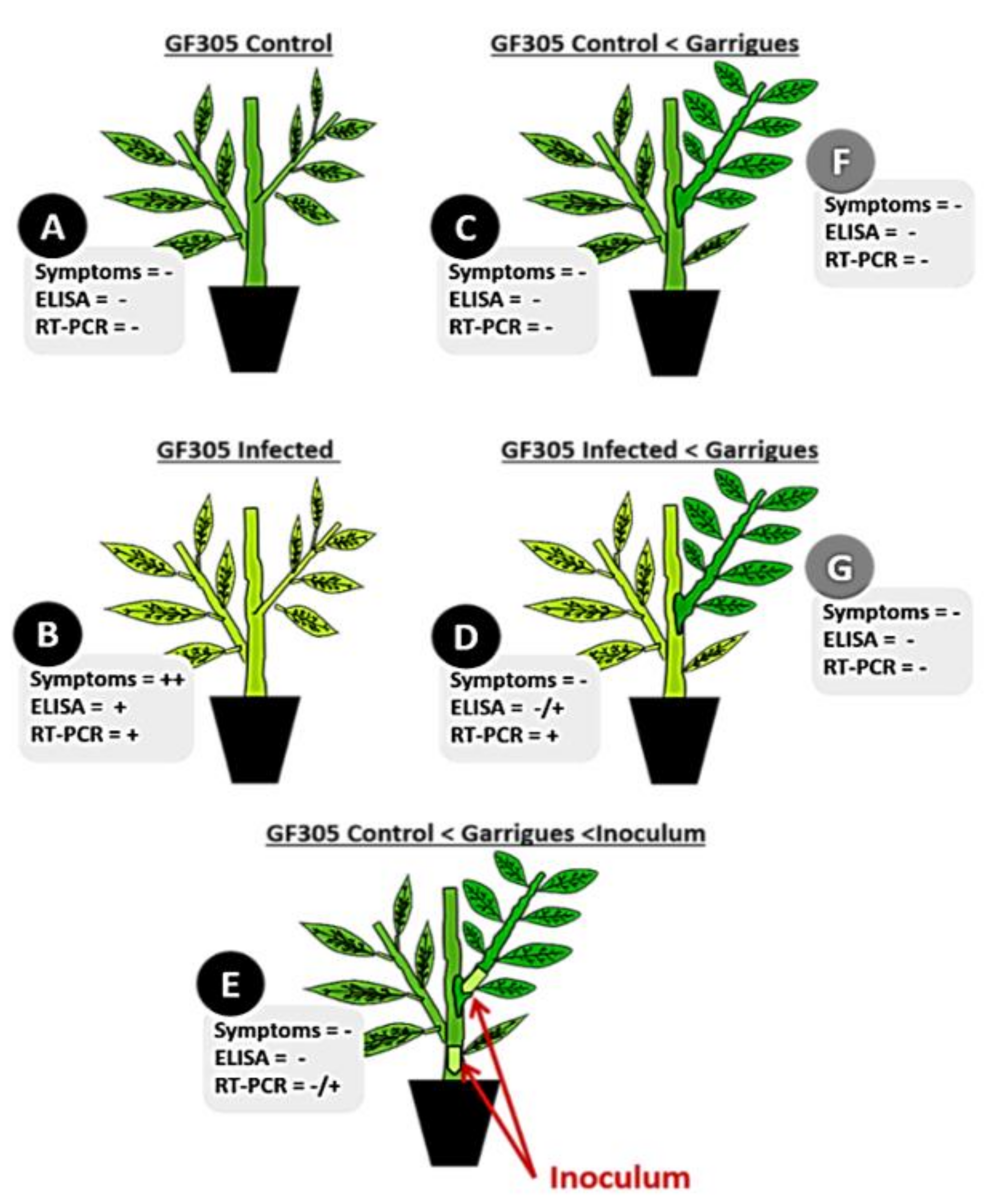
2.1. PPV Resistance Evaluation
2.2. RNA-Seq Transcriptome Profiles
2.3. Host Transcriptional Changes
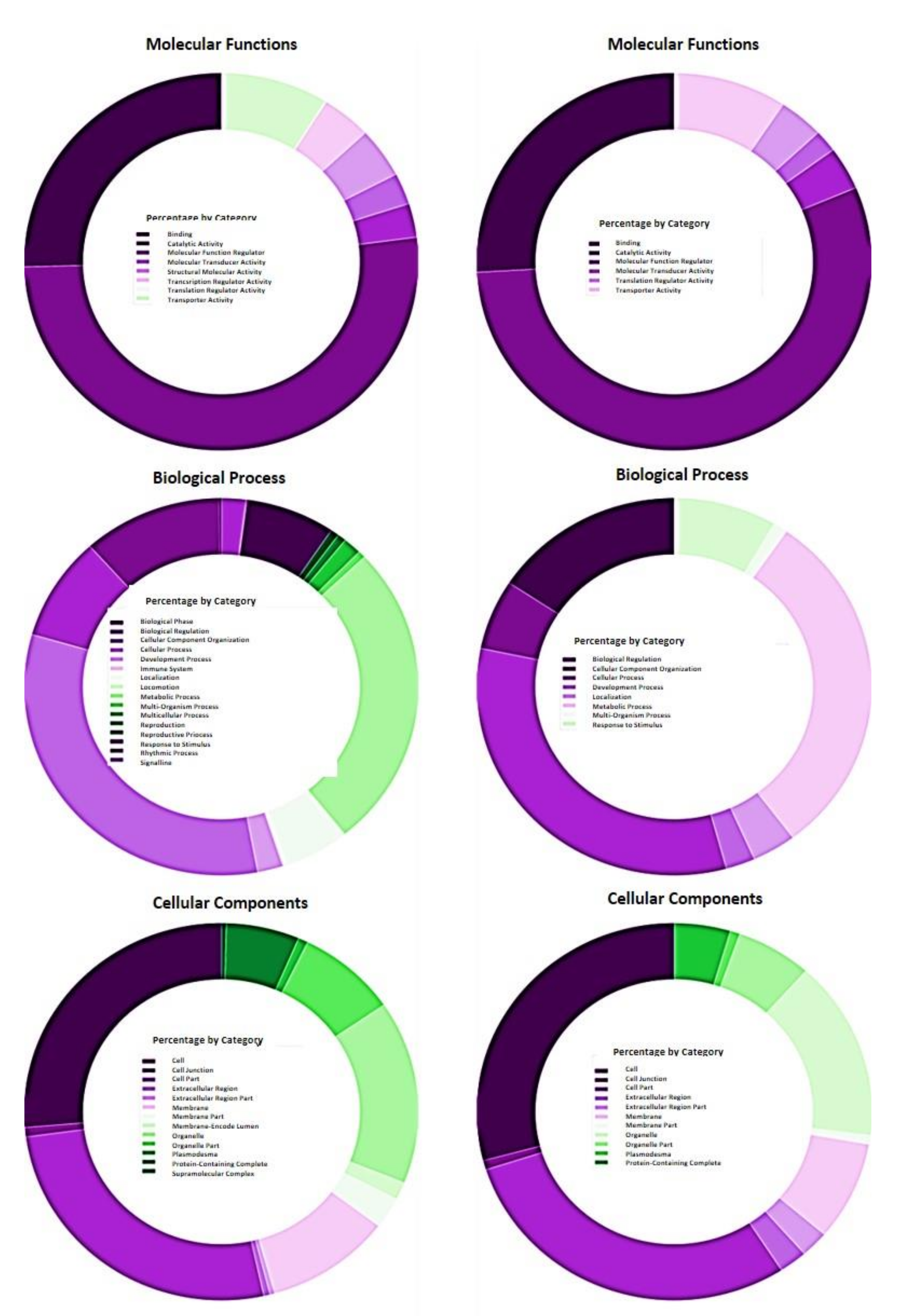
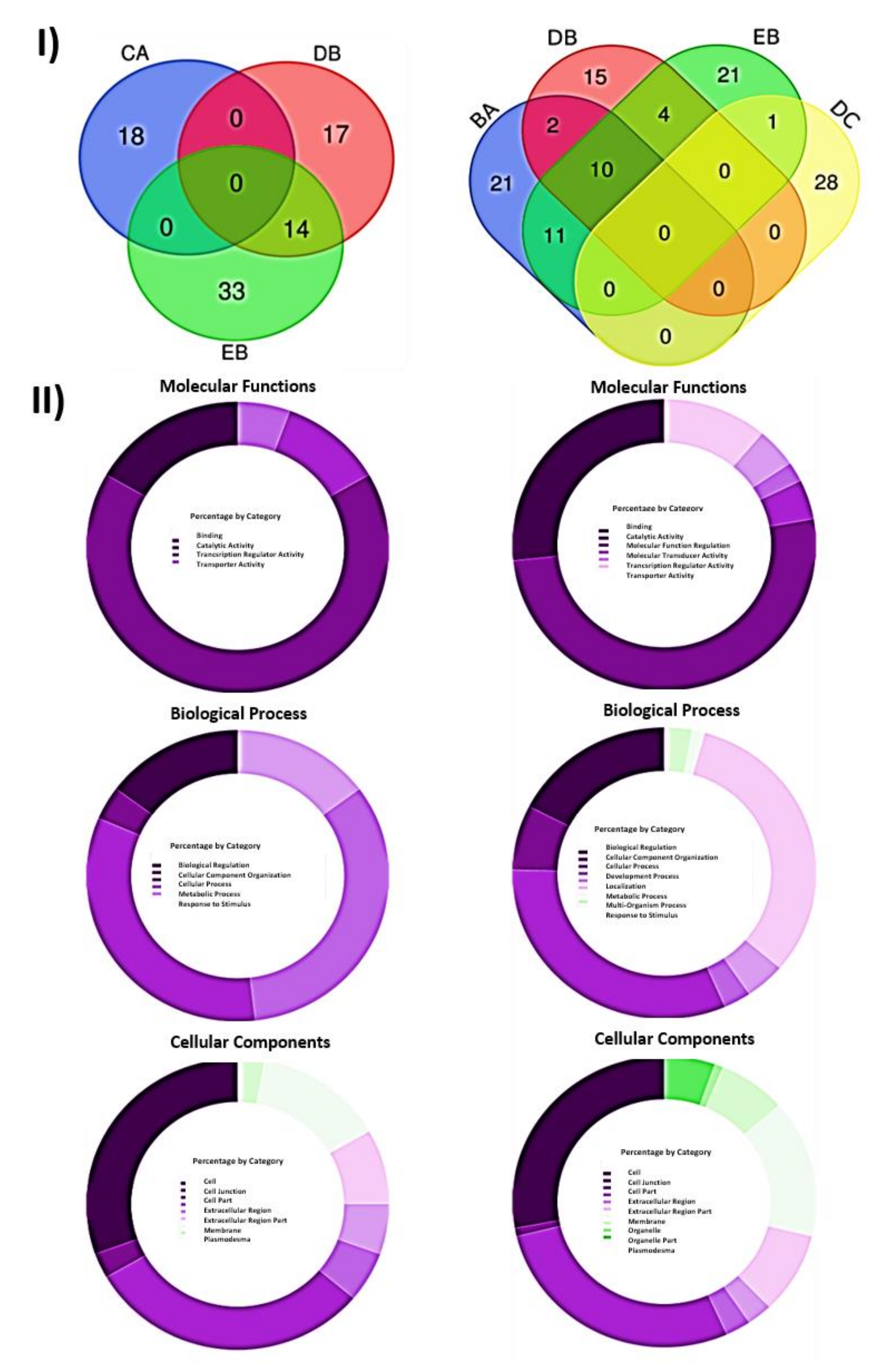
2.4. Functional Analysis of Differentially Expressed Genes
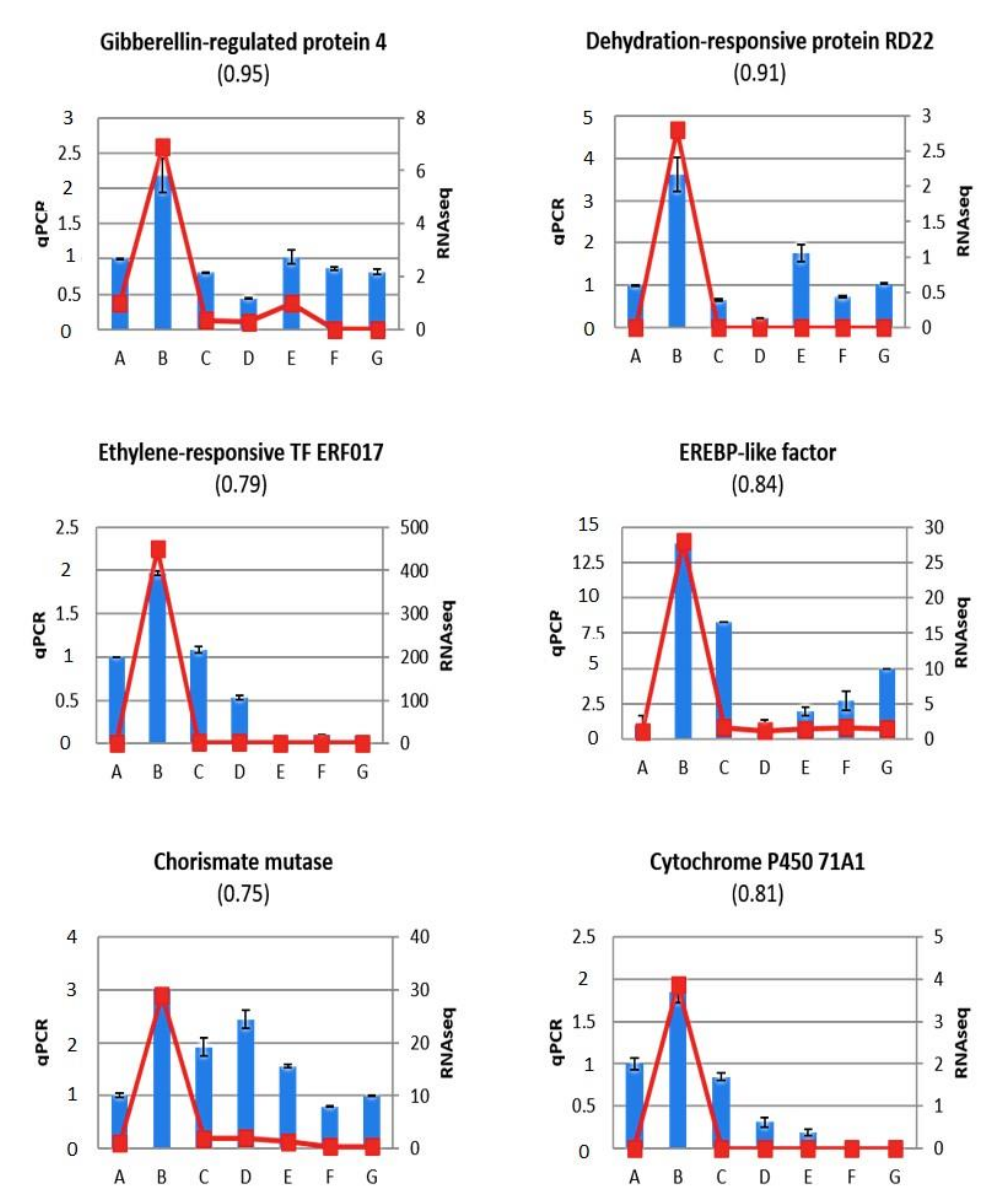
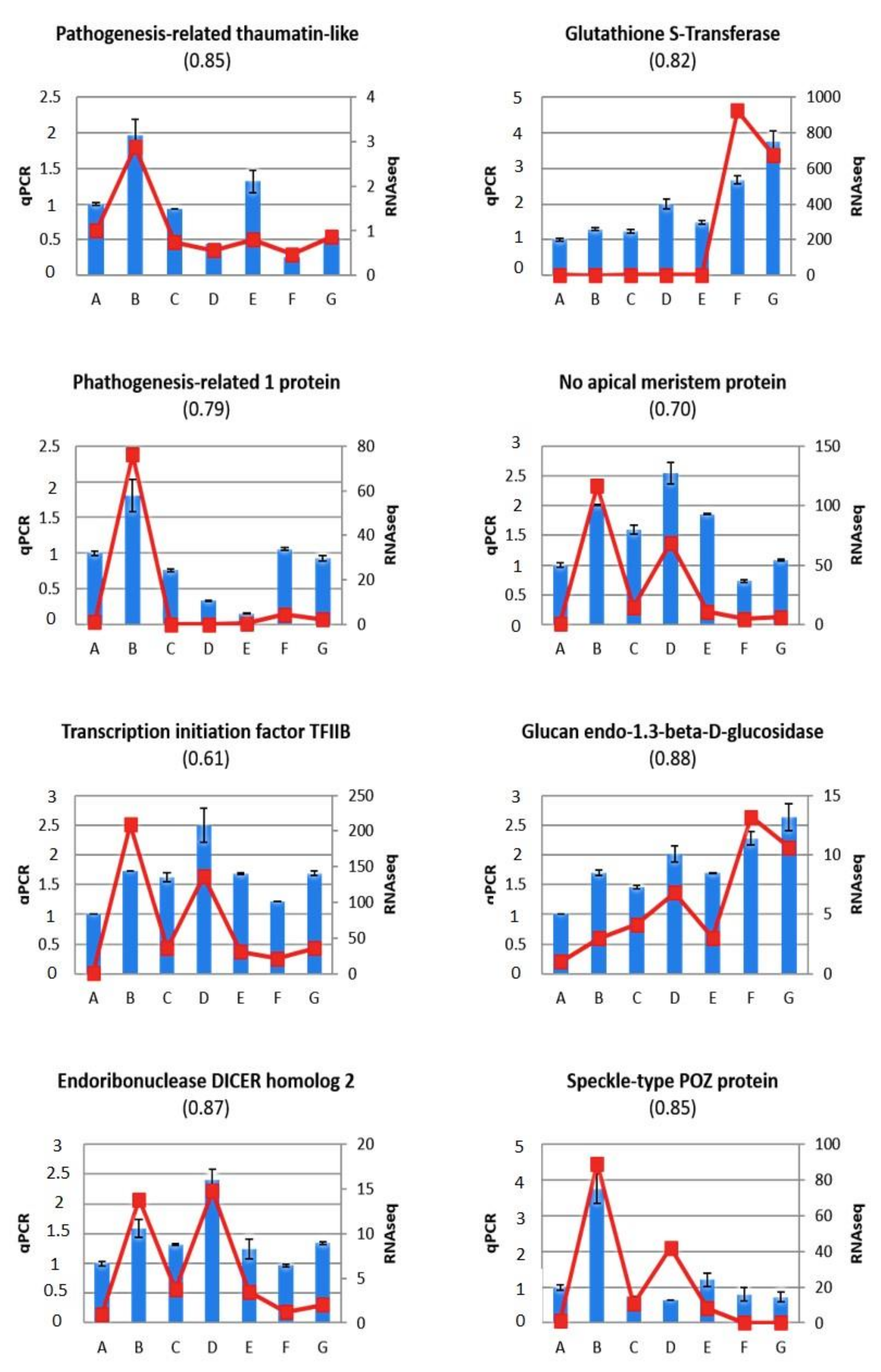
2.5. Validation of Gene Expression Profiles Using RT-qPCR
3. Discussion
4. Materials and Methods
4.1. Biological Materials
4.2. Sharka Phenotyping and PPV Detection
4.3. Experimental Design and High-Throughput mRNA Sequencing
4.4. Bioinformatic Analysis
4.5. RT-qPCR Validation of Selected Transcripts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CYP71A1 | Cytochrome P450 71A1 |
| DCL | Dicer-like |
| DEG | Differently expressed gene |
| ELISA-DASI | Enzyme-linked immunosorbent assay-double antibody sandwich mmunosorbent |
| GA | Gibberellic Acid |
| FPKM | Fragments per kilobase pair of transcript per million mapped reads |
| GO | Gene ontology |
| HCPro | Helper component proteinase |
| HEN1 | HUA ENHANCER 1 |
| JA | Jasmonic acid |
| ISR | Induced systemic resistance |
| MATH | Meprin and TRAF homology |
| OD | Optical density |
| PPV | Plum pox virus |
| PR | Pathogenesis-related |
| RDR | RNA-dependent RNA polymerase |
| RNA | Ribonucleic acid |
| RGE | Relative gene expression |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SA | Salicylic acid |
| SAR | Systemic acquired resistance |
| TF | Transcription factor |
| tZ | Trans-zeatine |
References
- Rubio, M.; Martínez-Gómez, P.; Marais, A.; Sánchez-Navarro, J.A.; Pallás, V.; Candresse, T. Recent advances and prospects in Prunus virology. Ann. Appl. Biol. 2017, 171, 125–138. [Google Scholar] [CrossRef]
- Sihelská, N.; Glasa, M.; Šubr, Z.W. Host preference of the major strains of Plum pox virus—Opinions based on regional and world-wide sequence data. J. Integr. Agric. 2017, 16, 510–515. [Google Scholar] [CrossRef]
- Scholthof, K.B.G.; Adkins, S.; Czosnek, H.; Saunders, K.; Candresse, T.; Ahlquist, P.; Hemenway, C.; Foster, G.D. Top 10 plant viruses in molecular plant pathology. Mol. Plant Patol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- García, J.A.; Glasa, M.; Cambra, C.; Candresse, T. Plum pox virus and sharka: A model potyvirus and a major disease. Mol. Plant Pathol. 2014, 15, 226–241. [Google Scholar] [CrossRef]
- Sochor, J.; Babula, P.; Adam, V.; Krska, B.; Kizek, R. Sharka: The past, the present and the future. Viruses 2012, 4, 2853–2901. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Gómez, P.; Dicenta, F.; Audergon, J.M. Behaviour of apricot (Prunus armeniaca L.) cultivars in presence of sharka (Plum pox virus): A review. Agronomie 2000, 20, 407–422. [Google Scholar] [CrossRef] [Green Version]
- Rubio, M.; Martínez-Gómez, P.; Dicenta, F. Resistance of almond cultivars to Plum pox virus (sharka). Plant Breed. 2003, 122, 462–464. [Google Scholar] [CrossRef]
- Rubio, M.; Martínez-Gómez, P.; Pascal, T.; Dicenta, F. Sensitivity of peach cultivars against a Dideron isolate of Plum pox virus. Sci. Hort. 2012, 144, 81–86. [Google Scholar] [CrossRef]
- Cirilli, M.; Geuna, F.; Babini, A.R.; Bozhkova, V.; Catalano, L.; Cavagna, B.; Dallot, S.; Decroocq, V.; Bondidni, L.; Foschi, S.; et al. Fighting sharka in peach: Current limitations and future perspectives. Front. Plant Sci. 2016, 7, 1290. [Google Scholar] [CrossRef]
- Rubio, M.; Pascal, T.; Bachellez, A.; Lambert, P. Quantitative trait loci analysis of PPV resistance in Prunus davidiana P1908. Tree Genet. Gen. 2010, 6, 291–304. [Google Scholar] [CrossRef]
- Martínez-Gómez, P.; Rubio, M.; Dicenta, F.; Gradziel, T.M. Resistance to Plum pox virus (RB3.30 isolate) in a group of California almonds and transfer of resistance to peach. J. Am. Soc. Hort. Sci. 2004, 129, 544–548. [Google Scholar] [CrossRef]
- Rubio, M.; Martínez-Gómez, P.; García, J.A.; Dicenta, F. Interspecific transfer of resistance to Plum pox virus from almond to peach by grafting. Ann. Appl. Biol. 2013, 163, 466–474. [Google Scholar]
- Rubio, M.; Rodríguez-Moreno, L.; Ballester, A.R.; Castro de Moura, M.; Bonghi, C.; Candresse, T.; Martínez-Gómez, P. Analysis of gene expression changes in peach leaf tissues in response to Plum pox virus infection using RNA-Seq. Mol. Plant Pathol. 2015, 16, 164–176. [Google Scholar] [CrossRef]
- Nikbakht-Dehkordi, A.; Rubio, M.; Babaeian, N.; Albacete, A.; Martínez-Gómez, P. Phytohormone Signaling of the Resistance to Plum pox virus (PPV, Sharka Disease) Induced by Almond (Prunus dulcis (Miller) Webb) Grafting to Peach (P. persica L. Batsch). Viruses 2018, 10, 238. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558–1560. [Google Scholar] [CrossRef]
- Guo, H.S.; Garcia, J.A. Delayed resistance to plum pox potyvirus mediated by a mutated RNA helicase gene: Involvement of a gene silencing mechanism. Mol. Plant-Microbe Interact. 1997, 10, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Pallás, V.; García, J.A. How do plant viruses induce disease? Interactions and interference with host componenents. J. General Virol. 2011, 92, 2691–2705. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, S.; Bock, R. Exchange of genetic material between cells in plant tissue grafts. Science 2009, 314, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Harada, J. Grafting and RNA transport via phloem tissue in horticultural plants. Sci. Hort. 2010, 125, 545–550. [Google Scholar] [CrossRef]
- Haroldsen, V.M.; Szczerba, M.W.; Aktas, H.; Lopez-Baltazar, J.; Odias, M.J. Mobility of Transgenic Nucleic Acids and Proteins within Grafted Rootstocks for Agricultural Improvement. Front. Plant Sci. 2012, 3, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.W.; Takahashi, N.; Hirata, Y.; Ronald, J.; Porco, S.; Davis, S.J.; Nusinow, D.A.; Kay, S.A.; Mas, P. A mobile ELF4 delivers circadian temperature information from shoots to roots. Nat. Plants 2020, 6, 416–426. [Google Scholar] [CrossRef]
- Molnar, A.; Melnyk, C.W.; Bassett, A.; Hardcastle, T.J.; Dunn, R. Small silencing RNAs in plants are mobile and direct epigenetic modification in recipient cells. Science 2000, 328, 872–875. [Google Scholar] [CrossRef] [Green Version]
- Lewsey, M.G.; Hardcastle, T.J.; Melnyk, C.W.; Molnar, A.; Valli, A.; Urich, M.A.; Nery, J.R.; Baulcombe, D.C.; Ecker, J.R. Mobile small RNAs regulate genome-wide DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, E801–E810. [Google Scholar] [CrossRef] [Green Version]
- Pagliarani, C.; Gambino, G. Small RNA Mobility: Spread of RNA Silencing Effectors and its Effect on Developmental Processes and Stress Adaptation in Plants. Int. J. Mol. Sci. 2019, 20, 4306. [Google Scholar] [CrossRef] [Green Version]
- Dandekar, A.M.; Jacobson, A.; Ibáñez, A.M.; Gouran, H.; Dolan, D.L.; Agüero, C.B.; Uratsu, S.L.; Just, R.; Zaini, P.A. Trans-Graft Protection Against Pierce’s Disease Mediated by Transgenic Grapevine Rootstocks. Front. Plant Sci. 2019, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Spanò, R.; Ferrara, M.; Montemurro, C.; Mulè, G.; Galitelli, D.; Mascia, T. Grafting alters tomato transcriptome and enhances tolerance to an airborne virus infection. Sci. Rep. 2020, 10, 2538. [Google Scholar] [CrossRef]
- Webster, A.D. Rootstock and interstock effects on deciduous fruit tree vigour, precocity, and yield productivity. N. Z. J. Crop Hort. Sci. 1995, 23, 373–382. [Google Scholar] [CrossRef]
- Rubio, M.; Ballester, A.R.; Olivares, P.M.; Castro de Moura, M.; Dicenta, F.; Martínez-Gómez, P. Gene expression analysis of Plum pox virus (Sharka) Susceptibility/Resistance in apricot (Prunus armeniaca L.). PLoS ONE 2015, 10, e0144670. [Google Scholar] [CrossRef]
- Yu, B.; Yang, Z.; Li, J.; Minakhina, S.; Yang, M.; Padgett, R.W.; Steward, R.; Chen, X. Methylation as a Crucial Step in Plant microRNA Biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collum, T.D.; Stone, A.L.; Sherman, D.J.; Rogers, E.E.; Dardick, C.; Culver, J.N. Translatome profiling of Plum pox virus infected leaves in European plum reveals temporal and spatial coordination of defense responses in phloem tissues. Mol. Plant-Microbe Interact. 2020, 33, 66–77. [Google Scholar] [CrossRef]
- Beffa, R.S.; Hofer, R.M.; Thomas, M.; Meins, F. Decreased susceptibility to virus disease of β-1,3-glucanase-deficient plants generated by antisense transformation. Plant Cell 1996, 8, 1001–1011. [Google Scholar] [CrossRef]
- Díaz-Vivancos, P.; Rubio, M.; Periago, P.; Martínez-Gómez, P.; Hernández, J.A. The apoplastic antioxidant system in Prunus: Response to long-term Plum pox virus infection. J. Exp. Bot. 2006, 57, 3813–3824. [Google Scholar] [CrossRef] [Green Version]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic Acid Biosynthesis in Plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.P.; Luo, T.; Fu, H.W.; Wang, L.; Tan, Y.Y.; Huang, J.Z.; Wang, Q.; Ye, G.Y.; Gatehouse, A.M.; Lou, Y.G.; et al. Resistance of rice to insect pests mediated by suppression of serotonin biosynthesis. Nat. Plants 2018, 4, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Pasin, F.; Shan, H.; García, B.; Müller, M.; San León, D.; Ludman, M.; Fresno, D.H.; Fátyol, K.; Munné-Bosch, S.; Rodrigo, G.; et al. Abscisic acid connects phytohormone signaling with RNA metabolic pathways and promotes an antiviral response that is evaded by a self-controlled RNA Virus. Plant Commun. 2009, 1, 100099. [Google Scholar] [CrossRef]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in Abscisic Acid signaling. Plant Cell 2013, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alazem, M.; Lin, N.S. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Ren, T.; Qu, F.; Morris, J. HRT Gene Function Requires Interaction between a NAC Protein and Viral Capsid Protein to Confer Resistance to Turnip Crinkle Virus. Plant Cell 2000, 12, 1917–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinoza, C.; Vega, A.; Schlauch, K.; Arce-Jhonson, P. Gene expression associated with compatible viral diseases in grapevine cultivars. Funct. Integr. Gen. 2017, 7, 95–110. [Google Scholar] [CrossRef]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolrà, R.; Poschenrieder, C. A Role for Zinc in plant defense against pathogens and herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Mani, R.S. The emerging role of speckle-type POZ protein (SPOP) in cancer development. Drug Discov. Today 2014, 19, 1498–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decroocq, V.; Salvador, B.; Sicard, O.; Glasa, M.; Cosson, P.; Svanella-Dumas, L.; Revers, F.; García, J.A.; Candresse, T. The determinant of potyvirus ability to overcome the RTM resistance of Arabidopsis thaliana maps to the N-terminal region of the coat protein. Mol. Plant-Microbe Intear. 2009, 22, 1302–1311. [Google Scholar] [CrossRef] [Green Version]
- Pagny, G.; Paulstephenraj, P.S.; Poque, S.; Sicard, O.; Cosson, P.; Eyquard, J.P.; Caballero, M.; Chague, A.; Gourdon, G.; Negrel, L.; et al. Family-based linkage and association mapping reveals novel genes affecting Plum pox virus infection in Arabidopsis thaliana. New Phytol. 2012, 196, 873–886. [Google Scholar] [CrossRef]
- Zuriaga, E.; Soriano, J.M.; Zhebentyayeva, T.N.; Romero, C.; Dardick, C.; Cañizares, J. Genomic analysis reveals MATH gene(s) as candidate for Plum pox virus (PPV) resistance in apricot (Prunus armeniaca L.). Mol. Plant Pathol. 2013, 13, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Zuriaga, E.; Romero, C.; Blanca, J.M.; Badenes, M.L. Resistance to Plum pox virus (PPV) in apricot (Prunus armeniaca L.) is associated with down-regulation of two MATHd genes. BMC Plant Biol. 2018, 18, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polo-Oltra, Á.; Romero, C.; López, I.; Badenes, M.L.; Zuriaga, E. Cost-Effective and Time-Efficient Molecular Assisted Selection for PPV Resistance in Apricot Based on ParPMC2 Allele-Specific PCR. Agronomy 2020, 10, 1292. [Google Scholar] [CrossRef]
- Bernhard, R.; Marénaud, C.; Sutic, D. Le pêcher GF305 indicateur polyvalent des virus des espèces à noyaux. Ann. Phytopathol. 1969, 1, 603–617. [Google Scholar]
- Rubio, M.; García-Ibarra, A.; Martínez-Gómez, P.; Dicenta, F. Analysis of the main factors involved in the evaluation of Prunus resistance to Plum pox virus (Sharka) in control greenhouse conditions. Sci. Hort. 2009, 123, 46–50. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Verde, I.; Jenkins, J.; Dondini, L.; Micali, S.; Pagliarani, G.; Vendramin, E.; Paris, R.; Aramini, V.; Gazza, L.; Rossini, L.; et al. The Peach v2.0 release: High-resolution linkage mapping and deep resequencing improve chromosome-scale assembly and contiguity. BMC Gen. 2017, 18, 225. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Sonali Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageu, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-Seq. Nat. Biotech. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Bi, C.L.; Lang, N.; Li, Y.Z.; Xu, C.; Zhang, Y.Q.; Zhai, A.X.; Cheng, Z.F. RNA-Seq methods for identifying diffrentially expressed gene in human pancreatic islet treated with pro-inflamatory cytokines. Mol. Biol. Rep. 2014, 41, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a newPANTHER GO-slim and improvements in enrichmentanalysis tools. Nucleic Acids Res. 2009, 47, D419–D426. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucl. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycle 1 | Cycle 2 | Cycle 3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Model | Sample | N 1 | Symptoms 2 | ELISA-DASI 3 | N 1 | Symptoms 2 | ELISA-DASI 3 | RT-PCR 4 | N 1 | Symptoms 2 | ELISA-DASI 3 | RT-PCR 4 |
| GF305 Control | A | 8 | 0 (0) | 0 (0.05) | 8 | 0 (0) | 0 (0.05) | 0 | 7 | 0 (0) | 0 (0.06) | 0 |
| GF305 + PPV | B | 8 | 8 (3.1) | 8 (3.51) | 8 | 8 (4) | 8 (3.12) | 8 | 7 | 7 (3.1) | 7 (1.35) | 7 |
| GF305 Control + Garrigues | C | 8 | 0 (0) | 0 (0.05) | 7 | 0 (0) | 0 (0.01) | 0 | 6 | 0 (0) | 0 (0.09) | 0 |
| GF305 + PPV + Garrigues | D | 8 | 2 (1) | 1 (1.23) | 6 | 1 (1) | 5 (1.59) | 5 | 5 | 0 (0) | 1 (0.28) | 1 |
| GF305 Control + Garrigues + PPV | E | 8 | 0 (0) | 0 (0.06) | 8 | 0 (0) | 0 (0.03) | 1 | 7 | 0 (0) | 0 (0.05) | 0 |
| GF305 Control + Garrigues | F | 8 | 0 (0) | 0 (0.06) | 6 | 0 (0) | 0 (0.05) | 0 | 6 | 0 (0) | 0 (0.06) | 0 |
| GF305 + PPV + Garrigues | G | 8 | 0 (0) | 0 (0.05) | 7 | 0 (0) | 0 (0.05) | 0 | 5 | 0 (0) | 0 (0.07) | 0 |
| Treatments | Sample | Total Reads | Clean Reads | Reads Mapped P. persica v 2.0 | Reads Mapped PPV Genome |
|---|---|---|---|---|---|
| GF305 Control | A (A’+A”) | 151,699,038 | 151,376,821 | 123,331,659 (81.5%) | 15 (0.00001%) |
| GF305 + PPV | B (B’+B”) | 159,272,116 | 158,812,159 | 125,187,942 (78.8%) | 2,071,915 (1.30%) |
| GF305 Control + Garrigues | C (C’+C”) | 142,956,238 | 142,296,978 | 113,078,192 (79.5%) | 152 (0.0001%) |
| GF305 + PPV + Garrigues | D (D’+D”) | 143,511,454 | 143,041,550 | 109,499,620 (76.6%) | 80,703 (0.06%) |
| GF305 Control + Garrigues + PPV | E (E’+E”) | 148,551,466 | 148,233,453 | 114,097,915 (77.0%) | 1904 (0.0013%) |
| GF305 Control + Garrigues | F (F’+F”) | 140,514,568 | 140,158,744 | 88,206,273 (62.9%) | 44 (0.00003%) |
| GF305 + PPV + Garrigues | G (G’+G”) | 138,038,412 | 137,569,619 | 101,163,774 (73.5%) | 45 (0.00003%) |
| Comparison of Transcriptomes | Sample Comparison | Total Identified Genes | Total Tested Genes | Total DEG Genes | Filtered DEG Genes | DEG Up | DEG Down |
|---|---|---|---|---|---|---|---|
| Effect of PPV inoculation/infection | |||||||
| GF305 + PPV vs. GF305 Control | B vs. A | 27,121 | 19,335 | 1102 | 44 | 9 | 35 |
| GF305 + PPV + Garrigues vs. GF305 Control + Garrigues | D vs. C | 27,036 | 18,832 | 548 | 29 | 20 | 9 |
| GF305 + PPV + Garrigues vs. GF305 Control + Garrigues | G vs. F | 27,203 | 18,808 | 439 | 0 | - | - |
| Effect of almond grafting | |||||||
| GF305 Control vs. GF305 Control + Garrigues | C vs. A | 27,011 | 19,906 | 823 | 18 | 10 | 8 |
| Effect of PPV infection and almond Grafting | |||||||
| GF305 + PPV + Garrigues vs. GF305 + PPV | D vs. B | 27,143 | 20,643 | 1163 | 31 | 23 | 8 |
| GF305 Control + Garrigues + PPV vs. GF305 + PPV | E vs. B | 27,131 | 19,362 | 1113 | 47 | 30 | 17 |
| Effect of shifting the order of inoculation and grafting | |||||||
| GF305 Control + Garrigues + PPV vs. GF305 + PPV + Garrigues | E vs. D | 26,589 | 18,865 | 627 | 28 | 6 | 22 |
| Total | 5815 | 197 | 98 | 99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio, M.; Martínez-García, P.J.; Nikbakht-Dehkordi, A.; Prudencio, Á.S.; Gómez, E.M.; Rodamilans, B.; Dicenta, F.; García, J.A.; Martínez-Gómez, P. Gene Expression Analysis of Induced Plum pox virus (Sharka) Resistance in Peach (Prunus persica) by Almond (P. dulcis) Grafting. Int. J. Mol. Sci. 2021, 22, 3585. https://doi.org/10.3390/ijms22073585
Rubio M, Martínez-García PJ, Nikbakht-Dehkordi A, Prudencio ÁS, Gómez EM, Rodamilans B, Dicenta F, García JA, Martínez-Gómez P. Gene Expression Analysis of Induced Plum pox virus (Sharka) Resistance in Peach (Prunus persica) by Almond (P. dulcis) Grafting. International Journal of Molecular Sciences. 2021; 22(7):3585. https://doi.org/10.3390/ijms22073585
Chicago/Turabian StyleRubio, Manuel, Pedro J. Martínez-García, Azam Nikbakht-Dehkordi, Ángela S. Prudencio, Eva M. Gómez, Bernardo Rodamilans, Federico Dicenta, Juan A. García, and Pedro Martínez-Gómez. 2021. "Gene Expression Analysis of Induced Plum pox virus (Sharka) Resistance in Peach (Prunus persica) by Almond (P. dulcis) Grafting" International Journal of Molecular Sciences 22, no. 7: 3585. https://doi.org/10.3390/ijms22073585
APA StyleRubio, M., Martínez-García, P. J., Nikbakht-Dehkordi, A., Prudencio, Á. S., Gómez, E. M., Rodamilans, B., Dicenta, F., García, J. A., & Martínez-Gómez, P. (2021). Gene Expression Analysis of Induced Plum pox virus (Sharka) Resistance in Peach (Prunus persica) by Almond (P. dulcis) Grafting. International Journal of Molecular Sciences, 22(7), 3585. https://doi.org/10.3390/ijms22073585