
Brachygnathia Inferior in Cloned Dogs Is Possibly Correlated with Variants of Wnt Signaling Pathway Initiators

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
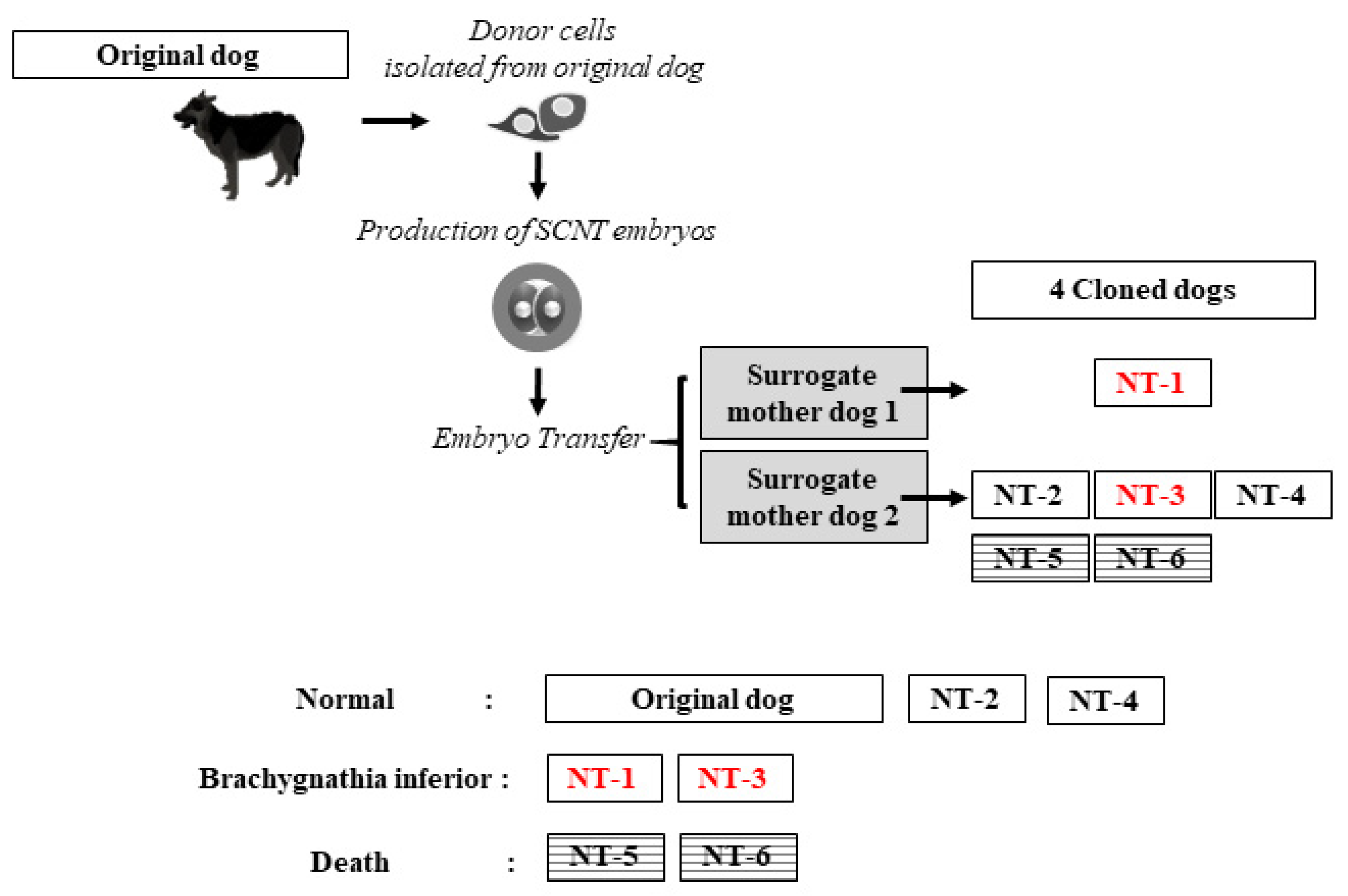
2.1. Production of Cloned Dogs
2.2. Care and Feeding of Cloned Offspring
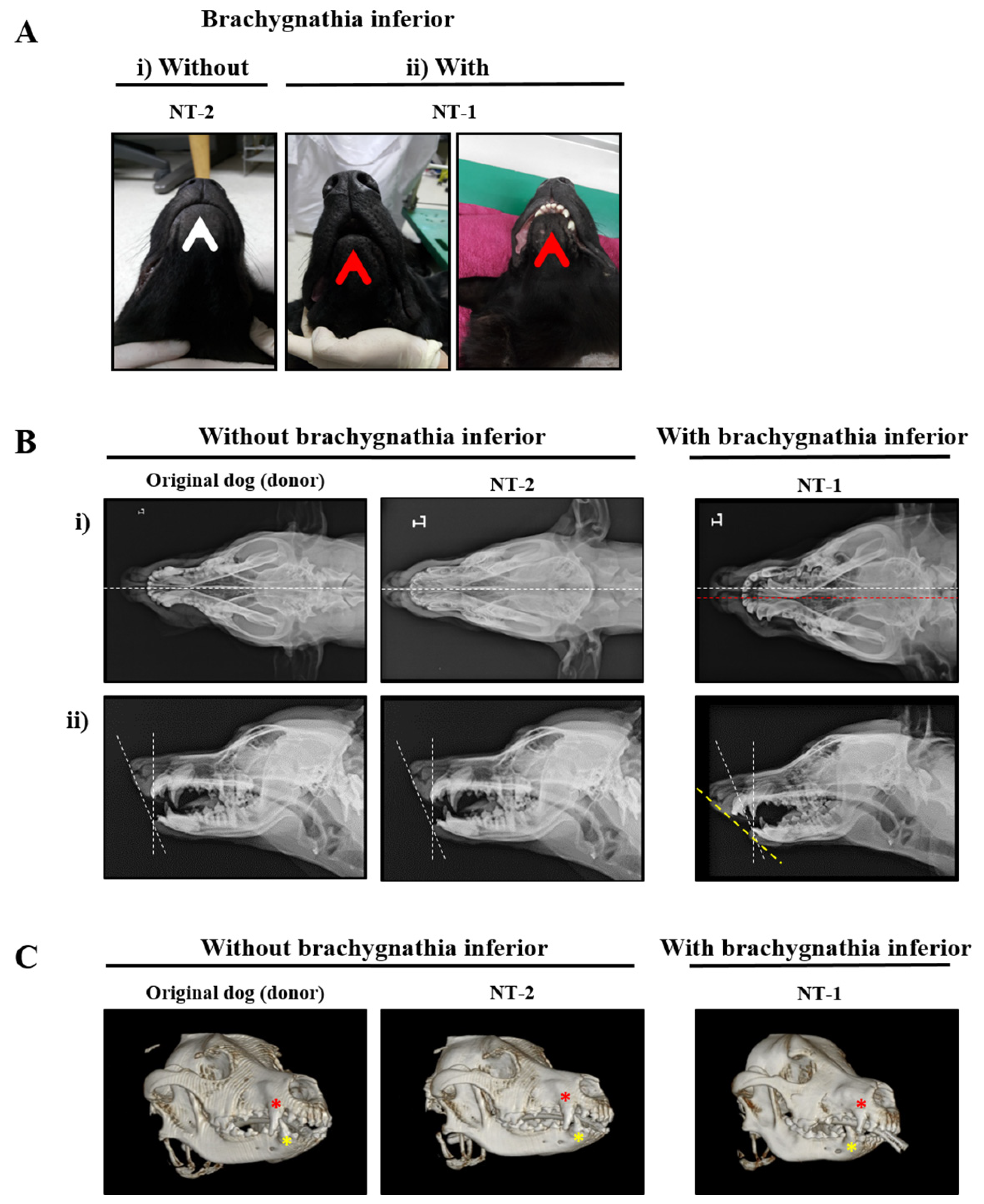
2.3. Clinical Diagnosis of Brachygnathia Inferior
2.4. Chromosomal Aberrations in the Donor and Cloned Dogs
2.5. Identification and Validation of Candidate Genes for Brachygnathia Inferior by Whole-Genome Sequencing
2.6. Interactive Network Analysis of Candidate Genes for Brachygnathia Inferior
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. Cloning of Dogs
5.3. Microsatellite Analysis for Confirmation of Paternity
5.4. Hematological and Biochemical Analysis of Blood
5.5. X-ray and CT Imaging
5.6. Karyotype Analysis
5.7. Whole-Genome Sequencing, Sequence Mapping, and Variant Calling
5.8. Predictive Functional and Interaction Analyses of Brachygnathia Inferior Candidate Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SCNT | Somatic cell nuclear transfer |
| CT | Computed tomography |
| WGS | Whole-genome sequencing |
| Wnt | Wingless-related integration site |
| DNA | Deoxyribonucleic acid |
| NT | Nuclear transfer |
| SMD | Surrogate mother dogs |
| CBC | Complete blood count |
| RBC | Red blood cell |
| MCV | Mean corpuscular volume |
| MCH | Mean corpuscular hemoglobin |
| MCHC | Mean corpuscular hemoglobin concentration |
| PDW | Platelet distribution width |
| WBC | White blood cell |
| MPV | Mean platelet volume |
| RDW | Red cell distribution width |
| PCT | Plateletcrit |
| BUN | Blood urea nitrogen |
| GGT | Gamma-glutamyl transferase |
| ALB | Albumin |
| TB | Total bilirubin |
| TP | Total protein |
| ALT | Alanine aminotransferase |
| AST | Aspartate aminotransferase |
| SNVs | Single nucleotide variations |
| Indels | Insertions/deletions |
| GO | Gene Ontology |
| PANTHER | Protein analysis through evolutionary relationships |
| DAVID | Database for annotation, visualization, and integrated discovery |
| GnRH | Gonadotropin-releasing hormone |
| STRING | Search tool for the retrieval of interacting genes/proteins |
| PCP | Planar cell polarity |
| FBS | Fetal bovine serum |
| BP | Biological process |
References
- Lee, B.C.; Kim, M.K.; Jang, G.; Oh, H.J.; Yuda, F.; Kim, H.J.; Shamim, M.H.; Kim, J.J.; Kang, S.K.; Schatten, G.; et al. Developmemtal technology: Dogs cloned from adult somatic cells. Nature 2005, 436, 641. [Google Scholar] [CrossRef] [PubMed]
- Keefer, C.L.; Baldassarre, H.; Keyston, R.; Wang, B.; Bhatia, B.; Bilodeau, A.S.; Zhou, J.F.; Leduc, M.; Downey, B.R.; Lazaris, A.; et al. Generation of dwarf goat (Capra hircus) clones following nuclear transfer with transfected and nontransfected fetal fibroblasts and in vitro-matured oocytes. Biol. Reprod. 2001, 64, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keefer, C.L. Artificial cloning of domestic animals. Proc. Natl. Acad. Sci. USA 2015, 112, 8874–8878. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Oh, H.J.; Hwang, S.Y.; Hur, T.Y.; Lee, B.C. Health and temperaments of cloned working dogs. J. Vet. Sci. 2018, 19, 585–591. [Google Scholar] [CrossRef]
- Kim, M.J.; Oh, H.J.; Kim, G.A.; Setyawan, E.M.N.; Choi, Y.B.; Lee, S.H.; Petersen-Jones, S.M.; Ko, C.M.J.; Lee, B.C. Birth of clones of the world’s first cloned dog. Sci. Rep. 2017, 7, 3–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Zhao, M.; No, J.; Nam, Y.; Im, G.S.; Hur, T.Y. Dog cloning with in vivo matured oocytes obtained using electric chemiluminescence immunoassay-predicted ovulation method. PLoS ONE 2017, 12, e0173735. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Cho, Y.S.; Kim, H.; Jho, S.; Son, B.; Choi, J.Y.; Kim, S.; Lee, B.C.; Bhak, J.; Jang, G. Whole genome comparison of donor and cloned dogs. Sci. Rep. 2013, 3, 1–4. [Google Scholar] [CrossRef]
- De Sousa, P.A.; King, T.; Harkness, L.; Young, L.E.; Walker, S.K.; Wilmut, I. Evaluation of gestational deficiencies in cloned sheep fetuses and placentae. Biol. Reprod. 2001, 65, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.R.; Burghardt, R.C.; Jones, K.; Long, C.R.; Looney, C.R.; Shin, T.; Spencer, T.E.; Thompson, J.A.; Winger, Q.A.; Westhusin, M.E. Evidence for placental abnormality as the major cause of mortality in first-trimester somatic cell cloned bovine fetuses. Biol. Reprod. 2000, 63, 1787–1794. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, T.; Ganter, M.; Distl, O.; Staszyk, C. Cranial morphology in the brachygnathic sheep. BMC Vet. Res. 2016, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shatab, M.S.; Kumar, N.; Singh, M.; Kumar, S.; Seckhon, M.S.; Dhindsa, S.S. Brachygnathia (parrot mouth) inferior calf in a murrah buffalo. Buffalo Bull. 2018, 37, 101–103. [Google Scholar]
- Dierks, C.; Hoffmann, H.; Heinrich, F.; Hellige, M.; Hewicker-Trautwein, M.; Distl, O. Suspected X-linked facial dysmorphia and growth retardation in related labrador retriever puppies. Vet. J. 2017, 220, 48–50. [Google Scholar] [CrossRef]
- Yin, R.; Kwoh, C.K.; Zheng, J. Whole genome sequencing analysis: Computational pipelines and workflows in bioinformatics. In Encyclopedia of Bioinformatics and Computational Biology: ABC of Bioinformatics; Shoba, R., Gribskov, M., Nakai, K., Christian, S., Eds.; Elsevier: Amsterda, The Netherlands, 2018; Volume 3, pp. 176–183. [Google Scholar]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H.S. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Wakayama, T.; Perry, A.C.F.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Tani, T.; Sotomaru, Y.; Kurokawa, K.; Kato, J.Y.; Doguchi, H.; Yasue, H.; Tsunoda, Y. Eight calves cloned from somatic cells of a single adult. Science 1998, 282, 2095–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baguisi, A.; Behboodi, E.; Melican, D.T.; Pollock, J.S.; Destrempes, M.M.; Cammuso, C.; Williams, J.L.; Nims, S.D.; Porter, C.A.; Midura, P.; et al. Production of goats by somatic cell nuclear transfer. Nat. Biotechnol. 1999, 17, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Polejaeva, I.A.; Chen, S.H.; Vaught, T.D.; Page, R.L.; Mullins, J.; Ball, S.; Dai, Y.; Boone, J.; Walker, S.; Ayares, D.L.; et al. Cloned pigs produced by nuclear transfer from adult somatic cells. Nature 2000, 407, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Oh, H.J.; Kim, G.A.; Jo, Y.K.; Choi, J.; Kim, H.J.; Choi, H.Y.; Kim, H.W.; Choi, M.C.; Lee, B.C. Reduced birth weight, cleft palate and preputial abnormalities in a cloned dog. Acta Vet. Scand. 2014, 56, 18–21. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Oda, M.; Toyoshima, Y.; Wakayama, T.; Tanaka, M.; Hattori, N.; Yoshida, N.; Ohgane, J.; Yanagimachi, R.; Shiota, K. Placentomegaly in cloned mouse concepti caused by expansion of the spongiotrophoblast layer. Biol. Reprod. 2001, 65, 1813–1821. [Google Scholar] [CrossRef]
- Dean, W.; Santos, F.; Stojkovic, M.; Zakhartchenko, V.; Walter, J.; Wolf, E.; Reik, W. Conservation of methylation reprogramming in mammalian development: Aberrant reprogramming in cloned embryos. Proc. Natl. Acad. Sci. USA 2001, 98, 13734–13738. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, C.; Huyser, C.; Egli, D.; Pepper, M.S. Lessons learned from somatic cell nuclear transfer. Int. J. Mol. Sci. 2020, 21, 2314. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Cloning: Experience from the mouse and other animals. Mol. Cell. Endocrinol. 2002, 187, 241–248. [Google Scholar] [CrossRef]
- Iannuzzi, L.; PiaDiMeo, G.; Leifsson, P.S.; Eggen, A.; Christensen, K. A case of trisomy 28 in cattle revealed by both banding and FISH-mapping techniques. Hereditas 2001, 134, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Mayr, B.; Krutzler, H.; Auer, H.; Schleger, W.; Sasshofer, K.; Glawischig, E. A viable calf with trisomy 22. Cytogenet. Genome Res. 1985, 39, 77–79. [Google Scholar] [CrossRef]
- Schwarzenbacher, H.; Wurmser, C.; Flisikowski, K.; Misurova, L.; Jung, S.; Langenmayer, M.C.; Schnieke, A.; Knubben-Schweizer, G.; Fries, R.; Pausch, H. A frameshift mutation in GON4L is associated with proportionate dwarfism in Fleckvieh cattle. Genet. Sel. Evol. 2016, 48, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Woolley, S.A.; Hayes, S.E.; Shariflou, M.R.; Nicholas, F.W.; Willet, C.E.; O’Rourke, B.A.; Tammen, I. Molecular basis of a new ovine model for human 3M syndrome-2. BMC Genet. 2020, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.G.; Piripi, S.A.; Dittmer, K.E. Inherited abnormalities of skeletal development in sheep. Vet. J. 2008, 177, 324–333. [Google Scholar] [CrossRef]
- Wolf, Z.T.; Leslie, E.J.; Arzi, B.; Jayashankar, K.; Karmi, N.; Jia, Z.; Rowland, D.J.; Young, A.; Safra, N.; Sliskovic, S.; et al. A LINE-1 insertion in DLX6 is responsible for cleft palate and mandibular abnormalities in a canine model of pierre robin sequence. PLoS Genet. 2014, 10, e1004257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenebeck, J.J.; Hutchinson, S.A.; Byers, A.; Beale, H.C.; Carrington, B.; Faden, D.L.; Rimbault, M.; Decker, B.; Kidd, J.M.; Sood, R.; et al. Variation of BMP3 contributes to dog breed skull diversity. PLoS Genet. 2012, 8, e1002849. [Google Scholar] [CrossRef] [Green Version]
- Dashti, M.J.S.; Gamieldien, J. A practical guide to filtering and prioritizing genetic variants. Biotechniques 2017, 62, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [Green Version]
- Brault, V.; Moore, R.; Kutsch, S.; Ishibashi, M.; Rowitch, D.H.; McMahon, A.P.; Sommer, L.; Boussadia, O.; Kemler, R. Inactivation of the β-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development 2001, 128, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Acebron, S.P.; Niehrs, C. β-catenin-independent roles of Wnt/LRP6 signaling. Trends Cell Biol. 2016, 26, 956–967. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Haseeb, M.; Pirzada, R.H.; Ain, Q.U.; Choi, S. Wnt signaling in the regulation of immune cell and cancer therapeutics. Cells 2019, 8, 1380. [Google Scholar] [CrossRef] [Green Version]
- Bonnans, C.; Flacelière, M.; Grillet, F.; Dantec, C.; Desvignes, J.P.; Pannequin, J.; Severac, D.; Dubois, E.; Bibeau, F.; Escriou, V.; et al. Essential requirement for β-arrestin2 in mouse intestinal tumors with elevated Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 3047–3052. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Chiang, Y.J.; Gao, C.; Xue, H.; Xu, J.; Ning, Y.; Hodes, R.J.; Gao, X.; Chen, Y.G. Loss of Dact1 disrupts planar cell polarity signaling by altering dishevelled activity and leads to posterior malformation in mice. J. Biol. Chem. 2010, 285, 11023–11030. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Xiang, T.; Li, L.L.; Su, X.; Shu, X.; Luo, X.; Huang, J.; Yuan, Y.; Peng, W.; Oberst, M.; et al. DACT1, an antagonist to Wnt/β-catenin signaling, suppresses tumor cell growth and is frequently silenced in breast cancer. Breast Cancer Res. 2013, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Topol, L.; Lee, H.; Wu, J. Wnt5a and Wnt5b exhibit distinct activities in coordinating chondrocyte proliferation and differentiation. Development 2003, 130, 1003–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suthon, S.; Perkins, R.S.; Bryja, V.; Miranda-Carboni, G.A.; Krum, S.A. WNT5B in physiology and disease. Front. Cell Dev. Biol. 2021, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, J.; Azuma, Y.; Hoshi, K.; Kii, I.; Takeshita, S.; Ohta, T.; Ozawa, H.; Takeichi, M.; Chisaka, O.; Kudo, A. Targeted disruption of cadherin-11 leads to a reduction in bone density in calvaria and long bone metaphyses. J. Bone Miner. Res. 2001, 16, 1265–1271. [Google Scholar] [CrossRef]
- Aberle, H.; Schwartz, H.; Kemler, R. Cadherin-catenin complex: Protein interactions and their implications for cadherin function. J. Cell. Biochem. 1996, 61, 514–523. [Google Scholar] [CrossRef]
- Hagman, R.; Rönnberg, E.; Pejler, G. Canine uterine bacterial infection induces upregulation of proteolysis-related genes and downregulation of homeobox and zinc finger factors. PLoS ONE 2009, 4, e8039. [Google Scholar] [CrossRef]
- Hansis, C.; Barreto, G.; Maltry, N.; Niehrs, C. Nuclear reprogramming of human somatic cells by xenopus egg extract requires BRG1. Curr. Biol. 2004, 14, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Bui, H.T.; Wakayama, S.; Kishigami, S.; Kim, J.H.; Van Thuan, N.; Wakayama, T. The cytoplasm of mouse germinal vesicle stage oocytes can enhance somatic cell nuclear reprogramming. Development 2008, 135, 3935–3945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, M.; Kwon, S. Genomic polymorphism analysis using microsatellites in the jeju dogs. J. Life Sci. 2019, 29, 637–644. [Google Scholar]
- Kim, S.; Jang, H.; Kim, L.; Lim, D.; Lee, S.; Cho, Y.; Kim, T.; Seong, H.; Oh, S.; Choi, B. Paternity diagnosis using the multiplex PCR with microsatellite markers in dogs. Reprod. Dev. Biol. 2011, 35, 399–405. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | FH2537 | FH3005 | FH3372 | FH3116 | REN51C16 | REN2770O5 | FH2834 | REN204K13 | FH2097 | FH2712 | FH2998 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Donor cells | 146 | 146 | 224 | 224 | 154 | 158 | 190 | 190 | 255 | 259 | 333 | 333 | 265 | 265 | 248 | 248 | 284 | 288 | 174 | 174 | 208 | 228 |
| NT-1 | 146 | 146 | 224 | 224 | 154 | 158 | 190 | 190 | 255 | 259 | 333 | 333 | 265 | 265 | 248 | 248 | 284 | 288 | 174 | 174 | 208 | 228 |
| NT-2 | 146 | 146 | 224 | 224 | 154 | 158 | 190 | 190 | 255 | 259 | 333 | 333 | 265 | 265 | 248 | 248 | 284 | 288 | 174 | 174 | 208 | 228 |
| NT-3 | 146 | 146 | 224 | 224 | 154 | 158 | 190 | 190 | 255 | 259 | 333 | 333 | 265 | 265 | 248 | 248 | 284 | 288 | 174 | 174 | 208 | 228 |
| NT-4 | 146 | 146 | 224 | 224 | 154 | 158 | 190 | 190 | 255 | 259 | 333 | 333 | 265 | 265 | 248 | 248 | 284 | 288 | 174 | 174 | 208 | 228 |
| Parameters (Unit) | Original Dog (Donor Cells) | Brachygnathia Inferior | ||
|---|---|---|---|---|
| without | with | |||
| NT2 | NT4 | NT-1 | ||
| RBC (1012/L) | 8.85 | 5.9 | 6.74 | 7.43 |
| Hematocrit [Hct] (%) | 56.7 | 37.7 | 41.2 | 48.1 |
| Hemoglobin [Hb] (g/dL) | 19.6 | 12.1 | 14.3 | 15.8 |
| MCV (fL) | 64.1 | 63.9 | 61.1 | 64.7 |
| MCH (pg) | 22.1 | 20.5 | 21.2 | 21.3 |
| MCHC (g/dL) | 34.6 | 32.1 | 34.7 | 32.8 |
| PDW (%) | 20.2 | 18.7 | 19.5 | 18.9 |
| Reticulocyte (%) | 0.2 | 1.8 | 0.5 | 1.1 |
| Reticulocyte (103/uL) | 18.6 | 108 | 30.3 | 81.7 |
| WBC (109/L) | 13.8 | 15.7 | 19.2 | 13.86 |
| WBC-Neut (%) | 70.6 | 56.9 | 64.6 | 60 |
| WBC-Lymph (%) | 15.3 | 29.5 | 24.9 | 25.2 |
| WBC-Mono (%) | 7.8 | 8.3 | 8.7 | 7 |
| WBC-Eos (%) | 6.2 | 5.2 | 1.7 | 7.8 |
| WBC-Baso (%) | 0.1 | 0.1 | 0.1 | 0 |
| WBC-Neut (109/L) | 9.75 | 8.94 | 12.4 | 8.32 |
| WBC-Lymph (109/L) | 2.11 | 4.63 | 4.76 | 3.49 |
| WBC-Mono (109/L) | 1.08 | 1.31 | 1.66 | 0.97 |
| WBC-Eos (109/L) | 0.85 | 0.81 | 0.33 | 1.08 |
| WBC-Baso (109/L) | 0.01 | 0.01 | 0.02 | 0 |
| Platelet (109/L) | 201 | 411 | 325 | 429 |
| MPV (fL) | 12.3 | 13.1 | 13.1 | |
| RDW (fL) | 19.36 | 18.8 | 18.8 | |
| PCT (%) | 0.5 | 0.56 | 0.56 | |
| Parameters (Unit) | Original Dog (Donor Cells) | Brachygnathia Inferior | |||
|---|---|---|---|---|---|
| without | with | ||||
| NT2 | NT4 | NT-1 | NT-3 | ||
| Glucose (mg/dL) | 68 | 31 | 104 | 62 | 90 |
| BUN (mg/dL) | 13 | 13 | 19 | 13 | 4 |
| Creatinine (mg/dL) | 1.2 | 1 | 0.7 | 1.1 | 0.3 |
| BUN: Creatinine (Ratio) | 11 | 13 | 25 | 11 | 12 |
| Phosphorus-Inorganic (mg/dL) | 3 | 8 | 8.7 | 5.7 | 8.3 |
| Calcium (mg/dL) | 10.8 | 11 | 9.9 | 10.9 | 11 |
| Protein-Total (g/dL) | 7.8 | 5.7 | 5 | 7.1 | 6.1 |
| Albumin (g/dL) | 4 | 2.9 | 2.7 | 3.4 | 2.9 |
| Globulin (g/dL) | 3.8 | 2.8 | 2.3 | 3.7 | 3.2 |
| A/G ratio | 1.1 | 1.1 | 1.2 | 0.9 | 0.9 |
| ALT (U/L) | 47 | 22 | 18 | 23 | 67 |
| ALKP (U/L) | 17 | 173 | 151 | 106 | 216 |
| GGT (U/L) | 0 | 0 | 2 | 0 | 8 |
| Bilirubin-Total (mg/dL) | 0.2 | 0.1 | 0.1 | 0.1 | 0.7 |
| Cholesterol-Total (mg/dL) | 191 | 119 | 171 | 170 | 237 |
| Amylase (U/L) | 727 | 539 | 299 | 863 | 323 |
| Lipase (U/L) | 341 | 331 | 352 | 322 | 93 |
| Na+ (mmol/L) | 153 | 150 | 147 | 151 | 140.9 |
| K+ (mmol/L) | 4.2 | 5 | 5.7 | 4.3 | 5.05 |
| Na+:K+ (Ratio) | 36 | 30 | 26 | 36 | 23.5 |
| Cl− (mmol/L) | 110 | 107 | 107 | 107 | 111.3 |
| Osmorality | 301 | 294 | 296 | 297 | |
| # | Gene Symbol (Gene Name) | Variant Information | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CHR | POS | REF | ALTS | Putative Impact | Genotype | ||||||
| Without Brachygnathia Inferior | With Brachygnathia Inferior | ||||||||||
| Original Dog | NT-2 | NT-4 | NT-1 | NT-3 | |||||||
| 1 | ARRB2 (arrestin beta 2) | 5 | 31834580 | T | G | + MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 2 | CCND1 (G1/S-specific cyclin-D1) | 18 | 48505585 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 48511836 | T | TG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 3 | CDH3 (cadherin 3) | 5 | 80888237 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 80917292 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 4 | CDH7 (cadherin 7) | 1 | 11751189 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 11765372 | CATATATATATATATATAT | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 5 | CDH8 (cadherin 8) | 5 | 86401758 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 86708399 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 86742629 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 86745597 | T | TATACAC | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 87225311 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 87225313 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 85863919 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 86114270 | C | CT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 86222689 | T | G | MOD | 0/0 | 0/1 | 0/0 | 1/1 | 1/1 | |||
| 86296806 | C | G | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 86390011 | TACCCC | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 6 | CDH9 (cadherin 9) | 4 | 78675775 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 78830300 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 78840862 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 79012127 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 7 | CDH10 (cadherin 10) | 4 | 81023137 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 81051478 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 80613492 | GTC | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 80892138 | GT | G | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 80904036 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 8 | CDH11 (cadherin 11) | 5 | 84404903 | AAGAG | AAGAGAG,A | MOD | 0/0 | 0/0 | 0/0 | 0/2 | 0/2 |
| 84473862 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 9 | CDH12 (cadherin 12) | 4 | 82754279 | T | TATATATAG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 82856048 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82947779 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 83011986 | C | CGT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 83066202 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 83101151 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 81762398 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 81811393 | TTGAAA | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 81895595 | G | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 81895601 | C | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 81934549 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 81942964 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82120066 | A | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82213787 | AT | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82245479 | C | CT | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82346773 | T | G | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 82391279 | GT | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82427446 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82427448 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82567248 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82591151 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 82672898 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 10 | CDH13 (cadherin 13) | 5 | 68789330 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 69134746 | GT | G | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 11 | CDH17 (cadherin 17) | 29 | 38980475 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 38980931 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 12 | CDH18 (cadherin 18) | 4 | 84572993 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 84609771 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 84777718 | CT | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 84815886 | CT | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 84845137 | AT | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 84917669 | TTA | T,TTATA | MOD | 0/2 | 0/2 | 0/2 | 2/2 | 2/2 | |||
| 85340830 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 13 | CDH19 (cadherin 19) | 1 | 11213176 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 11219146 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 11302080 | T | TA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 14 | CDH20 (cadherin 20) | 1 | 15018520 | CTTTTTTTTTTTTT | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 15026159 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 15385740 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 15397278 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 15 | CDH23 (cadherin 23) | 4 | 22217920 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 22268845 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 22296573 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 22308343 | G | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 22521773 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 16 | CHD1L (chromodomain helicase DNA-binding protein 1-like) | 17 | 57755822 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 57801618 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 17 | CTNND2 (catenin delta 2) | 34 | 2587961 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 2605251 | C | CAG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 2683554 | C | CT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 2729501 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2790163 | CAG | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2887443 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2926914 | GA | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 3386604 | T | TTTC | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 1984298 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2045599 | A | AG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 2211983 | T | TACACAC | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 2325817 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2340793 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 2420433 | T | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 2420448 | G | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 18 | DACT1 (Dishevelled binding antagonist of beta catenin 1) | 8 | 33957483 | G | A | HIGH | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 33957480 | C | T | LOW | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 19 | EN1 (engrailed homeobox 1) | 19 | 31181296 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 31191156 | TA | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 20 | ERBB4 (Erb-b2 receptor tyrosine kinase 4) | 37 | 19286254 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 19629719 | C | CT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 19767395 | TTTC | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 20133592 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21 | FBXW11 (F-box and WD repeat domain-containing 11) | 4 | 40367311 | CT | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 40278734 | T | TA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 40314614 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 22 | FYN (tyrosine-protein kinase) | 12 | 68231298 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 68274356 | T | TTA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 23 | FZD4 (frizzled class receptor 4) | 21 | 12700289 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 24 | FZD8 (frizzled class receptor 8) | 2 | 1329228 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 1333216 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 25 | INO80 (INO80 complex ATPase subunit) | 30 | 8165408 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 8165412 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 8165416 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 8206520 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 26 | ITPR2 (inositol 1,4,5-trisphosphate receptor type 2) | 27 | 20831508 | A | AG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 20872227 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 27 | MAP3K7 (mitogen-activated protein kinase kinase kinase 7) | 12 | 49749924 | A | AT | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 49769002 | C | CT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 28 | PCDH10 (protocadherin 10) | 19 | 8973713 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 9008967 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 9157583 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 29 | PCDH11X (protocadherin-11 X-linked) | X | 68666259 | G | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 68746381 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 30 | PCDH15 (protocadherin-related 15) | 26 | 34702015 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 35097300 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 33646637 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 33672867 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 33672897 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 31 | PCDH18 (protocadherin 18) | 19 | 5101338 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 5101376 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 32 | PCDH19 (protocadherin 19) | X | 74412666 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 74418677 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 74250517 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 74256962 | G | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 74260225 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 33 | PCDH7 (protocadherin 7) | 3 | 80140770 | TAAA | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 80356304 | G | GA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 79602331 | T | TGA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 79874335 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 79951643 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 34 | PCDH9 (protocadherin 9) | 22 | 22259657 | CA | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 22267695 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 20900771 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21327589 | T | TA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 21447120 | CAG | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21629025 | T | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21682718 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21741598 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21855444 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 21977854 | G | GA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 35 | PLCB1 (phospholipase C beta 4) | 24 | 13687020 | C | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 13703533 | CCCTCACACACA | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 13506621 | AT | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 13545968 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 36 | PLCB4 (phospholipase C beta 4) | 24 | 13106067 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 13222123 | C | CT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 37 | PPP3CC (protein phosphatase 3 catalytic subunit gamma) | 25 | 34764968 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 34812396 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 38 | PRKCA (protein kinase C alpha) | 9 | 13673919 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 13871044 | TAA | T,TA | MOD | 0/1 | 0/1 | 0/1 | 0/2 | 1/2 | |||
| 13872388 | G | GC | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 13906274 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 13963246 | G | GT | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 13987502 | GA | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 39 | PRKCE (protein kinase C epsilon type) | 10 | 48018924 | G | GT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 48021663 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 48362949 | A | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 48362950 | A | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 48362951 | A | T | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 48452967 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 40 | PRKCH (protein kinase C epsilon) | 8 | 36326004 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 36445470 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 41 | PRKCQ (protein kinase C theta) | 2 | 29254232 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 29254236 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 29246453 | C | CA | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 42 | TBL1X (transducin beta like 1 X-linke) | X | 6446386 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 6446390 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 43 | TBXT (T-box transcription factor T) | 1 | 53860297 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 53860299 | T | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 53916908 | G | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 54140252 | G | A | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 44 | TCF7L2 (transcription factor 7 like 2) | 28 | 23954797 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 23973312 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 24061992 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 45 | TLE1 (TLE family member 1, transcriptional corepressor) | 1 | 77969485 | G | GT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 78272418 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 77521907 | A | AT | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 77529973 | CATAT | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 46 | WNT5B (Wnt family member 5B) | 27 | 43705981 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 43718076 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 47 | ENSCAFG00000008115 | 17 | 44082433 | CTTTTTT | C,CTTTT | MOD | 0/2 | 0/2 | 0/2 | 0/1 | 0/1 |
| 44366744 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 44392169 | C | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 48 | CTNNA3 (catenin alpha 3) | 4 | 17723198 | C | CTCTG | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 |
| 18058494 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 18069599 | CT | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 16891381 | CA | C | MOD | 0/1 | 0/1 | 0/1 | 1/1 | 1/1 | |||
| 16989930 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 16989932 | A | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 17291872 | TA | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 49 | ENSCAFG00000023180 | X | 99421688 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 99421696 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421697 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421711 | G | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421728 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421731 | G | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421735 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421993 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99421995 | C | T | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99422007 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 99422009 | A | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 50 | LOC489647 (cadherin-1-like) | 5 | 63556214 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 |
| 63556216 | G | A | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 63556219 | C | G | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 63556227 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 63556230 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
| 63556233 | T | C | MOD | 0/0 | 0/0 | 0/0 | 0/1 | 0/1 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choe, Y.-h.; Hur, T.-Y.; Lee, S.-L.; Lee, S.; Lim, D.; Choi, B.-H.; Jeong, H.; No, J.-G.; Ock, S.A. Brachygnathia Inferior in Cloned Dogs Is Possibly Correlated with Variants of Wnt Signaling Pathway Initiators. Int. J. Mol. Sci. 2022, 23, 475. https://doi.org/10.3390/ijms23010475
Choe Y-h, Hur T-Y, Lee S-L, Lee S, Lim D, Choi B-H, Jeong H, No J-G, Ock SA. Brachygnathia Inferior in Cloned Dogs Is Possibly Correlated with Variants of Wnt Signaling Pathway Initiators. International Journal of Molecular Sciences. 2022; 23(1):475. https://doi.org/10.3390/ijms23010475
Chicago/Turabian StyleChoe, Yong-ho, Tai-Young Hur, Sung-Lim Lee, Seunghoon Lee, Dajeong Lim, Bong-Hwan Choi, Haeyun Jeong, Jin-Gu No, and Sun A Ock. 2022. "Brachygnathia Inferior in Cloned Dogs Is Possibly Correlated with Variants of Wnt Signaling Pathway Initiators" International Journal of Molecular Sciences 23, no. 1: 475. https://doi.org/10.3390/ijms23010475
APA StyleChoe, Y.-h., Hur, T.-Y., Lee, S.-L., Lee, S., Lim, D., Choi, B.-H., Jeong, H., No, J.-G., & Ock, S. A. (2022). Brachygnathia Inferior in Cloned Dogs Is Possibly Correlated with Variants of Wnt Signaling Pathway Initiators. International Journal of Molecular Sciences, 23(1), 475. https://doi.org/10.3390/ijms23010475