1. Introduction
Bone is a calcified tissue that provides a framework for the body [
1]. The integrity of bony structures is maintained by remodeling, whereby old or injured tissue is resorbed by osteoclasts and new bone is generated by osteoblasts [
1,
2]. However, under infectious conditions, excess osteoclast formation stemming from osteolytic diseases such as periodontitis and osteomyelitis induces high bone turnover and, ultimately, causes net bone loss and architectural bone decline [
3,
4]. Periodontitis in particular, which is associated with inflammatory alveolar bone loss, is a health threat at all ages [
5]. Pathogenic invasion from the oral environment can activate inflammation, which increases osteoclast activity in the alveolar bone and disrupts the balance between the activity of osteoclasts and osteoblasts [
6,
7]. It is thus conceivable that the external environment plays a crucial role in regulating bone remodeling.
Periodontitis is commonly initiated by accumulated dental deposits such as dental plaque and calculus [
5,
8]. In response to these deposits, inflammatory mediators such as prostaglandins, matrix metalloproteinases, and cytokines, are released in periodontal tissue [
5,
9]. Among them, proinflammatory cytokine IL-1β is a potent activator of bone resorption [
10]. Bacterial components contained in dental deposits can be recognized by host immune sensors such as Toll-like receptors (TLRs), leading to the nuclear translocation of nuclear factor (NF)-κB, resulting in pro-IL-1β production, which remains biologically inactive in the cytosol [
6,
9,
11]. For IL-1β maturation, 31 kDa pro-IL-1β must be proteolytically processed to generate the active 17 kDa form [
12]. During this process, bacterial components or dental calculus activate the nucleotide-binding oligomerization domain, leucine-rich repeat, and pyrin domain-containing 3 (NLRP3) inflammasome, which plays an important role in the immune system and consists of NLRP3, apoptosis-associated speck-like protein containing a caspase-1 recruitment domain (ASC), and pro-caspase-1 [
8,
13,
14]. During formation of the NLRP3 assembly, cysteine protease pro-caspase-1 converts into active caspase-1 via autocatalysis, promoting IL-1β maturation [
15,
16]. Mature IL-1β has the capacity to up-regulate osteoclast formation and promote bone resorption in inflammatory diseases such as periodontitis and osteomyelitis [
4,
5]. Although the balance between bone formation and resorption is maintained under healthy conditions, the NLRP3 inflammasome plays an important role in IL-1β production, which under infectious conditions shifts the balance of bone homeostasis toward greater bone resorption.
It is widely recognized that the NLRP3 inflammasome is involved in bone resorption following infection challenge [
17,
18]. However, the role of the NLRP3 inflammasome under physiological conditions remains unclear. A recent study revealed that bone matrix components activate the NLRP3 inflammasome and that bone particles cause exuberant osteoclastogenesis in wild-type (WT) cells, but less so in
Nlrp3−/− cells in the presence of receptor activator of nuclear factor-κB ligand (RANKL), a master regulator of osteoclast formation. This suggests that the NLRP3 inflammasome activation by bone matrix components, which can be generated in both physiological and pathological bone turnover, amplifies bone resorption [
19]. Under physiological conditions, bone resorption by osteoclasts and bone formation by osteoblasts are repeated continuously to maintain bone homeostasis [
1,
2]. During osteoclastogenic events, RANKL interacts with receptor activator of nuclear factor-κB (RANK) and mediates osteoclast formation. RANKL–RANK interaction can induce cellular stress, which markedly elevates levels of reactive oxygen species (ROS) [
20,
21]. ROS are thought to be one of the intracellular messengers responsible for NLRP3 inflammasome activation [
22]. Considering these interconnected links between RANKL, ROS, and NLRP3, we hypothesized that RANKL may activate the NLRP3 inflammasome in osteoclastogenesis under physiological conditions. We therefore investigated the role of the NLRP3 inflammasome in osteoclast differentiation in bone marrow macrophages (BMMs) under infectious and physiological conditions. In addition, we elucidated the role of RANKL in NLRP3 inflammasome activation during osteoclastogenesis.
For this study, we generated osteoclasts from BMMs under the presence or absence of NLRP3 inflammasome inhibitors to investigate the role of the NLRP3 inflammasome in osteoclastogenesis under physiological and infectious conditions. We used pit assays to assess the bone resorption capacity. Inflammasome assembly was confirmed via ASC speck formation in the presence or absence of a ROS inhibitor. IL-1β levels were analyzed via quantitative reverse-transcription–polymerase chain reaction (qRT-PCR) and enzyme-linked immunosorbent assay (ELISA). Finally, we performed cytotoxicity assays to further analyze the underlying mechanisms.
3. Discussion
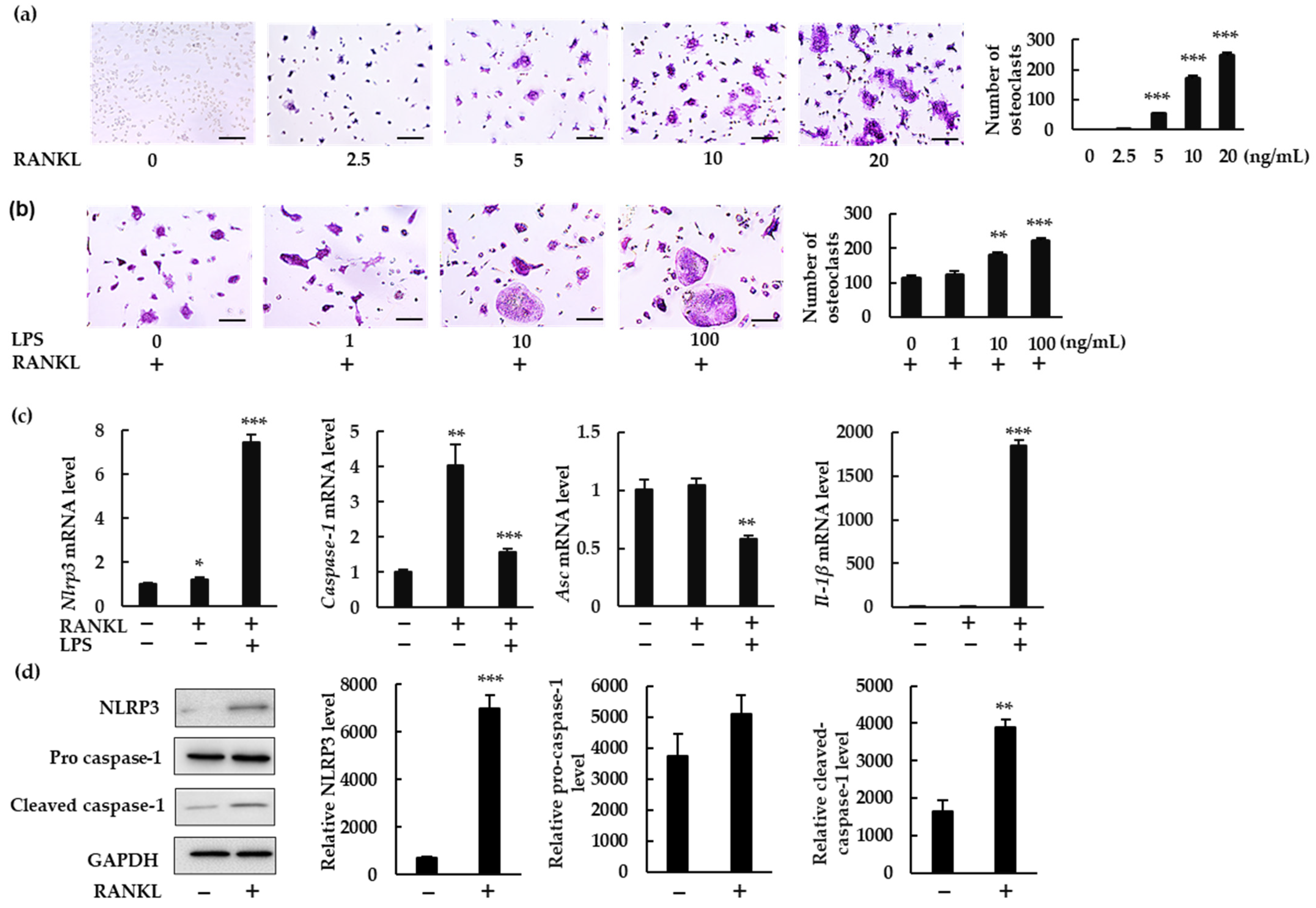
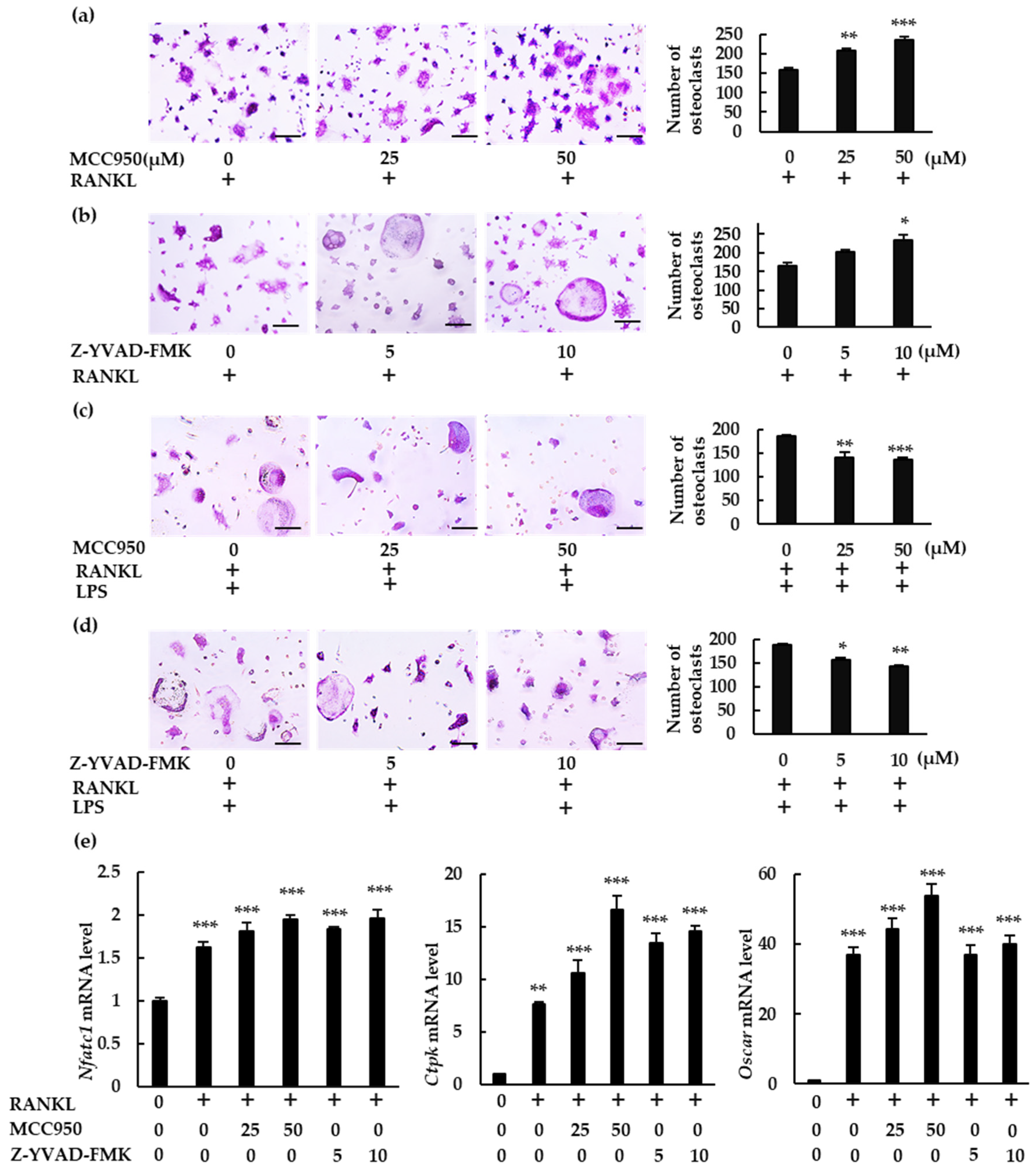
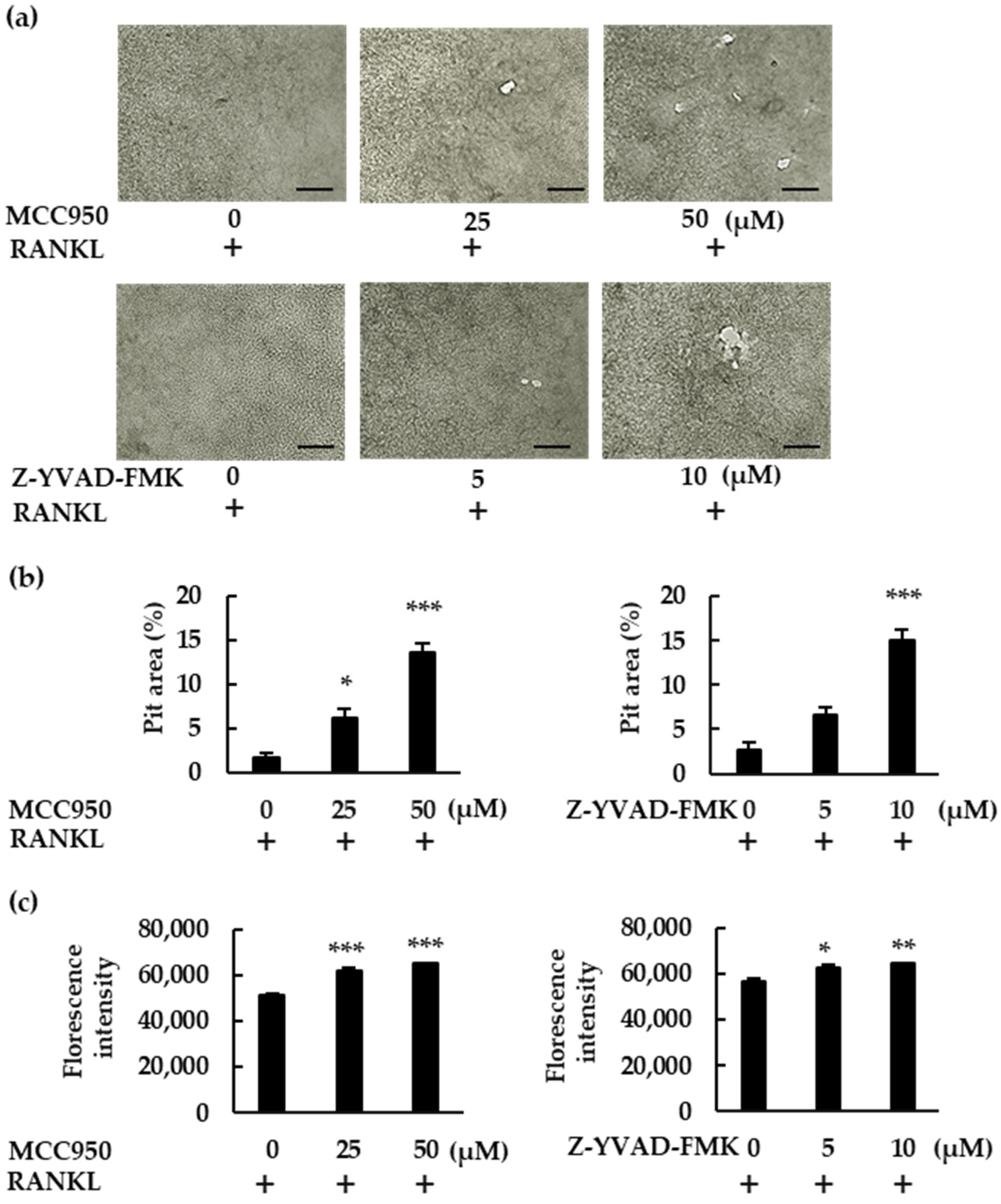
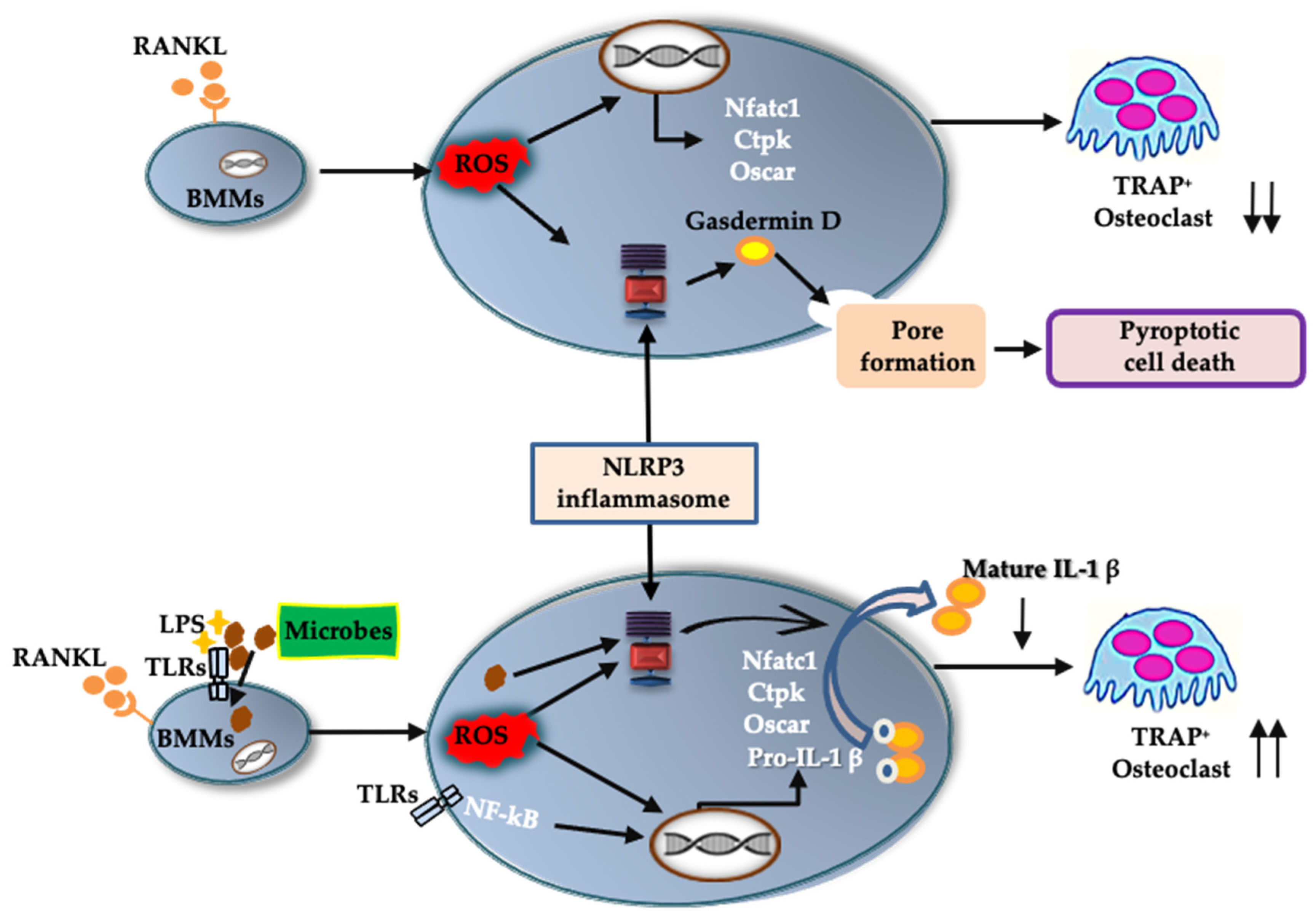
In this study, we demonstrated how the NLRP3 inflammasome is involved in osteoclast formation under various conditions. The abundance of osteoclasts increased in the presence of NLRP3 inflammasome inhibitors when BMMS were treated with RANKL in the absence of LPS. Moreover, these inhibitors generated more pit areas and increased the fluorescence intensity of bone substrates, indicating that the NLRP3 inflammasome negatively regulates osteoclast formation. However, osteoclast formation was suppressed in the presence of NLRP3 inflammasome inhibitors in BMMs treated with RANKL in the presence of LPS, suggesting that NLRP3 accelerates osteoclast formation. These results suggest that the NLRP3 inflammasome regulates osteoclast formation positively under infectious conditions but negatively under physiological conditions.
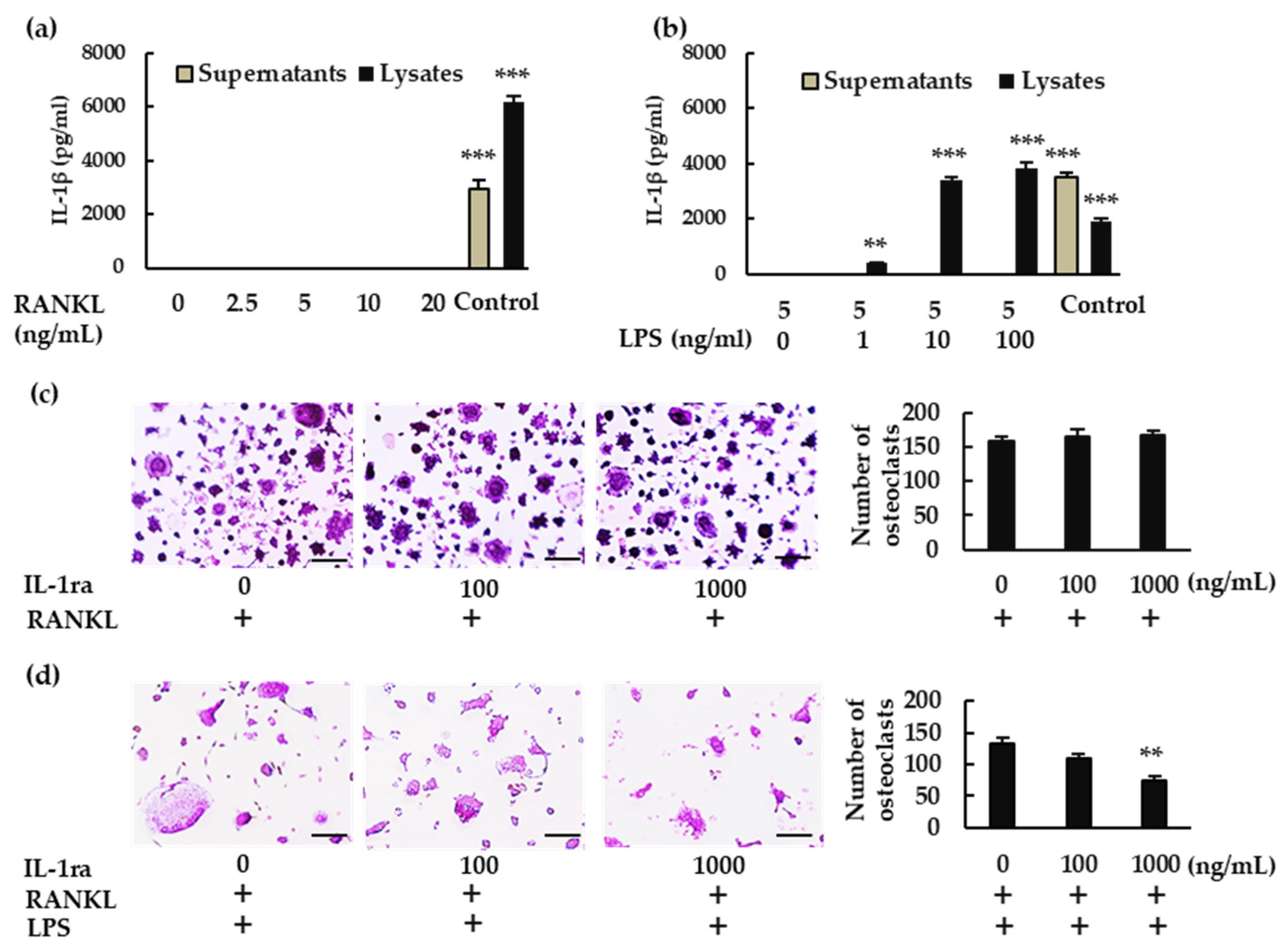
In response to RANKL and LPS treatment, mRNA levels of
Nlrp3,
Caspase-1, and (more importantly)
Il-1β were upregulated. Although we could not detect IL-1β in the culture supernatant of BMMs treated with RANKL and LPS, we did detect it in the cell lysate. IL-1β may be consumed in the culture supernatant of BMMs during differentiation to osteoclasts. Further, rIL-1ra suppressed osteoclast formation from BMMs treated with RANKL and LPS. These results indicate that IL-1β is involved in the osteoclastogenesis induced by RANKL in the presence of LPS. This is consistent with previous studies showing that IL-1β accelerates osteoclast formation in inflammatory diseases such as periodontitis and osteomyelitis [
4,
5]. These findings suggest that the NLRP3 inflammasome contributes positively to osteoclastogenesis under infectious conditions, via IL-1β production (
Figure 8).
In the absence of LPS, RANKL upregulated the expression of
Nlrp3 and
Caspase-1, but not
Il-1β. We detected no IL-1β production in either the BMM culture supernatant or the cell lysate. Further, rIL-1ra did not inhibit the osteoclastogenesis of BMMs treated with RANKL. Combined, these results indicate that IL-1β was not involved in the osteoclastogenesis that was induced by RANKL alone. However, the number of osteoclasts induced by RANKL increased in the presence of NLRP3 inflammasome inhibitors, revealing the negative role played by the NLRP3 inflammasome in osteoclastogenesis. As previously shown, ROS levels increase in BMMs treated with RANKL [
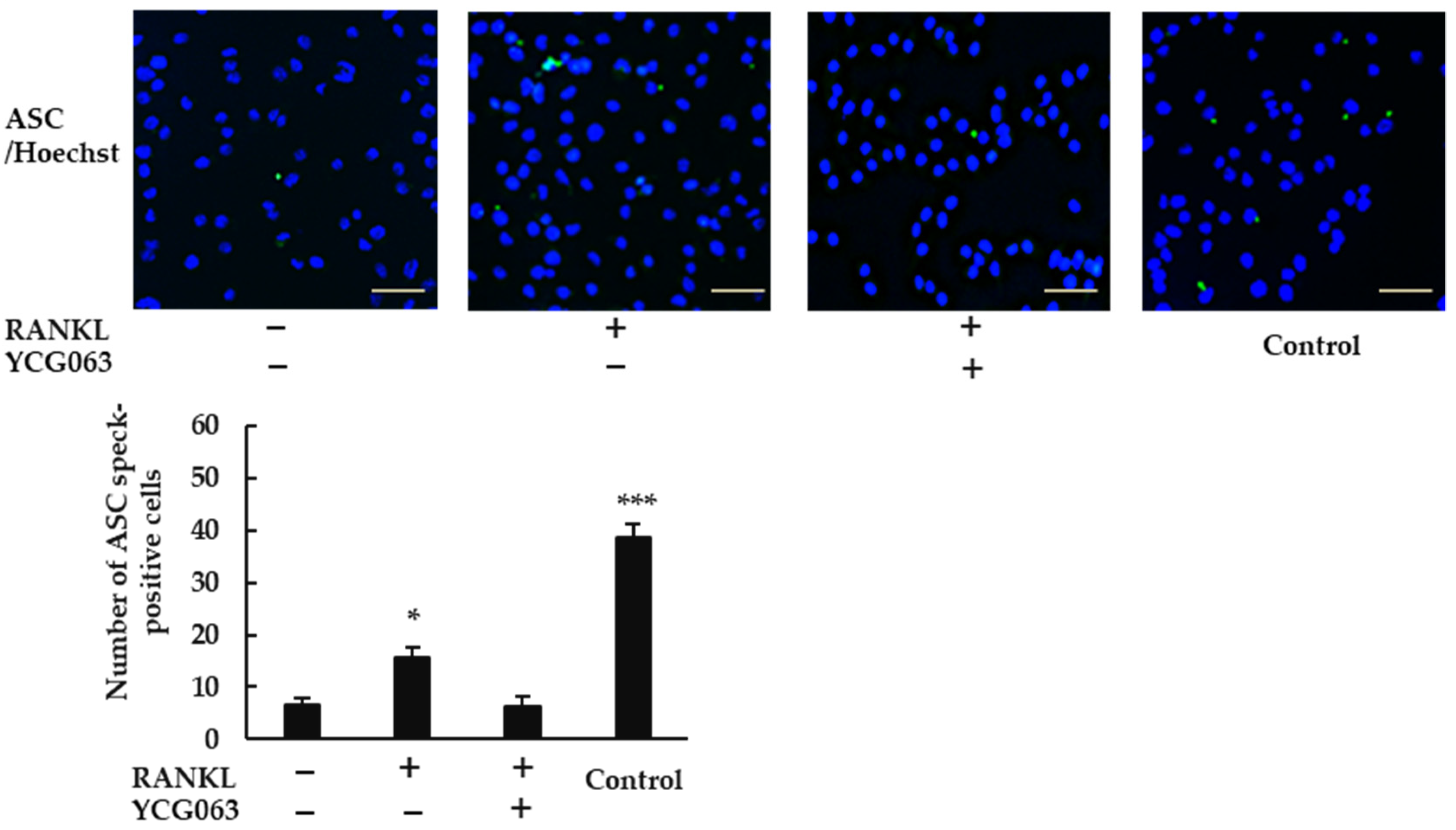
20,
21], and ROS are key activators of the NLRP3 inflammasome [
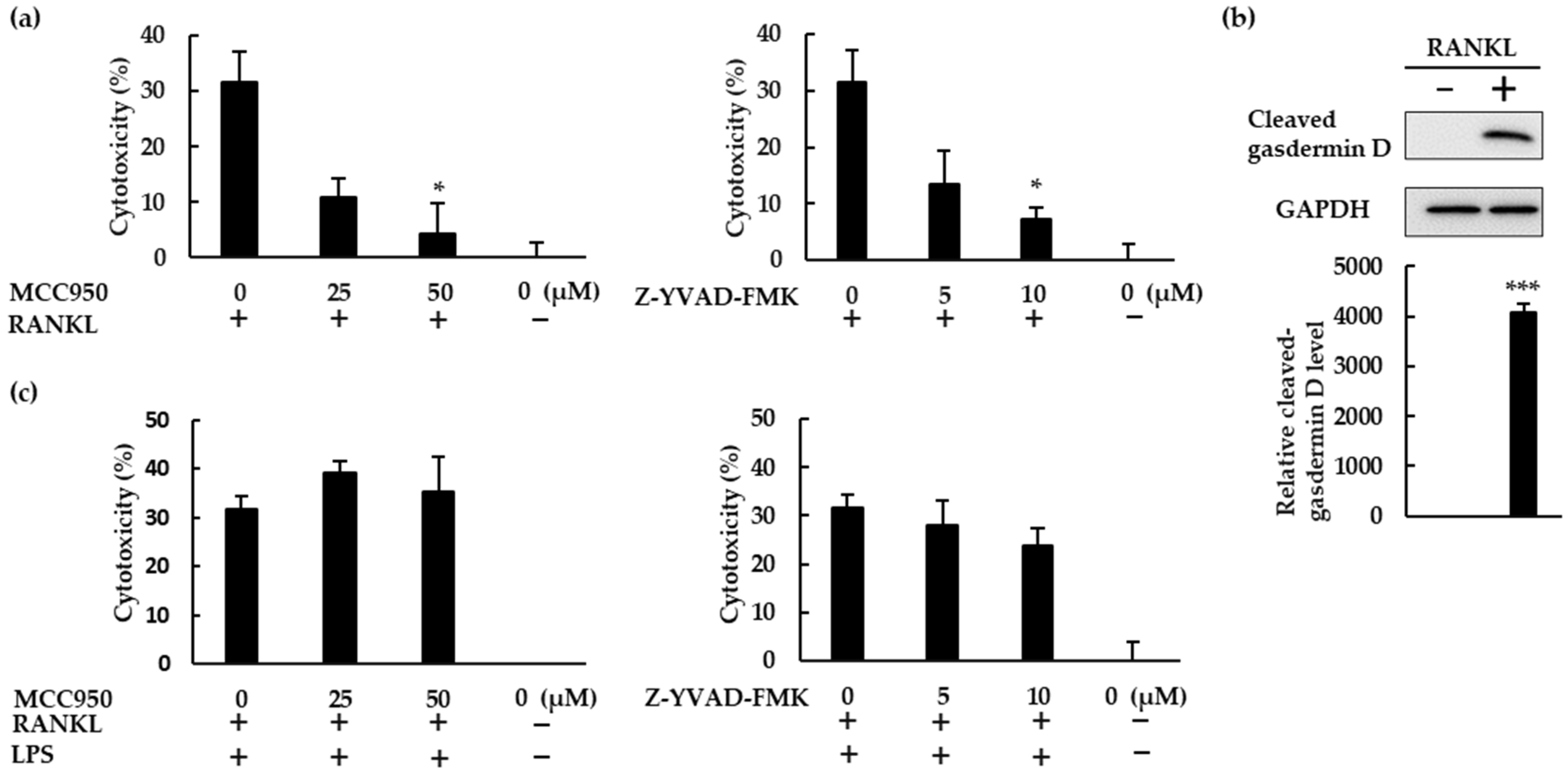
22]. Further, a ROS inhibitor suppressed ASC speck formation in BMMs treated with RANKL. These findings indicate that RANKL activates the NLRP3 inflammasome via ROS production. Assembly of the NLRP3 inflammasome results in the processing of caspase-1, which cleaves gasdermin D to its active form to induce pyroptosis [
23]. Inhibitors of the NLRP3 inflammasome suppressed the cell death of BMMs stimulated with RANKL, indicating that RANKL induces pyroptotic cell death via ROS and the NLRP3 inflammasome in BMMs. Based on these results, RANKL may negatively regulate osteoclastogenesis by inducing pyroptotic cell death (
Figure 8). To the best of our knowledge, this is the first evidence uncovering the mechanism by which the NLRP3 inflammasome regulates RANKL-induced osteoclastogenesis.
This study has demonstrated that the NLRP3 inflammasome can regulate osteoclast formation positively, and possibly also negatively, in BMMs, depending on the environment. Under infectious conditions, the NLRP3 inflammasome disrupts bone homeostasis by promoting osteoclast formation via IL-1β production, possibly to metabolize injured bone tissue and facilitate pathogen elimination around the bone. However, under physiological conditions, the NLRP3 inflammasome suppresses osteoclast formation via pyroptosis. Here, the NLRP3 inflammasome probably plays a crucial role in protecting bone from osteoclast overactivity, and thus helps to maintain bone homeostasis. In addition, this study uncovered the role of the NLRP3 inflammasome in RANKL-mediated physiological osteoclast differentiation. It is still necessary to clarify whether the NLRP3 inflammasome protects against excessive bone loss, using animal models. Future studies will elucidate the exact role of the NLRP3 inflammasome in regulating bone remodeling in infectious, as well as inflammatory, bone diseases such as rheumatoid arthritis and osteoporosis.
4. Materials and Methods
4.1. Reagents
Minimum Essential Media alpha (MEMα) and Dulbecco’s phosphate-buffered saline (D-PBS) were purchased from Fujifilm Wako Pure Chemical (Osaka, Japan). Fetal bovine serum (FBS) was purchased from Hyclone (Logan, UT, USA). Penicillin-streptomycin, anti-rabbit secondary antibody conjugated with Alexa Fluor 488 (A-11008) and CellROX Deep Red Reagent were purchased from Thermo Fisher Scientific (Waltham, MA, USA). ELISA kits for mouse IL-1β (DuoSet), recombinant mouse M-CSF, and recombinant mouse RANKL were purchased from R&D Systems (Minneapolis, MN, USA). Ultra-pure LPS from Escherichia coli O111:B4 was obtained from InvivoGen (San Diego, CA, USA). Erythrocyte lysis buffer BD Pharm Lyse were purchased from BD Biosciences (San Jose, CA, USA). Dimethyl sulfoxide (DMSO) and tartrate-resistant acid phosphatase (TRAP) staining kits were purchased from Sigma–Aldrich (St. Louis, MO, USA). NLRP3 inflammasome inhibitor MCC950 was purchased from Cayman Chemical (Ann Arbor, MI, USA) and caspase-1 inhibitor Z-YVAD-FMK was obtained from Calbiochem-EMD Millipore (Darmstadt, Germany). The Bone Resorption Assay Kit 48 was purchased from Iwai Chemicals Company (Tokyo, Japan). Mouse rIL-1ra was purchased from Biovision (Milpitas, CA, USA). Takara SYBR Premix Ex Taq (Tli RNase H Plus) was obtained from Takara Bio (Otsu, Japan). A rabbit anti-ASC antibody (AL177) was purchased from AdipoGen (San Diego, CA, USA) and CytoTox 96 nonradioactive assay kits were purchased from Promega (Madison, WI, USA). Antibodies for Western blot analysis, anti-NLRP3, anti-gasdermin D, anti-pro caspase-1, anti-caspase-1 rabbit monoclonal antibodies and horseradish peroxidase-conjugated secondary antibodies, were purchased from Cell Signaling Technology (Danvers, MA, USA).
4.2. BMM Isolation
Bone marrow (BM) cells were eluted from the femur and tibiae of six-week-old male BALB/c mice, as previously described [
24]. Briefly, BM was flushed from bones using MEMα, and BM cells were collected via centrifugation, treated with erythrocyte lysis buffer for 10 min at room temperature, and washed with D-PBS. The BM cells were then incubated in MEMα supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin in the presence of M-CSF (5 ng/mL) overnight. Nonadherent BM cells were then collected and incubated in 10 cm dishes for 48 h in MEMα containing M-CSF (30 ng/mL), 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. Cells attached to the dishes after 48 h were collected and used as BMMs. Cells were incubated at 37 °C and in a 5% CO
2 atmosphere.
4.3. In Vitro Osteoclastogenesis
To induce osteoclast differentiation, BMMs (1 × 104 cells/well) were seeded in 96-well plates, incubated with M-CSF (30 ng/mL) and RANKL (0, 2.5, 5, 10, or 20 ng/mL) in MEMα supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin for 72 h. For the inhibition assays, BMMs were incubated with M-CSF (30 ng/mL) and RANKL (10 ng/mL) in the presence or absence of rIL-1ra, MCC950, or Z-YVAD-FMK in the same concentration of M-CSF for 72 h.
Alternatively, BMMs (1 × 104 cells/well) were seeded in 96-well plates, incubated with M-CSF (30 ng/mL) and RANKL (5 ng/mL) in MEMα supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin for 48 h. The cells were then stimulated with ultrapure LPS (0, 1, 10, or 100 ng/mL) in the presence of the same concentrations of RANKL and M-CSF for another 48 h. For the inhibition assays, the cells were incubated with the same concentrations of M-CSF, RANKL, and LPS in the presence or absence of rIL-1ra, MCC950, or Z-YVAD-FMK for the final 48 h.
Cells in 96-well plates were fixed in 4% paraformaldehyde (PFA) for 15 min at 4 °C, treated with 0.2% Triton X-100 in PBS at room temperature for approximately 8–10 min, treated with TRAP staining solution for 10 min at 37 °C, and rinsed three times with distilled water. TRAP-positive cells with three or more nuclei were counted as osteoclasts.
4.4. IL-1β Concentration Measurements
BMMs (5 × 104 cells/well) were seeded in 24-well plates, incubated with M-CSF (30 ng/mL), and RANKL (0, 2.5, 5, 10, or 20 ng/mL) for 7 h. Alternatively, BMMs (5 × 104 cells/well) were seeded in 24-well plates, then incubated with M-CSF (30 ng/mL) and RANKL (5 ng/mL) for 48 h. The cells were then stimulated with LPS (0, 1, 10, or 100 ng/mL) for 7 h in combination with the same concentrations of M-CSF and RANKL. For positive controls, cells were primed with LPS (1 μg/mL) for 3 h, then stimulated with adenosine triphosphate (ATP; 4 mM) for another 45 min. The culture supernatant was collected after centrifugation. The cell lysate was extracted using ice-cold 1% Triton X-100 diluted in D-PBS for 30 min at 4 °C and collected after centrifugation. The concentration of IL-1β was then measured via ELISA, according to the manufacturer’s protocol.
4.5. qRT-PCR Analysis
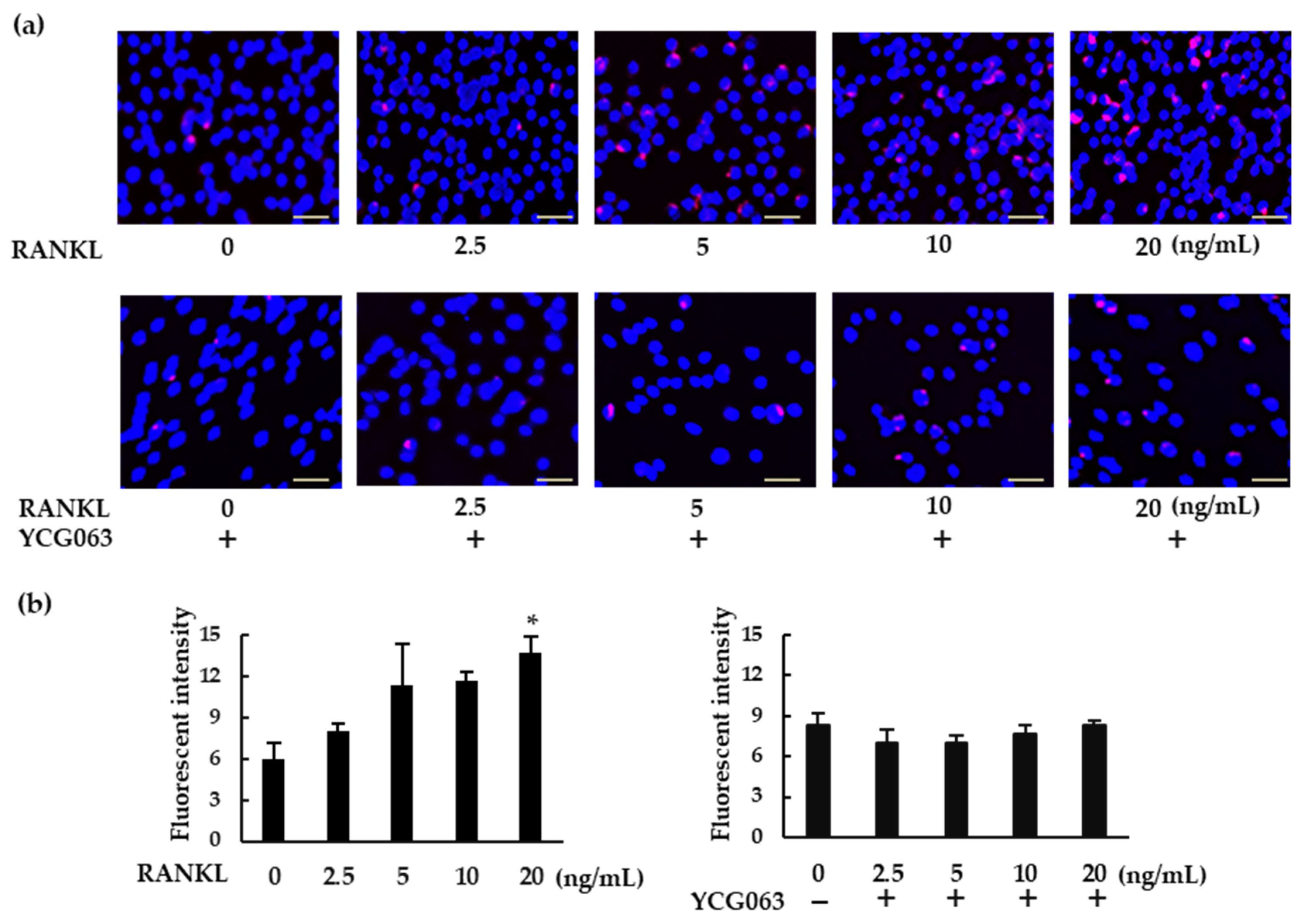
To analyze the mRNA levels of inflammasome-related genes such as Nlrp3, Caspase-1, Asc, and Il-1β, BMMs (1 × 106 cells/dish) were incubated with M-CSF (30 ng/mL) in 60 cm dishes. Following overnight incubation, the cells were treated with or without RANKL (20 ng/mL) in the presence or absence of YCG063 (20 μM) for 7 h. To analyze the mRNA levels of osteoclast-related genes such as Nfatc1, Ctpk and Oscar, BMMs (1 × 106 cells/dish) were incubated with M-CSF (30 ng/mL) and RANKL (20 ng/mL) in the presence or absence of MCC950 or Z-YVAD-FMK for 60 h. Total RNA was extracted from cells using a NucleoSpin RNA purification kit (Takara Bio, Otsu, Japan) with on-column DNase treatment, according to the manufacturer’s instructions. For each sample, 2 μg of total RNA was converted to first-strand cDNA using avian myeloblastosis virus reverse transcriptase (Promega, Madison, WI, USA) at 25 °C for 10 min, followed by 50 min at 42 °C, then 15 min at 70 °C using a Takara PCR thermal cycler (Takara Bio, Otsu, Japan). The cDNA was treated with RNase H and purified using the QIAprep Spin Miniprep Kit (Qiagen, Hilden, Germany). Primer sequences used were designed using the Primer3 Input (v0.4.0) software and obtained from Hokkaido System Science (Sapporo, Japan). Primer sequences were as follows: Nlrp3 forward, 5′-ATGCTGCTTCGACATCTCCT-3′ and reverse, 5′-AACCAATGCGAGATCCTGAC-3′; Caspase-1 forward, 5′-CACAGCTCTGGAGATGGTGA-3′ and reverse, 5′-TCTTTCAAGCTTGGGCACTT-3′; Asc forward, 5′-TCACAGAAGTGGACGGAGTG-3′ and reverse, 5′-CTCCAGGTCCATCACCAAGT-3′; Il-1β forward, 5′-AACCTGCTGGTGTGTGACGTTC-3′ and reverse, 5′-CAGCACGAGGCTTTTTTGTTGT-3′; Nfatc1 forward, 5′-GGGTCAGTGTGACCGAAGAT-3′ and reverse, 5′-GGAAGTCAGAAGTGGGTGGA-3′; Ctpk forward, 5′-CCAGTGGGAGCTATGGAAGA-3′ and reverse, 5′-AAGTGGTTCATGGCCAGTTC-3′; Oscar forward, 5′-TGCTGTGCCAATCACAAGTA-3′ and reverse, 5′-AGGGAAACCTCATCCGTTT-3′; GAPDH forward, 5′-GGAGGAACCTGCCAAGTATG-3′, and reverse, 5′-TGGGAGTTGCTGTTGAAGTC-3′. Comparative quantification for Nlrp3, Caspase-1, Asc, and Il-1β was completed with SYBR Premix Ex Taq using the Mx3000 P qPCR System (Agilent Technologies, Santa Clara, CA, USA). Amplification conditions were as follows: 95 °C for 10 s, followed by 40 cycles of 95 °C for 5 s, 54 °C (Oscar), 56 °C (Nlrp3 and Caspase-1) or 58 °C (Il-1β, Asc, Nfatc1 and Ctpk) for 20 s, and a final cycle of 95 °C for 1 min, 55 °C for 30 s, and 95 °C for 30 s. A melting curve analysis was used to confirm that the proper PCR products had been amplified in all samples. The relative expression ratio of Nlrp3, Caspase-1, Asc, and Il-1β mRNA was calculated based on PCR efficiency and threshold cycle differences between test samples (stimulated cells) and a calibrator (unstimulated cells). Target gene expression was normalized using GAPDH gene expression. Calibrator mRNA levels were set to 1.
4.6. ASC Speck Formation
BMMs (5 × 104 cells/well) were incubated with M-CSF (30 ng/mL) in 96-well plates overnight, and then treated with RANKL (0 or 20 ng/mL) in the presence or absence of YCG063 (20 μM) for 24 h. For positive controls, cells were primed with LPS (1 μg/mL) for 3 h and further stimulated with ATP (5 mM) for 90 min, then fixed with 4% PFA in PBS for 30 min at 4 °C and permeabilized with 0.2% Triton X-100 in PBS at room temperature for 10 min. A blocking solution containing 0.2% gelatin was applied for 60 min at room temperature. The cells were then incubated with a rabbit anti-ASC antibody (1:250 dilution) overnight at 4 °C. After the cells had been washed with blocking buffer thrice, goat anti-rabbit secondary antibody conjugated with Alexa Fluor 488 (1:500 dilution) was applied for 60 min. The cells were then counterstained with Hoechst 33342 for 30 min. Finally, ASC speck formation was analyzed via fluorescent microscopy (BZ-X800, All-in-One Fluorescence Microscope, Keyence, Osaka, Japan).
4.7. Cytotoxicity Assay
BMMs (1 × 10
4 cells/well) were seeded in 96-well plates and incubated with M-CSF (30 ng/mL), RANKL (0 or 10 ng/mL), and 4% FBS in the presence or absence of MCC950 or Z-YVAD-FMK for 72 h. Alternatively, BMMs (1 × 10
4 cells/well) were seeded in 96-well plates and incubated with M-CSF (30 ng/mL), RANKL (0 or 5 ng/mL), and 10% FBS for 48 h. The cells were then stimulated with LPS (10 ng/mL) with 1% FBS and the same concentrations of M-CSF and RANKL, in the presence or absence of MCC950 or Z-YVAD-FMK, for 48 h. As a negative control, cells were treated with 30 ng/mL M-CSF alone. As a positive control, cells were frozen (−80 °C) and thawed for two cycles. After stimulation, the plates were centrifuged and the cell culture supernatant was harvested. Lactate dehydrogenase (LDH) in the supernatant was then measured using a CytoTox 96 nonradioactive assay, according to the manufacturer’s protocol. Cytotoxicity percentage was determined based on the following calculation [
25]:
4.8. Bone-Resorption Assay
BMMs (5 × 10
4 cells/well) were seeded in 48-well plates and cultured in MEMα (without phenol red) supplemented with 10% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin containing M-CSF (30 ng/mL), and RANKL (20 ng/mL), in the presence or absence of MCC950 and Z-YVAD-FMK, for 4 d. The medium was then replaced with fresh medium containing the same concentrations of these reagents and incubated for another 6 d. After incubation, a bone-resorption pit assay was performed, according to the manufacturer’s instructions. Briefly, culture supernatant was transferred to a black plate. Bone-resorption assay buffer was added and fluorescence was measured using a fluorescent plate reader (BMG FLUOstar OPTIMA, BMG LABTECH, Ortenberg, Germany). After the supernatant had been removed, the calcium-coated plates were treated with 5% sodium hypochlorite to remove the cells. Images of the pit areas were then captured using a microscope and analyzed using ImageJ software (v1.52a;
http://imagej.nih.gov/ij/, accessed on 28 March 2022) [
24].
4.9. ROS Detection Assay
To detect intracellular ROS production, BMMs (5 × 104 cells/well) were seeded in 96-well plates and cultured overnight with M-CSF (30 ng/mL). The cells were then treated with RANKL (0, 2.5, 5, 10, or 20 ng/mL) in the presence of CellROX Deep Red Reagent, at a final concentration of 5 μM, for 30 min. For inhibition assays, BMMs were pretreated with YCG063 (20 μM) for 1 h. The cells were then counterstained with Hoechst 33342 for 15 min, washed with PBS, and the fluorescence was measured using a fluorescent plate reader (BMG FLUOstar OPTIMA). ROS-expressing cells were also analyzed via fluorescence microscopy (BZ-X800, All-in-One Fluorescence Microscope).
4.10. Western Blot Analysis
BMMs (5 × 10
6 cells/dish) were treated with RANKL (0, 20 ng/mL) and M-CSF (30 ng/mL) for 24 h and subjected to Western blot analysis as previously described [
26]. Briefly, cells were rinsed twice with ice-cold PBS, and lysed in a cell lysis buffer (50 mM Tris-HCl (pH 8.0), 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), 150 mM NaCl, 1 mM PMSF, and proteinase inhibitor cocktail). An equal amount of protein (5 µg) was applied to each lane of SDS-polyacrylamide gel electrophoresis, followed by transfer onto a polyvinylidene difluoride membrane. The blots were blocked with 3% skim milk in Tris-buffered saline for 1 h at 25 °C. Blots were then incubated with anti-NLRP3, anti-pro Caspase-1, anti-cleaved caspase-1, anti-gasdermin D, or anti-GAPDH primary antibodies overnight at 4 °C, washed, incubated with horseradish peroxidase-conjugated secondary antibodies, and finally detected with Immobilon Forte Western HRP substrate (Merck, Darmstadt, Germany). The immunoreactive bands were analyzed using a LAS-4000mini (Fujifilm, Tokyo, Japan) and relative level of proteins were determined using ImageJ software.
4.11. Statistical Analysis
Statistical differences among groups were assessed using a one-factor analysis of variance (ANOVA) with Tukey–Kramer tests. Differences between two groups were analyzed using t-tests. All data were analyzed using Stat Mate V (ATMS, Tokyo, Japan).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







