
Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts

Abstract
:1. Introduction
2. Results
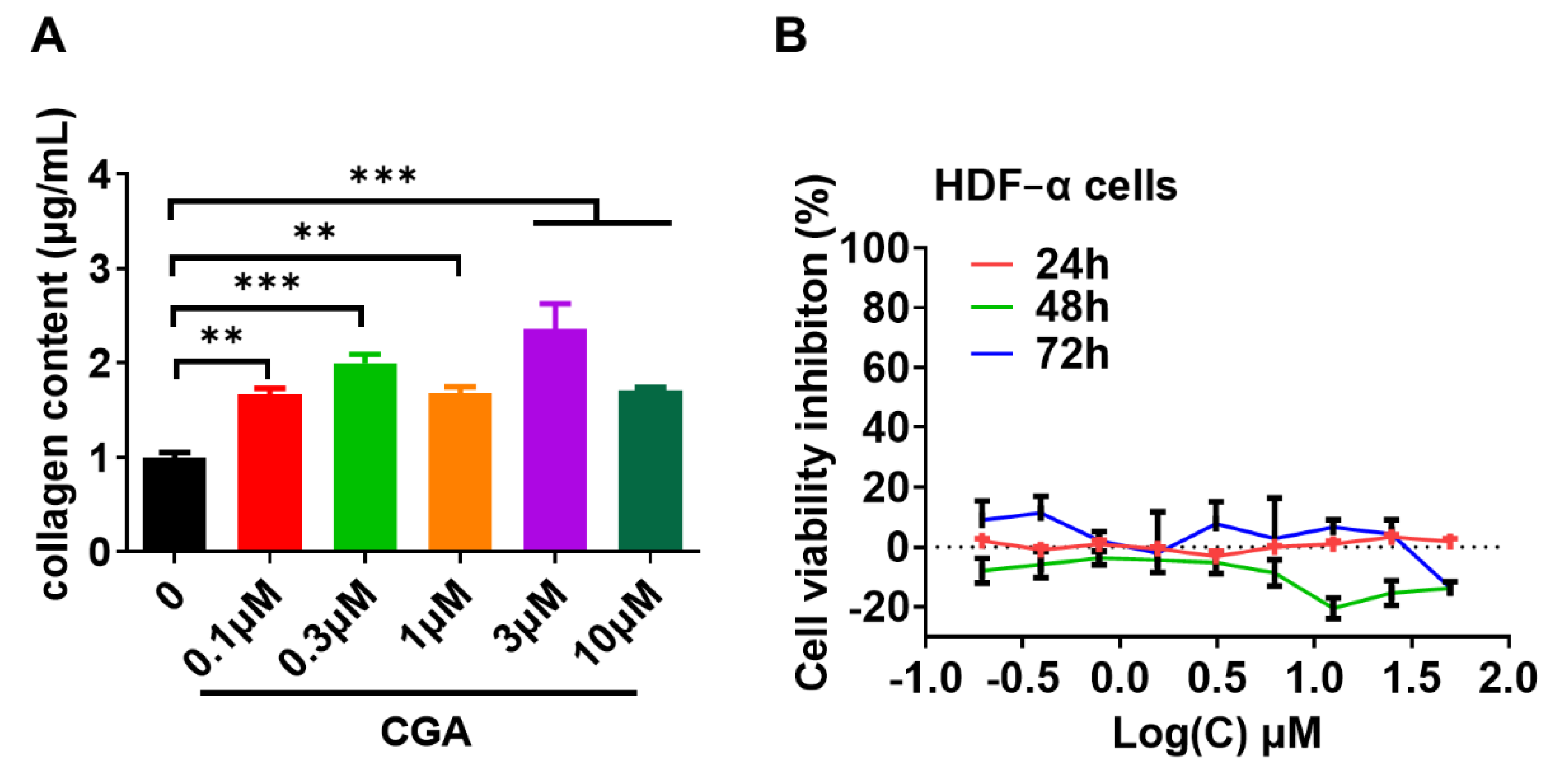
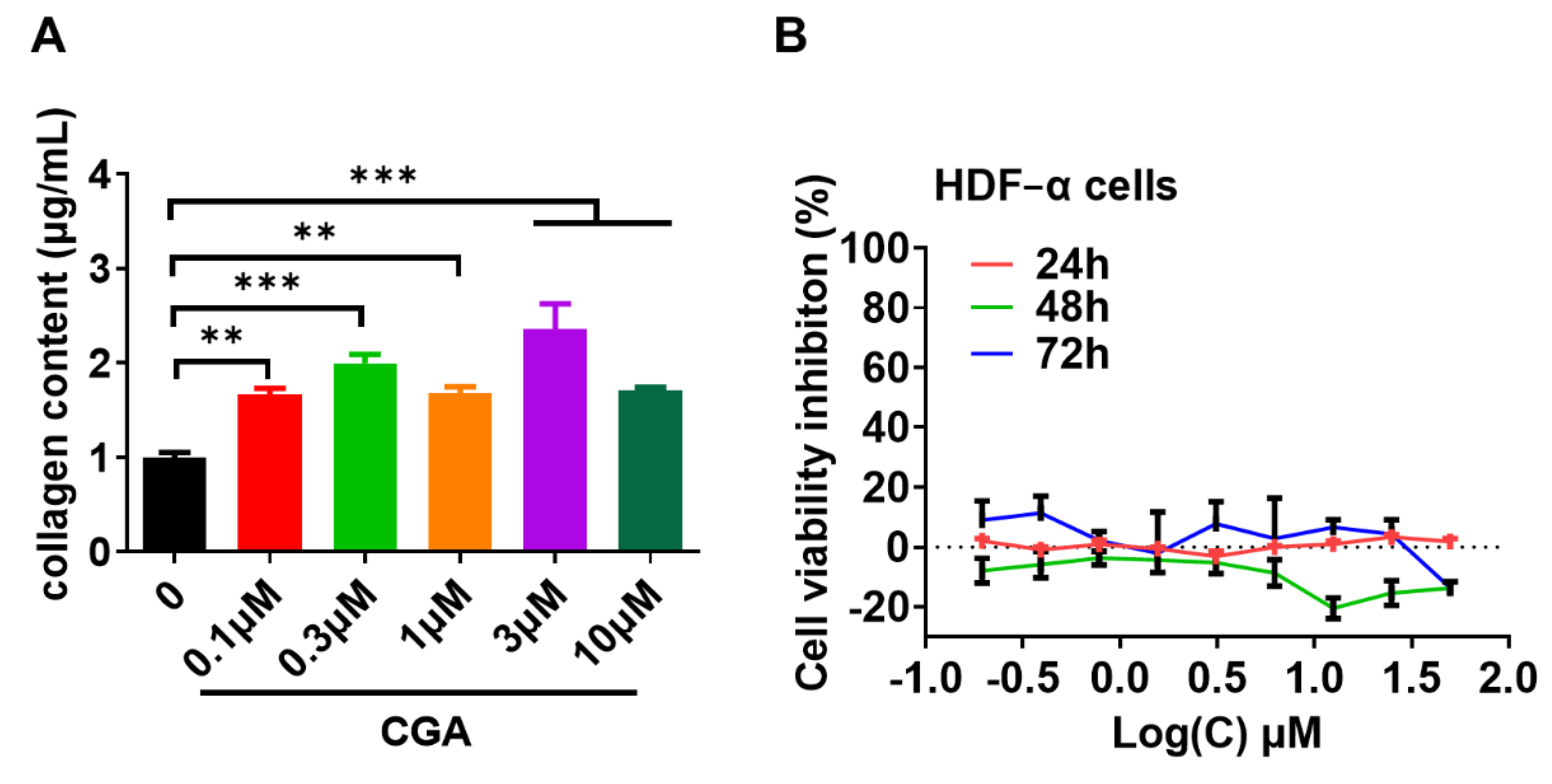
2.1. Effect of CGA on Collagen Expression in Human Skin Fibroblasts
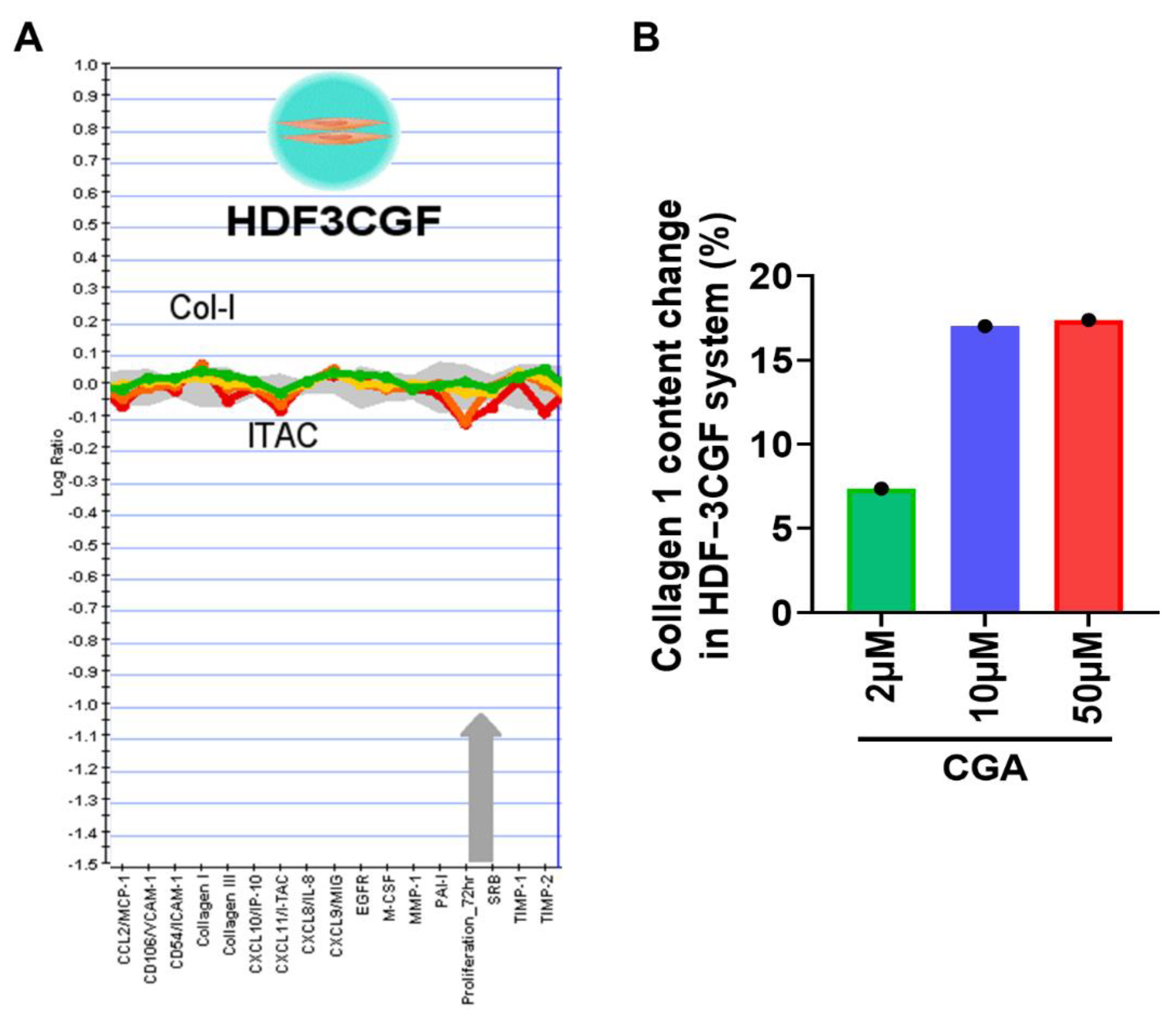
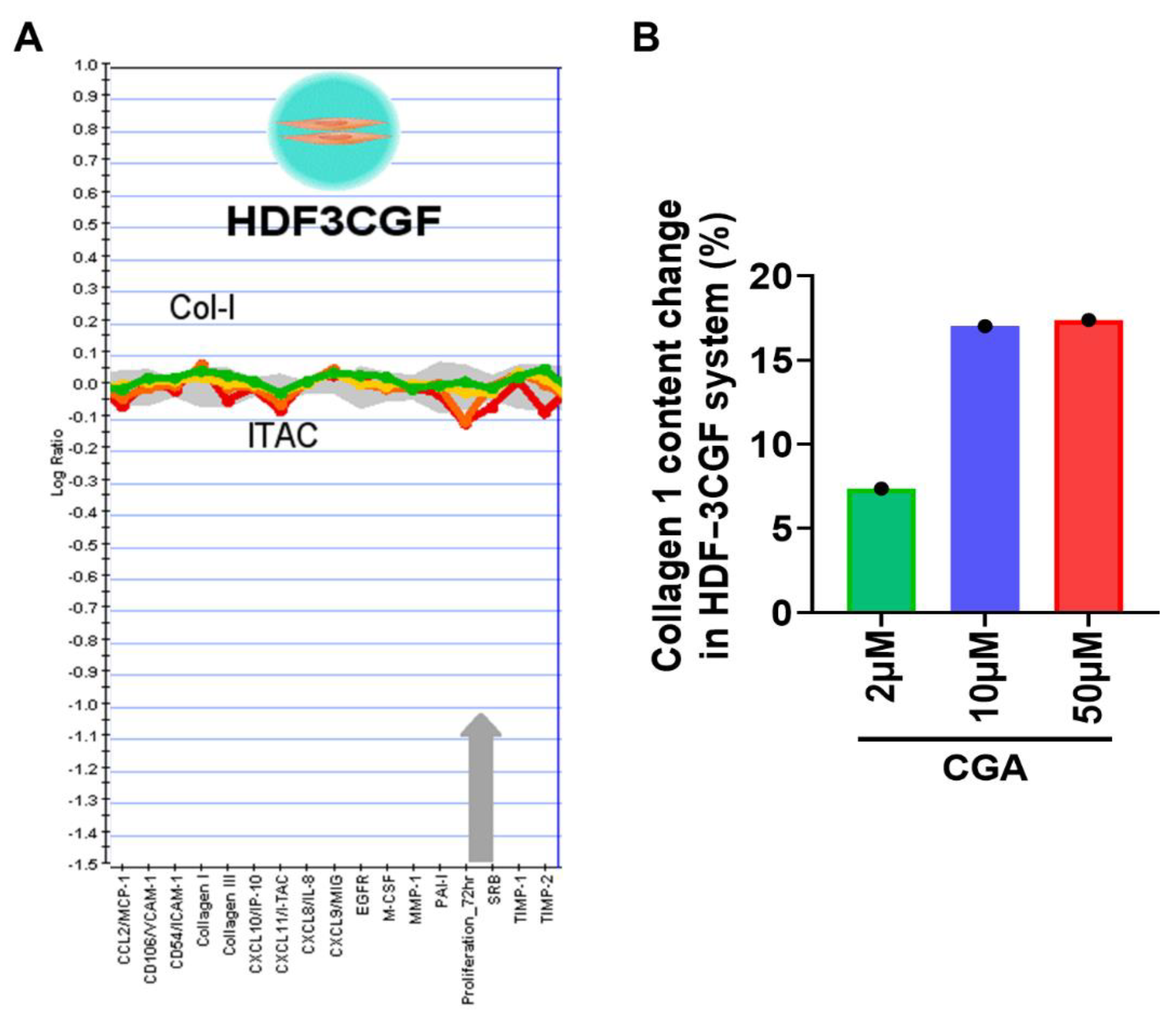
2.2. Effect of CGA on Collagen Secretion in HDFs
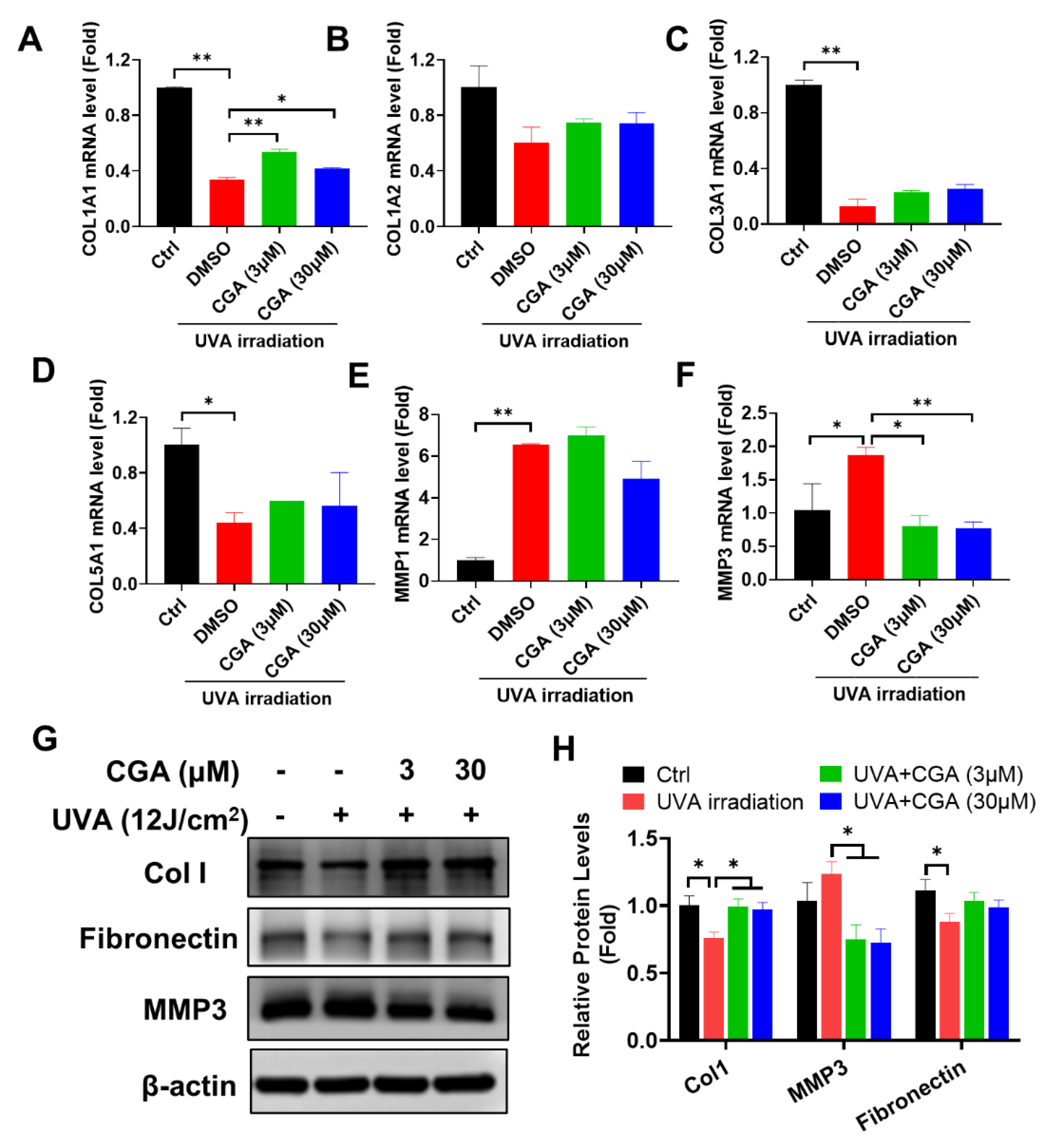
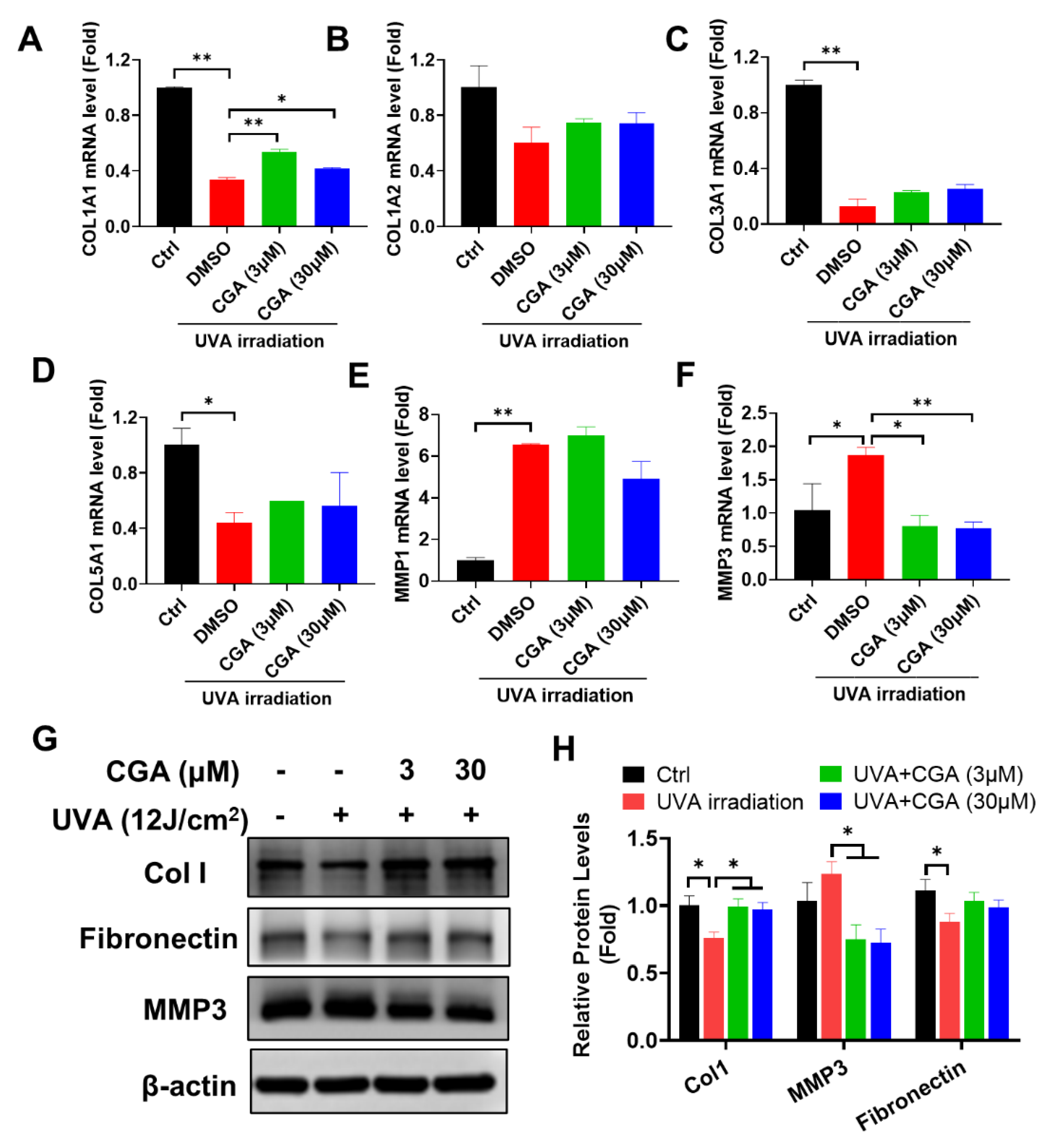
2.3. Effect of CGA on Collagen Metabolism in UVA-Irradiated HDFs
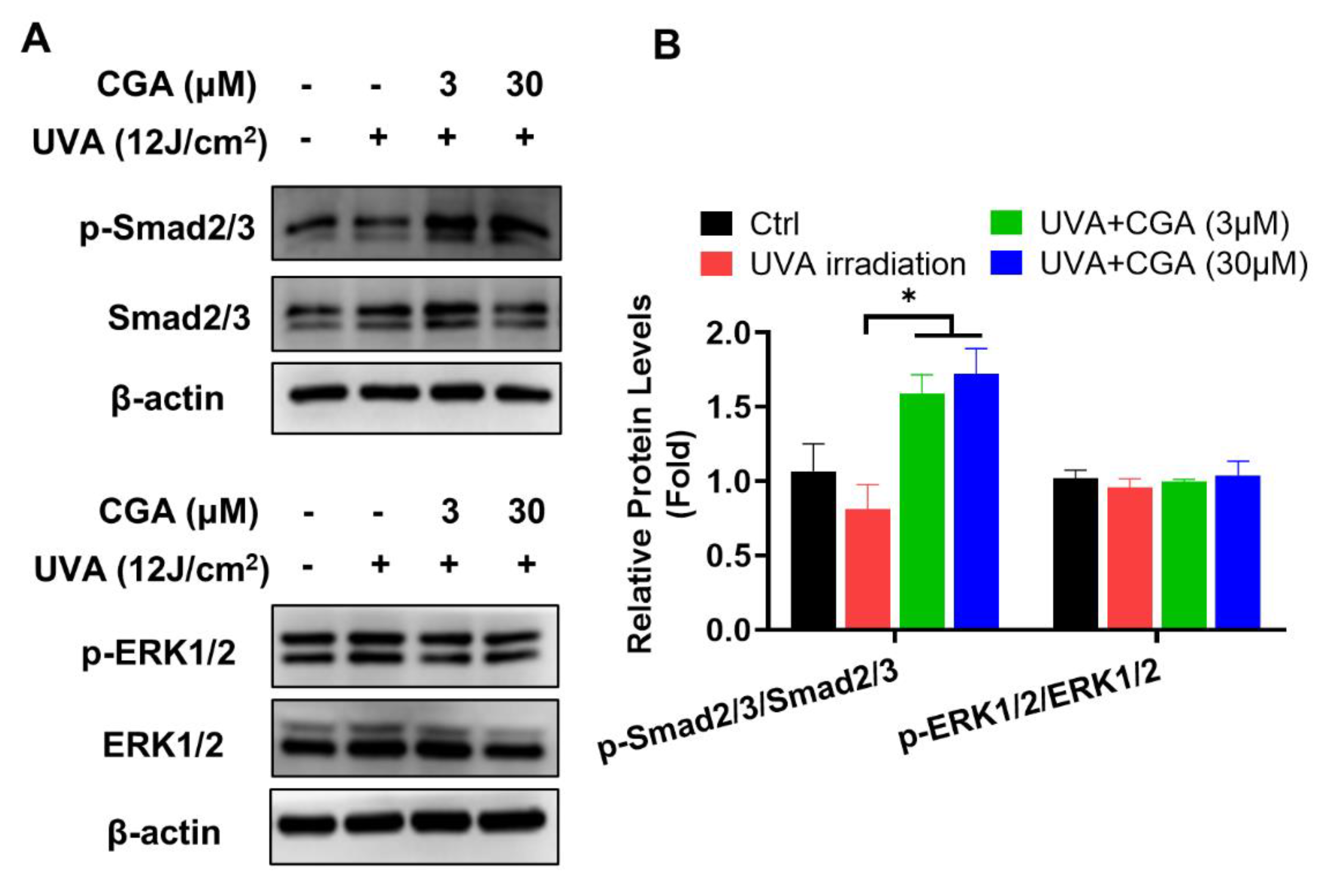
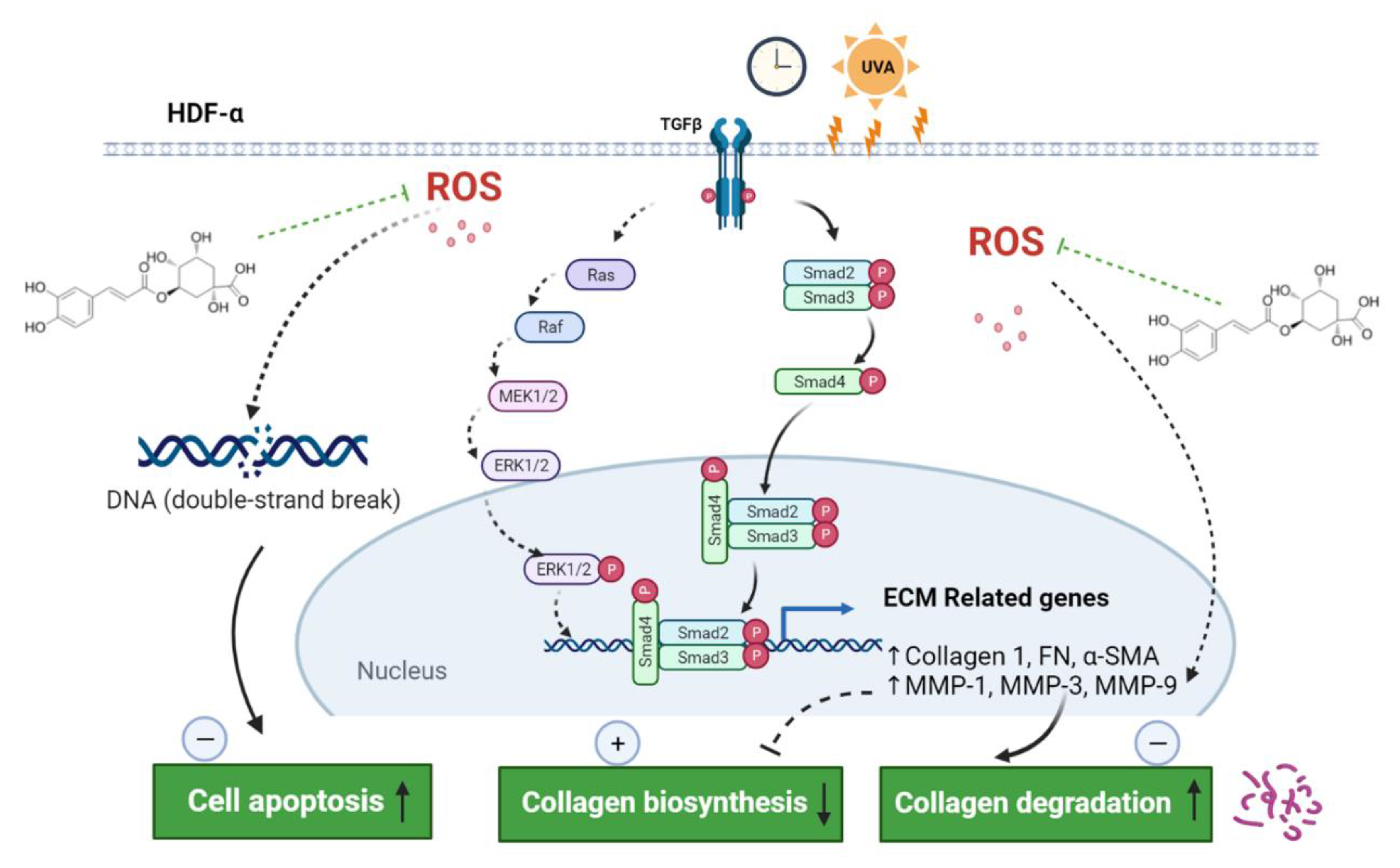
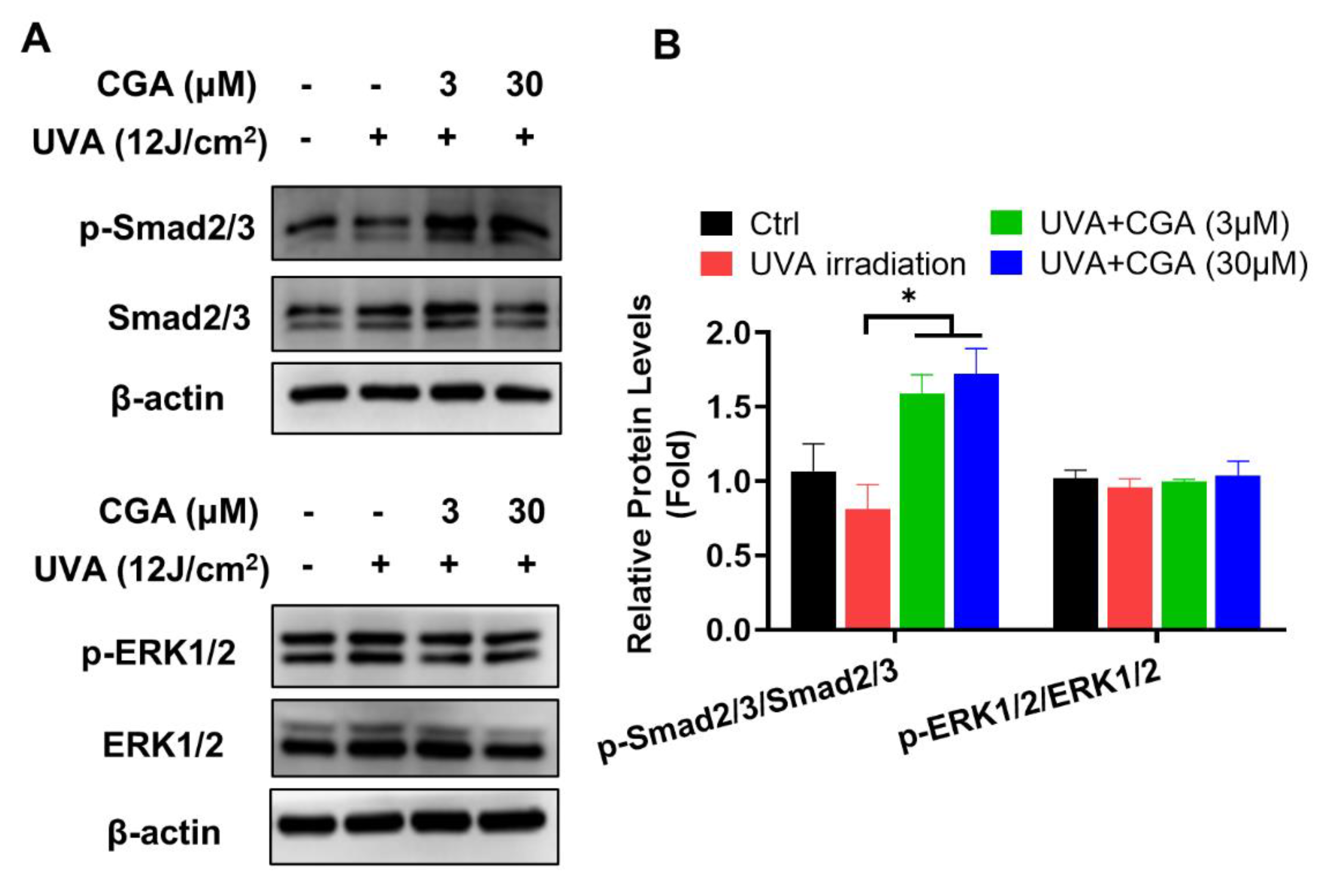
2.4. Effect of CGA on TGF-β/Smad Signaling in UVA-Irradiated HDFs
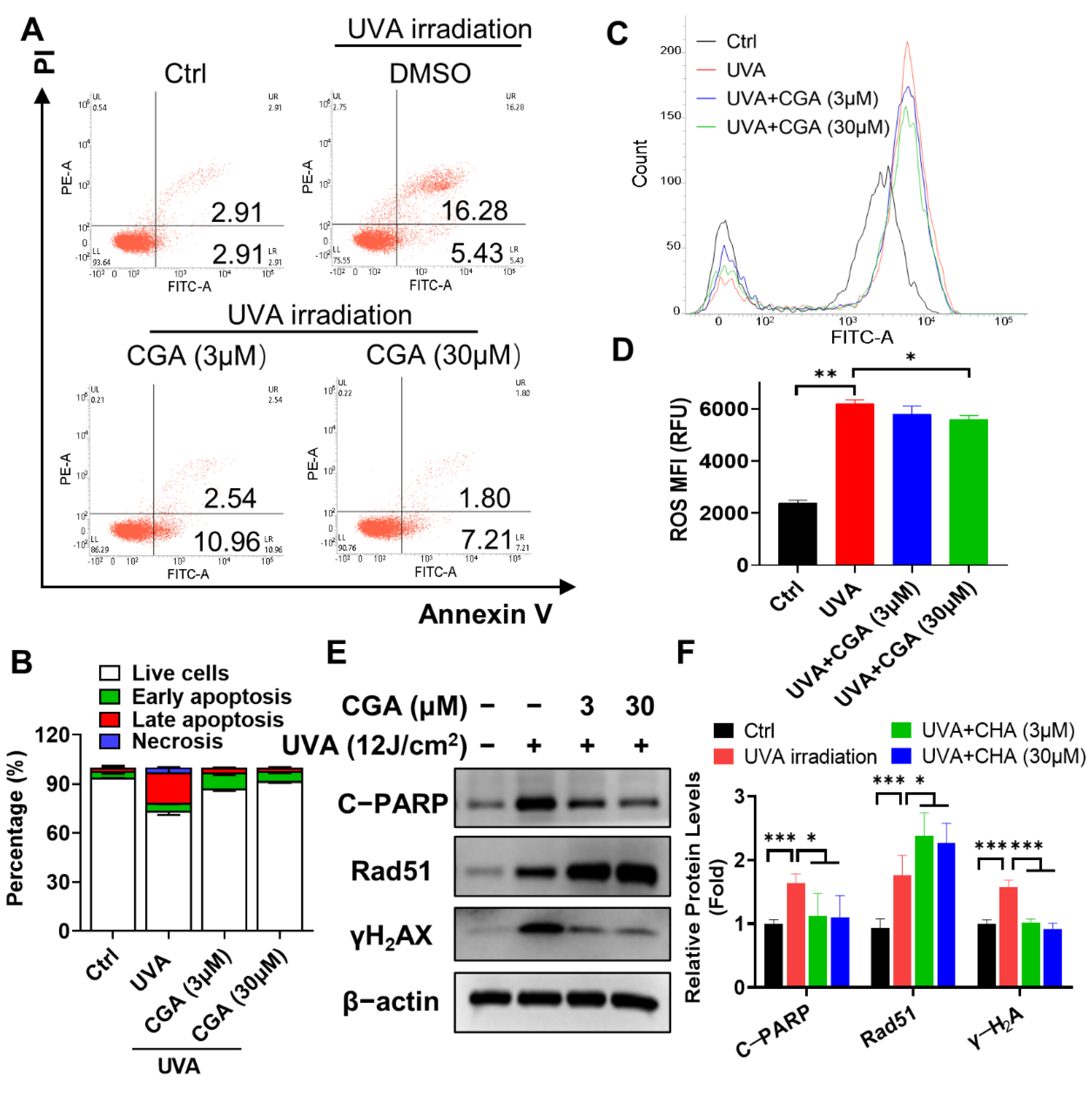
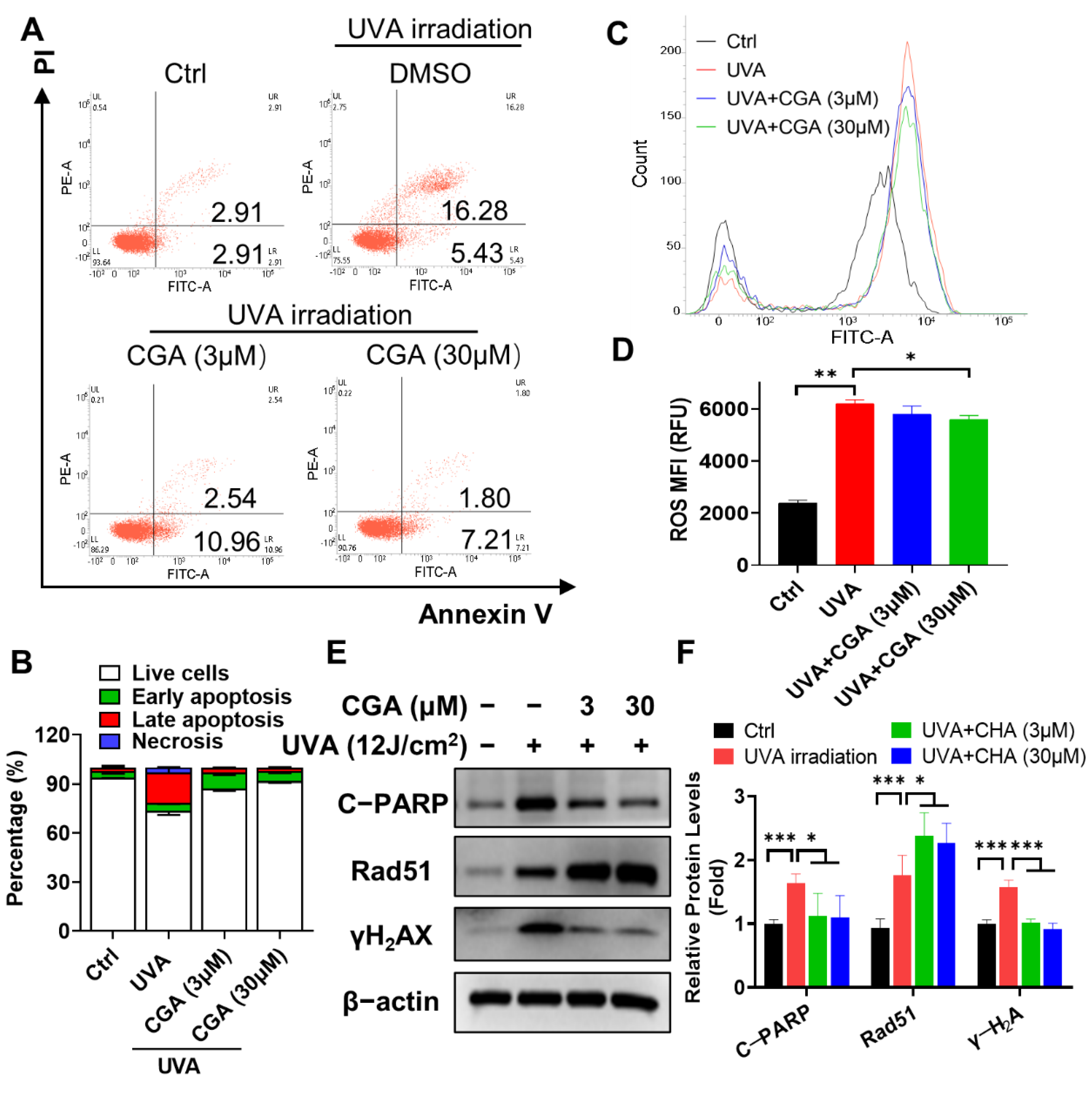
2.5. CGA Protects against the UVA-Induced Apoptosis in HDFs
2.6. CGA Inhibits the UVA-Induced ROS Production in HDFs
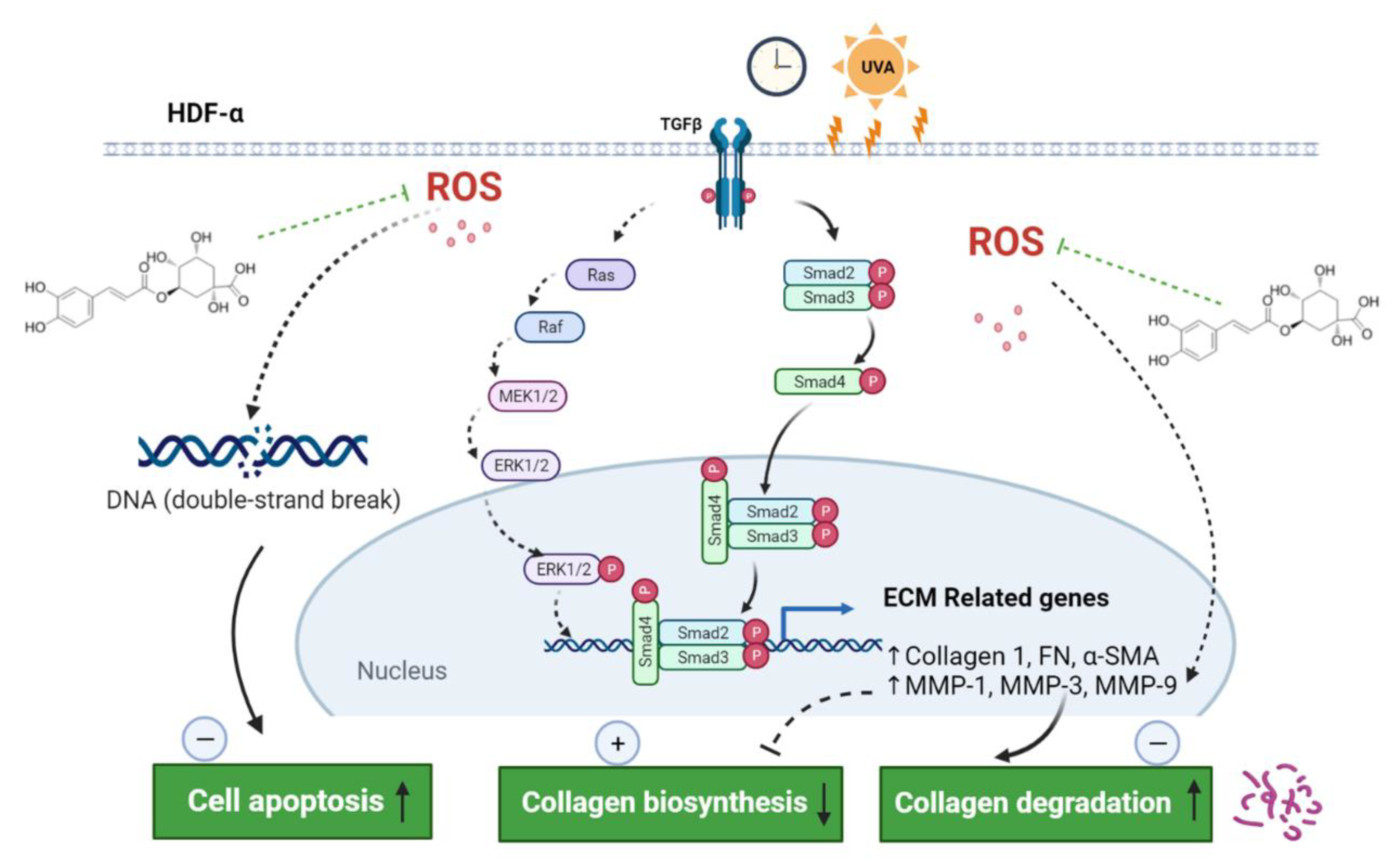
3. Discussion
4. Materials and Methods
4.1. Cells Culture
4.2. UVA Irradiation
4.3. Antibodies and Chemicals
4.4. Cell Viability Assay
4.5. Quantitative RT-PCR (qRT-PCR) Assay
4.6. Western Blot Assays
4.7. Soluble Collagen Detection
4.8. BioMAP Profiling Analysis
4.9. Apoptosis Assay
4.10. Cellular ROS Detection Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CGA | Chlorogenic acid |
| HDFs | Human dermal fibroblasts |
| Col | Collagen |
| DSBs | Double strand breaks |
| MMPs | Matrix metalloproteinases |
| ECM | Extracellular matrix |
| UVA | Ultraviolet A |
| ROS | Reactive oxygen species |
| TGF-β | Transforming growth factor-β |
References
- Rose, M.R.; Flatt, T.; Graves, J.L.; Greer, L.F.; Martinez, D.E.; Matos, M.; Mueller, L.D.; Reis, R.J.S.; Shahrestani, P. What is Aging? Front. Genet. 2012, 3, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases—From induction via signaling to initial events. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T.; Osawa, M.; Nishikawa, S.I. Steel factor controls the proliferation and differentiation of neonatal mouse epidermal melanocytes in culture. Pigment. Cell Res. 2003, 16, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.T.; Holbrook, K.A.; Madri, J.A. Collagen types I, III, and V in human embryonic and fetal skin. Am. J. Anat. 1986, 175, 507–521. [Google Scholar] [CrossRef]
- Rittie, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Verma, R.P.; Hansch, C. Matrix metalloproteinases (MMPs): Chemical-biological functions and (Q)SARs. Bioorganic Med. Chem. 2007, 15, 2223–2268. [Google Scholar] [CrossRef]
- Erman, H.; Gelisgen, R.; Cengiz, M.; Tabak, O.; Erdenen, F.; Uzun, H. The association of vascular endothelial growth factor, metalloproteinases and their tissue inhibitors with cardiovascular risk factors in the metabolic syndrome. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1015–1022. [Google Scholar]
- Kohl, E.; Steinbauer, J.; Landthaler, M.; Szeimies, R.M. Skin ageing. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 873–884. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Zaid, A.N.; Al Ramahi, R. Depigmentation and Anti-aging Treatment by Natural Molecules. Curr. Pharm. Des. 2019, 25, 2292–2312. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N. Chlorogenic acids and other cinnamates—Nature, occurrence and dietary burden. J. Sci. Food Agr. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Xu, Y.X.; Chen, J.W.; Yu, X.A.; Tao, W.W.; Jiang, F.R.; Yin, Z.M.; Liu, C. Protective effects of chlorogenic acid on acute hepatotoxicity induced by lipopolysaccharide in mice. Inflamm. Res. 2010, 59, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; He, J.; Ren, X.; Cai, W.S.; Fang, Y.C.; Feng, X.Z. Development of functional biointerfaces by surface modification of polydimethylsiloxane with bioactive chlorogenic acid. Colloid Surf. B 2014, 116, 700–706. [Google Scholar] [CrossRef]
- Xue, N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.; Chen, J.; Zhang, M.; Jia, J.; Yang, H.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef] [Green Version]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; Xia, F.F.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Li, H.R.; Habasi, M.; Xie, L.Z.; Aisa, H.A. Effect of Chlorogenic Acid on Melanogenesis of B16 Melanoma Cells. Molecules 2014, 19, 12940–12948. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.W.; Piao, M.J.; Kim, K.C.; Yao, C.W.; Zheng, J.; Kim, S.M.; Hyun, C.L.; Ahn, Y.S.; Hyun, J.W. The Polyphenol Chlorogenic Acid Attenuates UVB-mediated Oxidative Stress in Human HaCaT Keratinocytes. Biomol. Ther. 2014, 22, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Her, Y.; Lee, T.K.; Kim, J.D.; Kim, B.; Sim, H.; Lee, J.C.; Ahn, J.H.; Park, J.H.; Lee, J.W.; Hong, J.; et al. Topical Application of Aronia melanocarpa Extract Rich in Chlorogenic Acid and Rutin Reduces UVB-Induced Skin Damage via Attenuating Collagen Disruption in Mice. Molecules 2020, 25, 4577. [Google Scholar] [CrossRef]
- Girsang, E.; Ginting, C.N.; Lister, I.N.E.; Gunawan, K.Y.; Widowati, W. Anti-inflammatory and antiaging properties of chlorogenic acid on UV-induced fibroblast cell. Peer J. 2021, 9, e11419. [Google Scholar] [CrossRef]
- Chung, J.H.; Seo, J.Y.; Lee, M.K.; Eun, H.C.; Lee, J.H.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Ultraviolet modulation of human macrophage metalloelastase in human skin In Vivo. J. Investig. Dermatol. 2002, 119, 507–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossetti, D.; Kielmanowicz, M.G.; Vigodman, S.; Hu, Y.P.; Chen, N.; Nkengne, A.; Oddos, T.; Fischer, D.; Seiberg, M.; Lin, C.B. A novel anti-ageing mechanism for retinol: Induction of dermal elastin synthesis and elastin fibre formation. Int. J. Cosmet. Sci. 2011, 33, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Purohit, T.; He, T.; Qin, Z.; Li, T.; Fisher, G.J.; Yan, Y.; Voorhees, J.J.; Quan, T. Smad3-dependent regulation of type I collagen in human dermal fibroblasts: Impact on human skin connective tissue aging. J. Dermatol. Sci. 2016, 83, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Bhogal, R.K.; Bona, C.A. Regulatory effect of extracellular signal-regulated kinases (ERK) on type I collagen synthesis in human dermal fibroblasts stimulated by IL-4 and IL-13. Int. Rev. Immunol. 2008, 27, 472–496. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, E.R. Role of UV light in photodamage, skin aging, and skin cancer: Importance of photoprotection. Am. J. Clin. Dermatol. 2009, 10 (Suppl. 1), 19–24. [Google Scholar] [CrossRef]
- Landau, M. Exogenous factors in skin aging. Curr. Probl. Dermatol. 2007, 35, 1–13. [Google Scholar]
- Inagaki, Y.; Truter, S.; Ramirez, F. Transforming growth factor-beta stimulates alpha 2(I) collagen gene expression through a cis-acting element that contains an Sp1-binding site. J. Biol. Chem. 1994, 269, 14828–14834. [Google Scholar] [CrossRef]
- Choi, S.I.; Jung, T.D.; Cho, B.Y.; Choi, S.H.; Sim, W.S.; Han, X.; Lee, S.J.; Kim, Y.C.; Lee, O.H. Antiphotoaging effect of fermented agricultural byproducts on ultraviolet Birradiated hairless mouse skin. Int. J. Mol. Med. 2019, 44, 559–568. [Google Scholar]
- Liu, X.; Zhang, R.; Shi, H.; Li, X.; Li, Y.; Taha, A.; Xu, C. Protective effect of curcumin against ultraviolet A irradiationinduced photoaging in human dermal fibroblasts. Mol. Med. Rep. 2018, 17, 7227–7237. [Google Scholar]
- Hwang, B.M.; Noh, E.M.; Kim, J.S.; Kim, J.M.; You, Y.O.; Hwang, J.K.; Kwon, K.B.; Lee, Y.R. Curcumin inhibits UVB-induced matrix metalloproteinase-1/3 expression by suppressing the MAPK-p38/JNK pathways in human dermal fibroblasts. Exp. Dermatol. 2013, 22, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Berg, E.L.; Yang, J.; Melrose, J.; Nguyen, D.; Privat, S.; Rosler, E.; Kunkel, E.J.; Ekins, S. Chemical target and pathway toxicity mechanisms defined in primary human cell systems. J. Pharmacol. Toxicol. Methods 2010, 61, 3–15. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Cell Type | Environment | Readouts |
|---|---|---|---|
HDF3CGF  | Dermal fibroblasts | IL-1β + TNF-α + IFNγ+ bFGF + EGF + PDGF-BB | MCP-1, VCAM-1, ICAM-1, Collagen Ⅰ, Collagen Ⅲ, IP-10, I-TAC, IL-8, MIG, EGFR, M-CSF, MMP-1, PAI-1, Proliferation-72 h, SRB, TIMP-1, TIMP-2 |
| Target | Primer Sequence (5′-3′) |
|---|---|
| hCol1A1-F | GTGCGATGACGTGATCTGTGA |
| hCol1A1-R | CGGTGGTTTCTTGGTCGGT |
| hCol1A2-F | GGCCCTCAAGGTTTCCAAGG |
| hCol1A2-R | CACCCTGTGGTCCAACAACTC |
| hCol3A1-F | TTGAAGGAGGATGTTCCCATCT |
| hCol3A1-R | ACAGACACATATTTGGCATGGTT |
| hCol5A1-F | TACCCTGCGTCTGCATTTCC |
| hCol5A1-R | GCTCGTTGTAGATGGAGACCA |
| hMMP1-F | AAAATTACACGCCAGATTTGCC |
| hMMP1-R | GGTGTGACATTACTCCAGAGTTG |
| hMMP3-F | CTGGACTCCGACACTCTGGA |
| hMMP3-R | CAGGAAAGGTTCTGAAGTGACC |
| hMMP9-F | TGTACCGCTATGGTTACACTCG |
| hMMP9-R | GGCAGGGACAGTTGCTTCT |
| hGAPDH-F | GGAGCGAGATCCCTCCAAAAT |
| hGAPDH-R | GGCTGTTGTCATACTTCTCATGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, N.; Liu, Y.; Jin, J.; Ji, M.; Chen, X. Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2022, 23, 6941. https://doi.org/10.3390/ijms23136941
Xue N, Liu Y, Jin J, Ji M, Chen X. Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. International Journal of Molecular Sciences. 2022; 23(13):6941. https://doi.org/10.3390/ijms23136941
Chicago/Turabian StyleXue, Nina, Ying Liu, Jing Jin, Ming Ji, and Xiaoguang Chen. 2022. "Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts" International Journal of Molecular Sciences 23, no. 13: 6941. https://doi.org/10.3390/ijms23136941
APA StyleXue, N., Liu, Y., Jin, J., Ji, M., & Chen, X. (2022). Chlorogenic Acid Prevents UVA-Induced Skin Photoaging through Regulating Collagen Metabolism and Apoptosis in Human Dermal Fibroblasts. International Journal of Molecular Sciences, 23(13), 6941. https://doi.org/10.3390/ijms23136941