
Molecular Basis for Luteolin as a Natural TatD DNase Inhibitor in Trueperella pyogenes

Abstract
:1. Introduction
2. Results
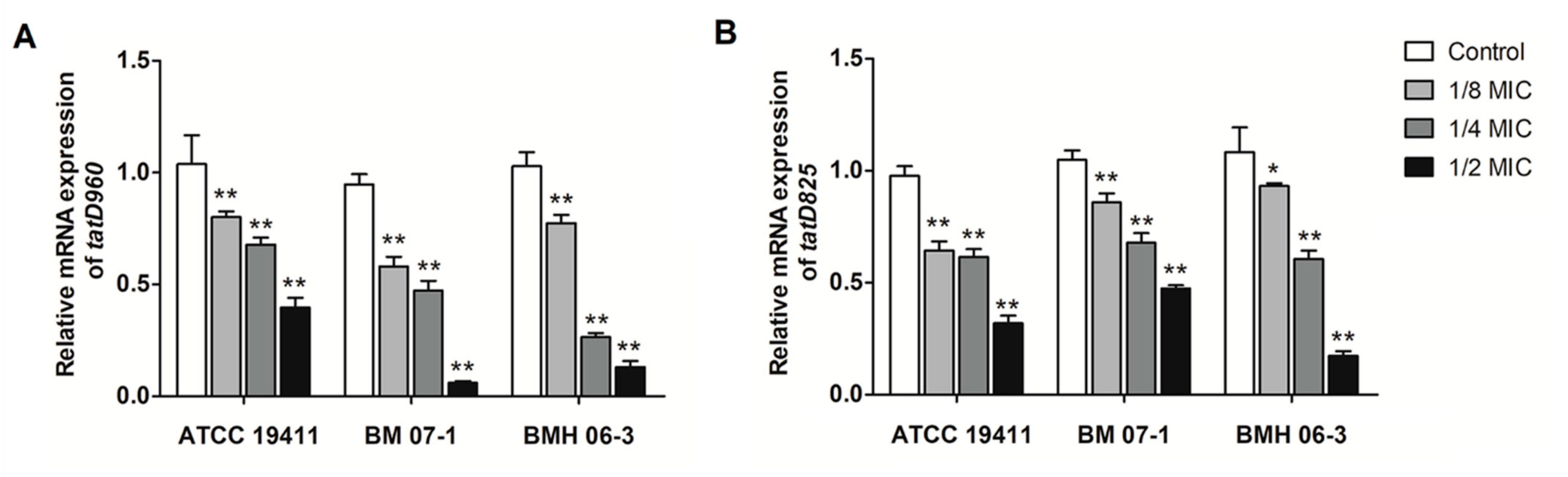
2.1. Effects of Luteolin on the Transcription of tatD960 and tatD825
2.2. Effects of Luteolin on the Protein Expression of TatD960 and TatD825
2.3. Treatment with Luteolin Leads to Inhibition of TatD DNases Activity
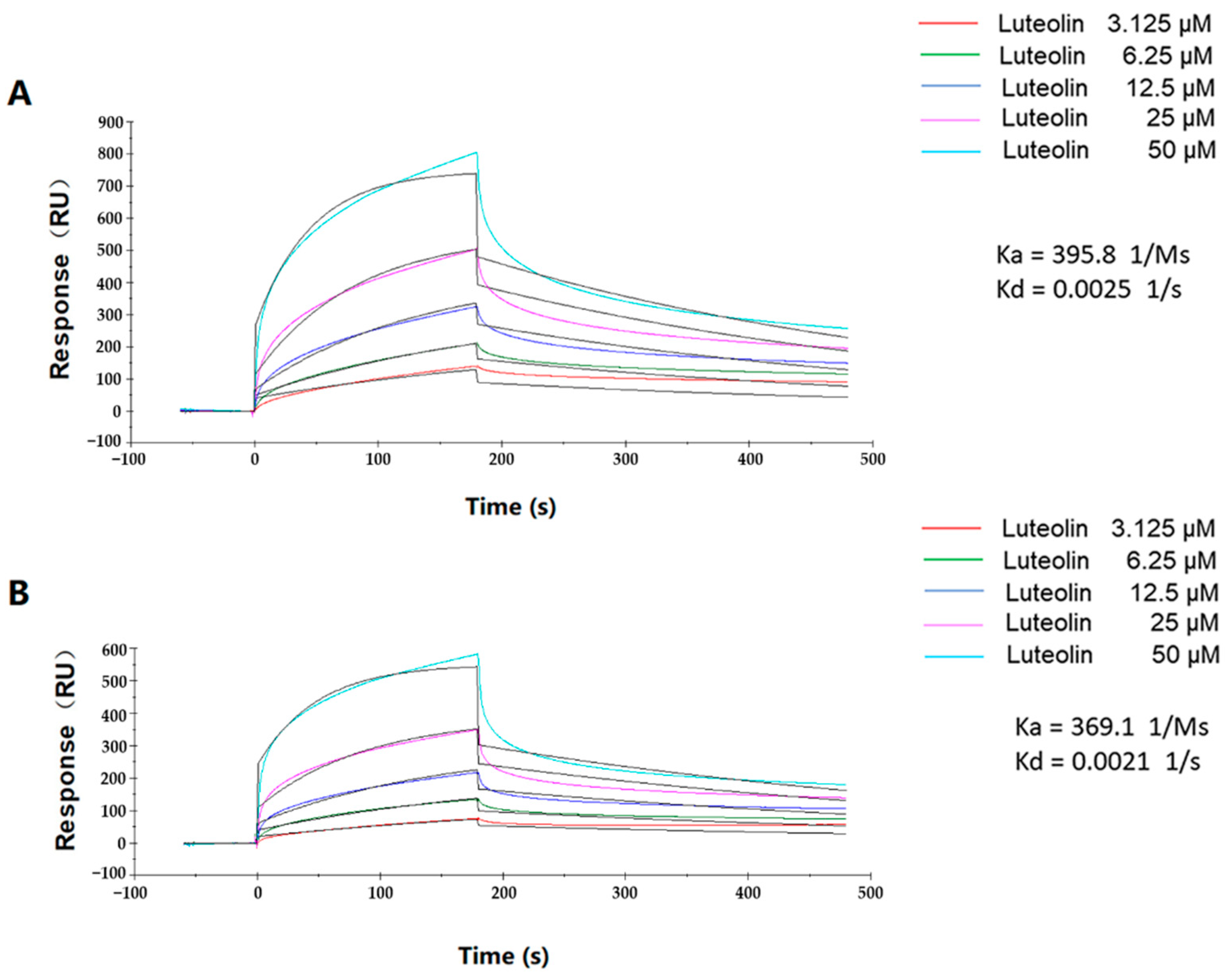
2.4. Binding Kinetics of Luteolin and TatD DNases
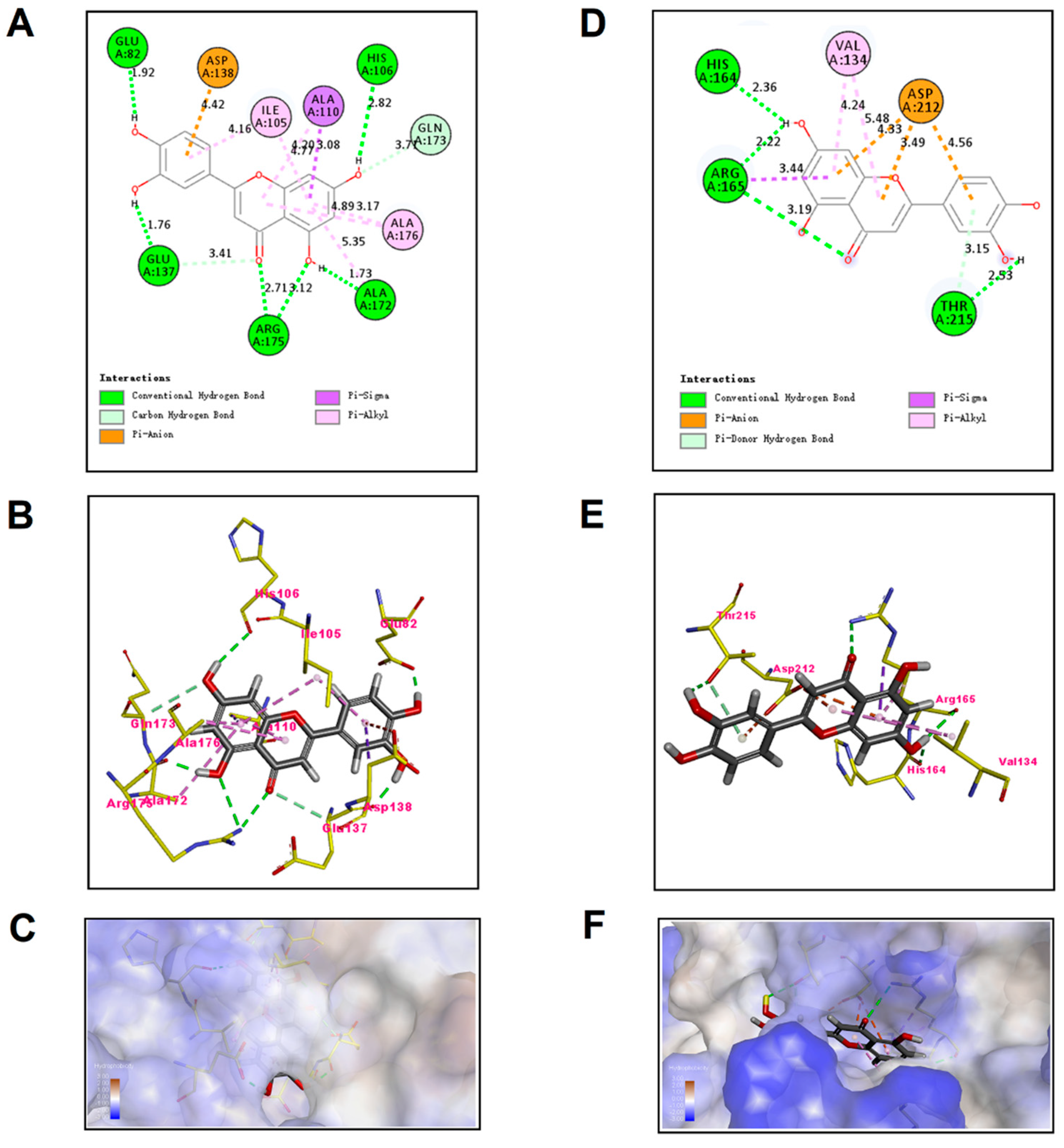
2.5. Molecular Docking Analysis of Luteolin and TatD DNases
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Strains and Reagents
4.3. Detection of the Expression of tatD Genes by Quantitative Real-Time PCR
4.4. Analysis of the Expression of TatD DNases Using Western Blot
4.5. Detection of the Activity of TatD DNases Treated with Luteolin
4.6. Measurement of the Kinetics of Luteolin with TatD DNases Using SPR
4.7. Molecular Docking of Luteolin to TatD DNases
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rzewuska, M.; Kwiecień, E.; Chrobak-Chmiel, D.; Kizerwetter-Świda, M.; Stefańska, I.; Gieryńska, M. Pathogenicity and Virulence of Trueperella pyogenes: A Review. Int. J. Mol. Sci. 2019, 20, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, M.G.; Risseti, R.M.; Bolaños, C.A.; Caffaro, K.A.; de Morais, A.C.; Lara, G.H.; Zamprogna, T.O.; Paes, A.C.; Listoni, F.J.; Franco, M.M. Trueperella pyogenes multispecies infections in domestic animals: A retrospective study of 144 cases (2002 to 2012). Vet. Q. 2015, 35, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Kwiecień, E.; Stefańska, I.; Chrobak-Chmiel, D.; Kizerwetter-Świda, M.; Moroz, A.; Olech, W.; Spinu, M.; Binek, M.; Rzewuska, M. Trueperella pyogenes Isolates from Livestock and European Bison (Bison bonasus) as a Reservoir of Tetracycline Resistance Determinants. Antibiotics 2021, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi Tamai, I.; Mohammadzadeh, A.; Zahraei Salehi, T.; Mahmoodi, P.; Pakbin, B. Investigation of antimicrobial susceptibility and virulence factor genes in Trueperella pyogenes isolated from clinical mastitis cases of dairy cows. Food Sci. Nutr. 2021, 9, 4529–4538. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, Ł.S.; Gradzki, Z.; Kalinowski, M. Trueperella pyogenes infections in swine: Clinical course and pathology. Pol. J. Vet. Sci. 2014, 17, 395–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Wang, B.; Wang, J.; Ma, B.; Zhang, W. Pyolysin of Trueperella pyogenes Induces Pyroptosis and IL-1β Release in Murine Macrophages Through Potassium/NLRP3/Caspase-1/Gasdermin D Pathway. Front. Immunol. 2022, 13, 832458. [Google Scholar] [CrossRef] [PubMed]
- Deliwala, S.; Beere, T.; Samji, V.; Mcdonald, P.J.; Bachuwa, G. When Zoonotic Organisms Cross Over-Trueperella pyogenes Endocarditis Presenting as a Septic Embolic Stroke. Cureus 2020, 12, e7740. [Google Scholar] [CrossRef] [Green Version]
- Meili, Z. Trueperella pyogenes pharyngitis in an immunocompetent 40-year-old man. BMJ Case Rep. 2020, 13, e236129. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Zhao, K.; Tian, Y.; Yue, B.; Wang, H.; Zhang, X. Virulence determinants and biofilm production among Trueperella pyogenes recovered from abscesses of captive forest musk deer. Arch. Microbiol. 2013, 195, 203–209. [Google Scholar] [CrossRef]
- Alkasir, R.; Wang, J.; Gao, J.; Ali, T.; Zhang, L.; Szenci, O.; Bajcsy, Á.C.; Han, B. Properties and antimicrobial susceptibility of Trueperella pyogenes isolated from bovine mastitis in China. Acta Vet. Hung. 2016, 64, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Duarte, V.; Dias, R.S.; Kropinski, A.M.; da Silva Xavier, A.; Ferro, C.G.; Vidigal, P.M.P.; da Silva, C.C.; de Paula, S.O. A T4virus prevents biofilm formation by Trueperella pyogenes. Vet. Microbiol. 2018, 218, 45–51. [Google Scholar] [CrossRef]
- Chen, Y.C.; Li, C.L.; Hsiao, Y.Y.; Duh, Y.; Yuan, H.S. Structure and function of TatD exonuclease in DNA repair. Nucleic Acids Res. 2014, 42, 10776–10785. [Google Scholar] [CrossRef] [Green Version]
- Jhelum, H.; Sori, H.; Sehgal, D. A novel extracellular vesicle-associated endodeoxyribonuclease helps Streptococcus pneumoniae evade neutrophil extracellular traps and is required for full virulence. Sci. Rep. 2018, 8, 7985. [Google Scholar] [CrossRef]
- Chang, Z.; Jiang, N.; Zhang, Y.; Lu, H.; Yin, J.; Wahlgren, M.; Cheng, X.; Cao, Y.; Chen, Q. The TatD-like DNase of Plasmodium is a virulence factor and a potential malaria vaccine candidate. Nat. Commun. 2016, 7, 11537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liang, Y.; Yu, L.; Chen, M.; Guo, Y.; Kang, Z.; Qu, C.; Tian, C.; Zhang, D.; Liu, M. TatD DNases Contribute to Biofilm Formation and Virulence in Trueperella pyogenes. Front. Microbiol. 2021, 12, 758465. [Google Scholar] [CrossRef]
- Zhang, T.; Qiu, Y.; Luo, Q.; Zhao, L.; Yan, X.; Ding, Q.; Jiang, H.; Yang, H. The Mechanism by Which Luteolin Disrupts the Cytoplasmic Membrane of Methicillin-Resistant Staphylococcus aureus. J. Phys. Chem. B. 2018, 122, 1427–1438. [Google Scholar] [CrossRef]
- Usman Amin, M.; Khurram, M.; Khan, T.A.; Faidah, H.S.; Ullah Shah, Z.; Ur Rahman, S.; Haseeb, A.; Ilyas, M.; Ullah, N.; Umar Khayam, S.M.; et al. Effects of Luteolin and Quercetin in Combination with Some Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2016, 17, 1947. [Google Scholar] [CrossRef] [Green Version]
- Siriwong, S.; Thumanu, K.; Hengpratom, T.; Eumkeb, G. Synergy and Mode of Action of Ceftazidime plus Quercetin or Luteolin on Streptococcus pyogenes. Evid.-Based Complement. Alternat. Med. 2015, 2015, 759459. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Fu, Y.; Liu, M.; Zhang, J.; Wang, W.; Li, J.; Zeng, Q.; Wang, T.; Li, Y. Mechanisms of Action of Luteolin Against Single- and Dual-Species of Escherichia coli and Enterobacter cloacae and Its Antibiofilm Activities. Appl. Biochem. Biotechnol. 2021, 193, 1397–1414. [Google Scholar] [CrossRef]
- Qiu, J.; Li, H.; Meng, H.; Hu, C.; Li, J.; Luo, M.; Dong, J.; Wang, X.; Wang, J.; Deng, Y.; et al. Impact of luteolin on the production of alpha-toxin by Staphylococcus aureus. Lett. Appl. Microbiol. 2011, 53, 238–243. [Google Scholar] [CrossRef]
- Shen, X.F.; Ren, L.B.; Teng, Y.; Zheng, S.; Yang, X.L.; Guo, X.J.; Wang, X.Y.; Sha, K.H.; Li, N.; Xu, G.Y.; et al. Luteolin decreases the attachment, invasion and cytotoxicity of UPEC in bladder epithelial cells and inhibits UPEC biofilm formation. Food Chem. Toxicol. 2014, 72, 204–211. [Google Scholar] [CrossRef]
- Rivera, M.L.C.; Hassimotto, N.M.A.; Bueris, V.; Sircili, M.P.; de Almeida, F.A.; Pinto, U.M. Effect of Capsicum Frutescens Extract, Capsaicin, and Luteolin on Quorum Sensing Regulated Phenotypes. J. Food. Sci. 2019, 84, 1477–1486. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, Y.; Zhang, Z.; Chen, M.; Zhang, D.; Tian, C.; Liu, M.; Jiang, G. The Antibacterial Activity and Mechanism of Action of Luteolin Against Trueperella pyogenes. Infect. Drug Resist. 2020, 13, 1697–1711. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, X.; Song, X.; Zhou, W.; Hong, W.; Tian, C.; Liu, Y.; Liu, M. Luteolin Showed a Resistance Elimination Effect on Gentamicin by Decreasing MATE mRNA Expression in Trueperella pyogenes. Microb. Drug Resist. 2019, 25, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Zuo, R.; de Oliveira, A.; Bullita, E.; Torino, M.I.; Padgett-Pagliai, K.A.; Gardner, C.L.; Harrison, N.A.; da Silva, D.; Merli, M.L.; Gonzalez, C.F.; et al. Identification of flavonoids as regulators of YbeY activity in Liberibacter asiaticus. Environ. Microbiol. 2019, 21, 4822–4835. [Google Scholar] [CrossRef] [PubMed]
- Sanna, C.; Marengo, A.; Acquadro, S.; Caredda, A.; Lai, R.; Corona, A.; Tramontano, E.; Rubiolo, P.; Esposito, F. In Vitro Anti-HIV-1 Reverse Transcriptase and Integrase Properties of Punica granatum L. Leaves, Bark, and Peel Extracts and Their Main Compounds. Plants 2021, 10, 2124. [Google Scholar] [CrossRef] [PubMed]
- Lv, P.C.; Li, H.Q.; Xue, J.Y.; Shi, L.; Zhu, H.L. Synthesis and biological evaluation of novel luteolin derivatives as antibacterial agents. Eur. J. Med. Chem. 2009, 44, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.F.; Yang, C.; Zhang, Y.; Tao, S.N.; Mei, J.; Zhang, X.C.; Sun, Y.J.; Zhao, B.T. An innovative role for luteolin as a natural quorum sensing inhibitor in Pseudomonas aeruginosa. Life Sci. 2021, 274, 119325. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, L.; Ji, S.; Hou, S.; Luo, L.; Li, C.; Liu, M.; Liu, Y.; Jiang, L. Structural Basis for the Inhibition of the Autophosphorylation Activity of HK853 by Luteolin. Molecules 2019, 24, 933. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Huang, C.; Su, H.; Zhang, Z.; Chen, M.; Wang, R.; Zhang, D.; Zhang, L.; Liu, M. Luteolin increases susceptibility to macrolides by inhibiting MsrA efflux pump in Trueperella pyogenes. Vet. Res. 2022, 53, 3. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S.; Liu, B.; Niu, X.; Deng, X. Luteolin Inhibits Listeriolysin O Translation by Directly Targeting the Coding Region of the hly mRNA. Front. Microbiol. 2019, 10, 1496. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, W.; Zeng, Q.; Wang, T.; Qian, W. Antibiofilm Efficacy of Luteolin Against Single and Dual Species of Candida albicans and Enterococcus faecalis. Front. Microbiol. 2021, 12, 715156. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′-3′) | Product Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| tatD960-F | GTGGACCTTCTGCTGCGTGAC | 103 | 60 |
| tatD960-R | CATACCAGCCGTGCTCCTTGC | ||
| tatD825-F | GCCGCCTGCTTGACCATATCG | 119 | 60 |
| tatD825-R | GGTGCTGGAGCCAGATGATTCG | ||
| 16S rRNA-F | ATGCAACGCGAAGAACCTTACC | 127 | 60 |
| 16S rRNA-R | TTAACCCAACATCTCACGACAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Guo, Y.; Guo, Y.; Zhang, L.; Niu, S.; Tian, C.; Han, L.; Zhang, D.; Liu, M. Molecular Basis for Luteolin as a Natural TatD DNase Inhibitor in Trueperella pyogenes. Int. J. Mol. Sci. 2022, 23, 8374. https://doi.org/10.3390/ijms23158374
Zhang Z, Guo Y, Guo Y, Zhang L, Niu S, Tian C, Han L, Zhang D, Liu M. Molecular Basis for Luteolin as a Natural TatD DNase Inhibitor in Trueperella pyogenes. International Journal of Molecular Sciences. 2022; 23(15):8374. https://doi.org/10.3390/ijms23158374
Chicago/Turabian StyleZhang, Zehui, Yuru Guo, Yueting Guo, Luyao Zhang, Shengli Niu, Chunlian Tian, Limei Han, Dexian Zhang, and Mingchun Liu. 2022. "Molecular Basis for Luteolin as a Natural TatD DNase Inhibitor in Trueperella pyogenes" International Journal of Molecular Sciences 23, no. 15: 8374. https://doi.org/10.3390/ijms23158374
APA StyleZhang, Z., Guo, Y., Guo, Y., Zhang, L., Niu, S., Tian, C., Han, L., Zhang, D., & Liu, M. (2022). Molecular Basis for Luteolin as a Natural TatD DNase Inhibitor in Trueperella pyogenes. International Journal of Molecular Sciences, 23(15), 8374. https://doi.org/10.3390/ijms23158374