
Root Foraging Strategy Improves the Adaptability of Tea Plants (Camellia sinensis L.) to Soil Potassium Heterogeneity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Nutrition and Growth of Tea Plants in the Multi-Layer Split-Root System
2.2. Temporal and Spatial Variations of Soil K Concentrations and Root Development
2.3. Release of Organic Acid and Solubilization of Soil K by Root Exudates
2.4. Potential Key Genes Involved in Root K Foraging
3. Discussion
4. Methods and Materials
4.1. Plant and Soil Materials
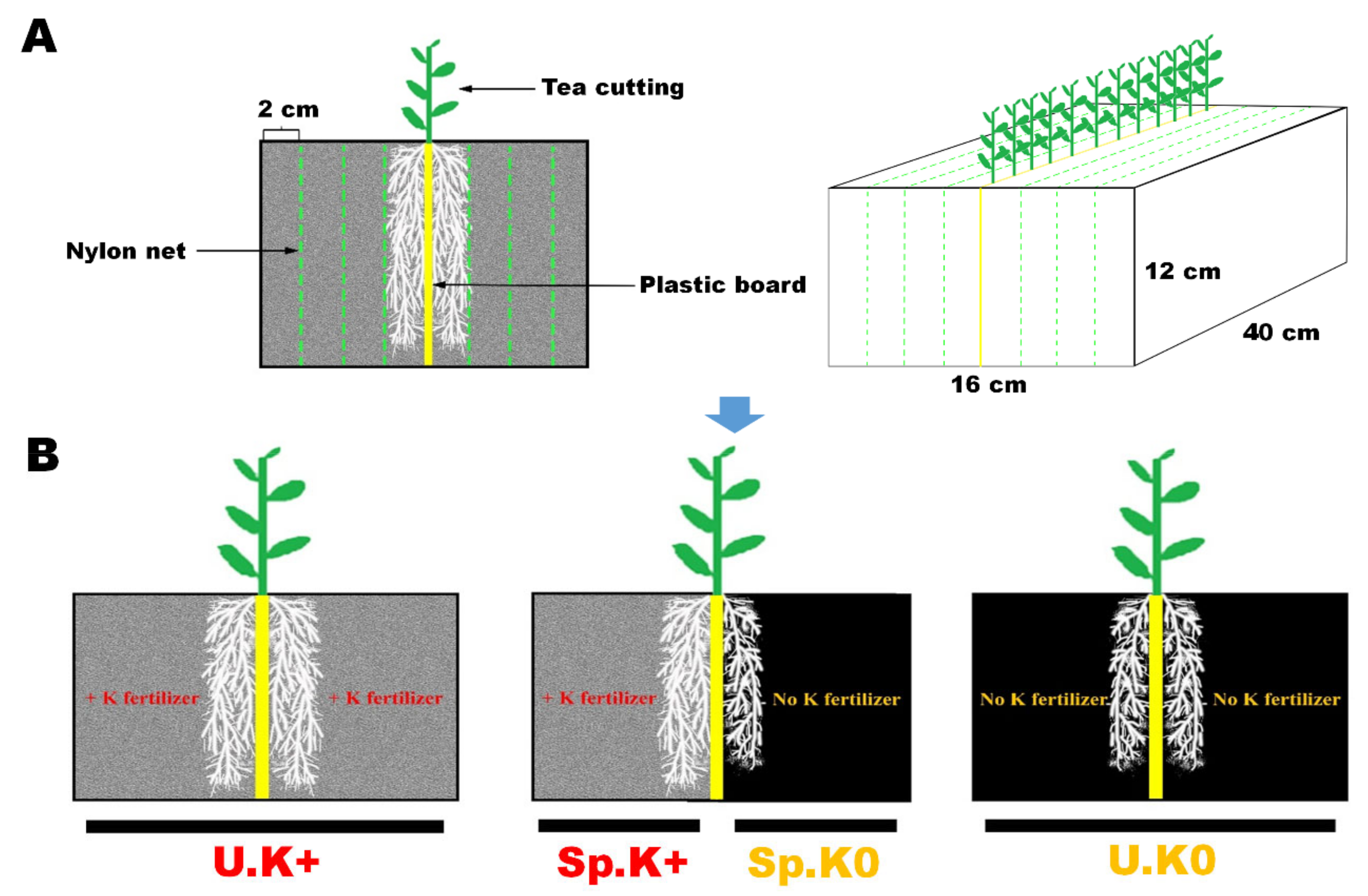
4.2. Experiment on Soil Culture with Multi-Layer Split-Root Box
4.3. Determinations of Soil K Contents, Root Morphology, and Plant K Contents
4.4. Root Exudate Collection and Soil K Solubilization Test
4.5. Culture Experiment with a Split-Root System
4.6. Transcriptome Analysis and qRT-PCR Verification
4.7. Determinations of Antioxidant Enzyme Activities and Polyphenol Contents
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Römheld, V.; Kirkby, E. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture-status and perspectives. J. Plant Physio. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.Y.; Ma, L.F.; Shi, Y.Z. Potassium management in tea plantations: Its uptake by field plants, status in soils, and efficacy on yields and quality of teas in China. J. Plant Nutr. Soil Sci. 2013, 176, 450–459. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; Wirén, N.V. Root nutrient foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Lotfollah, A.; Munkholm, L.J. Tillage system and cover crop effects on soil quality: I. chemical, mechanical, and biological properties. Soil Sci. Soc. Am. J. 2014, 78, 262–270. [Google Scholar] [CrossRef]
- Fernández, F.G.; Sorensen, B.A.; Villamil, M.B.A. Comparison of soil properties after five years of no-till and strip-till. Agron. J. 2015, 107, 1339–1346. [Google Scholar] [CrossRef]
- Williams, A.; Davis, A.S.; Jilling, A.; Grandy, A.S.; Koide, R.T.; Mortensen, D.A.; Smith, R.G.; Snapp, S.S.; Spokas, K.A.; Yannarell, A.C.; et al. Reconciling opposing soil processes in row-crop agroecosystems via soil functional zone management. Agr. Ecosyst. Environ. 2017, 236, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; McCormack, M.L.; Ma, Z.; Guo, D. Diverse belowground resource strategies underlie plant species coexistence and spatial distribution in three grasslands along a precipitation gradient. New Phytol. 2017, 216, 1140–1150. [Google Scholar] [CrossRef] [Green Version]
- Kellermeier, F.; Amtmann, C.A. Natural variation of Arabidopsis root architecture reveals complementing adaptive strategies to potassium starvation. Plant Physiol. 2013, 161, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Penuelas, J. Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood. Global Ecol. Biogeogr. 2013, 22, 494–507. [Google Scholar] [CrossRef]
- Wang, H.Y.; Shen, Q.H.; Zhou, J.M.; Wang, J.; Du, C.W.; Chen, X.Q. Plants use alternative strategies to utilize nonexchangeable potassium in minerals. Plant Soil 2011, 343, 209–220. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Ruyter-Spira, C.; Bouwmeester, H.J. Engineering the plant rhizosphere. Curr. Opin. Biotech. 2016, 32, 136–142. [Google Scholar] [CrossRef]
- Zhao, X.H.; Yu, H.Q.; Wen, J.; Wang, X.G.; Du, Q.; Wang, J.; Wang, Q. Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency. J. Integr. Agr. 2016, 15, 785–794. [Google Scholar] [CrossRef]
- Lee, S.; Sergeeva, L.I.; Vreugdenhil, D. Natural variation of hormone levels in Arabidopsis roots and correlations with complex root architecture. J. Integr. Plant Bio. 2017, 60, 292–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Wang, W.Z.; Wei, K.; Ruan, L.; Wang, L.Y.; Cheng, H.; Zhang, F.; Wu, L.Y.; Bai, P.X. Differential transcriptomic changes in low-potassium sensitive and low-potassium tolerant tea plant (Camellia sinensis) genotypes under potassium deprivation. Sci. Hortic. 2019, 256, 108570. [Google Scholar] [CrossRef]
- Chen, W.; Koide, R.T.; Eissenstat, D.M. Root morphology and mycorrhizal type strongly influence root production in nutrient hot spots of mixed forests. J. Ecol. 2017, 106, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Koide, R.T.; Eissenstat, D.M. Nutrient foraging by mycorrhizas: From species functional traits to ecosystem processes. Funct. Ecol. 2018, 32, 1365–2435. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.P. Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Whalley, W.R.; Miller, A.J.; White, P.J.; Shen, J. Sustainable cropping requires adaptation to a heterogeneous rhizosphere. Trends Plant Sci. 2020, 25, 1194–1202. [Google Scholar] [CrossRef]
- Li, J.W.; Li, X.; Hao, G.S.; Zhang, F.F.; Ruan, L.; Wang, W.Z. Rhizosphere processes of tea (Camellia sinensis) plants under spatial heterogeneity of soil potassium. Rhizosphere 2021, 17, 100299. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Z.C.; Yang, Q. Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions. Plant Soil 2013, 364, 93–104. [Google Scholar] [CrossRef]
- Li, W.; Xiang, F.; Zhong, M.C.; Zhou, L.Y.; Liu, H.Y.; Li, S.J.; Wang, X.W. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Flores, R.; Winkel, T.; Nguyen-Thi-Truc, A.; Joffre, R. Root foraging capacity depends on root system architecture and ontogeny in seedlings of three Andean Chenopodium species. Plant Soil 2014, 380, 415–428. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, B.; Ge, X.; Cao, Y.; Li, M.H. Species-specific responses of root morphology of three co-existing tree species to nutrient patches reflect their root foraging strategies. Front Plant Sci. 2021, 11, 618222. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Shin, R. Nutrient sensing and signaling: NPKS. Annu. Rev. Plant Bio. 2007, 58, 47–69. [Google Scholar] [CrossRef]
- Fernández, F.G.; Brouder, S.M.; Volenec, J.J.; Beyrout, C.A.; Hoyum, R. Soybean shoot and root response to localized water and potassium in a split-pot study. Plant Soil 2011, 344, 197–212. [Google Scholar] [CrossRef]
- Ruan, L.; Xin, X.L.; Zhang, J.B.; Zhao, B.Z.; Cheng, H.; Zhang, C.Z.; Ma, D.H.; Chen, L. Potential root foraging strategy of wheat (Triticum aestivum L.) for potassium heterogeneity. Front Plant Sci. 2018, 9, 1755. [Google Scholar] [CrossRef]
- Bouain, N.; Krouk, G.; Lacombe, B.; Rouached, H. Getting to the root of plant mineral nutrition: Combinatorial nutrient stresses reveal emergent properties. Trends Plant Sci. 2019, 24, 542–552. [Google Scholar] [CrossRef]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef]
- Freschet, G.T.; Valverde-Barrantes, O.J.; Tucker, C.M.; Craine, J.M.; McCormack, M.L.; Violle, C.; Fort, F.; Blackwood, C.B.; Urban-Mead, K.R.; Iversen, C.M.; et al. Climate, soil and plant functional types as drivers of global fine-root trait variation. J. Ecol. 2017, 105, 1182–1196. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Jia, C.G.; Zhuo, W.Z.; Zheng, Q.W. Responses of fine root mass, length, production and turnover to soil nitrogen fertilization in Larix gmelinii and Fraxinus mandshurica forests in Northeastern China. J. Forest Res. 2010, 15, 194–201. [Google Scholar] [CrossRef]
- Lambers, H.; Martinoia, E.; Renton, M. Plant adaptations to severely phosphorus-impoverished soils. Curr. Opin. Plant Biol. 2015, 25, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Nasto, M.K.; Osborne, B.B.; Lekberg, Y.; Asner, G.P.; Balzotti, C.S.; Porder, S.; Taylor, P.G.; Townsend, A.R.; Cleveland, C.C. soil phosphorus partitioning and competition among trees in a lowland tropical rain forest. New Phytol. 2017, 214, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.S.; Shen, J.B.; Zhang, J.L.; Zuo, Y.M.; Li, L.; Chen, X.P. Rhizosphere processes and management for improving nutrient use efficiency and crop productivity: Implications for China. Adv. Agron. 2010, 107, 1–32. [Google Scholar] [CrossRef]
- Dreyer, I.; Gomez-Porras, J.L.; Riedelsberger, J. The potassium battery: A mobile energy source for transport processes in plant vascular tissues. New Phytol. 2017, 216, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, W.H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency. Curr. Opin. Plant Biol. 2015, 25, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, D.J. Diffuse growth of plant cell walls. Plant Physiol. 2018, 176, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Cheddadi, I.; Mosca, G. Cellular heterogeneity in pressure and growth emerges from tissue topology and geometry. Curr. Biol. 2020, 30, 1504–1516. [Google Scholar] [CrossRef]
- Wang, Y.; Jiao, Y.L. Cellulose microfibril-mediated directional plant cell expansion: Gas and Brake. Mol. Plant 2020, 13, 1670–1672. [Google Scholar] [CrossRef]
- Mishra, B.S.; Singh, M.; Aggrawal, P.A. Glucose and auxin signaling interaction in controlling Arabidopsis thaliana seedlings root growth and development. PLoS ONE 2009, 4, e4502. [Google Scholar] [CrossRef] [PubMed]
- Leclere, S.S.E.; Chourey, P.S. Sugar levels regulate tryptophan-dependent auxin biosynthesis in developing maize kernels. Plant Physiol. 2010, 153, 306–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zhao, J.; Yuan, J.; Hale, L.; Wen, T.; Huang, Q.W.; Vivanco, J.M.; Zhou, J.Z.; Kowalchuk, G.A.; Shen, Q.R. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2020, 44, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Chinese Soil Taxonomy Research Group (CST). Chinese Soil Taxonomy; Science Press: Beijing, China, 2001. [Google Scholar]
- Wu, K.N.; Zhao, R. Soil texture classification and its application in China. Acta Petrol. Sin. 2019, 56, 227–241. (In Chinese) [Google Scholar] [CrossRef]
- Ruan, J.Y.; Zhang, F.S.; Ming, H.W. Effect of nitrogen form and phosphorus source on the growth, nutrient uptake and rhizosphere soil property of Camellia sinensis L. Plant Soil 2000, 223, 65–73. [Google Scholar] [CrossRef]
- Yang, Y.J. Tea Cultivation in China; Shanghai Science and Technology Press: Shanghai, China, 2004. [Google Scholar]
- Bao, S.D. Soil and Agricultura Chemistry Analysis; China Agricultura Press: Beijing, China, 2000. [Google Scholar]
- Singh, U.; Ladha, J.K.; Castillo, E.G.; Punzalan, G.; Duqueza, M. Genotypic variation in nitrogen use efficiency in medium- and long-duration rice. Field Crop Res. 1998, 58, 35–53. [Google Scholar] [CrossRef]
- Chen, R.F.; Shen, R.F.; Yang, X.D.; Wang, X.L. Effects of buckwheat growth on variation of aluminum and major metals in root-zone soil solutions. J. Plant Nutr. Soil Sci. 2010, 173, 788–794. [Google Scholar] [CrossRef]
- Lin, Z.H.; Chen, L.S.; Chen, R.B.; Zhang, F.Z.; Jiang, H.X.; Tang, N.; Smithd, B.R. Root release and metabolism of organic acids in tea plants in response to phosphorus supply. J. Plant Physiol. 2011, 168, 644–652. [Google Scholar] [CrossRef]
- Ruan, L.; Zhang, J.; Xin, X.; Miller, A.J.; Tong, Y. Elymus dahuricus H+-PPase EdVP1 enhances potassium uptake and utilization of wheat through the development of root system. J. Soil Sci. Plant Nutr. 2013, 13, 716–729. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.Y.; Gerendás, J.; Haerdter, R.; Sattelmacher, B. Effect of root-zone pH and form and concentration of nitrogen on the accumulation of quality-related components in green tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Liu, M.Y.; Burgos, A.; Zhang, Q.F.; Tang, D.D.; Shi, Y.Z.; Ma, L.F.; Yi, X.Y.; Ruan, J.Y. Analyses of transcriptome profiles and selected metabolites unravel the metabolic response to NH4+ and NO3− as signaling molecules in tea plant (Camellia sinensis L.). Sci. Hortic. 2017, 218, 293–303. [Google Scholar] [CrossRef]
- Wei, C.L.; Yang, H.; Wang, S.B.; Zhao, J.; Liu, C.; Gao, L. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, 4151–4158. [Google Scholar]
- Xiong, L.G.; Li, J.; Li, Y.H.; Yuan, L.; Liu, S.Q.; Huang, J.A.; Liu, Z.H. Dynamic changes in catechin levels and catechin biosynthesis-related gene expression in albino tea plants (Camellia sinensis L.). Plant Physiol. Biochem. 2013, 71, 132–143. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Qian, H.; Chen, L.; Sun, B.; Chang, J.; Miao, H.; Cai, C.; Wang, Q. Influence of pre-harvest red light irradiation on main phytochemicals and antioxidant activity of Chinese kale sprouts. Food Chem. 2017, 222, 1–5. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, L.; Cheng, H.; Ludewig, U.; Li, J.; Chang, S.X. Root Foraging Strategy Improves the Adaptability of Tea Plants (Camellia sinensis L.) to Soil Potassium Heterogeneity. Int. J. Mol. Sci. 2022, 23, 8585. https://doi.org/10.3390/ijms23158585
Ruan L, Cheng H, Ludewig U, Li J, Chang SX. Root Foraging Strategy Improves the Adaptability of Tea Plants (Camellia sinensis L.) to Soil Potassium Heterogeneity. International Journal of Molecular Sciences. 2022; 23(15):8585. https://doi.org/10.3390/ijms23158585
Chicago/Turabian StyleRuan, Li, Hao Cheng, Uwe Ludewig, Jianwu Li, and Scott X. Chang. 2022. "Root Foraging Strategy Improves the Adaptability of Tea Plants (Camellia sinensis L.) to Soil Potassium Heterogeneity" International Journal of Molecular Sciences 23, no. 15: 8585. https://doi.org/10.3390/ijms23158585
APA StyleRuan, L., Cheng, H., Ludewig, U., Li, J., & Chang, S. X. (2022). Root Foraging Strategy Improves the Adaptability of Tea Plants (Camellia sinensis L.) to Soil Potassium Heterogeneity. International Journal of Molecular Sciences, 23(15), 8585. https://doi.org/10.3390/ijms23158585