
Lipid-Based Molecules on Signaling Pathways in Autism Spectrum Disorder


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
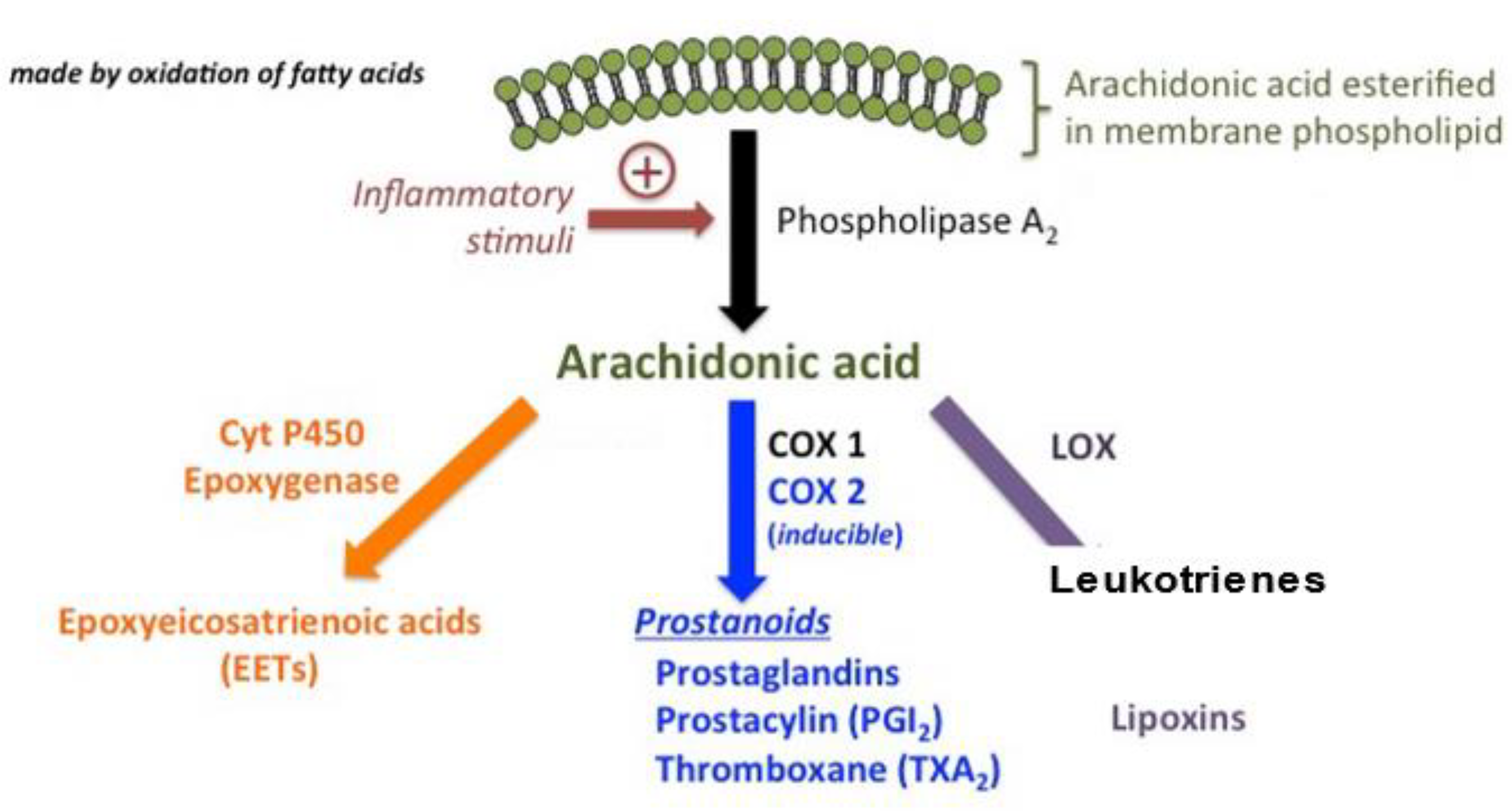
2. Signaling Pathways Associated with Lipid Metabolites
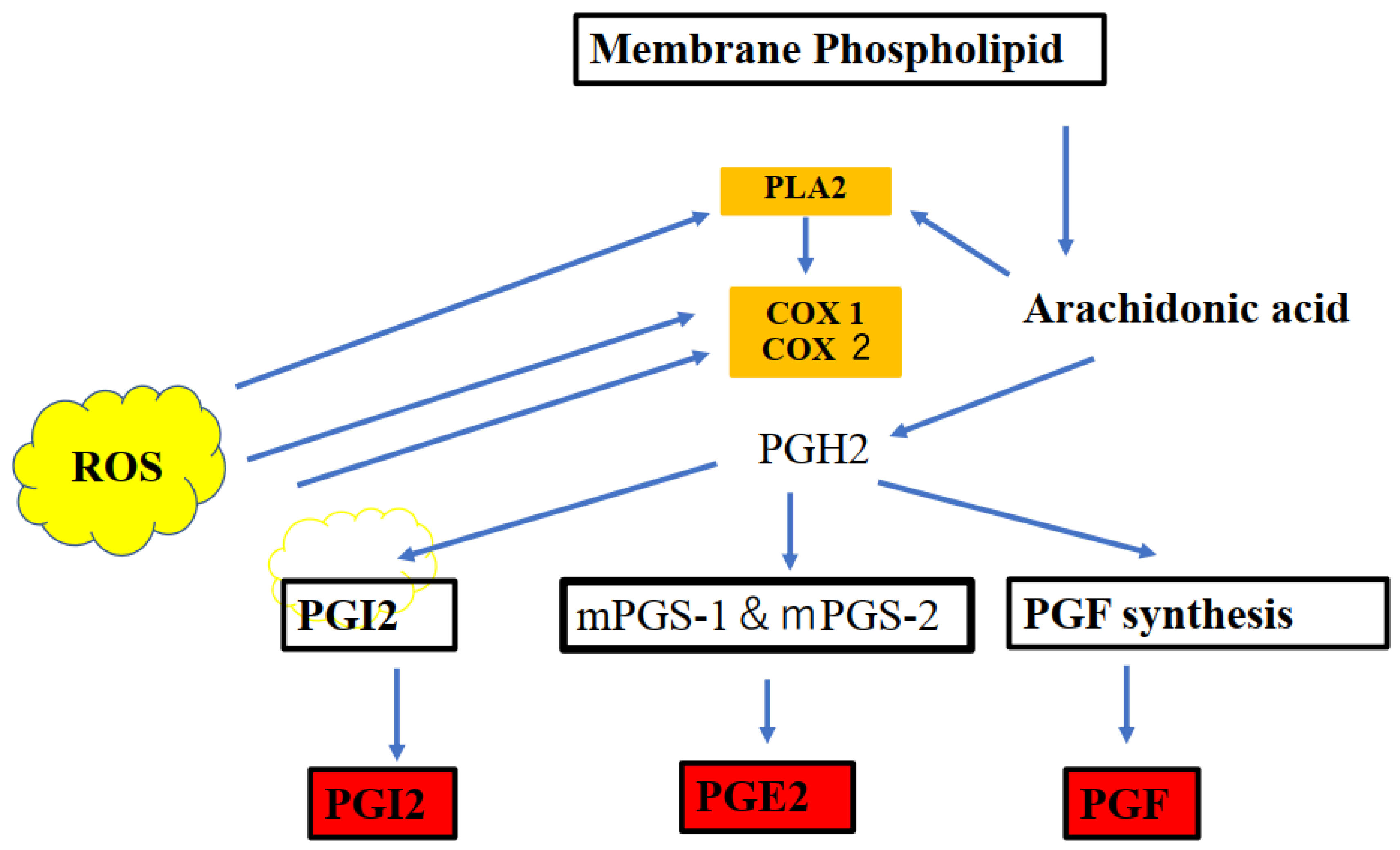
3. Prostaglandin E2 (PGE2)
3.1. Characteristics
3.2. PGE2 in ASD
3.3. Perspectives on ASD Therapy
4. Cyclooxygenase-2 (COX2)
5. The Association between PGE2 and COX-2 in ASD
6. Endocannabinoid Signaling
6.1. The Role of Endocannabinoids in Brain Function
6.2. The Role of eCBs in the Development of Brain
6.3. The Role of Cannabinoid Compounds of Cannabigerol, CB1 and CB2 Receptors, and Cannabidivarin in ASD Symptoms
6.4. ASD and Endocannabinoids: The Role of Cannabidivarin and 2-AG
6.5. Therapeutic Potential of the Endocannabinoid System for ASD
6.6. Endocannabinoid and Valproic Acid-Induced Model of ASD
7. Redox Signaling
7.1. The Role of Redox Signaling
7.2. Reactive Nitrogen and Oxygen Species
7.3. The Association between Redox Signaling and Iron
8. Copper Signaling
8.1. The Role of Copper Signaling
8.2. Copper Signaling and ASD
9. Iron Signaling and ASD
10. The Association between the Endocannabinoid Systems and Redox Signaling
11. Modulation of Endocannabinoid Action in Neuroinflammation Is Related to ASD
12. The Association between Lipid Metabolism and the Endocannabinoid System and Its Related Signaling Pathways
13. Dysregulation of the ECS in Fragile X Syndrome
14. Limitation of This Review
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lai, M.C.; Lombardo, M.V.; Baron-Cohen, S. Austism. Lancet 2014, 383, 896–910. [Google Scholar] [CrossRef]
- Lord, C.; Elsabbagh, M.; Baird, G.; Veenstra-Vanderweele, J. Autismspectrum disorder. Lancet 2018, 392, 508–520. [Google Scholar] [CrossRef]
- Levy, S.E.; Mandell, D.S.; Schultz, R.T. Autism. Lancet 2009, 374, 1627–1638. [Google Scholar] [CrossRef]
- Trifonova, E.A.; Klimenko, A.I.; Mustafin, Z.S.; Lashin, S.A.; Kochetov, A.V. Do autism spectrum and autoimmune disorders share predisposition gene signature due to mTOR signaling pathway controlling expression? Int. J. Mol. Sci. 2021, 22, 5248. [Google Scholar] [CrossRef]
- Kissoondoyal, A.; Crawford, D.A. Prostaglandin E2 Increases neurite length and the formation of axonal loops, and regulates cone turning in differentiating NE4C cells via PKA. Cell Mol. Neurobiol. 2022, 42, 1385–1397. [Google Scholar] [CrossRef]
- Chakrabarti, B.; Persico, A.; Battista, N.; Maccarrone, M. Endocannabinoid signaling in autism. Neurotherapeutics 2015, 12, 837–847. [Google Scholar] [CrossRef]
- Wong, C.T.; Ahmad, E.; Li, H.; Crawford, D.A. Prostaglandin E2 alters Wnt-dependent migration and proliferation in neuroectodermal stem cells: Implications for autism spectrum disorders. Cell Commun. Signal. 2014, 12, 19. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal. Transduct. Target Ther. 2021, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.T.; Bestard-Lorigados, I.; Crawford, D.A. Autism-related behaviors in the cyclooxygenase-2-deficient mouse model. Genes Brain Behav. 2019, 18, e12506. [Google Scholar] [CrossRef]
- Pradhan, S.S.; Salinas, K.; Garduno, A.C.; Johansson, J.U.; Wang, Q.; Manning-Bog, A.; Andreasson, K.I. Anti-inflammatory and neuroprotective effects of PGE (2) EP4 signaling in models of parkinson’s disease. Neuroimmune Pharmacol. 2017, 2, 92–304. [Google Scholar] [CrossRef]
- Rai-Bhogal, R.; Wong, C.; Kissoondoyal, A.; Davidson, J.; Li, H.; Crawford, D.A. Maternal exposure to prostaglandin E(2) modifies expression of Wnt genes in mouse brain—An autism connection. Biochem. Biophys. Rep. 2018, 14, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Yan, Y.; Li, Q.; Ye, J.; Pei, L. Endocannabinoid system jnlocks the puzzle of autism treatment via microglia. Front. Psychiatry 2021, 12, 734837. [Google Scholar] [CrossRef] [PubMed]
- Schultz, S.; Gould, G.G.; Antonucci, N.; Brigida, A.L.; Siniscalco, D. Endocannabinoid system dysregulation from acetaminophen use may lead to autism spectrum disorder: Could cannabinoid treatment be efficacious? Molecules 2021, 26, 1845. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.G.; Rocha, J.M.; Rossi, G.N.; Osório, F.L.; Ona, G.; Bouso, J.C.; Silveira, G.O.; Yonamine, M.; Marchioni, C.; Crevelin, E.J.; et al. Effects of ayahuasca on the endocannabinoid system of healthy volunteers and in volunteers with social anxiety disorder: Results from two pilot, proof-of-concept, randomized, placebo-controlled trials. Hum. Psychopharmacol. 2022, 37, e2834. [Google Scholar] [CrossRef]
- Davidson, J.M.; Wong, C.T.; Li, H.; Craford, D.A. Prostaglandin E2 facilitates subcellular translocation of the EP4 receptor in neuroectodermal NE-4C stem cells. Biochem. Biophys. Rep. 2016, 7, 173–179. [Google Scholar] [CrossRef]
- Tithof, P.K.; Roberts, M.P.; Guan, W.; Elgayyar, M.; Godkin, J.D. Distinct phospholipase A2 enzymes regulate prostaglandin E2 and F2 alpha production by bovine endometrial epithelial cells. Reprod. Biol. Endocrinol. 2007, 5, 16. [Google Scholar] [CrossRef]
- Ong, W.Y.; Farooqui, T.; Kokotos, G.; Farooqui, A.A. Synthetic and natural inhibitors of phospholipases A2: Their importance for understanding and treatment of neurological disorders. ACS. Chem. Neurosci. 2015, 6, 814–831. [Google Scholar] [CrossRef]
- Yang, H.; Chen, C. Cyclooxygenase-2 in synaptic signaling. Curr. Pharm. Des. 2008, 14, 1443–1451. [Google Scholar] [CrossRef]
- Sethi, R.; Gómez-Coronado, N.; Walker, A.J.; Robertson, O.D.; Agustini, B.; Berk, M.; Dodd, S. Neurobiology and Therapeutic Potential of Cyclooxygenase-2 (COX-2) inhibitors for inflammation in neuropsychiatric disorders. Front. Psychiatry 2019, 10, 605. [Google Scholar] [CrossRef]
- Siniscalco, D.; Sapone, A.; Giordano, C.; Cirillo, A.; de Magistris, L.; Rossi, F.; Fasano, A.; Bradstreet, J.J.; Maione, S.; Antonucci, N. Cannabinoid receptor type 2, but not type 1, is up-regulated in peripheral blood mononuclear cells of children affected by autistic disorders. J. Autism Dev. Disord. 2013, 43, 686–695. [Google Scholar] [CrossRef]
- Font-Nieves, M.; Sans-Fons, M.G.; Gorina, R.; Bonfill-Teixidor, E.; Salas-Pérdomo, A.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. J Induction of COX-2 enzyme and down-regulation of COX-1 expression by lipopolysaccharide (LPS) control prostaglandin E2 production in astrocytes. Biol. Chem. 2012, 287, 6454–6468. [Google Scholar] [CrossRef] [PubMed]
- Rai-Bhogal, R.; Ahmad, E.; Li, H.; Crawford, D.A. Microarray analysis of gene expression in the cyclooxygenase knockout mice—A connection to to autism spectrum disorder. Eur. J. Neurosci. 2018, 47, 750–766. [Google Scholar] [CrossRef] [PubMed]
- El-Ansary, A.; Alhakbany, M.; Aldbass, A.; Qasem, H.; Al-Mazidi, S.; Bhat, R.S.; Al-Ayadhi, L. Alpha-Synuclein, cyclooxygenase-2 and prostaglandins-EP2 receptors as neuroinflammatory biomarkers of autism spectrum disorders: Use of combined ROC curves to increase their diagnostic values. Lipids Health Dis. 2021, 20, 55. [Google Scholar] [CrossRef]
- Komarnytsky, S.; Rathinasabapathy, T.; Wagner, C.; Metzger, B. Encocannabinoid system and its regulation by polyunsaturated fatty acids and full spectrum hemp oils. Int. J. Mol. Sci. 2021, 22, 5479. [Google Scholar] [CrossRef] [PubMed]
- Freitas, H.R.; Isaac, A.R.; Malcher-Lopes, R.; Diaz, B.L.; Trevenzoli, I.H.; De Mel Reis, R.A. Polyunsaturated fatty acids and endocannabinoids in health and disease. Nut. Neurosci. 2018, 2, 695–714. [Google Scholar] [CrossRef]
- Lu, H.C.; Mackie, K. Review of the endocannabinoid system. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef]
- Navarro, G.; Varani, K.; Reyes-Resina, I.R.; Sánchez de Medina, V.; Rivas-Santisteban, R.; Sánchez-Carnerero Callado, C.; Vincenzi, F.; Casano, S.; Ferreiro-Vera, C.; Canela, E.I.; et al. Cannabigerol action at cannabinoid CB1 and CB2 Receptors and at CB1-CB2 heteroreceptor complexes. Front. Pharmacol. 2018, 9, 632. [Google Scholar] [CrossRef]
- Garcia-Arencibia, M.; Molina-Holgado, E.; Molina-Holgado, F. Effect of endocannabinoid signalling on cell fate: Life, death, differentiation and proliferation of brain cells. Br. J. Pharmacol. 2019, 176, 1361–1369. [Google Scholar] [CrossRef]
- Wu, H.F.; Lu, T.Y.; Chu, M.C.; Chen, P.S.; Lee, C.W.; Lin, H.C. Targeting the inhibition of fatty acid amide hydrolase ameliorate the endocannabinoid-mediated synaptic dysfunction in a valproic acid-induced rat model of autism. Neuropharmacology 2020, 162, 107736. [Google Scholar] [CrossRef]
- Gomes, T.M.; Dias da Silva, D.; Carmo, H.; Carvalho, F.; Silva, J.P. Epigenetics and the endocannabinoid system signaling: An intricate interplay modulating neurodevelopment. Pharmacol. Res. 2020, 162, 105237. [Google Scholar] [CrossRef]
- Folkes, O.M.; Báldi, R.; Kondev, V.; Marcus, D.J.; Hartley, N.D.; Turner, B.D.; Ayers, J.K.; Baechle, J.J.; Misra, M.P.; Altemus, M.; et al. An endocannabinoid-regulated basolateral amygdala-nucleus accumbens circuit modulates sociability. J. Clin. Investig. 2020, 130, 728–742. [Google Scholar] [CrossRef]
- Wei, D.; Allsop, S.; Tye, K.; Piomelli, D. Endocannabinoid signaling in the control of social behavior. Trends Neurosci. 2017, 40, 385–396. [Google Scholar] [CrossRef]
- Fleury-Teixeira, P.; Caixeta, F.V.; Ramires da Silva, L.C.; Brasil-Neto, J.P.; Malcher-Lopes, R. Effects of CBD-Enriched cannabis sativa extract on autismspectrum disorder symptoms: An observational study of 18 participants undergoing compassionate use. Front. Neurol. 2019, 10, 1145. [Google Scholar] [CrossRef]
- Nachnani, R.; Raup-Konsavage, W.M.; Vrana, K.E. The pharmacological case for cannabigerol. J. Pharmacol. Exp. Ther. 2021, 376, 204–212. [Google Scholar] [CrossRef]
- Giacoppo, S.; Rajan, T.S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. The α-cyclodextrin complex of the Moringa isothiocyanate suppresses lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells through Akt and p38 inhibition. Inflamm. Res. 2017, 66, 487–503. [Google Scholar] [CrossRef]
- Ożarowski, M.; Karpiński, T.M.; Zielińska, A.; Souto, E.B.; Wielgus, K. Cannabidiol in neurological and neoplastic diseases: Latest developments on the molecular mechanism of action. Int. J. Mol. Sci. 2021, 22, 4294. [Google Scholar] [CrossRef]
- Schonhofen, P.; Bristot, I.J.; Crippa, J.A.; Hallak, J.E.C.; Zuardi, A.W.; Parsons, R.B.; Klamt, F. Cannabinoid-Based Therapies and Brain Development: Potential Harmful Effect of Early Modulation of the Endocannabinoid System. CNS Drugs 2018, 32, 697–712. [Google Scholar] [CrossRef]
- Melas, P.A.; Scherma, M.; Fratta, W.; Cifani, C.; Fadda, P. Cannabidiol as a potential treatment foranxiety and mood disorders: Molecular targets and epigenetic insights from preclinical Research. Int. J. Mol. Sci. 2021, 22, 1863. [Google Scholar] [CrossRef]
- Habib, S.S.; Al-Regaiey, K.; Bashir, S.; Iqbal, M. Role of endocannabinoids on neuroinflammation in autism spectrum disorder prevention. J. Clin. Diagn. Res. 2017, 11, CE01–CE03. [Google Scholar] [CrossRef]
- Zamberletti, E.; Gabaglio, M.; Woolley-Roberts, M.; Bingham, S.; Rubino, T.; Parolaro, D. Cannabidivarin treatment ameliorates autism-like behaviors and restores hippocampal endocannabinoid system and glia alterations induced by prenatal valproic acid exposure in rats. Front. Cell Neurosci. 2019, 13, 367. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R.; Burke, S.L.; Maddux, M. Current state of evidence of cannabis utilization for treatment of autism spectrum disorders. BMC Psychiatry 2019, 19, 328. [Google Scholar] [CrossRef]
- Aran, A.; Harel, M.; Cassuto, H.; Polyansky, L.; Schnapp, A.; Wattad, N.; Shmueli, D.; Golan, D.; Castellanos, F.X. Cannabinoid treatment for autism: A proof-of-concept randomized trial. Mol. Autism 2021, 12, 6. [Google Scholar] [CrossRef]
- Pretzsch, C.M.; Schäfer, T.; Lombardo, M.V.; Warrier, V.; Mann, C.; Bletsch, A.; Chatham, C.H.; Floris, D.L.; Tillmann, J.; Yousaf, A.; et al. Neurobiological correlates of change in adaptive behavior in autism. Am. J. Psychiatry 2022, 179, 336–349. [Google Scholar] [CrossRef]
- Fusar-Poli, L.; Cavone, V.; Tinacci, S.; Concas, I.; Petralia, A.; Signorelli, M.S.; Díaz-Caneja, C.M.; Aguglia, E. Cannabinoids for people with ASD: A systematic review of published and ongoing studies. Brain Sci. 2020, 10, 572. [Google Scholar] [CrossRef]
- Shonesy, B.C.; Parrish, W.P.; Haddad, H.K.; Stephenson, J.R.; Báldi, R.; Bluett, R.J.; Marks, C.R.; Centanni, S.W.; Folkes, O.M.; Spiess, K.; et al. Role of striatal direct pathway 2-arachidonoylglycerol signaling in sociability and repetitive behavior. Biol. Psychiatry 2018, 84, 304–315. [Google Scholar] [CrossRef]
- Zou, M.; Liu, Y.; Xie, S.; Wang, L.; Li, D.; Li, L.; Wang, F.; Zhang, Y.; Xia, W.; Sun, C.; et al. Alterations of the endocannabinoid system and its therapeutic potential in autism spectrum disorder. Open Biol. 2021, 1, 200306. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, C. Alleviation of neuropathology by inhibition of monoacylglycerol lipase in APP transgenic mice lacking CB2 receptors. Mol. Neurobiol. 2018, 55, 4802–4810. [Google Scholar] [CrossRef]
- Burgoyne, J.R.; Mongue-Din, H.; Eaton, P.; Shah, A.M. Redox signaling in cardiac physiology and pathology. Circ. Res. 2012, 111, 1091–1106. [Google Scholar] [CrossRef]
- Le Belle, J.E.; Sperry, J.; Ngo, A.; Ghochani, Y.; Laks, D.R.; López-Aranda, M.; Silva, A.J.; Kornblum, H.I. Maternal inflammation contributes to brain overgrowth and autism-associated behaviors through altered redox signaling in stem and progenitor cells. Stem Cell Reports 2014, 3, 725–734. [Google Scholar] [CrossRef]
- Taylor, J.P.; Tse, H.M. The role of oxidases in infectious and inflammatory diseases. Redox. Biol. 2021, 48, 102159. [Google Scholar] [CrossRef]
- Franco, M.C.; Carreras, M.C.; Hannibal, L. Molecular basis of redox signaling Molecular basis of redox signaling. Oxid. Med. Cell Longev. 2019, 2019, 6414975. [Google Scholar] [CrossRef]
- Fernando, V.; Zheng, X.; Walia, Y.; Sharma, V.; Letson, J.; Furuta, S.S. Nitrosylation: An emerging paradigm of redox signaling. Antioxidants 2019, 8, 404. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Lipton, S.A. S-Nitrosylation in neurogenesis and neuronal development. Biochim. Biophys. Acta 2015, 1850, 1588–1593. [Google Scholar] [CrossRef] [PubMed]
- Doenyas, C. Potential role of epigenetics and redox signaling in the gut-brain communication and the case of autism spectrum disorder. Cell Mol. Neurobiol. 2022, 42, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.S.; Costa, V.; Pereira, C. Signaling pathways governing iron homeostasis in budding yeast. Mol. Microbiol. 2018, 109, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.D.; Tan, E.K. Iron regulatory protein (IRP)-iron responsive element (IRE) signaling pathway in human neurodegenerative diseases. Mol. Neurodegener. 2017, 12, 75. [Google Scholar] [CrossRef]
- Hasan, N.M.; Lutsenko, S. Regulation of copper transporters in human cells. Curr. Top. Membr. 2012, 69, 137–161. [Google Scholar] [CrossRef]
- Patel, B.N.; Dunn, R.J.; Jeong, S.Y.; Zhu, Q.; Julien, J.P.; David, S. Ceruloplasmin regulates iron levels in the CNS and prevents free radical injury. J. Neurosci. 2002, 22, 6578–6586. [Google Scholar] [CrossRef]
- Barbariga, M.; Curnis, F.; Andolfo, A.; Zanardi, A.; Lazzaro, M.; Conti, A.; Magnani, G.; Volontè, M.A.; Ferrari, L.; Comi, G.; et al. Ceruloplasmin functional changes in Parkinson’s disease-cerebrospinal fluid. Mol. Neurodegener. 2015, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, C.M.; Chang, C.J. Copper signaling in the brain and beyond. Biol. Chem. 2018, 293, 4628–4635. [Google Scholar] [CrossRef] [Green Version]
- Kardos, J.; Héja, L.; Simon, Á.; Jablonkai, I.; Kovács, R.; Jemnitz, K. Copper signalling: Causes and consequences. Cell Commun. Signal. 2018, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Santiago González, D.A.; Cheli, V.T.; Rosenblum, S.L.; Denaroso, G.; Paez, P.M. Ceruloplasmin deletion in myelinating glial cells induces myelin disruption and oxidative stress in the central and peripheral nervous systems. Redox. Biol. 2021, 46, 102118. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Sasaki, H.; Kawasaki, Y.; Yoshihara, S. Contribution of transferrin and ceruloplasmin neurotransmission and oxidant/antioxidant status to the effects of everolimus: A Case Series. Cureus 2020, 12, e6920. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Kawasak, Y.; Yamada, H. Increased ω-3 polyunsaturated fatty acid/arachidonic acid ratios and upregulation of signaling mediator in individuals with autism spectrum disorders. Life Sci. 2016, 145, 205–212. [Google Scholar] [CrossRef]
- Yui, K.; Imataka, G.; Kawasaki, Y.; Yamada, H. Down-regulation of a signaling mediator in association with lowered plasma arachidonic acid levels in individuals with autism spectrum disorders. Neurosci. Lett. 2016, 610, 223–228. [Google Scholar] [CrossRef]
- De Giacomo, A.; Medicamento, S.; Pedac, I.C.; Giambersio, D.; Giannico, O.V.; Petruzzelli, M.G.; Simone, M.; Corsalini, M.; Marzulli, L.; Matera, E. Peripheral Iron Levels in Autism Spectrum Disorders vs. Other Neurodevelopmental Disorders: Preliminary Data. Int. J. Environ. Res. Public Health 2022, 19, 4006. [Google Scholar] [CrossRef]
- Bener, A.; Khattab, A.O.; Bhugra, D.; Hoffmann, G.F. Iron and vitamin D levels among autism spectrum disorders children. Ann. Afr. Med. 2017, 16, 186–191. [Google Scholar] [CrossRef]
- Lipina, C.; Hundal, H.S. Modulation of cellular redox homeostasis by the endocannabinoid system. Open Biol. 2016, 6, 150276. [Google Scholar] [CrossRef]
- Scalvini, L.; Vacondio, F.; Bassi, M.; Pala, D.; Lodola, A.; Rivara, S.; Jung, K.M.; Piomelli, D.; Mor, M. Free-energy studies reveal a possible mechanism for oxidation-dependent inhibition of MGL. Sci. Rep. 2016, 6, 31046. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Hackett, B.; O’Driscoll, D.N.; Sun, M.C.; Downer, J. Cannabidiol modulation of oxidative stress and signalling. Neuronal Signal. 2021, 5, NS20200080. [Google Scholar] [CrossRef] [PubMed]
- Sunda, F.; Arowolo, A. A molecular basis for the anti-inflammatory and anti-fibrosis properties of cannabidiol. FASEB J. 2020, 34, 14083–14092. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Romero, J.; Ramos, J.A. Endocannabinoids and neurodegenerative disorders: Parkinson’s disease, Huntington’s Chorea, Alzheimer’s disease, and Others. Handb. Exp. Pharmacol. 2015, 231, 233–259. [Google Scholar] [CrossRef]
- Sisk, L.M.; Rapuano, K.M.; Conley, M.I.; Greene, A.S.; Horien, C.; Rosenberg, M.D.; Scheinost, D.; Constable, R.T.; Glatt, C.E.; Casey, B.J.; et al. Genetic variation in endocannabinoid signaling is associated with differential network-level functional connectivity in youth. J. Neurosci. Res. 2021, 100, 731–741. [Google Scholar] [CrossRef]
- Meccariello, R.; Santoro, A.; D’Angelo, S.; Morrone, R.; Fasano, S.; Viggiano, A.; Pierantoni, R. The epigenetics of the endocannabinoid system. Int. J. Mol. Sci. 2020, 21, 1113. [Google Scholar] [CrossRef]
- Morris, G.; Walder, K.; Berk, M.; Carvalho, A.F.; Marx, W.; Bortolasci, C.C.; Yung, A.R.; Puri, B.K.; Maes, M. Intertwined associations between oxidative and nitrosative stress and endocannabinoid system pathways: Relevance for neuropsychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 114, 110481. [Google Scholar] [CrossRef]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.S.; Redegeld, F.A. Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef]
- Viader, A.; Blankman, J.L.; Zhong, P.; Liu, X.; Schlosburg, J.E.; Joslyn, C.M.; Liu, Q.S.; Tomarchio, A.J.; Lichtman, A.H.; Selley, D.E.; et al. Metabolic interplay between astrocytes and neurons reglates endocannabinoid action. Cell Rep. 2015, 12, 798–808. [Google Scholar] [CrossRef]
- Tanaka, M.; Sackett, S.; Zhang, Y. Endocannabinoid modulation of microglial phenotypes in neuropathology. Front. Neurol. 2020, 11, 87. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Zhang, H. Modulation of astrocytic glutamine synthetase by endocannabinoid 2-arachidonoylglycerol in JNK-independent pathway. Front. Pain. Res. (Lausanne) 2021, 2, 682051. [Google Scholar] [CrossRef]
- Sheppe, A.E.F.; Santelices, J.; Czyz, D.M.; Edelmann, M.J. Yersinia pseudotuberculosis Yop, J. Limits. Macrophage response by downregulating COX-2-mediated biosynthesis of PGE2 in a MAPK/ERK-dependent manner. Microbiol. Spectr. 2021, 9, e0049621. [Google Scholar] [CrossRef]
- Petroni, V.; Subashi, E.; Premoli, M.; Wöhr, M.; Crusio, W.E.; Lemaire, V.; Pietropaolo, S. Autistic-like behavioral effects of prenatal stress in juvenile Fmr1 mice: The relevance of sex differences and gene-environment interactions. Sci. Rep. 2022, 12, 7269. [Google Scholar] [CrossRef]
- Niu, M.; Han, Y.; Dy, A.B.C.; Du, J.; Jin, H.; Qin, J.; Zhang, J.; Li, Q.; Hagerman, R.J. Autism symptoms in Fragile X syndrome. J. Child. Neurol. 2017, 32, 903–909. [Google Scholar] [CrossRef]
- Roberts, J.E.; Ezell, J.E.; Fairchild, A.J.; Klusek, J.; Thurman, A.J.; McDuffie, A.; Abbeduto, L. Biobehavioral composite of social aspects of anxiety in young adults with fragile X syndrome contrasted to autism spectrum disorder. Am J Med Genet B Neuropsychiatr. Genet. 2018, 177, 665–675. [Google Scholar] [CrossRef]
- Liu, X.; Kumar, V.; Tsai, N.P.; Auerbach, B.D. Hyperexcitability and homeostasis in Fragile X syndrome. Front Mol Neurosci 2022, 14, 805929. [Google Scholar] [CrossRef]
- Gross, C.; Hoffmann, A.; Bassell, G.J.; Berry-Kravis, E.M. Therapeutic Strategies in Fragile X Syndrome: From Bench to Bedside and Back. Neurotherapeutics 2015, 12, 584–608. [Google Scholar] [CrossRef]
- Busquets-Garcia, A.; Gomis-González, M.; Guegan, T.; Agustín-Pavón, C.; Pastor, A.; Mato, S.; Pérez-Samartín, A.; Matute, C.; de la Torre, R.; Dierssen, M.; et al. Targeting the endocannabinoid system in the treatment of fragile X syndrome. Nat. Med. 2013, 19, 603–607. [Google Scholar] [CrossRef]
- Qin, M.; Zeider, Z.; Moulton, K.; Krych, L.; Xia, Z.; Smith, C.B. Endocannabinoid-mediated improvement on a test of aversive memory in a mouse model of fragile X syndrome. Behav. Brain. Res. 2015, 291, 164–171. [Google Scholar] [CrossRef]
- Bordeleau, M.; Fernández de Cossío, L.; Chakravarty, M.M.; Tremblay, M.È. From maternaldiet to neurodevelopmental disorders: A story of neuroinflammation. Front. Cell. Neurosci. 2021, 14, 612705. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A. Palmitoylethanolamide: A nutritional approach to keep neuroinflammation within physiological boundaries—A dystematic Review. Int. J. Mol. Sci. 2020, 21, 9526. [Google Scholar] [CrossRef]
- Hafizi, S.; Tabatabaei, D. Review of clinical studies targeting inflammatory pathways for individuals with autism. Front. Psychiatry 2019, 10, 849. [Google Scholar] [CrossRef]
- Hill, K.P.; Gold, M.S.; Nemeroff, C.B.; McDonald, W.; Grzenda, A.; Widge, A.S.; Rodriguez, C.; Kraguljac, N.V.; Krystal, J.H.; Carpenter, L.L. Risks and benefits of cannabis and cannabinoids in psychiatry. Am. J. Psychiatry 2022, 179, 98–109. [Google Scholar] [CrossRef]
- Silva, E.A.D., Jr.; Medeiros, W.M.B.; Torro, N.; Sousa, J.M.M.; Almeida, I.B.C.M.; Costa, F.B.D.; Pontes, K.M.; Nunes, E.L.G.; Rosa, M.D.D.; Albuquerque, K.L.G.D. Cannabis and cannabinoid use in autism spectrum disorder: A systematic review. Trends Psychiatry Psychother. 2022, 44, e20200149. [Google Scholar] [CrossRef]
- Barchel, D.; Stolar, O.; De-Haan, T.; Ziv-Baran, T.; Saban, N.; Fuchs, D.O.; Koren, G.; Berkovitch, M. Oral cannabidiol use in children with autism spectrum disorder to treat related symptoms and co-morbidities. Front Pharmacol 2019, 9, 1521. [Google Scholar] [CrossRef]
- Nezgovorova, V.; Ferretti, C.J.; Taylor, B.P.; Shanahan, E.; Uzunova, G.; Hong, K.; Devinsky, O.; Hollander, E. Potential of cannabinoids as treatments for autism spectrum disorders. J. Psychiatr. Res. 2021, 137, 194–201. [Google Scholar] [CrossRef]
- Bellocchio, L.; Inchingolo, A.D.; Inchingolo, A.M.; Lorusso, F.; Malcangi, G.; Santacroce, L.; Scarano, A.; Bordea, I.R.; Hazballa, D.; D’Oria, M.T.; et al. Cannabinoids drugs and oral health-from recreational side-effects to medicinal purposes: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 8329. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yui, K.; Imataka, G.; Yoshihara, S. Lipid-Based Molecules on Signaling Pathways in Autism Spectrum Disorder. Int. J. Mol. Sci. 2022, 23, 9803. https://doi.org/10.3390/ijms23179803
Yui K, Imataka G, Yoshihara S. Lipid-Based Molecules on Signaling Pathways in Autism Spectrum Disorder. International Journal of Molecular Sciences. 2022; 23(17):9803. https://doi.org/10.3390/ijms23179803
Chicago/Turabian StyleYui, Kunio, George Imataka, and Shigemi Yoshihara. 2022. "Lipid-Based Molecules on Signaling Pathways in Autism Spectrum Disorder" International Journal of Molecular Sciences 23, no. 17: 9803. https://doi.org/10.3390/ijms23179803
APA StyleYui, K., Imataka, G., & Yoshihara, S. (2022). Lipid-Based Molecules on Signaling Pathways in Autism Spectrum Disorder. International Journal of Molecular Sciences, 23(17), 9803. https://doi.org/10.3390/ijms23179803