
Human Amnion-Derived Mesenchymal Stromal Cells: A New Potential Treatment for Carbapenem-Resistant Enterobacterales in Decompensated Cirrhosis

 ,
,  ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
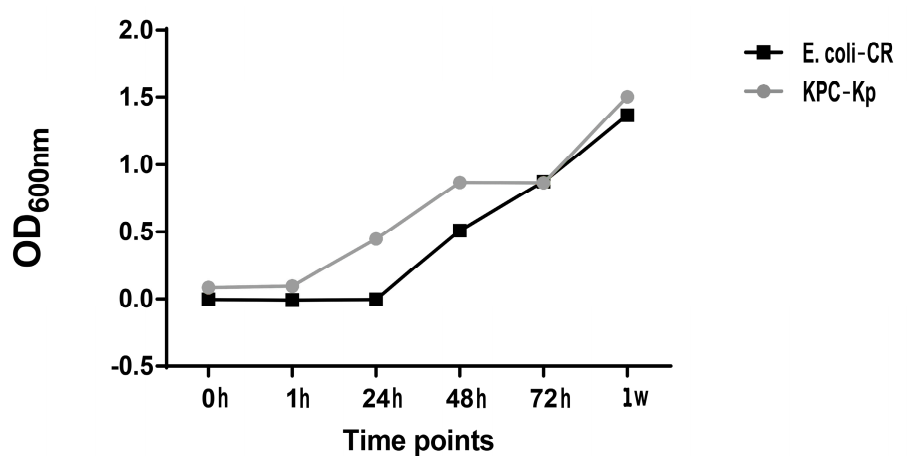
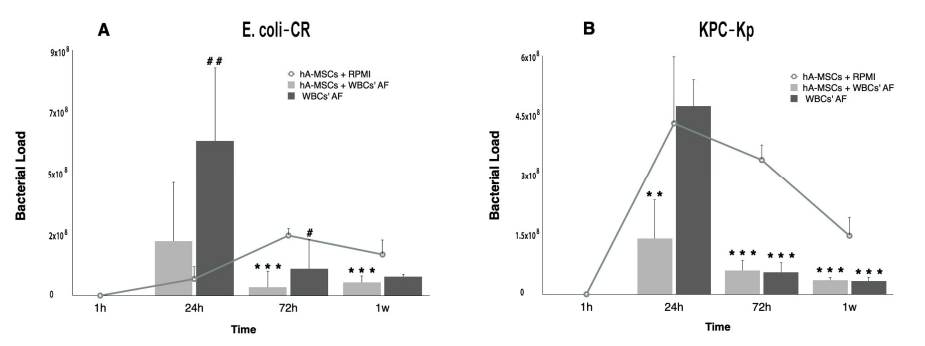
2.1. Determination of Bacterial Load after hA-MSCs Co-Culture Shows a Better Trend in AF Infected with Carbapenem-Resistant Enterobacterales
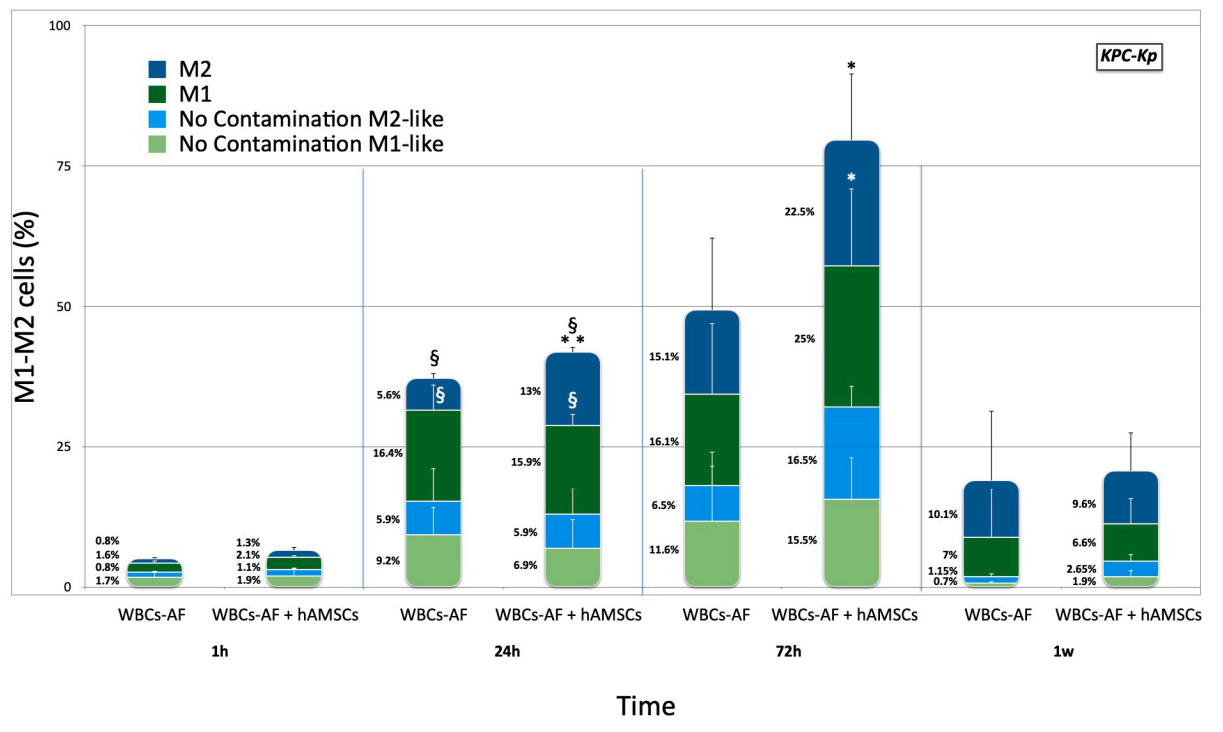
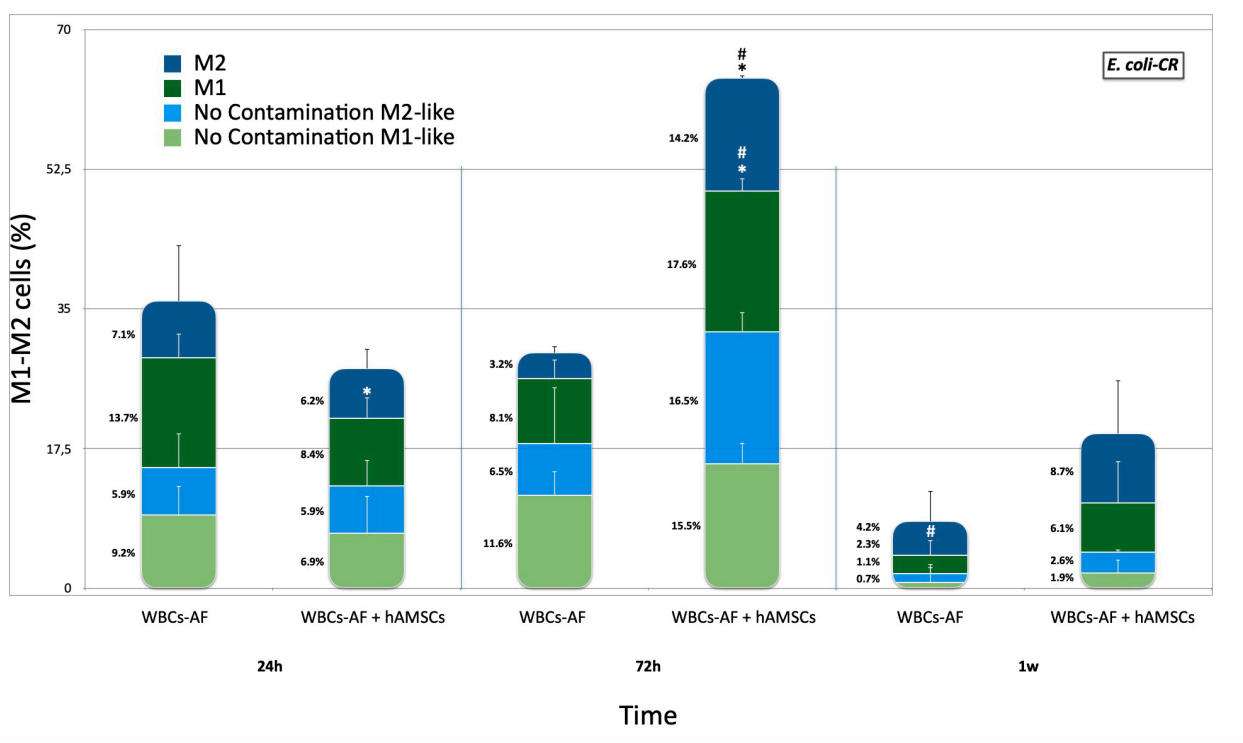
2.2. Effects of hA-MSCs on Macrophage Phenotypes in Presence of Carbapenem-Resistant Enterobacterales
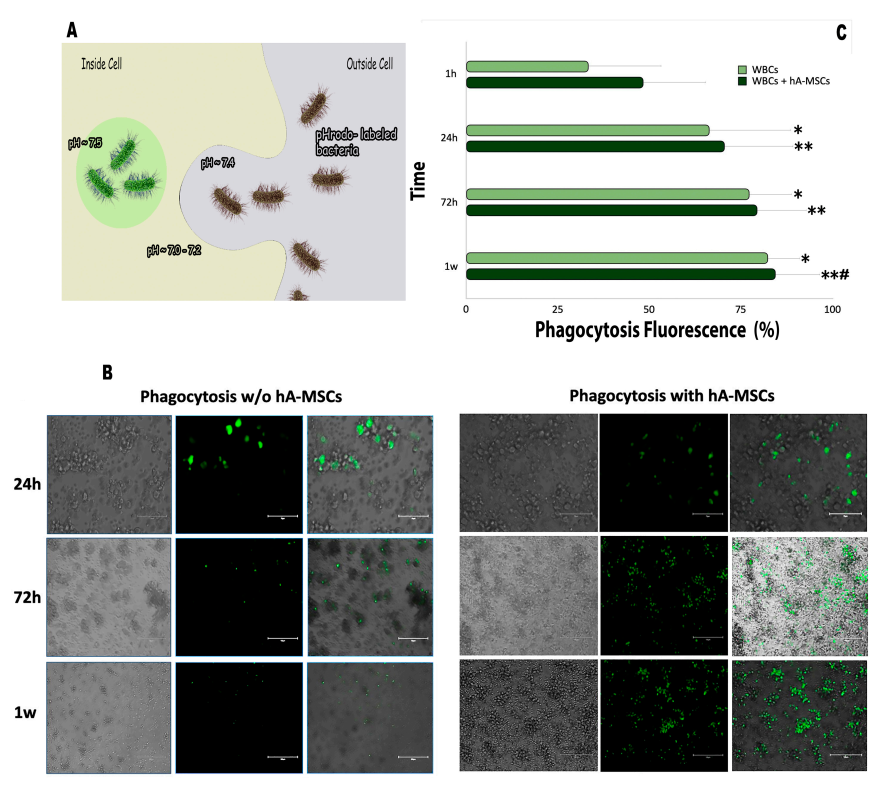
2.3. Phagocytic Capacity of the WBCs’ AF Component in Co-Culture with hA-MSCs
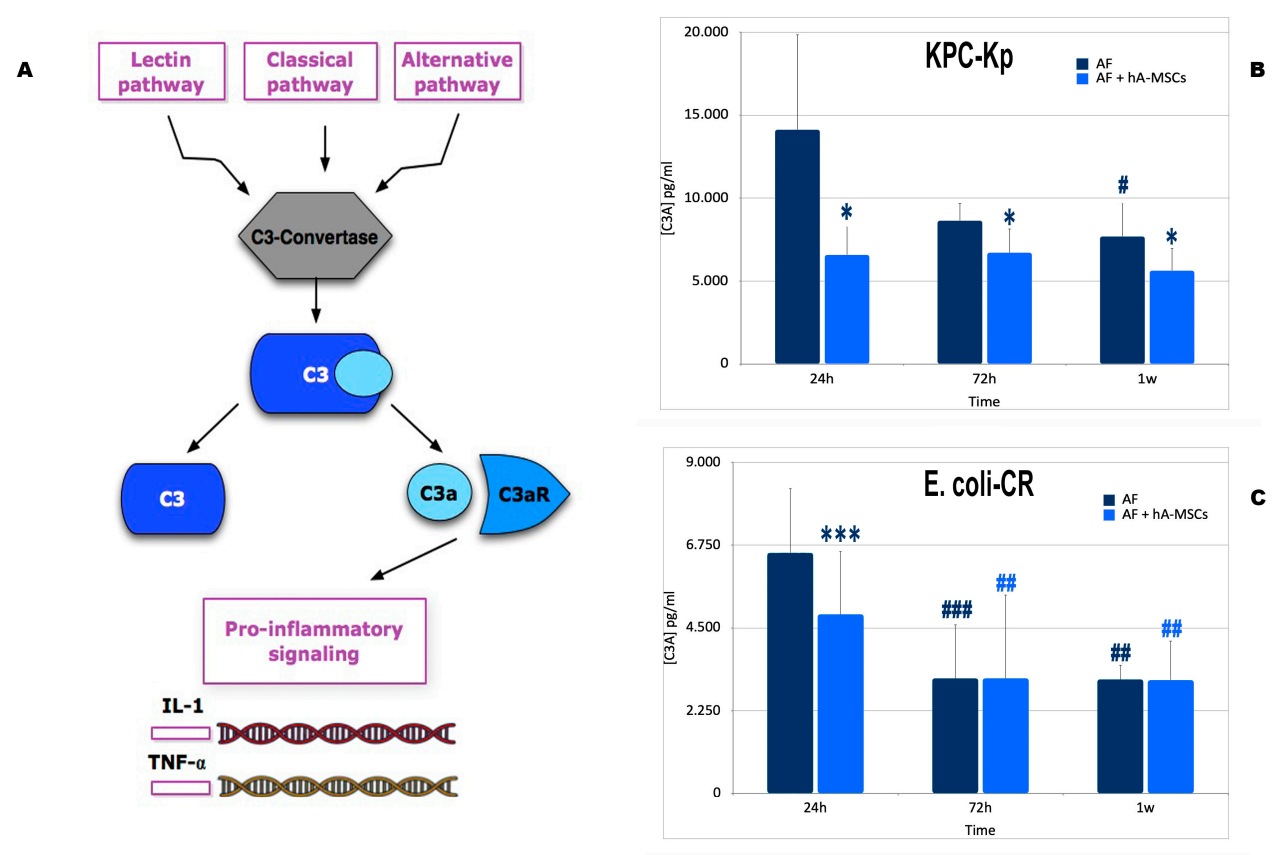
2.4. Mesenchymal Stromal Cells Induce Greater Activation of the Complement System with a Lower Formation of Pro-Inflammatory Molecules
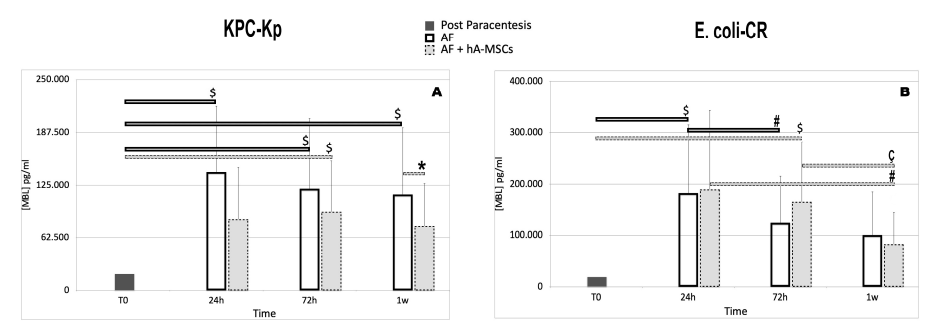
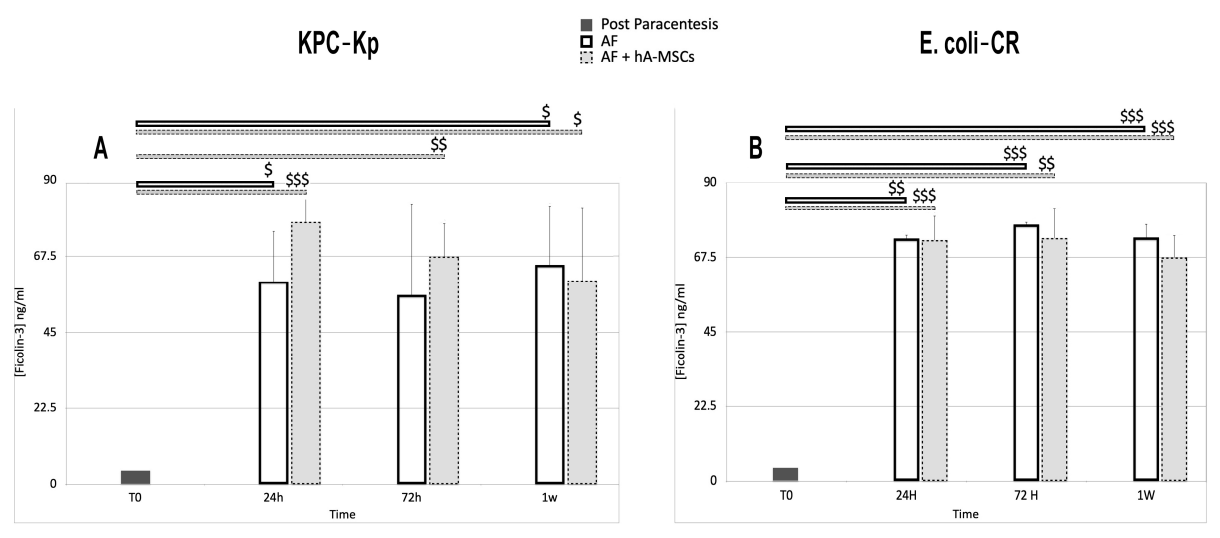
2.5. Levels of Pattern Recognition Molecules (PRMs), Mannan-Binding Lectin (MBL) and Ficolins (FNCs)
3. Discussion
4. Materials and Methods
4.1. Patients and Ascitic Fluid Collection
4.2. Isolation and Culture of Human Amnion-Derived Mesenchymal Stromal Cells (hA-MSCs)
4.3. Bacterial Strains
4.4. Exposure of hA-MSCs to Infected Ascitic Fluid
4.4.1. Analyses of Macrophage Polarization WBCs from Infected Ascetic Fluid in Co-Culture with hA-MSCs
4.4.2. Evaluation of Bacterial Load in Ascitic Fluid
4.4.3. Phagocytosis Assay
4.4.4. Assessment of the Rate of C3a and Mannose Binding Lectin
4.4.5. Ficolin-3 and Ficolin-2 Enzyme-Linked Immunosorbent Assay
5. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Tsao, G. Bacterial translocation: Cause or consequence of decompensation in cirrhosis? J. Hepatol. 2001, 34, 150–155. [Google Scholar] [CrossRef]
- Wiest, R.; Garcia-Tsao, G. Bacterial translocation (BT) in cirrhosis. Hepatology 2005, 41, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Northup, P.G.; Intagliata, N.M.; Shah, N.L.; Pelletier, S.J.; Berg, C.L.; Argo, C.K. Excess mortality on the liver transplant waiting list: Unintended policy consequences and Model for End-Stage Liver Disease (MELD) inflation. Hepatology 2015, 61, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Yu, Y.; Chen, M.; Wang, C.; Kang, Y.; Lou, J. Causative agents and outcome of spontaneous bacterial peritonitis in cirrhotic patients: Community-acquired versus nosocomial infections. BMC Infect. Dis. 2019, 19, 463. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Prado, V.; Trebicka, J.; Amoros, A.; Gustot, T.; Wiest, R.; Deulofeu, C.; Garcia, E.; Acevedo, J.; Fuhrmann, V.; et al. Multidrug-resistant bacterial infections in patients with decompensated cirrhosis and with acute-on-chronic liver failure in Europe. J. Hepatol. 2019, 70, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Lutz, P.; Jeffery, H.C.; Jones, N.; Birtwistle, J.; Kramer, B.; Nattermann, J.; Spengler, U.; Strassburg, C.P.; Adams, D.H.; Oo, Y.H. NK Cells in Ascites from Liver Disease Patients Display a Particular Phenotype and Take Part in Antibacterial Immune Response. Front. Immunol. 2019, 10, 1838. [Google Scholar] [CrossRef] [Green Version]
- Lung, T.; Sakem, B.; Risch, L.; Wurzner, R.; Colucci, G.; Cerny, A.; Nydegger, U. The complement system in liver diseases: Evidence-based approach and therapeutic options. J. Transl. Autoimmun. 2019, 2, 100017. [Google Scholar] [CrossRef]
- Oladimeji, A.A.; Temi, A.P.; Adekunle, A.E.; Taiwo, R.H.; Ayokunle, D.S. Prevalence of spontaneous bacterial peritonitis in liver cirrhosis with ascites. Pan Afr. Med. J. 2013, 15, 128. [Google Scholar]
- Irvine, K.M.; Ratnasekera, I.; Powell, E.E.; Hume, D.A. Causes and Consequences of Innate Immune Dysfunction in Cirrhosis. Front. Immunol. 2019, 10, 293. [Google Scholar] [CrossRef]
- Glargaard, S.; Boysen, T.; Pilely, K.; Garred, P.; Ytting, H. Prognostic value of lectin pathway molecules and complement proteins in ascitic fluid and blood in patients with liver cirrhosis. Scand. J. Gastroenterol. 2018, 53, 64–69. [Google Scholar] [CrossRef]
- Garred, P.; Genster, N.; Pilely, K.; Bayarri-Olmos, R.; Rosbjerg, A.; Ma, Y.J.; Skjoedt, M.O. A journey through the lectin pathway of complement-MBL and beyond. Immunol. Rev. 2016, 274, 74–97. [Google Scholar] [CrossRef]
- Foldi, I.; Tornai, T.; Tornai, D.; Sipeki, N.; Vitalis, Z.; Tornai, I.; Dinya, T.; Antal-Szalmas, P.; Papp, M. Lectin-complement pathway molecules are decreased in patients with cirrhosis and constitute the risk of bacterial infections. Liver Int. 2017, 37, 1023–1031. [Google Scholar] [CrossRef] [Green Version]
- Pampalone, M.; Corrao, S.; Amico, G.; Vitale, G.; Alduino, R.; Conaldi, P.G.; Pietrosi, G. Human Amnion-Derived Mesenchymal Stromal Cells in Cirrhotic Patients with Refractory Ascites: A Possible Anti-Inflammatory Therapy for Preventing Spontaneous Bacterial Peritonitis. Stem Cell Rev. Rep. 2021, 17, 981–998. [Google Scholar] [CrossRef]
- Netea, M.G.; Dominguez-Andres, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Wu, W.; Millman, A.; Craft, J.F.; Chen, E.; Patel, N.; Boucher, J.L.; Urban, J.F., Jr.; Kim, C.C.; Gause, W.C. Neutrophils prime a long-lived effector macrophage phenotype that mediates accelerated helminth expulsion. Nat. Immunol. 2014, 15, 938–946. [Google Scholar] [CrossRef] [Green Version]
- Magatti, M.; Vertua, E.; De Munari, S.; Caro, M.; Caruso, M.; Silini, A.; Delgado, M.; Parolini, O. Human amnion favours tissue repair by inducing the M1-to-M2 switch and enhancing M2 macrophage features. J. Tissue Eng. Regen. Med. 2017, 11, 2895–2911. [Google Scholar] [CrossRef]
- Malyshev, I.; Malyshev, Y. Current Concept and Update of the Macrophage Plasticity Concept: Intracellular Mechanisms of Reprogramming and M3 Macrophage “Switch” Phenotype. Biomed. Res. Int. 2015, 2015, 341308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of macrophage polarization in autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Sa Pessoa, J. Klebsiella pneumoniae infection biology: Living to counteract host defences. FEMS Microbiol. Rev. 2019, 43, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Schraufstatter, I.U.; Khaldoyanidi, S.K.; DiScipio, R.G. Complement activation in the context of stem cells and tissue repair. World J. Stem Cells 2015, 7, 1090–1108. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.Q.; Lambris, J.D.; Ricklin, D. Protection of host cells by complement regulators. Immunol. Rev. 2016, 274, 152–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubbers, R.; van Essen, M.F.; van Kooten, C.; Trouw, L.A. Production of complement components by cells of the immune system. Clin. Exp. Immunol. 2017, 188, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Wong, E.K.S.; Kavanagh, D. Diseases of complement dysregulation-an overview. Semin. Immunopathol. 2018, 40, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Ricklin, D.; Reis, E.S.; Mastellos, D.C.; Gros, P.; Lambris, J.D. Complement component C3—The “Swiss Army Knife” of innate immunity and host defense. Immunol. Rev. 2016, 274, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Reis, E.S.; Mastellos, D.C.; Hajishengallis, G.; Lambris, J.D. New insights into the immune functions of complement. Nat. Rev. Immunol. 2019, 19, 503–516. [Google Scholar] [CrossRef]
- Dobo, J.; Kocsis, A.; Gal, P. Be on Target: Strategies of Targeting Alternative and Lectin Pathway Components in Complement-Mediated Diseases. Front. Immunol. 2018, 9, 1851. [Google Scholar] [CrossRef]
- Ray, T.D.; Mekasha, S.; Liang, Y.; Lu, B.; Ram, S.; Ingalls, R.R. Species-specific differences in regulation of macrophage inflammation by the C3a-C3a receptor axis. Innate Immun. 2018, 24, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Beltrame, M.H.; Boldt, A.B.; Catarino, S.J.; Mendes, H.C.; Boschmann, S.E.; Goeldner, I.; Messias-Reason, I. MBL-associated serine proteases (MASPs) and infectious diseases. Mol. Immunol. 2015, 67, 85–100. [Google Scholar] [CrossRef]
- Rothfuchs, A.G.; Roffe, E.; Gibson, A.; Cheever, A.W.; Ezekowitz, R.A.; Takahashi, K.; Steindel, M.; Sher, A.; Bafica, A. Mannose-binding lectin regulates host resistance and pathology during experimental infection with Trypanosoma cruzi. PLoS ONE 2012, 7, e47835. [Google Scholar]
- Sastry, K.; Zahedi, K.; Lelias, J.M.; Whitehead, A.S.; Ezekowitz, R.A. Molecular characterization of the mouse mannose-binding proteins. The mannose-binding protein A but not C is an acute phase reactant. J. Immunol. 1991, 147, 692–697. [Google Scholar]
- Liu, H.; Jensen, L.; Hansen, S.; Petersen, S.V.; Takahashi, K.; Ezekowitz, A.B.; Hansen, F.D.; Jensenius, J.C.; Thiel, S. Characterization and quantification of mouse mannan-binding lectins (MBL-A and MBL-C) and study of acute phase responses. Scand. J. Immunol. 2001, 53, 489–497. [Google Scholar] [CrossRef]
- Hayashi, M.; Abe, K.; Fujita, M.; Takahashi, A.; Sekine, H.; Ohira, H. Association between serum ficolin-1 level and disease progression in primary biliary cholangitis. PLoS ONE 2020, 15, e0238300. [Google Scholar] [CrossRef]
- Genster, N.; Ostrup, O.; Schjalm, C.; Eirik Mollnes, T.; Cowland, J.B.; Garred, P. Ficolins do not alter host immune responses to lipopolysaccharide-induced inflammation in vivo. Sci. Rep. 2017, 7, 3852. [Google Scholar] [CrossRef] [Green Version]
- Rueschenbaum, S.; Ciesek, S.; Queck, A.; Widera, M.; Schwarzkopf, K.; Brune, B.; Welsch, C.; Wedemeyer, H.; Zeuzem, S.; Weigert, A.; et al. Dysregulated Adaptive Immunity Is an Early Event in Liver Cirrhosis Preceding Acute-on-Chronic Liver Failure. Front. Immunol. 2020, 11, 534731. [Google Scholar] [CrossRef]
- Runyon, B.A.; Morrissey, R.L.; Hoefs, J.C.; Wyle, F.A. Opsonic activity of human ascitic fluid: A potentially important protective mechanism against spontaneous bacterial peritonitis. Hepatology 1985, 5, 634–637. [Google Scholar] [CrossRef]
- Marciano, S.; Diaz, J.M.; Dirchwolf, M.; Gadano, A. Spontaneous bacterial peritonitis in patients with cirrhosis: Incidence, outcomes, and treatment strategies. Hepatic Med. Evid. Res. 2019, 11, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Navasa, M.; Planas, R.; Montoliu, S.; Monfort, D.; Soriano, G.; Vila, C.; Pardo, A.; Quintero, E.; Vargas, V.; et al. Primary prophylaxis of spontaneous bacterial peritonitis delays hepatorenal syndrome and improves survival in cirrhosis. Gastroenterology 2007, 133, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Piano, S.; Singh, V.; Caraceni, P.; Maiwall, R.; Alessandria, C.; Fernandez, J.; Soares, E.C.; Kim, D.J.; Kim, S.E.; Marino, M.; et al. Epidemiology and Effects of Bacterial Infections in Patients with Cirrhosis Worldwide. Gastroenterology 2019, 156, 1368–1380.e10. [Google Scholar] [CrossRef] [Green Version]
- Chow, L.; Johnson, V.; Impastato, R.; Coy, J.; Strumpf, A.; Dow, S. Antibacterial activity of human mesenchymal stem cells mediated directly by constitutively secreted factors and indirectly by activation of innate immune effector cells. Stem Cells Transl. Med. 2020, 9, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Wong, F.; Piano, S.; Singh, V.; Bartoletti, M.; Maiwall, R.; Alessandria, C.; Fernandez, J.; Soares, E.C.; Kim, D.J.; Kim, S.E.; et al. Clinical features and evolution of bacterial infection-related acute-on-chronic liver failure. J. Hepatol. 2021, 74, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, L.Y.; Garcia-Tsao, G. A Historical Overview of Spontaneous Bacterial Peritonitis: From Rare to Resistant. Clin. Liver Dis. 2021, 18 (Suppl. 1), 63–75. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, Y.; Zhang, L.; Zhang, F.; Li, L. The Application of Mesenchymal Stem Cells in the Treatment of Liver Diseases: Mechanism, Efficacy, and Safety Issues. Front. Med. 2021, 8, 655268. [Google Scholar] [CrossRef]
- You, J.; Fu, Z.; Zou, L. Mechanism and Potential of Extracellular Vesicles Derived from Mesenchymal Stem Cells for the Treatment of Infectious Diseases. Front. Microbiol. 2021, 12, 761338. [Google Scholar] [CrossRef]
- Varkouhi, A.K.; Jerkic, M.; Ormesher, L.; Gagnon, S.; Goyal, S.; Rabani, R.; Masterson, C.; Spring, C.; Chen, P.Z.; Gu, F.X.; et al. Extracellular Vesicles from Interferon-gamma-primed Human Umbilical Cord Mesenchymal Stromal Cells Reduce Escherichia coli-induced Acute Lung Injury in Rats. Anesthesiology 2019, 130, 778–790. [Google Scholar] [CrossRef]
- Hao, Q.; Gudapati, V.; Monsel, A.; Park, J.H.; Hu, S.; Kato, H.; Lee, J.H.; Zhou, L.; He, H.; Lee, J.W. Mesenchymal Stem Cell-Derived Extracellular Vesicles Decrease Lung Injury in Mice. J. Immunol. 2019, 203, 1961–1972. [Google Scholar] [CrossRef]
- Marrazzo, P.; Crupi, A.N.; Alviano, F.; Teodori, L.; Bonsi, L. Exploring the roles of MSCs in infections: Focus on bacterial diseases. J. Mol. Med. 2019, 97, 437–450. [Google Scholar] [CrossRef]
- Hosseiniyan Khatibi, S.M.; Kheyrolahzadeh, K.; Barzegari, A.; Rahbar Saadat, Y.; Zununi Vahed, S. Medicinal signaling cells: A potential antimicrobial drug store. J. Cell Physiol. 2020, 235, 7731–7746. [Google Scholar] [CrossRef]
- Bhalerao, A.; Raut, S.; Noorani, B.; Mancuso, S.; Cucullo, L. Molecular Mechanisms of Multi-Organ Failure in COVID-19 and Potential of Stem Cell Therapy. Cells 2021, 10, 2878. [Google Scholar] [CrossRef]
- Pietrosi, G.; Fernandez-Iglesias, A.; Pampalone, M.; Ortega-Ribera, M.; Lozano, J.J.; Garcia-Caldero, H.; Abad-Jorda, L.; Conaldi, P.G.; Parolini, O.; Vizzini, G.; et al. Human amniotic stem cells improve hepatic microvascular dysfunction and portal hypertension in cirrhotic rats. Liver Int. 2020, 40, 2500–2514. [Google Scholar] [CrossRef]
- Komolafe, O.; Roberts, D.; Freeman, S.C.; Wilson, P.; Sutton, A.J.; Cooper, N.J.; Pavlov, C.S.; Milne, E.J.; Hawkins, N.; Cowlin, M.; et al. Antibiotic prophylaxis to prevent spontaneous bacterial peritonitis in people with liver cirrhosis: A network meta-analysis. Cochrane Database Syst. Rev. 2020, 1, CD013125. [Google Scholar] [CrossRef] [Green Version]
- Garred, P.; Tenner, A.J.; Mollnes, T.E. Therapeutic Targeting of the Complement System: From Rare Diseases to Pandemics. Pharmacol. Rev. 2021, 73, 792–827. [Google Scholar] [CrossRef]
- Kalia, N.; Singh, J.; Kaur, M. The ambiguous role of mannose-binding lectin (MBL) in human immunity. Open Med. 2021, 16, 299–310. [Google Scholar] [CrossRef]
- Bulati, M.; Busa, R.; Carcione, C.; Iannolo, G.; Di Mento, G.; Cuscino, N.; Di Gesu, R.; Piccionello, A.P.; Buscemi, S.; Carreca, A.P.; et al. Klebsiella pneumoniae Lipopolysaccharides Serotype O2afg Induce Poor Inflammatory Immune Responses Ex Vivo. Microorganisms 2021, 9, 1317. [Google Scholar] [CrossRef]
- Monaco, F.; Mento, G.D.; Cuscino, N.; Conaldi, P.G.; Douradinha, B. Infant colonisation with Escherichia coli and Klebsiella pneumoniae strains co-harbouring blaOXA-48 and blaNDM-1 carbapenemases genes: A case report. Int. J. Antimicrob. Agents 2018, 52, 121–122. [Google Scholar] [CrossRef]
- Metzger, M.L.; Michelfelder, I.; Goldacker, S.; Melkaoui, K.; Litzman, J.; Guzman, D.; Grimbacher, B.; Salzer, U. Low ficolin-2 levels in common variable immunodeficiency patients with bronchiectasis. Clin. Exp. Immunol. 2015, 179, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Man-Kupisinska, A.; Michalski, M.; Maciejewska, A.; Swierzko, A.S.; Cedzynski, M.; Lugowski, C.; Lukasiewicz, J. A New Ligand-Based Method for Purifying Active Human Plasma-Derived Ficolin-3 Complexes Supports the Phenomenon of Crosstalk between Pattern-Recognition Molecules and Immunoglobulins. PLoS ONE 2016, 11, e0156691. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Clone | Isotype | Dilution | Conjugation | Manufacturer |
|---|---|---|---|---|---|
| CD14 | MφP9 | Mouse BALB/c IgG2b, κ | 1:20 | APC Cy7 | BD Biosciences, CA, USA |
| CD16 | B73.1 | Mouse BALB/c IgG1, κ | 1:20 | PE | BD Biosciences, CA, USA |
| CD206 | 19.2 | Mouse IgG1, κ | 1:20 | APC | BD Biosciences, CA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pampalone, M.; Vitale, G.; Gruttadauria, S.; Amico, G.; Iannolo, G.; Douradinha, B.; Mularoni, A.; Conaldi, P.G.; Pietrosi, G. Human Amnion-Derived Mesenchymal Stromal Cells: A New Potential Treatment for Carbapenem-Resistant Enterobacterales in Decompensated Cirrhosis. Int. J. Mol. Sci. 2022, 23, 857. https://doi.org/10.3390/ijms23020857
Pampalone M, Vitale G, Gruttadauria S, Amico G, Iannolo G, Douradinha B, Mularoni A, Conaldi PG, Pietrosi G. Human Amnion-Derived Mesenchymal Stromal Cells: A New Potential Treatment for Carbapenem-Resistant Enterobacterales in Decompensated Cirrhosis. International Journal of Molecular Sciences. 2022; 23(2):857. https://doi.org/10.3390/ijms23020857
Chicago/Turabian StylePampalone, Mariangela, Giampiero Vitale, Salvatore Gruttadauria, Giandomenico Amico, Gioacchin Iannolo, Bruno Douradinha, Alessandra Mularoni, Pier Giulio Conaldi, and Giada Pietrosi. 2022. "Human Amnion-Derived Mesenchymal Stromal Cells: A New Potential Treatment for Carbapenem-Resistant Enterobacterales in Decompensated Cirrhosis" International Journal of Molecular Sciences 23, no. 2: 857. https://doi.org/10.3390/ijms23020857
APA StylePampalone, M., Vitale, G., Gruttadauria, S., Amico, G., Iannolo, G., Douradinha, B., Mularoni, A., Conaldi, P. G., & Pietrosi, G. (2022). Human Amnion-Derived Mesenchymal Stromal Cells: A New Potential Treatment for Carbapenem-Resistant Enterobacterales in Decompensated Cirrhosis. International Journal of Molecular Sciences, 23(2), 857. https://doi.org/10.3390/ijms23020857