
The Small GTPase FgRab1 Plays Indispensable Roles in the Vegetative Growth, Vesicle Fusion, Autophagy and Pathogenicity of Fusarium graminearum

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
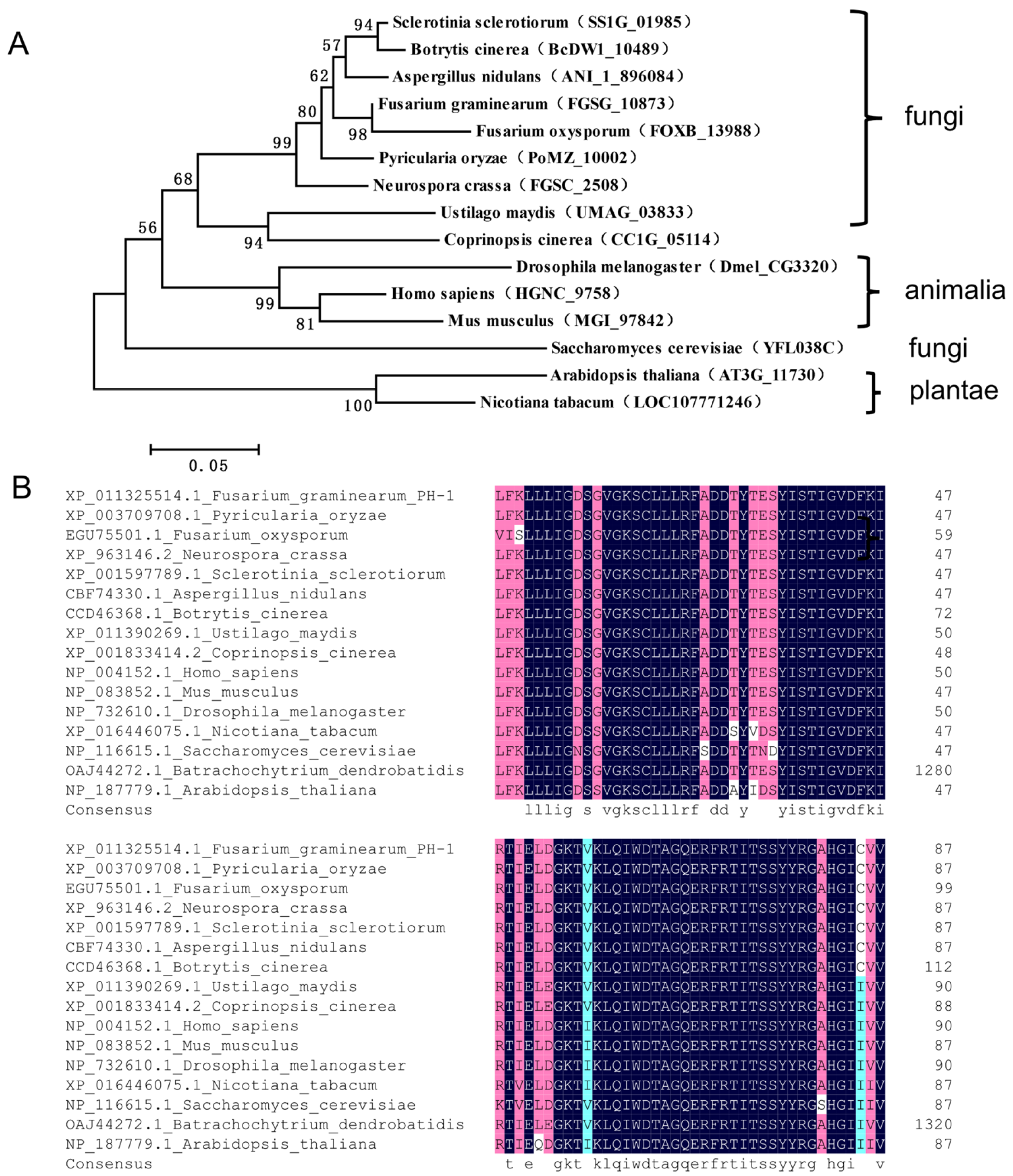
2.1. Identification of FgRab1 in F. graminearum
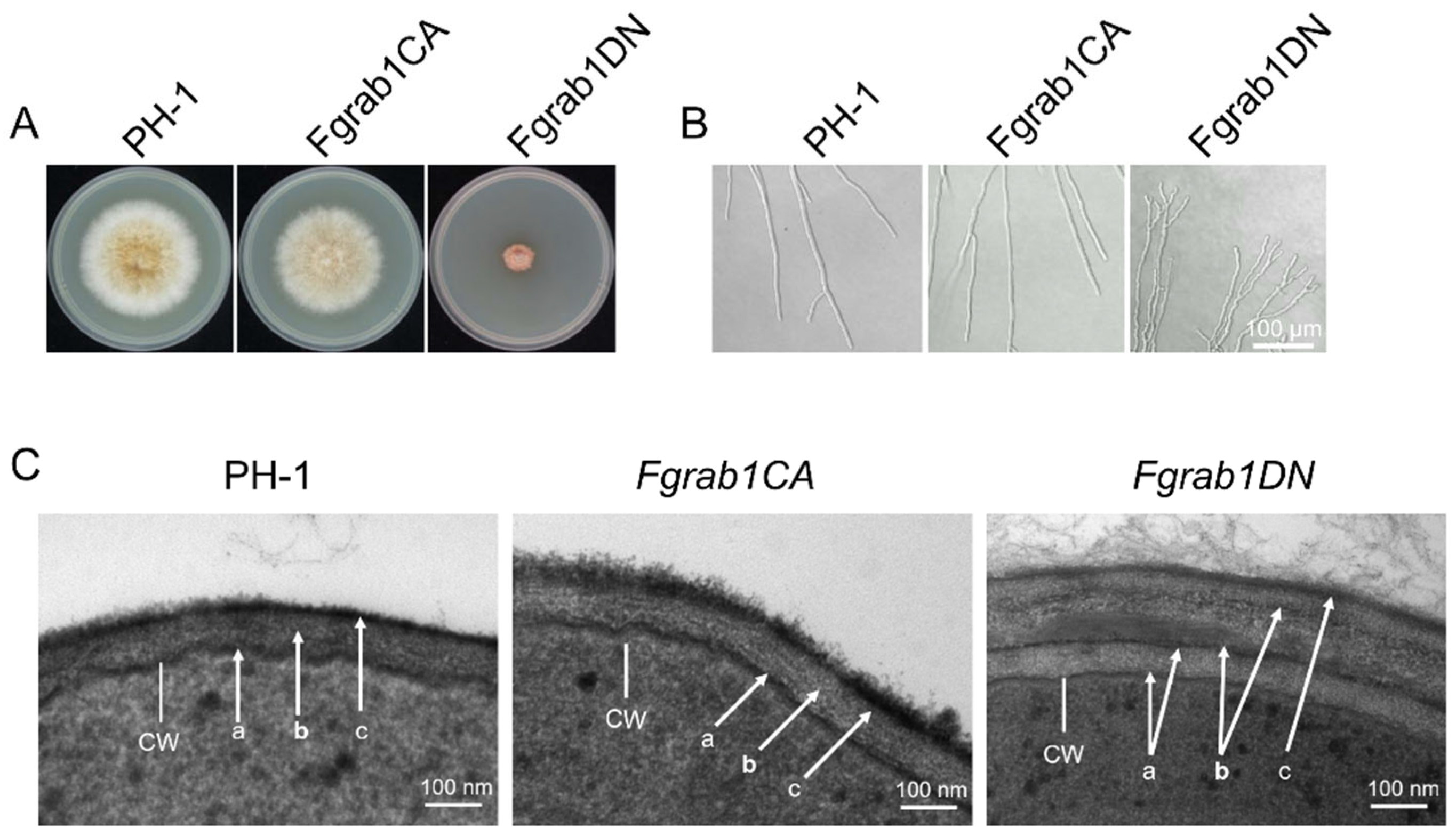
2.2. FgRab1 Is Critical for Vegetative Growth and Is Involved in the Maintenance of Cell Wall Integrity of F. graminearum
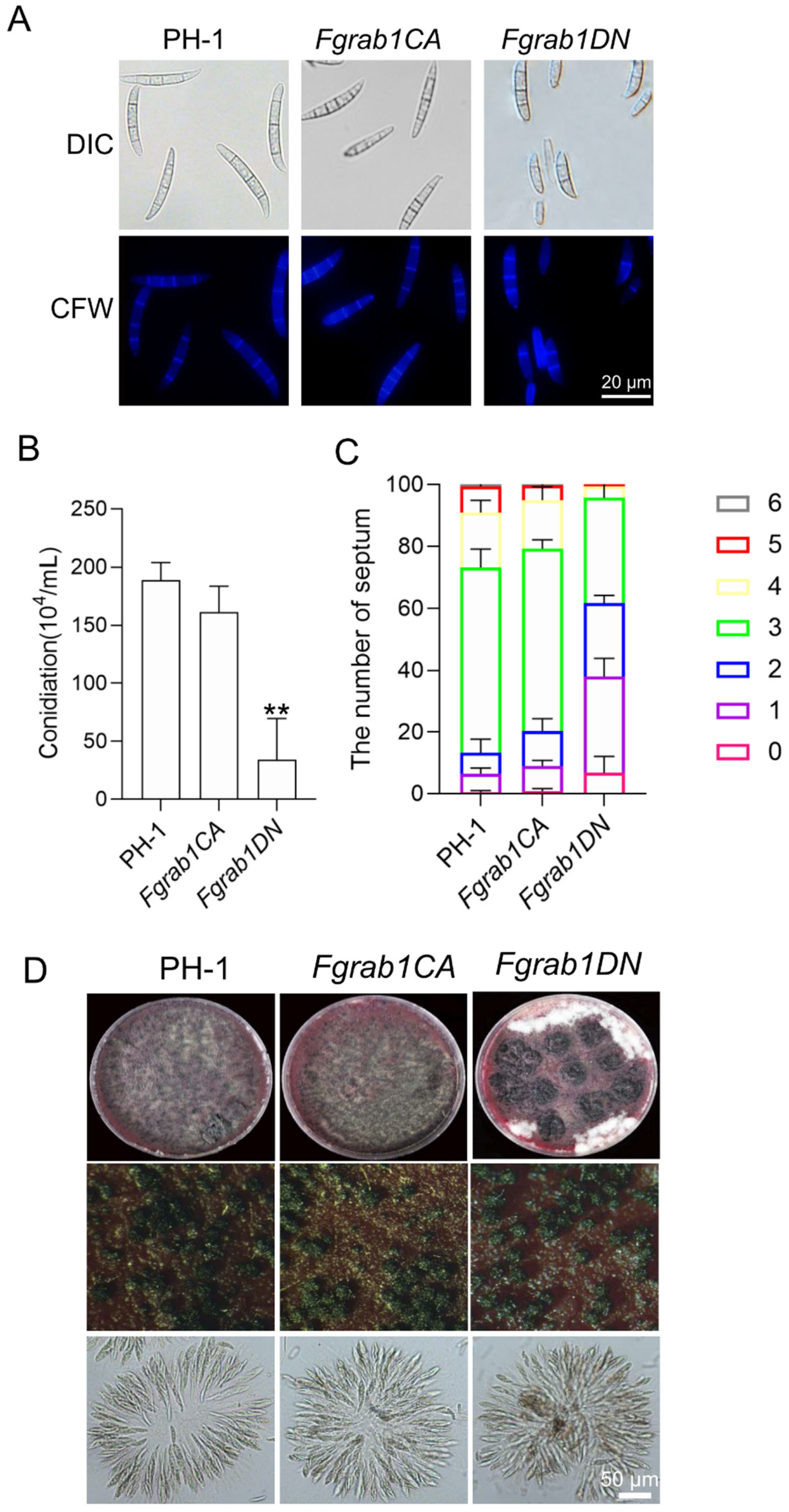
2.3. FgRab1 Is Involved in Regulating Conidiogenesis but Not Sexual Reproduction in F. graminearum
2.4. FgRab1 Localizes to the Golgi, ER and Spitzenkörper
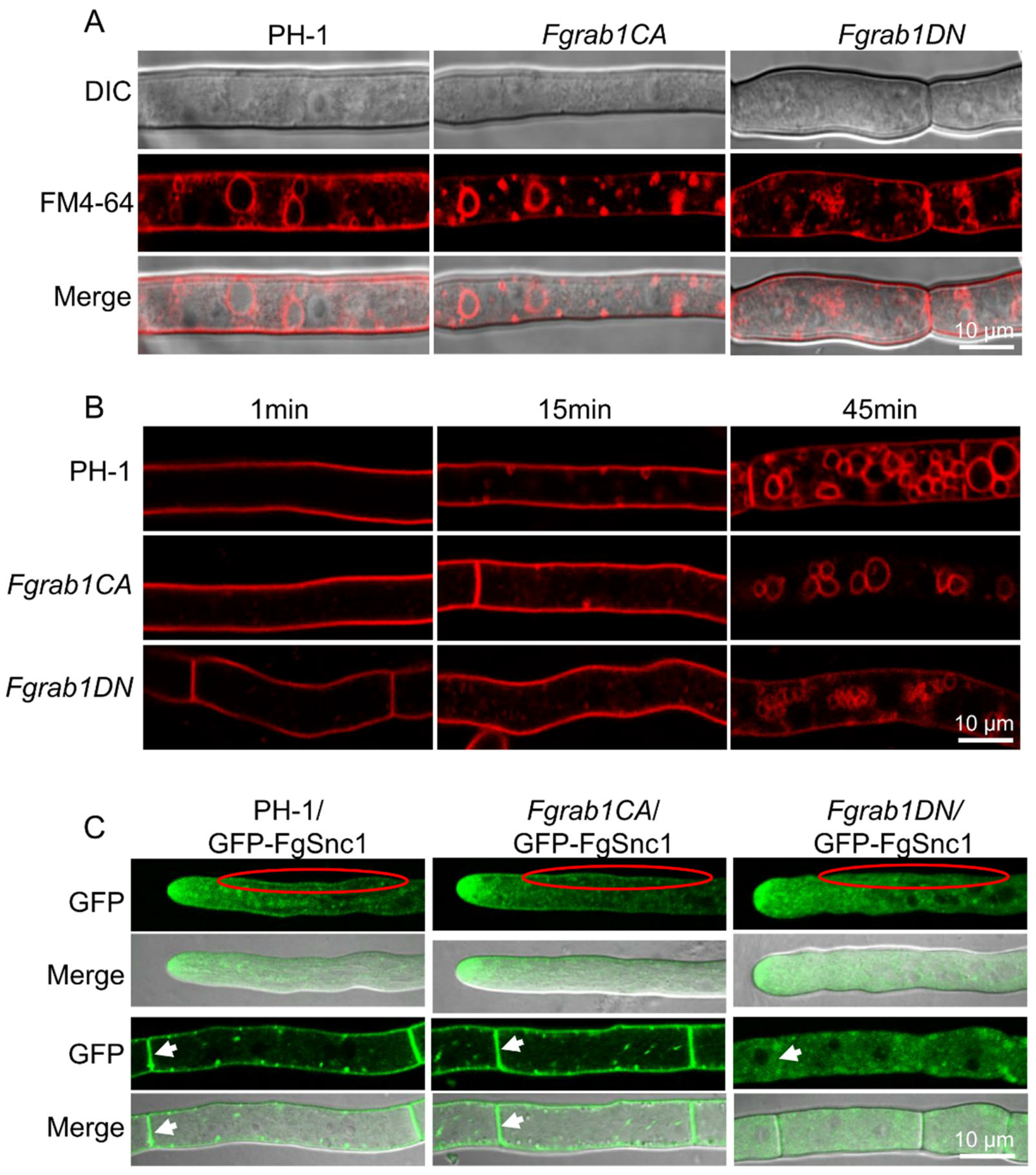
2.5. FgRab1 Is Required for Vesicle Trafficking and Fusion Process in F. graminearum
2.6. FgRab1 Is Required for Pathogenicity
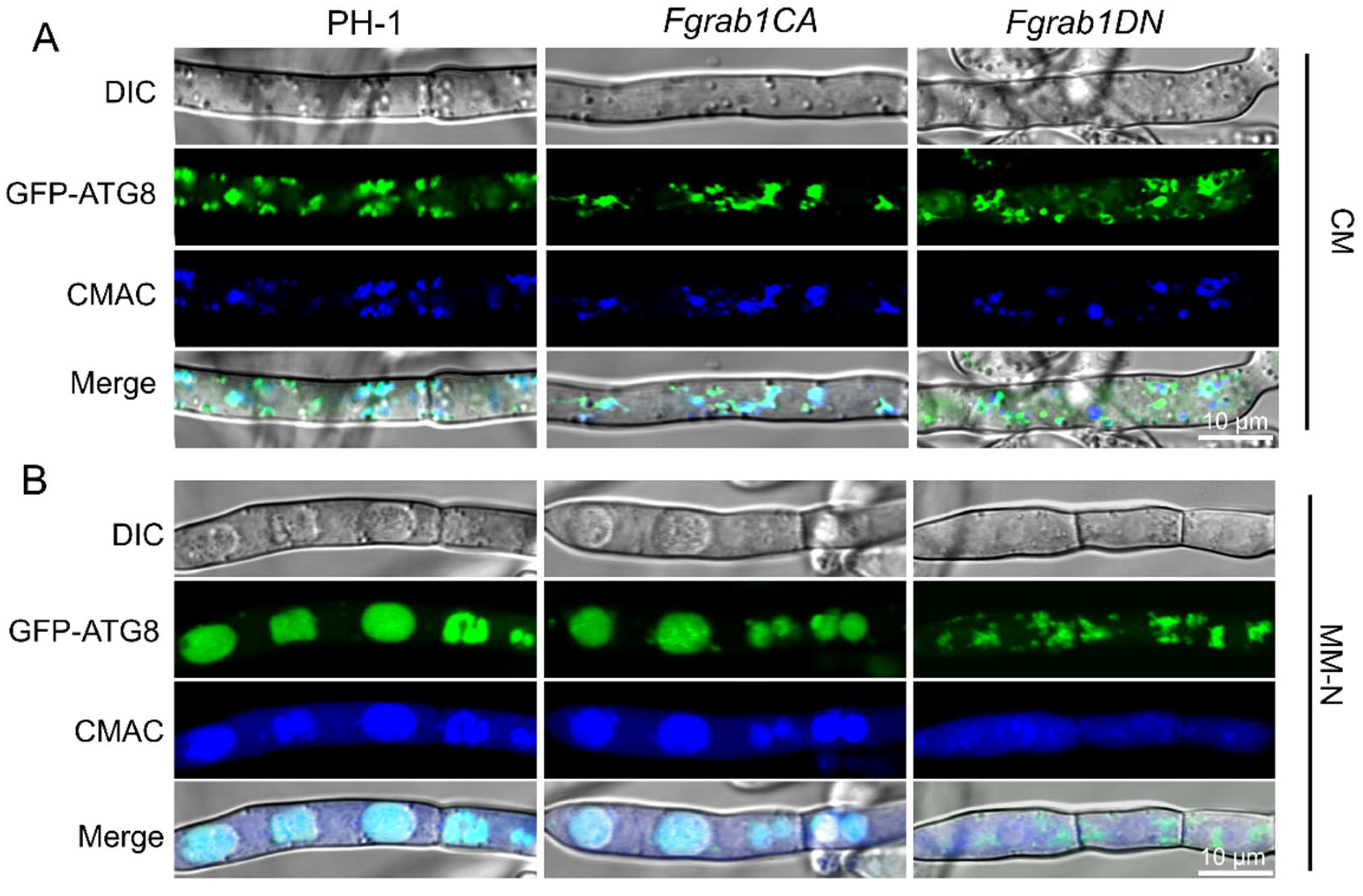
2.7. Inactivation of FgRab1 Prevents Autophagy in F. graminearum
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Culture Conditions
4.2. Construction of Constitutively Active and Dominant-Negative Mutants
4.3. Quantitative Real-Time PCR
4.4. Asexual and Sexual Reproduction Assays
4.5. Pathogenicity Assays
4.6. Autophagy Assay
4.7. Live-Cell Imaging of F. graminearum
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinar, M.; Peñalva, M.A. The fungal RABOME: RAB GTPases acting in the endocytic and exocytic pathways of Aspergillus nidulans (with excursions to other filamentous fungi). Mol. Microbiol. 2021, 116, 53–70. [Google Scholar] [CrossRef] [PubMed]
- Mizuno-Yamasaki, E.; Rivera-Molina, F.; Novick, P. GTPase Networks in Membrane Traffic. Annu. Rev. Biochem. 2012, 81, 637–659. [Google Scholar] [CrossRef] [Green Version]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Marlin, M.C. Rab family of GTPases. Methods Mol. Biol. 2015, 1298, 1–15. [Google Scholar]
- Dautt-Castro, M.; Rosendo-Vargas, M.; Casas-Flores, S. The Small GTPases in Fungal Signaling Conservation and Function. Cells 2021, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: Conserved structure and molecular mechanism. Nature 1991, 349, 117–127. [Google Scholar] [CrossRef]
- Pereira-Leal, J.B. The Ypt/Rab family and the evolution of trafficking in fungi. Traffic 2008, 9, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lei, M.; Zhou, Y.; Chen, F. A Comprehensive Analysis of the Small GTPases Ypt7 Involved in the Regulation of Fungal Development and Secondary Metabolism in Monascus ruber M7. Front. Microbiol. 2019, 10, 452. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zheng, W.; Wu, C.; Yang, J.; Xi, Y.; Xie, Q.; Zhao, X.; Deng, X.; Lu, G.; Li, G.; et al. Rab GTPases are essential for membrane trafficking-dependent growth and pathogenicity in Fusarium graminearum. Environ. Microbiol. 2015, 17, 4580–4599. [Google Scholar] [CrossRef]
- Irieda, H.; Maeda, H.; Akiyama, K.; Hagiwara, A.; Saitoh, H.; Uemura, A.; Terauchi, R.; Takano, Y. Colletotrichum orbiculare Secretes Virulence Effectors to a Biotrophic Interface at the Primary Hyphal Neck via Exocytosis Coupled with SEC22-Mediated Traffic. Plant Cell 2014, 26, 2265–2281. [Google Scholar] [CrossRef] [Green Version]
- Bielska, E.; Higuchi, Y.; Schuster, M.; Steinberg, N.; Kilaru, S.; Talbot, N.J.; Steinberg, G. Long-distance endosome trafficking drives fungal effector production during plant infection. Nat. Commun. 2014, 5, 5097. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.; Qin, G.Z.; Li, B.Q.; Tian, S.P. Knocking Out Bcsas1 in Botrytis cinerea Impacts Growth, Development, and Secretion of Extracellular Proteins, Which Decreases Virulence. Mol. Plant Microbe Interact. 2014, 27, 590–600. [Google Scholar] [CrossRef]
- Yan, H.; Huang, J.; Zhang, H.; Shim, W.B. A Rab GTPase protein FvSec4 is necessary for fumonisin B1 biosynthesis and virulence in Fusarium verticillioides. Curr. Genet. 2020, 66, 205–216. [Google Scholar] [CrossRef]
- Yang, C.D.; Dang, X.; Zheng, H.W.; Chen, X.F.; Lin, X.L.; Zhang, D.M.; Abubakar, Y.S.; Chen, X.; Lu, G.; Wang, Z.; et al. Two Rab5 Homologs Are Essential for the Development and Pathogenicity of the Rice Blast Fungus Magnaporthe oryzae. Front. Plant. Sci. 2017, 8, 620. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Marlin, M.C.; Liang, Z.; Berry, W.L.; Janknecht, R.; Zhou, J.; Wang, Z.; Lu, G.; Li, G. Distinct Biochemical and Functional Properties of Two Rab5 Homologs from the Rice Blast Fungus Magnaporthe oryzae. J. Biol. Chem. 2014, 289, 28299–282309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.H.; Chen, S.M.; Gao, H.M.; Ning, G.A.; Shi, H.B.; Wang, Y.; Dong, B.; Qi, Y.Y.; Zhang, D.M.; Lu, G.D.; et al. The small GTPase MoYpt7 is required for membrane fusion in autophagy and pathogenicity of Magnaporthe oryzae. Environ. Microbiol. 2015, 17, 4495–4510. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lin, Y.; Zheng, H.; Abubakar, Y.S.; Peng, M.; Li, J.; Yu, Z.; Wang, Z.; Naqvi, N.I.; Li, G.; et al. The retromer CSC subcomplex is recruited by MoYpt7 and sequentially sorted by MoVps17 for effective conidiation and pathogenicity of the rice blast fungus. Mol. Plant Pathol. 2021, 22, 284–298. [Google Scholar] [CrossRef]
- Zheng, H.; Chen, S.; Chen, X.; Liu, S.; Dang, X.; Yang, C.; Giraldo, M.C.; Oliveira-Garcia, E.; Zhou, J.; Wang, Z.; et al. The Small GTPase MoSec4 Is Involved in Vegetative Development and Pathogenicity by Regulating the Extracellular Protein Secretion in Magnaporthe oryzae. Front. Plant. Sci. 2016, 7, 1458. [Google Scholar] [CrossRef] [Green Version]
- Siriputthaiwan, P.; Jauneau, A.; Herbert, C.; Garcin, D.; Dumas, B. Functional analysis of CLPT1, a Rab/GTPase required for protein secretion and pathogenesis in the plant fungal pathogen Colletotrichum lindemuthianum. J. Cell Sci. 2005, 118, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, H.D.; Puzicha, M.; Gallwitz, D. Study of a temperature-sensitive mutant of the ras-related YPT1 gene product in yeast suggests a role in the regulation of intracellular calcium. Cell 1988, 53, 635–647. [Google Scholar] [CrossRef]
- Schmitt, H.D.; Wagner, P.; Pfaff, E.; Gallwitz, D. The ras-related YPT1 gene product in yeast: A GTP-binding protein that might be involved in microtubule organization. Cell 1986, 47, 401–412. [Google Scholar] [CrossRef]
- Segev, N.; Botstein, D. The ras-like yeast YPT1 gene is itself essential for growth, sporulation, and starvation response. Mol. Cell. Biol. 1987, 7, 2367–2377. [Google Scholar]
- Bacon, R.A.; Salminen, A.; Ruohola, H.; Novick, P.; Ferro-Novick, S. The GTP-binding protein Ypt1 is required for transport in vitro: The Golgi apparatus is defective in ypt1 mutants. J. Cell Biol. 1989, 109, 1015–1022. [Google Scholar] [CrossRef]
- Miyake, S.; Yamamoto, M. Identification of ras-related, YPT family genes in Schizosaccharomyces pombe. EMBO J. 1990, 9, 1417–1422. [Google Scholar] [CrossRef]
- Lee, S.A.; Mao, Y.; Zhang, Z.; Wong, B. Overexpression of a dominant-negative allele of YPT1 inhibits growth and aspartyl protease secretion in Candida albicans. Microbiology 2001, 147, 1961–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pind, S.N.; Nuoffer, C.; McCaffery, J.M.; Plutner, H.; Davidson, H.W.; Farquhar, M.G.; Balch, W.E. Rab1 and Ca2+ are required for the fusion of carrier vesicles mediating endoplasmic reticulum to Golgi transport. J. Cell Biol. 1994, 125, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, W.; Hazen, B.; Pritsch, C.; Leonard, K. Histology and physiology of Fusarium head blight. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; APS Press: St. Paul, MN, USA, 2003; pp. 44–83. [Google Scholar]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Rusu, A.; Barrett, L.; Hunter, G.C.; Kazan, K. Can natural gene drives be part of future fungal pathogen control strategies in plants? New Phytol. 2020, 228, 1431–1439. [Google Scholar] [CrossRef]
- Lofgren, L.A.; LeBlanc, N.R.; Certano, A.K.; Nachtigall, J.; LaBine, K.M.; Riddle, J.; Broz, K.; Dong, Y.; Bethan, B.; Kafer, C.W.; et al. Fusarium graminearum: Pathogen or endophyte of North American grasses? New Phytol. 2018, 217, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Bönnighausen, J.; Schauer, N.; Schäfer, W.; Bormann, J. Metabolic profiling of wheat rachis node infection by Fusarium graminearum—Decoding deoxynivalenol-dependent susceptibility. New Phytol. 2019, 221, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Zhang, Y.K.; Kremling, K.A.; Ding, Y.; Bennett, J.S.; Bae, J.S.; Kim, D.K.; Ackerman, H.H.; Kolomiets, M.V.; Schmelz, E.A.; et al. Ethylene signaling regulates natural variation in the abundance of antifungal acetylated diferuloylsucroses and Fusarium graminearum resistance in maize seedling roots. New Phytol. 2019, 221, 2096–2111. [Google Scholar] [CrossRef]
- Boddu, J.; Cho, S.; Kruger, W.M.; Muehlbauer, G.J. Transcriptome analysis of the barley-Fusarium graminearum interaction. Mol. Plant Microbe Interact 2006, 19, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Van der Lee, T.; Yang, X.; Yu, D.; Waalwijk, C. Fusarium populations on Chinese barley show a dramatic gradient in mycotoxin profiles. Phytopathology 2008, 98, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, J.; Jia, L.J.; Yuan, T.L.; Zhang, D.; Guo, Y.; Wang, Y.; Tang, W.H. Cellular Tracking and Gene Profiling of Fusarium graminearum during Maize Stalk Rot Disease Development Elucidates Its Strategies in Confronting Phosphorus Limitation in the Host Apoplast. PLoS Pathog. 2016, 12, e1005485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Li, J.; Chen, X.; Zhang, X.; Liao, D.; Yun, Y.; Zheng, W.; Abubakar, Y.S.; Li, G.; Wang, Z.; et al. FgVps9, a Rab5 GEF, Is Critical for DON Biosynthesis and Pathogenicity in Fusarium graminearum. Front Microbiol 2020, 11, 1714. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, B.; Liu, L.; Chen, H.; Zhang, H.; Zheng, X.; Zhang, Z. FgMon1, a guanine nucleotide exchange factor of FgRab7, is important for vacuole fusion, autophagy and plant infection in Fusarium graminearum. Sci. Rep. 2015, 5, 18101. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Li, L.; Miao, P.; Wu, C.; Chen, X.; Yuan, M.; Fang, T.; Norvienyeku, J.; Li, G.; Zheng, W.; et al. FgSec2A, a guanine nucleotide exchange factor of FgRab8, is important for polarized growth, pathogenicity and deoxynivalenol production in Fusarium graminearum. Environ Microbiol 2018, 20, 3378–3392. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Li, L.; Yu, Z.; Yuan, Y.; Zheng, Q.; Xie, Q.; Li, G.; Abubakar, Y.S.; Zhou, J.; Wang, Z.; et al. FgSpa2 recruits FgMsb3, a Rab8 GAP, to the polarisome to regulate polarized trafficking, growth and pathogenicity in Fusarium graminearum. New Phytol. 2021, 229, 1665–1683. [Google Scholar] [CrossRef]
- Zheng, Q.; Yu, Z.; Yuan, Y.; Sun, D.; Abubakar, Y.S.; Zhou, J.; Wang, Z.; Zheng, H. The GTPase-Activating Protein FgGyp1 Is Important for Vegetative Growth, Conidiation, and Virulence and Negatively Regulates DON Biosynthesis in Fusarium graminearum. Front. Microbiol. 2021, 12, 621519. [Google Scholar] [CrossRef]
- Pereira-Leal, J.B.; Seabra, M.C. The mammalian Rab family of small GTPases: Definition of family and subfamily sequence motifs suggests a mechanism for functional specificity in the Ras superfamily. J. Mol. Biol. 2000, 301, 1077–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira-Leal, J.B.; Strom, M.; Godfrey, R.F.; Seabra, M.C. Structural determinants of Rab and Rab Escort Protein interaction: Rab family motifs define a conserved binding surface. Biochem. Biophys. Res. Commun. 2003, 301, 92–97. [Google Scholar] [CrossRef]
- Ali, B.R.; Wasmeier, C.; Lamoreux, L.; Strom, M.; Seabra, M.C. Multiple regions contribute to membrane targeting of Rab GTPases. J. Cell Sci. 2004, 117, 6401–6412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Miao, P.; Lin, X.; Li, L.; Wu, C.; Chen, X.; Abubakar, Y.S.; Norvienyeku, J.; Li, G.; Zhou, J.; et al. Small GTPase Rab7-mediated FgAtg9 trafficking is essential for autophagy-dependent development and pathogenicity in Fusarium graminearum. PLoS Genet. 2018, 14, e1007546. [Google Scholar] [CrossRef]
- Josefsen, L.; Droce, A.; Sondergaard, T.E.; Sorensen, J.L.; Bormann, J.; Schafer, W.; Giese, H.; Olsson, S. Autophagy provides nutrients for nonassimilating fungal structures and is necessary for plant colonization but not for infection in the necrotrophic plant pathogen Fusarium graminearum. Autophagy 2012, 8, 326–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Chen, L.; Li, W.; Zhang, L.; Zhang, L.; Zou, S.; Liang, Y.; Yu, J.; Dong, H. Endocytic FgEde1 regulates virulence and autophagy in Fusarium graminearum. Fungal Genet. Biol. 2020, 141, 103400. [Google Scholar] [CrossRef]
- Thomas, L.L.; Fromme, J.C. Extensive GTPase crosstalk regulates Golgi trafficking and maturation. Curr. Opin. Cell Biol. 2020, 65, 1–7. [Google Scholar] [CrossRef]
- Yang, X.Z.; Li, X.X.; Zhang, Y.J.; Rodriguez-Rodriguez, L.; Xiang, M.Q.; Wang, H.Y.; Zheng, X.F. Rab1 in cell signaling, cancer and other diseases. Oncogene 2016, 35, 5699–5704. [Google Scholar] [CrossRef] [Green Version]
- Laura, M.; Westrate, M.J.H.; Nash, M.J.; Voeltz, G.K. Vesicular and uncoated Rab1-dependent cargo carriers facilitate ER to Golgi transport. J. Cell Sci. 2020, 133, jcs239814. [Google Scholar]
- Sechi, S.; Frappaolo, A.; Fraschini, R.; Capalbo, L.; Gottardo, M.; Belloni, G.; Glover, D.M.; Wainman, A.; Giansanti, M.G. Rab1 interacts with GOLPH3 and controls Golgi structure and contractile ring constriction during cytokinesis in Drosophila melanogaster. Open Biol. 2017, 7, 160257. [Google Scholar] [CrossRef] [PubMed]
- Thomas Weide, M.K.; Barnekow, A. Inactive and active mutants of rab1b are not tightly integrated into target membranes. Int. J. Oncol. 1999, 15, 727–736. [Google Scholar]
- Best, J.T.; Xu, P.; McGuire, J.G.; Leahy, S.N.; Graham, T.R. Yeast synaptobrevin, Snc1, engages distinct routes of postendocytic recycling mediated by a sorting nexin, Rcy1-COPI, and retromer. Mol. Biol. Cell 2020, 31, 944–962. [Google Scholar] [CrossRef]
- Zheng, W.; Lin, Y.; Fang, W.; Zhao, X.; Lou, Y.; Wang, G.; Zheng, H.; Liang, Q.; Abubakar, Y.S.; Olsson, S.; et al. The endosomal recycling of FgSnc1 by FgSnx41-FgSnx4 heterodimer is essential for polarized growth and pathogenicity in Fusarium graminearum. New Phytol. 2018, 219, 654–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, N.; Dumur, C.I.; Martinez, H.; Garcia, I.A.; Monetta, P.; Slavin, I.; Sampieri, L.; Koritschoner, N.; Mironov, A.A.; De Matteis, M.A.; et al. Rab1b overexpression modifies Golgi size and gene expression in HeLa cells and modulates the thyrotrophin response in thyroid cells in culture. Mol. Biol. Cell 2013, 24, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.; Garcia-Mata, R.; Brandon, E.; Sztul, E. COPI recruitment is modulated by a Rab1b-dependent mechanism. Mol. Biol. Cell 2003, 14, 2116–2127. [Google Scholar] [CrossRef] [Green Version]
- Campoy, E.M.; Zoppino, F.C.; Colombo, M.I. The early secretory pathway contributes to the growth of the Coxiella-replicative niche. Infect. Immun. 2011, 79, 402–413. [Google Scholar] [CrossRef] [Green Version]
- Pérez González, P.R.P.; Sergio, A. Rho GTPases: Regulation of cell polarity and growth in yeasts. Biochem. J. 2010, 426, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, X.; Zou, L.; Wu, Y. Regulation of autophagy by the Rab GTPase network. Cell Death Differ. 2014, 21, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.H.; Gao, H.M.; Xu, F.; Lu, J.P.; Devenish, R.J.; Lin, F.C. Autophagy vitalizes the pathogenicity of pathogenic fungi. Autophagy 2012, 8, 1415–1425. [Google Scholar] [CrossRef]
- Mitter, A.L.; Schlotterhose, P.; Krick, R. Gyp1 has a dual function as Ypt1 GAP and interaction partner of Atg8 in selective autophagy. Autophagy 2019, 15, 1031–1050. [Google Scholar] [CrossRef] [PubMed]
- Du, L.L.; Novick, P. Yeast rab GTPase-activating protein Gyp1p localizes to the Golgi apparatus and is a negative regulator of Ypt1p. Mol. Biol. Cell 2001, 12, 1215–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joiner, A.M.; Phillips, B.P.; Yugandhar, K.; Sanford, E.J.; Smolka, M.B.; Yu, H.; Miller, E.A.; Fromme, J.C. Structural basis of TRAPPIII-mediated Rab1 activation. EMBO J. 2021, 40, e107607. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Zhang, M.; Li, J.; Yang, C.; Abubakar, Y.S.; Chen, X.; Zheng, W.; Wang, Z.; Zheng, H.; Zhou, J. The Small GTPase FgRab1 Plays Indispensable Roles in the Vegetative Growth, Vesicle Fusion, Autophagy and Pathogenicity of Fusarium graminearum. Int. J. Mol. Sci. 2022, 23, 895. https://doi.org/10.3390/ijms23020895
Yuan Y, Zhang M, Li J, Yang C, Abubakar YS, Chen X, Zheng W, Wang Z, Zheng H, Zhou J. The Small GTPase FgRab1 Plays Indispensable Roles in the Vegetative Growth, Vesicle Fusion, Autophagy and Pathogenicity of Fusarium graminearum. International Journal of Molecular Sciences. 2022; 23(2):895. https://doi.org/10.3390/ijms23020895
Chicago/Turabian StyleYuan, Yanping, Meiru Zhang, Jingjing Li, Chengdong Yang, Yakubu Saddeeq Abubakar, Xin Chen, Wenhui Zheng, Zonghua Wang, Huawei Zheng, and Jie Zhou. 2022. "The Small GTPase FgRab1 Plays Indispensable Roles in the Vegetative Growth, Vesicle Fusion, Autophagy and Pathogenicity of Fusarium graminearum" International Journal of Molecular Sciences 23, no. 2: 895. https://doi.org/10.3390/ijms23020895