
Identification of Low-Nitrogen-Related miRNAs and Their Target Genes in Sugarcane and the Role of miR156 in Nitrogen Assimilation

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
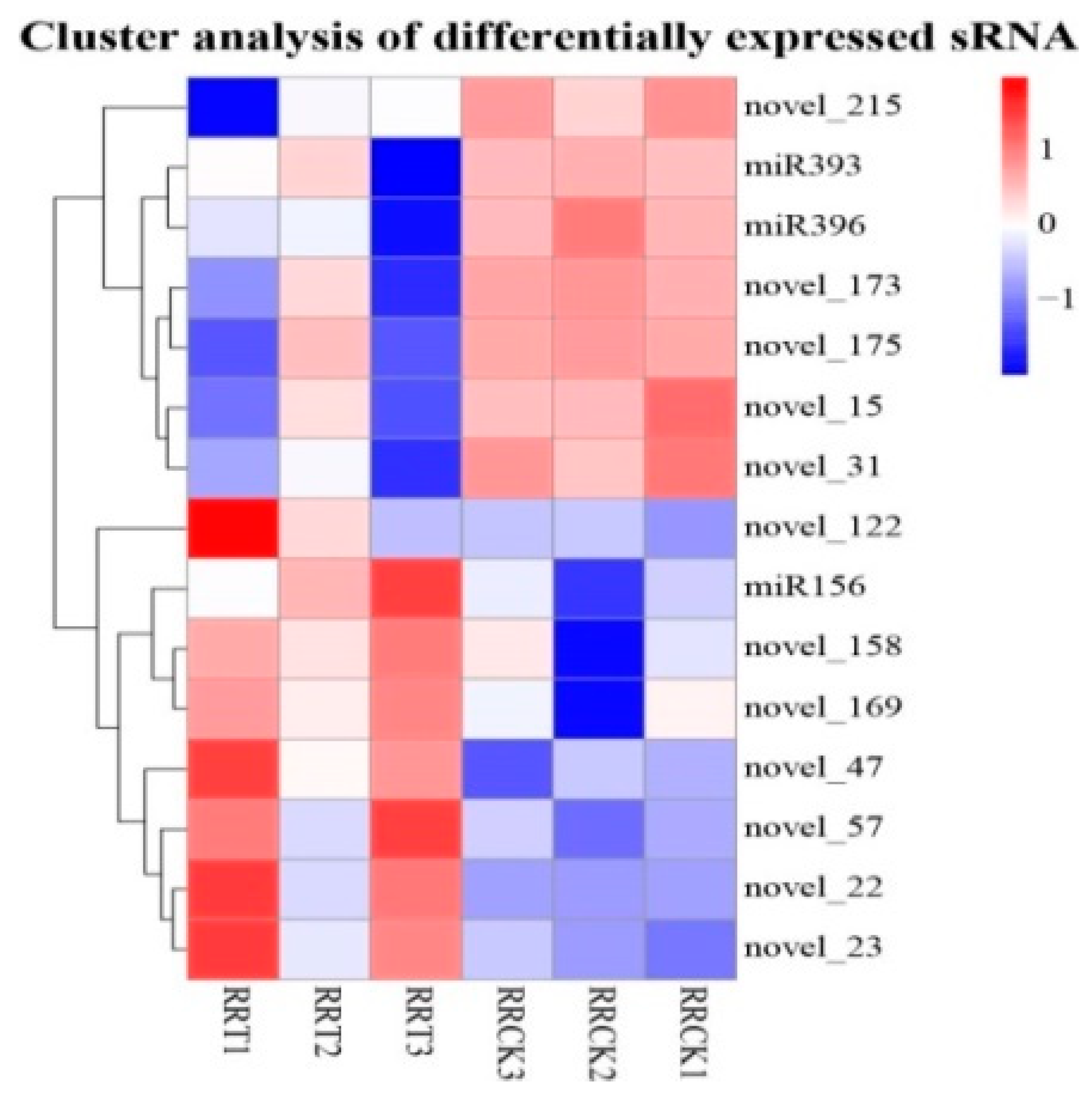
2.1. sRNA Sequencing Data
2.2. Weighted Gene Co-Expression Network Construction and Module Identification
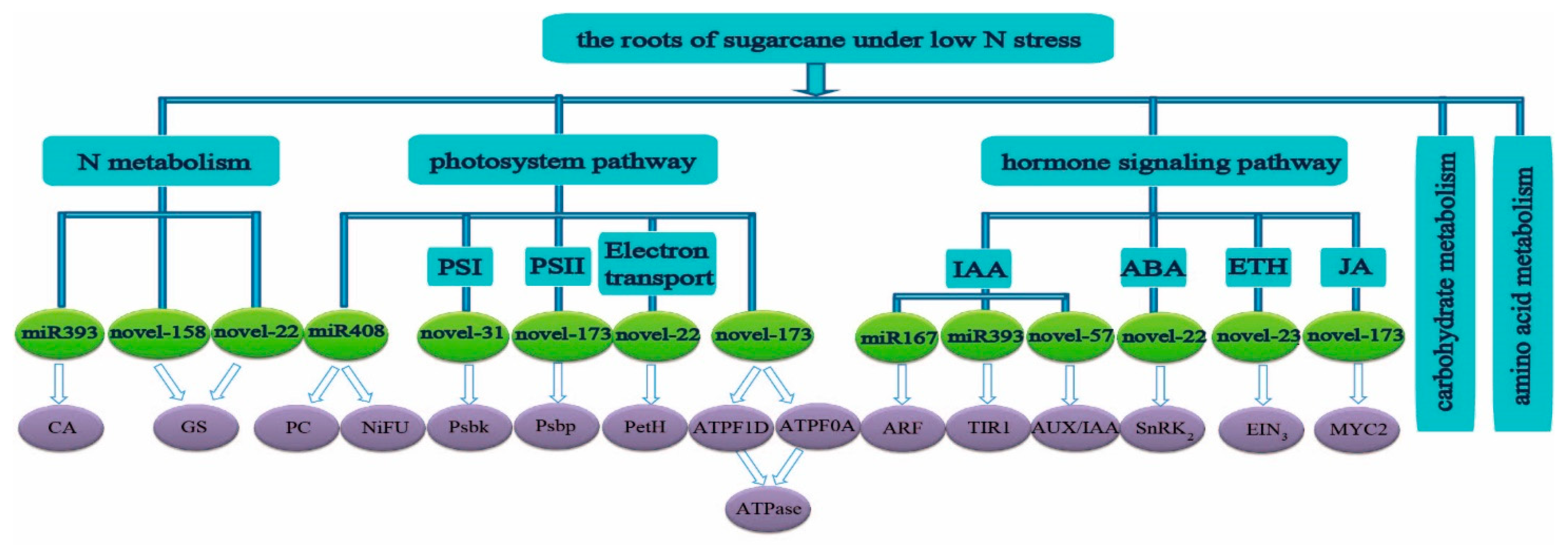
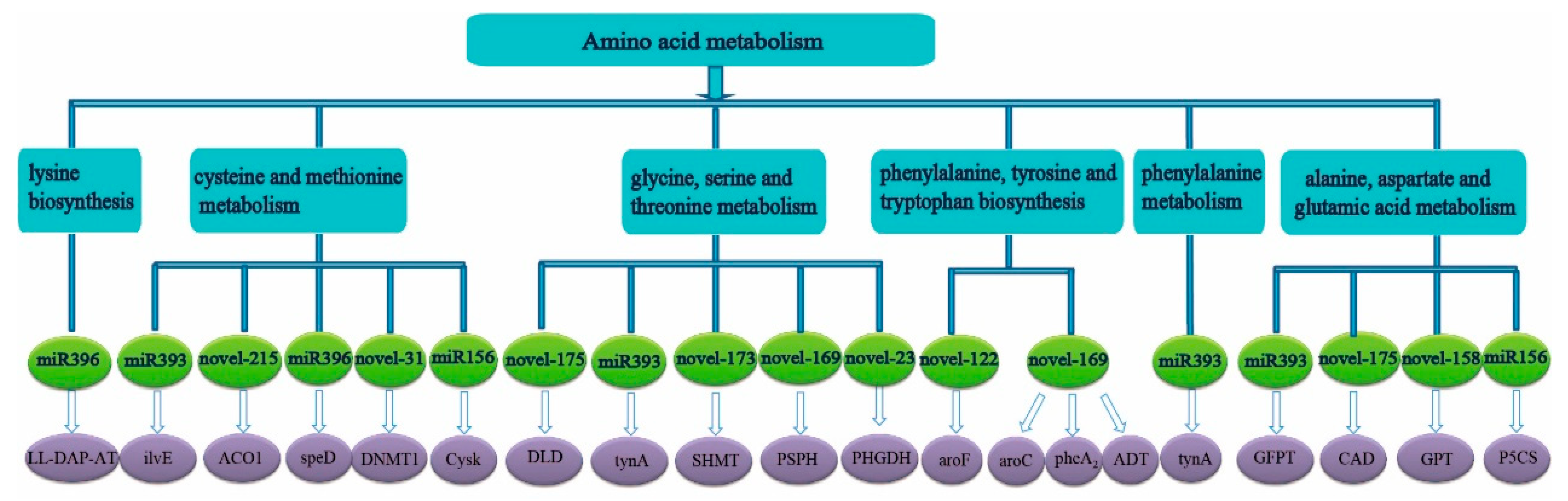
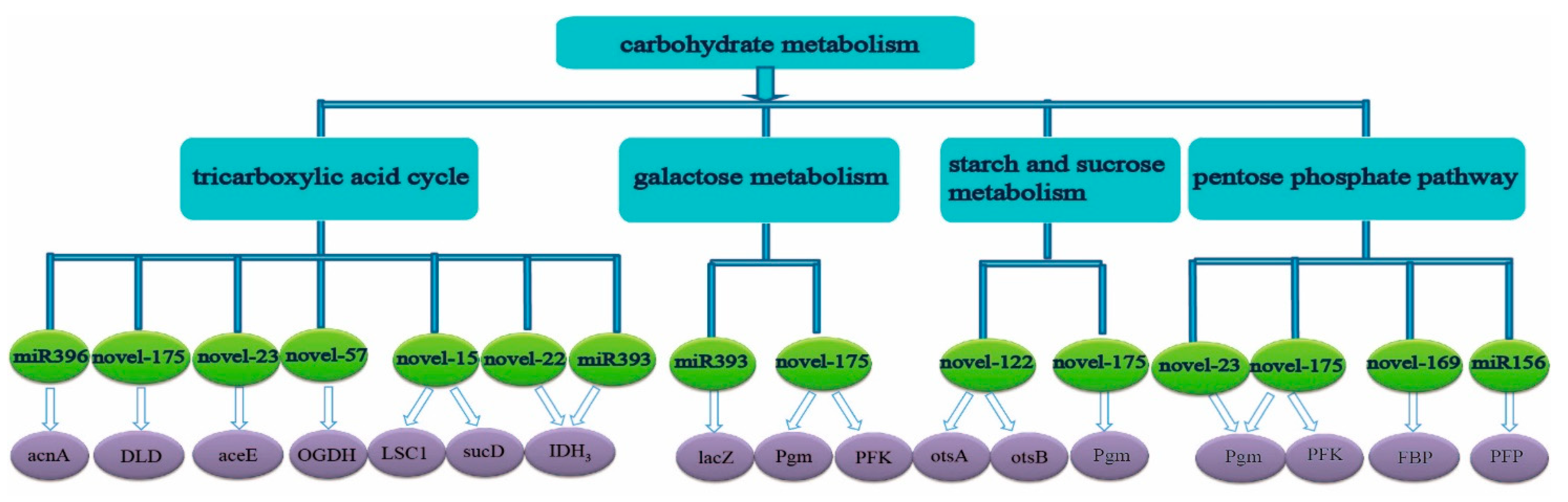
2.3. Prediction and Function of Target Genes of miRNA
2.4. Crosstalk with miRNA and mRNA Transcriptome Sequencing Data
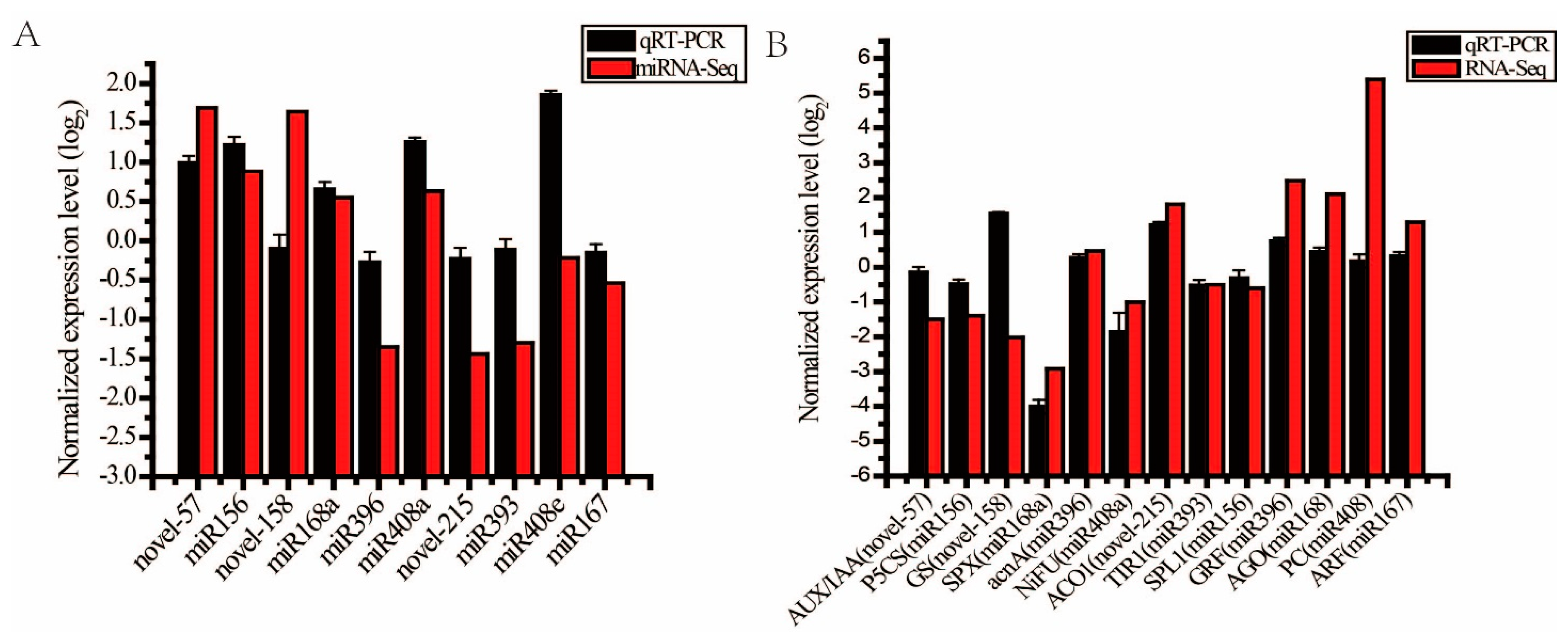
2.5. Verification of the Transcriptome Sequencing Data of miRNAs and Candidate Target Genes by qRT-PCR
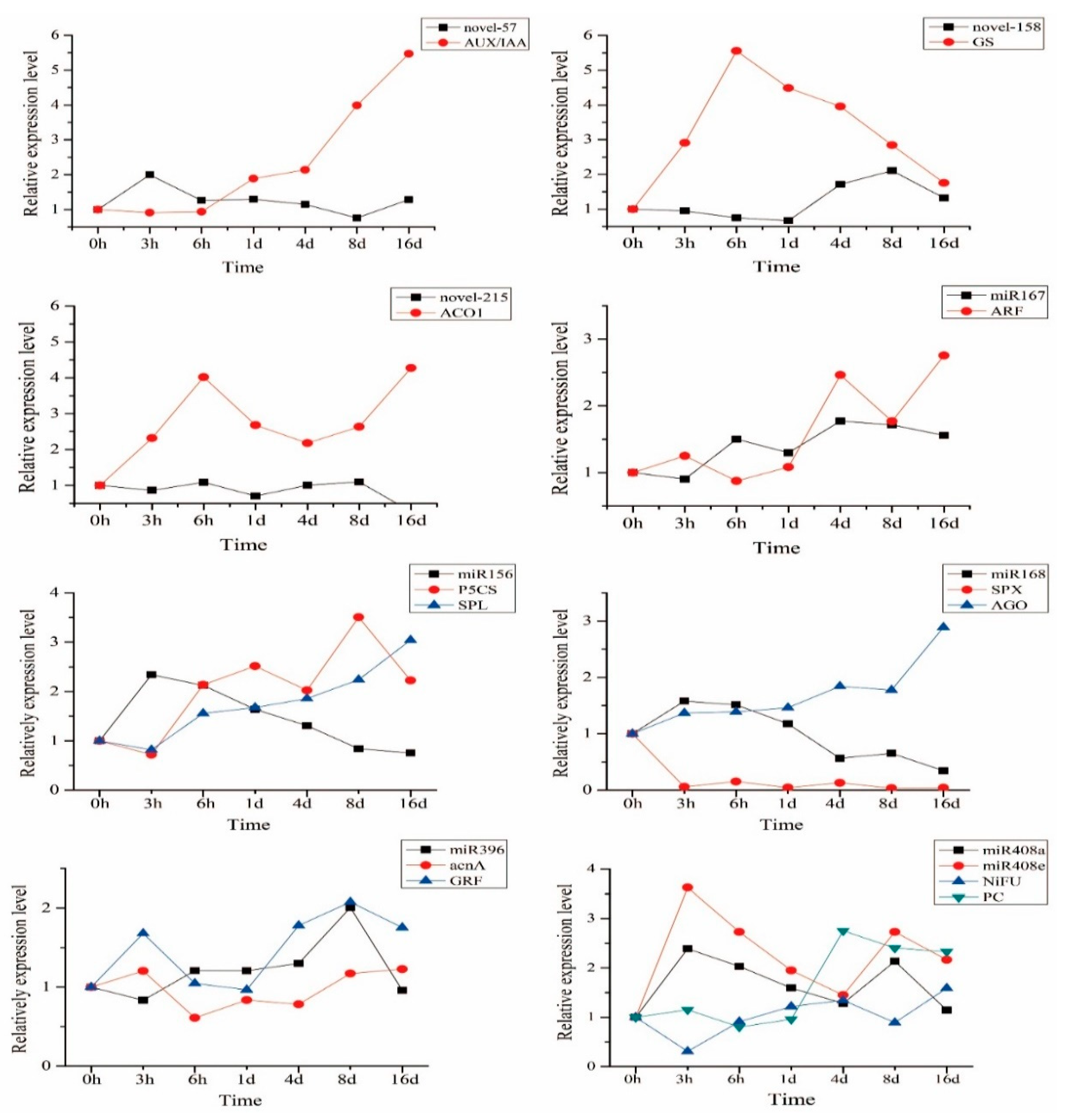
2.6. Expression Trend of miRNA and Candidate Target Genes in Sugarcane under Low N Stress
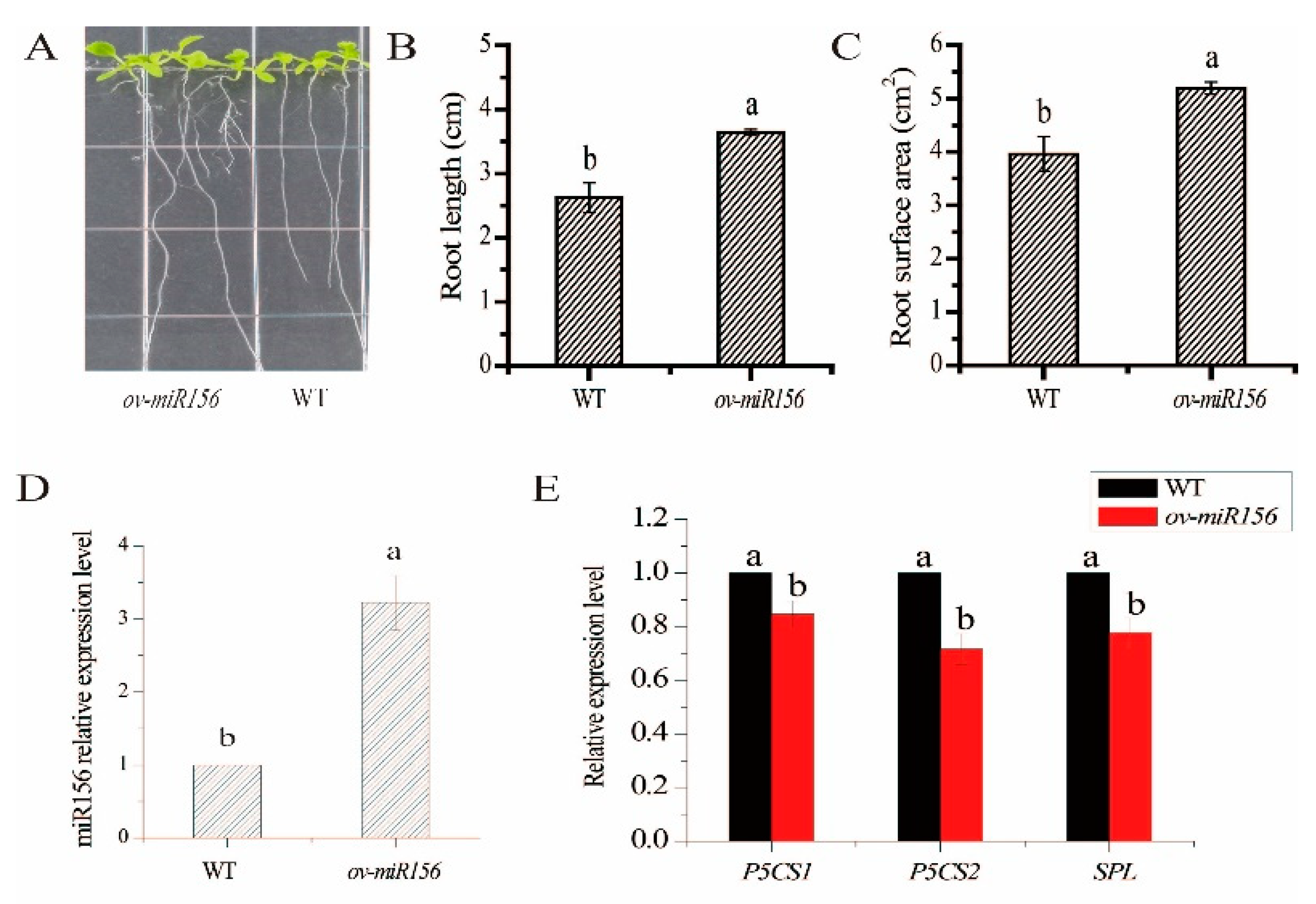
2.7. Characterization of the miR156-Overexpressed Arabidopsis Plants under Low N Treatment
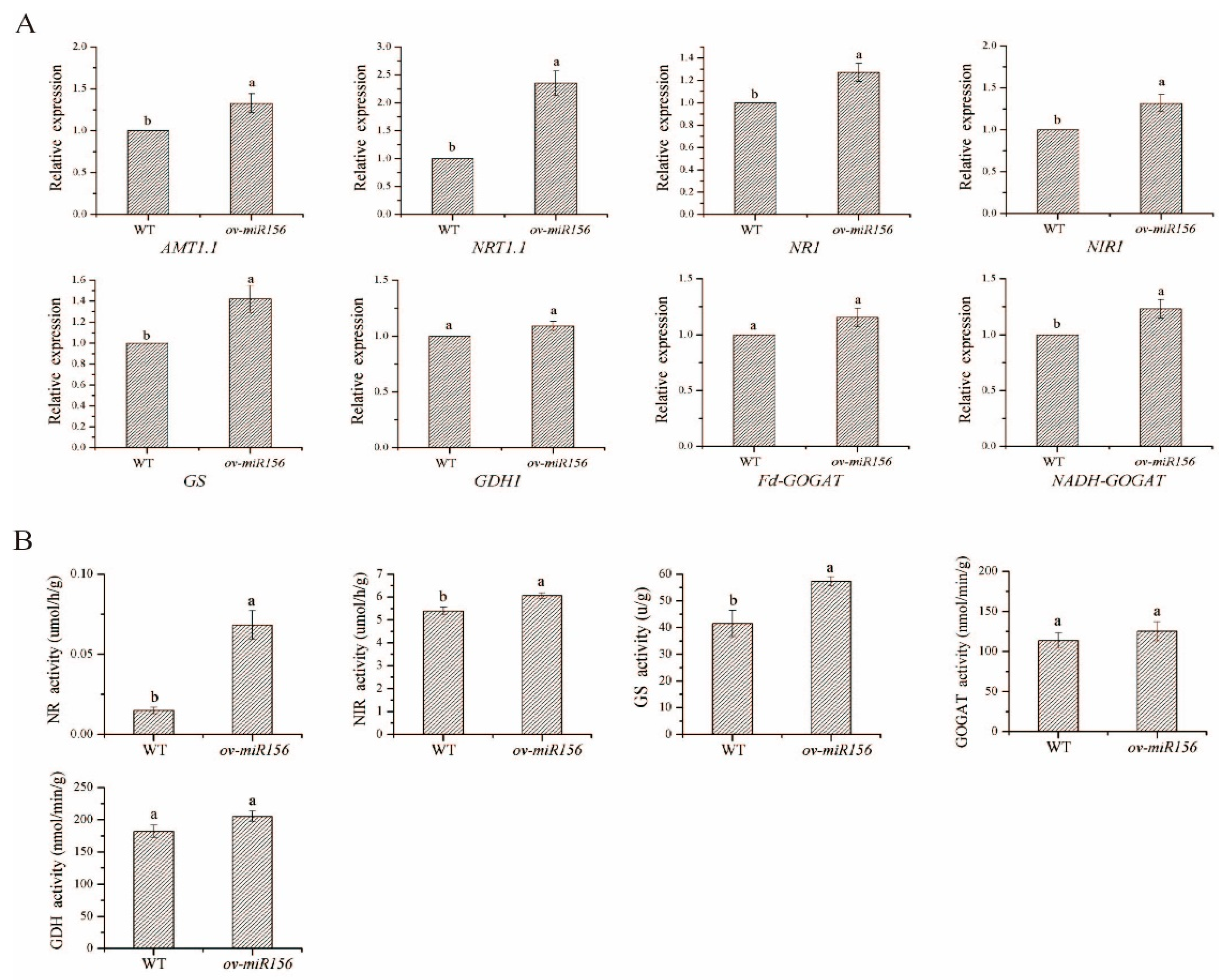
2.8. Changes in the N Assimilation Key Genes Expressions and Enzymes Activities in the miR156-Overexpressed Arabidopsis under Low N Treatment
3. Discussion
3.1. The miRNA Data of Sugarcane Root under Low N Stress Were Reliable
3.2. The Predicted Target Genes of Differentially Expressed miRNA Were Mainly Concentrated in N Metabolism, Carbohydrate Metabolism, and Amino Acid Metabolism
3.3. miRNA Was Differentially Expressed in Sugarcane under Low N Stress
3.4. Diverse Expression Trends of miRNA and Predicted Target Genes in Sugarcane under Low N Stress
3.5. The miR156 from Sugarcane Roots Could Enhance the N Assimilation Ability of Arabidopsis under Low N Application
4. Materials and Methods
4.1. Cultivation of Plant Materials
4.2. cDNA Library Construction and Small RNA (sRNA) Sequencing
4.3. Detection of miRNA and Target Gene Expression under Low N Stress
4.4. Acquisition of the miR156-Overexpressed Arabidopsis
4.5. Low N Treatment of the miR156-Overexpressed Arabidopsis
4.6. Physiological Changes and qRT-PCR Detection of the miR156-Overexpressed Arabidopsis under Low N Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.N.; Feng, X.M.; Zeng, Q.Y.; Lin, H.Z.; Wu, Z.L.; Gao, X.N.; Huang, Y.H.; Wu, J.Y.; Qi, Y.W. Integrated analysis of miRNAs associated with sugarcane responses to low-potassium stress. Front. Plant Sci. 2022, 12, 750805. [Google Scholar] [CrossRef]
- Qiu, L.; Luo, H.; Zhou, H.; Yan, H.; Fan, Y.; Zhou, Z.; Chen, R.; Liu, J.; Luo, T.; Deng, Y.; et al. MicroSugar: A database of comprehensive miRNA target prediction framework for sugarcane (Saccharum officinarum L.). Genomics 2022, 114, 110420. [Google Scholar] [CrossRef]
- Li, Y.R.; Yang, L.T. Sugarcane agriculture and sugar industry in China. Sugar Tech. 2015, 17, 1–8. [Google Scholar] [CrossRef]
- Martinez-Feria, R.A.; Castellano, M.J.; Dietzel, R.N.; Helmers, M.J.; Liebman, M.; Huber, I.; Archontoulis, S.V. Linking crop- and soil-based approaches to evaluate system nitrogen-use efficiency and tradeoffs. Agr. Ecosyst. Environ. 2018, 256, 131–143. [Google Scholar] [CrossRef]
- Sunkar, R. MicroRNAs with macro-effects on plant stress responses. Semin. Cell Dev. Biol. 2010, 21, 805–811. [Google Scholar]
- Reyes, J.L.; Chua, N.H. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant J. 2007, 49, 592–606. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and transcriptional responses of two contrasting Populus clones to nitrogen stress. Tree Physiol. 2016, 36, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Tai, H.; Sun, S.; Zhang, F.; Xu, Y.; Li, W.X. Cloning and characterization of maize miRNAs involved in responses to nitrogen deficiency. PLoS ONE 2012, 7, e29669. [Google Scholar]
- Lin, S.I.; Santi, C.; Jobet, E.; Lacut, E.; Kholti, N.E.; Karlowski, W.M.; Verdeil, J.L.; Breitler, J.C.; Périn, C.; Ko, S.S.; et al. Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation. Plant Cell Physiol. 2010, 51, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Achkar, N.P.; Cambiagno, D.A.; Manavella, P.A. miRNA biogenesis: A dynamic pathway. Trends Plant Sci. 2016, 21, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Wang, L.; Chen, S.; Jiang, J.; Guan, Z.; Li, P.; Chen, F. Identification of nitrogen starvation-responsive microRNAs in Chrysanth. Nankingense. Plant Physiol. Biochem. 2015, 91, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Moyano, T.C.; Krouk, G.; Katari, M.S.; Tanurdzic, M.; McCombie, W.R.; Coruzzi, G.M.; Gutiérrez, R.A. Integrated RNA-seq and sRNA-seq analysis identifies novel nitrate-responsive genes in Arabidopsis thaliana roots. BMC Genom. 2013, 14, 701. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.K.; Shin, C. Transcriptomic analyses of rice (Oryza sativa) genes and non-coding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef]
- Xu, Z.; Zhong, S.; Li, X.; Li, W.; Rothstein, S.J.; Zhang, S.; Bi, Y.; Xie, C. Genome-wide identification of microRNAs in response to low nitrate availability in maize leaves and roots. PLoS ONE 2011, 6, e28009. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.B.D.; Soares, J.D.M.; Lima, J.E.; Silva, J.C.; Ivamoto, S.T.; Baba, V.Y.; Souza, S.G.H.; Lorenzetti, A.P.R.; Paschoal, A.R.; Meda, A.R.; et al. An integrated analysis of mRNA and sRNA transcriptional profiles in Coffea arabica L. roots: Insights on nitrogen starvation responses. Funct. Integr. Genomic. 2019, 19, 151–169. [Google Scholar] [CrossRef] [Green Version]
- Nischal, L.; Mohsin, M.; Khan, I.; Kardam, H.; Wadhwa, A.; Abrol, Y.P.; Iqbal, M.; Ahmad, A. Identification and comparative analysis of microRNAs associated with low-N tolerance in rice genotypes. PLoS ONE 2012, 7, e50261. [Google Scholar] [CrossRef]
- Xie, K.; Shen, J.; Hou, X.; Yao, J.; Li, X.; Xiao, J.; Xiong, L. Gradual increase of miR156 regulates temporal expression changes of numerous genes during leaf development in rice. Plant Physiol. 2012, 158, 1382–1394. [Google Scholar] [CrossRef] [Green Version]
- Pant, B.D.; Musialak-Lange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.R. Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Sun, F.; Hou, J.; Chen, L.; Zhang, Y.; Kang, X.; Wang, Y. Differential profiling analysis of miRNAs reveals a regulatory role in low N stress response of Populus. Funct. Integr. Genomic. 2015, 15, 93–105. [Google Scholar] [CrossRef]
- Shen, J.; Xie, K.; Xiong, L. Global expression profiling of rice microRNAs by one-tube stem-loop reverse transcription quantitative PCR revealed important roles of microRNAs in abiotic stress responses. Mol. Genet. Genom. 2010, 284, 477–488. [Google Scholar] [CrossRef]
- Kong, Y.M.; Elling, A.A.; Chen, B.; Deng, X. Differential expression of microRNAs in maize inbred and hybrid lines during salt and drought stress. Am. J. Plant Sci. 2010, 1, 8. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Davis, K.E.; Patterson, C.; Oo, S.; Liu, W.; Liu, J.; Wang, G.; Fontana, J.E.; Thornburg, T.E.; et al. Response of root growth and development to nitrogen and potassium deficiency as well as microRNA-mediated mechanism in peanut (Arachis hypogaea L.). Front. Plant Sci. 2021, 12, 695234. [Google Scholar] [CrossRef]
- Aung, B.; Gao, R.; Gruber, M.Y.; Yuan, Z.C.; Sumarah, M.; Hannoufa, A. MsmiR156 affects global gene expression and promotes root regenerative capacity and nitrogen fixation activity in alfalfa. Transgenic Res. 2017, 26, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Y.; Gao, S.W.; Su, Y.C.; Lin, Z.L.; Guo, J.L.; Li, M.J.; Wang, Z.T.; Que, Y.X.; Xu, L.P. Transcripts and low nitrogen tolerance: Regulatory and metabolic pathways in sugarcane under low nitrogen stress. Environ. Exp. Bot. 2019, 163, 97–111. [Google Scholar] [CrossRef]
- Bartel, D.P. microRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.T.; Yu, Q.; Yang, Y.Y.; Su, Y.C.; Waqar, A.; Guo, J.L.; Gao, S.W.; Xu, L.P.; Que, Y.X. Identification of cold-related miRNAs in sugarcane by small RNA sequencing and functional analysis of a cold inducible ScmiR393 to cold stress. Environ. Exp. Bot. 2018, 155, 464–476. [Google Scholar] [CrossRef]
- Que, Y.X.; Su, Y.C.; Guo, J.L.; Wu, Q.B.; Xu, L.P. A global view of transcriptome dynamics during Sporisorium scitamineum challenge in sugarcane by RNA-seq. PLoS ONE 2014, 9, e106476. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, C.; Hao, Q.; Sha, A.; Zhou, R.; Zhou, X.; Yuan, L. Elucidation of miRNAs-mediated responses to low nitrogen stress by deep sequencing of two soybean genotypes. PLoS ONE 2013, 8, e67423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauffer, E.; Maizel, A. Post-transcriptional regulation in root development. Wiley Interdiscip. Rev. RNA. 2014, 5, 679–696. [Google Scholar] [PubMed]
- Xu, P.; Vernooy, S.Y.; Guo, M.; Hay, B.A. The Drosophila microRNA Mir-14 suppresses cell death and is required for normal fat metabolism. Curr. Biol. 2003, 13, 790–795. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Huang, H. Roles of small RNAs in plant disease resistance. J. Integr. Plant Boil. 2014, 56, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.M.; Song, R.; Peng, H.; Luo, M.; Shen, Y.O.; Liu, L.; Zhao, M.J.; Pan, G.T. Bioinformatic prediction of microRNAs and their target genes in maize. Acta. Agron. Sin. 2010, 36, 1324–1335. [Google Scholar] [CrossRef]
- Liang, G.; He, H.; Yu, D.Q. Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS ONE 2012, 7, e48951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Cheng, X.; Liu, P.; Sun, J. miR156-targeted SBP-Box transcription factors interact with DWARF53 to regulate TEOSINTE BRANCHED1 and BARREN STALK1 expression in bread wheat. Plant Physiol. 2017, 174, 1931–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.M.; Wang, Y.J.; Zhang, N.N.; Zhang, X.B.; Wu, J.Y.; Huang, Y.H.; Ruan, M.H.; Zhang, J.S.; Qi, Y.W. Systematic identification, evolution and expression analysis of the SPL gene family in sugarcane (Saccharum spontaneum). Trop. Plant Biol. 2021, 14, 313–328. [Google Scholar] [CrossRef]
- Zhao, Y.; Wen, H.; Teotia, S.; Du, Y.; Zhang, J.; Li, J.; Sun, H.; Tang, G.; Peng, T.; Zhao, Q. Suppression of microRNA159 impacts multiple agronomic traits in rice (Oryza sativa L.). BMC Plant Biol. 2017, 17, 215. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Tian, X.; Li, Y.J.; Wu, C.A.; Zheng, C.C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.A.; Chen, C.; Shireen, F.; Zheng, Z.; Sohail, H.; Afzal, M.; Ali, M.A.; Bie, Z.; Huang, Y. Genome-wide expression profiling of leaves and roots of watermelon in response to low nitrogen. BMC Genom. 2018, 19, 456. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.; Tuteja, N. NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 2016, 253, 767–786. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.C.; Bartel, D.P.; Barrel, B. MicroRNA-directed regulation of Arabidopsis A UXIN RESPONSE FACTORl 7 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [Green Version]
- Gifford, M.L.; Dean, A.; Gutierrez, R.A.; Coruzzi, G.M.; Birnbaum, K.D. Cell specific nitrogen responses mediate developmental plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghany, S.E.; Pilon, M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J. Biol. Chem. 2008, 283, 15932–15945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Poethig, R.S. Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development 2006, 133, 3539–3547. [Google Scholar] [CrossRef] [Green Version]
- Kantar, M.; Lucas, S.; Budak, H. miRNA expression patterns of triticum dicoccoides in response to shock drought stress. Planta 2011, 233, 471–484. [Google Scholar] [CrossRef]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Xing, S.; Salinas, M.; Höhmann, S.; Berndtgen, R.; Huijser, P. miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell. 2010, 22, 3935–3950. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Sunkar, R.; Zhou, C.; Shen, H.; Zhang, J.Y.; Matts, J.; Wolf, J.; Mann, D.G.J.; Stewart, C.N., Jr.; Tang, Y.; et al. Overexpression of miR156 in switchgrass (Panicum virgatum L.) results in various morphological alterations and leads to improved biomass production. Plant Biotechnol. J. 2012, 10, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An insight into microRNA156 role in salinity stress responses of alfalfa. Front. Plant Sci. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL16 transcription factor. Plant Cell. 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Sun, X.C.; Hussain, S.; Ali, U.; Rana, M.S.; Rasul, F.; Saleem, M.H.; Moussa, M.G.; Bhantana, P.; Afzal, J.; et al. Molybdenum-induced effects on nitrogen metabolism enzymes and elemental profile of winter wheat (Triticum aestivum L.) under different nitrogen sources. Int. J. Mol. Sci. 2019, 20, 3009. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Zhou, J.; Li, H.; Shi, W.; Polle, A.; Lu, M.; Sun, X.; Luo, Z.B. Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess. Tree Physiol. 2015, 35, 1283–1302. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Y.; Gao, S.W.; Jiang, Y.; Lin, Z.L.; Luo, J.; Li, M.J.; Guo, J.L.; Su, Y.C.; Xu, L.P.; Que, Y.X. The physiological and agronomic responses to nitrogen dosage in different sugarcane varieties. Front. Plant Sci. 2019, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, X.; Chen, Y.; Guo, J.; Ling, H.; Gao, S.; Su, Y.; Que, Y.; Xu, L. Selection of reference genes for normalization of microRNA expression by RT-qPCR in sugarcane buds under cold stress. Front. Plant Sci. 2016, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.; Wu, Q.; Guo, J.; Xu, L.; Que, Y. Comprehensive selection of reference genes for gene expression normalization in sugarcane by real time quantitative RT-PCR. PLoS ONE. 2014, 9, e97469. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, X.; Su, Y.; Zou, J.; Wang, Z.; Xu, L.; Que, Y. miRNA alteration is an important mechanism in sugarcane response to low temperature environment. BMC Genom. 2017, 18, 833. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, S.C.; Lin, C.S.; Hsu, P.K.; Lin, S.H.; Tsay, Y.F. The Arabidopsis nitrate transporter NRT1.7, expressed in phloem, is responsible for source-to-sink remobilization of nitrate. Plant Cell. 2009, 21, 2750–2761. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Yang, Y.; Yang, Y.; Zhang, X.; Su, Y.; Guo, J.; Que, Y.; Xu, L. Identification of Low-Nitrogen-Related miRNAs and Their Target Genes in Sugarcane and the Role of miR156 in Nitrogen Assimilation. Int. J. Mol. Sci. 2022, 23, 13187. https://doi.org/10.3390/ijms232113187
Gao S, Yang Y, Yang Y, Zhang X, Su Y, Guo J, Que Y, Xu L. Identification of Low-Nitrogen-Related miRNAs and Their Target Genes in Sugarcane and the Role of miR156 in Nitrogen Assimilation. International Journal of Molecular Sciences. 2022; 23(21):13187. https://doi.org/10.3390/ijms232113187
Chicago/Turabian StyleGao, Shiwu, Yingying Yang, Yuting Yang, Xu Zhang, Yachun Su, Jinlong Guo, Youxiong Que, and Liping Xu. 2022. "Identification of Low-Nitrogen-Related miRNAs and Their Target Genes in Sugarcane and the Role of miR156 in Nitrogen Assimilation" International Journal of Molecular Sciences 23, no. 21: 13187. https://doi.org/10.3390/ijms232113187
APA StyleGao, S., Yang, Y., Yang, Y., Zhang, X., Su, Y., Guo, J., Que, Y., & Xu, L. (2022). Identification of Low-Nitrogen-Related miRNAs and Their Target Genes in Sugarcane and the Role of miR156 in Nitrogen Assimilation. International Journal of Molecular Sciences, 23(21), 13187. https://doi.org/10.3390/ijms232113187