Abstract
MicroRNAs are hidden players in complex psychophysical phenomena such as depression and anxiety related disorders though the activation and deactivation of multiple proteins in signaling cascades. Depression is classified as a mood disorder and described as feelings of sadness, loss, or anger that interfere with a person’s everyday activities. In this review, we have focused on exploration of the significant role of miRNAs in depression by affecting associated target proteins (cellular and synaptic) and their signaling pathways which can be controlled by the attachment of miRNAs at transcriptional and translational levels. Moreover, miRNAs have potential role as biomarkers and may help to cure depression through involvement and interactions with multiple pharmacological and physiological therapies. Taken together, miRNAs might be considered as promising novel therapy targets themselves and may interfere with currently available antidepressant treatments.
1. Introduction
Anxiety is a disturbed mental health condition in which a person responds to stimuli and situations with fear, increased heart rate, and sweating [1]. There are different types of anxiety disorders at diverse age ranges (16–85 years) such as generalized anxiety (GA; 2.7%), obsessive-compulsive anxiety (OC; 1.9%), panic (2.6%), social anxiety (4.7%), post-traumatic stress disorder (PTSD; 6.4%) [2]. Globally, it has been estimated that 264 million individuals are facing anxiety problems which are mostly affecting females as compared to males (4.6%, vs. 2.6%). The major cause of anxiety disorder is disturbance in brain chemistry, environmental stress, and genetic inheritance from parents [3]. Similarly, as anxiety, depression is also a neurological disease in which person feels hopeless, sleeps too much, thinks about death and suicide, and feels guilty [4]. There are different types of depression such as major depressive disorder (MDD), persistent depressive disorder (PDD)—a depression stage which lasts for two or more years, and disruptive mood dysregulation disorder (DMDD) that frequently becomes aggressive [5]. The major causes of depression are: genetic predispositions, poor nutrition, stress, excessive intake of drugs, and brain unbalancing. However, it has also been observed that around 85% of depression patients experience significant symptoms of anxiety [6].
2. Neurobiology and Molecular Basis of Anxiety and Depression
The etiology and underpinning of anxiety and depression depends upon the neuronal correlations in brain and their responses to different biological conditions and environmental factors [7]. The disturbed behavior such as fear and stress depends upon the neuronal activity and communication between the related brain areas such as hippocampus (the memory center), anterior cingulate (the brain’s conflict-resolution area), and prefrontal cortex (involved in planning and executing activities), respectively [8,9]. The basic features of anxiety are identification of potential threat or harm, the psychological features of alarm, dread or fear, and physiological response that includes autonomic discharge and motor activity, respectively. However, each feature is controlled by different brain regions such as cortical, subcortical and hypothalamus, respectively. It has been also observed that hypothalamus is particularly involved in alerting the person of impending harm, and brain stem responses [10]. Moreover, the individuals with panic disorder may have subtle developmental abnormalities of the right cerebral hemisphere. The left cerebral hemisphere specializes in discrete functions like language, while the right hemisphere specializes in multimodal sensory function and emotions. Lesions in right hemisphere, i.e., mesial temporal cortex are commonly associated with panic disorder [10]. Amygdala consists of two sub-regions such as basolateral amygdala (BLA) and central amygdala (CeA) which is usually linked with emotional response to environmental information [9]. The medial prefrontal cortex (MPFC) is connected to the hippocampus and encodes anxiety related behavior [9].
3. Disruptions of Synaptic Circuits and Their Associated Function
Disturbances in synaptic circuits are caustic factors of depression and related disorders [11,12]. Reduction in prefrontal cortex and hippocampus volume associates with ailment and time of action and chronic conditions in depression. When stress is introduced, it results in the reduction or loss of neurons and glia in the hippocampus and prefrontal cortex (PFC) [13,14,15]. The synaptic plasticity of the brain (the change that occurs at synapses, the junctions between neurons that allow them to communicate) plays significant function in signaling pathways and synaptogenesis. The synaptic plasticity is also involved in decrease or loss of neurotrophic factors (NTFs), disturbance of estradiol cycle, and increased levels of inflammatory cytokines which are ultimately causes of depression [13]. Glia (non-neuronal cells) also plays an important role in the regulation and reuptake of glutamate which is also involved in the depression [16,17].
4. Hypothalamic-Pituitary-Adrenal (HPA) Axis and Glucocorticoid
In depression, acute traumatic or chronic stress is considered the most common factor [18], however, long-lasting stress can cause changes in DNA and histones which results in abnormal behavior in adulthood [19,20,21]. Stress responses can be activated by the hypothalamic-pituitary-adrenal (HPA) axis due to increase in the circulating levels of glucocorticoids. High levels of glucocorticoid affect multiple organs and lead to negative feedback mechanism [22,23,24].
The increased glucocorticoid levels act on multiple levels of neuronal functions (decreasing levels or numbers of synapses) and behaviors [25,26]. The prior research also showed that acute stress increases the levels of glutamate and neuronal transmission in the prefrontal cortex and hippocampus, thereby influencing some aspects of cognitive processing [17,27,28]. Moreover, the molecular studies from mouse and human reveal the role of glucocorticoid in the regulation of signaling pathways, gene transcription, and epigenetic mechanism of regulation of glucocorticoid receptors [29]. Additionally, genetic variations in the genes encoding the FKBP5 and CRF-1, have been linked with different neuropsychiatric diseases and stressful events of life such as childhood abuse and trauma [30]. Figure 1 illustrates the mechanistic pathway of stress induced disturbance in brain.
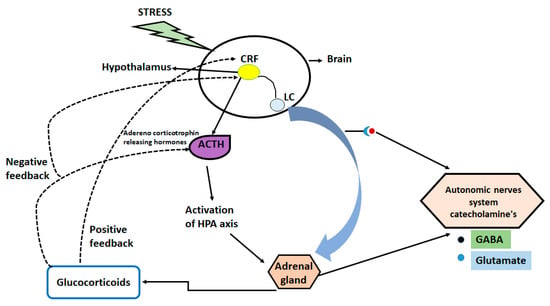
Figure 1.
Mechanistic pathway of stress induced disturbance in brain.
5. Neurotropic Factors and Associated Signaling Pathways in Stress and Depression
There are several growth factors, including brain-derived neurotrophic factors (BDNFs), vascular endothelial growth factors (VEGFs), fibroblast growth factors 2 (FGF-2), and insulin growth factor 1 (IGF1) that are involved in depression. The BDNF growth factors are necessary for synaptic networks [31,32]. Depression is negatively correlated with expression of BDNFs in patients’ blood. The enhanced stress and depression, decrease the concentration of BDNFs in blood in the prefrontal cortex and hippocampus [33,34]. The reduction in neurotrophic growth factors are responsible to the disruptions in brain neuroplasticity caused by stress and depression [35]. Therefore, to overcome this situation, different antidepressants-like selective serotonin reuptake inhibitors (SSRIs) can be used to increase the levels of BDNF and modulate the neurotrophic growth factors expression. A frequent polymorphism of the BDNF gene (val(66)met) is suggested to modulate hippocampal neuronal plasticity and has been associated with individual variations in emotional reactivity traits and episodic memory [36]. Moreover, alleles (val(66)met) block the process of releasing of BDNF causing the shrinkage of neurons in the hippocampus and MPFC [37]. Furthermore, decreased levels of BDNF decrease the spine densities and dendritic lengths in hippocampus and PFC neurons and decrease volume of hippocampus [38,39,40,41].
Additionally, neurotropic factors are also linked with intracellular pathways and control the synaptic connectivity and neural function through the activation of Phosphoinositide 3-Kinase (PI3K), AKT, Raf-1 Proto-Oncogene, Serine/Threonine Kinase (RAF), Mitogen-activated protein kinase (MEK), and Extracellular signal-Regulated Kinases (ERK), respectively [42,43]. These signaling pathways relate to many downstream targets which affect many neuronal functions including protection and survival of neurons and initiation of synaptic plasticity.
Rapamycin targets the protein kinase forming the mammalian Target Of Rapamycin (mTOR) that regulates the cell growth, survival, metabolism, and immunity. The mTOR exists in two forms mTORC1-2, which is formed by binding to two distinct accessory proteins such as Raptor and Rictor, respectively, with diverse substrates. The mTORC1 is an important kinase that activates the translation of synaptic proteins and is involved in regulation of synaptic plasticity [42,44]. The mTORC1 is composed of couple of downstream substrates like p70-S6 kinase 1 (S6K1) and eukaryotic initiation factor 4E binding protein 1 (4E-BP1). Moreover, activation of mTORC1 and enhanced synaptogenesis in the prefrontal cortex appear to be important in mediating the antidepressant effects of ketamine [45]. Therefore, faster-acting antidepressants stimulate the mTORC1signaling pathway in the prefrontal cortex [46,47,48,49].
Regulated in Development and DNA Damage Responses 1 protein (REDD1) is a conserved protein encoded by REDD1 gene produced during cellular stresses [50]. The REDD1 gene is present and mostly expressed in most adult tissues of human, mouse and drosophila, [51]. The REDD1 protein is composed of 232 amino acids which lack functional domains in the structure. The prior reported studies showed that REDD1 plays either a pro- or an anti-apoptotic role during the stress response, depending on the cell context [52]. REDD1 is a negative regulator of mTORC1; the expression of REDD1 is elevated in prefrontal cortex (PFC) causing depression in subjects during hypoxic stress [53]. The mice studies also showed that the chronic stress can decrease the level of mTORC1 signaling proteins causing the loss of neurotransmitters in the PFC [54,55].
6. Role of Neurotransmitters in Depression
Neurosciences focus on brain regions (PFC, hippocampus, amygdala, anterior cingulate) and different neurotransmitters which control the regulation of normal emotions and stress-induced situations. Multiple neurotransmitters and neuromodulators like gamma-aminobutyric acid (GABA), glutamate, monoamines including serotonin, norepinephrine, dopamine, several neuropeptides are unique players in the regulation of depressive disorders and signaling pathways [56]. Most of the studies have reported biological abnormalities in parents with mood disorders but nowadays neurotransmitters, such as norepinephrine, dopamine, histamine, serotonin are the focus of research and theories of the etiology of these diseases. Disturbance of the signal neurotransmission system is the major reason to study the neurobehavioral system circuit and neuroregulatory mechanisms. Monoamines are key neuromodulators involved in brain functions. Norepinephrine and serotonin are the two most important neurotransmitters which are implicated in the pathophysiology of mood disorders [57] (Figure 2).
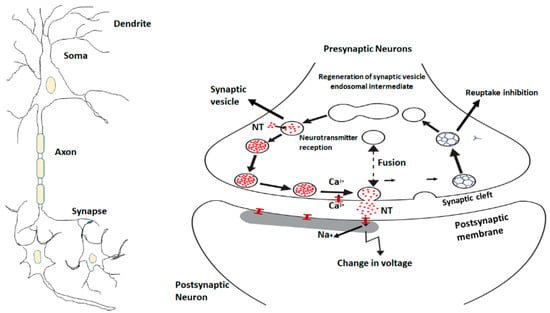
Figure 2.
The neuronal signaling pathway and the movement of neurotransmitters through synaptic vesicles.
6.1. Norepinephrine
Norepinephrine (noradrenaline) is monoamine neurotransmitter involved in depression [58]. Norepinephrine is functionally associated with multiple functions such as memory, stress reactions, and in emotions regulation. Norepinephrine acts as a hormone and is synthesized from dopamine by the catalytic reaction of β-hydroxylase. It has also been observed that norepinephrine producing neurons are mostly present in the pons and medulla parts of central nervous system (CNS).
The norepinephrine neurotransmitters are usually used by the sympathetic nerves system and are found in the form of clusters in sympathetic neurons which are located near the spinal cord called sympathetic ganglia [59]. Norepinephrine acts on GPCRs, as adrenergic or adrenoceptors such as α-1,2 and β-adrenergic receptor, respectively and is frequently associated with arousal, alertness, and attention. However, the full extent of these actions is more complex and release in the sympathetic nerves system is typically associated with increased heart rate and blood pressure [60] (Figure 3).
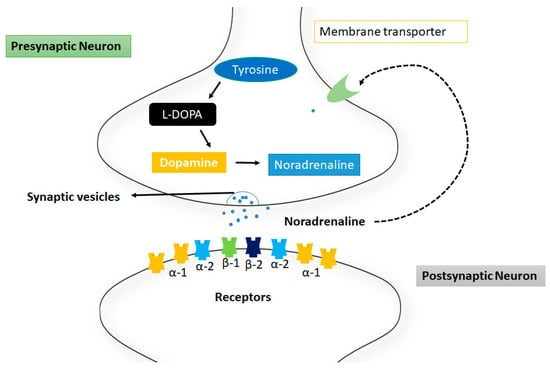
Figure 3.
Mechanism of action of norepinephrine at synapsis. The tyrosine is converted into noradrenaline through intermediates such as L-dopa and dopamine respectively. These neurotransmitters are released from the membrane through synaptic vesicles. There are different receptors represented in different colors at postsynaptic neuron.
6.2. Serotonin
Serotonin (5-hydroxytryptamine) is a monoamine neurotransmitter involved in many brain functions modulating mood, and in different physiological processes [61]. Serotonin is produced in specialized neurons mostly in raphe nuclei and in major subdivision in the CNS [62,63]. To synthesize serotonin, the amino acid tryptophan is converted into 5HTP, and the 5HTP is converted into 5HT. There are seven different serotonin receptors (5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6, and 5-HT7), which include six belonging to GPCRs and one associated with ligand-gated ion channels receptors [64,65]. Functional study showed that serotonin is associated with the pathophysiological depression and low concentration of serotonin in CSF may originate suicidal thoughts in patients. Therefore, many antidepressants are used to rise the levels of serotonin [66,67] (Figure 4).
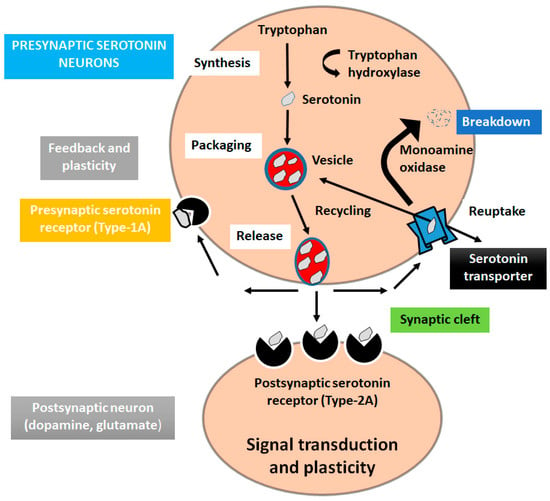
Figure 4.
Serotonin signaling pathway in depression. The amino acid tryptophan is converted into 5HTP, and the 5HTP is converted into 5HT.
6.3. Dopamine
Dopamine is a monoamine neurotransmitter involved in feeling pleasure, satisfaction and motivation [68]. Dopamine synthesis begins with the amino acid phenylalanine, and proceeds sequentially through tyrosine, DOPA, and then dopamine [69]. Dopamine is produced in the ventral tegmental area in the midbrain and acts as a messenger in brain. Dopamine works in various parts such as the PFC, amygdala, nucleus accumbens, hippocampus, and olfactory bulb in the brain. It has also been observed that dopamine pathways carry dopamine from areas of higher concentration to other parts of brain [70,71]. The bilateral dopaminergic pathway is a major dopamine nigrostriatal pathway in the brain that connects the substantia nigra pars compacta (SNc) in the midbrain with the dorsal striatum in the forebrain [72]. Another biochemical study showed that homovanillic acid (HVA) which is dopamine metabolite is responsible for dopamine turnover in depression patients. Moreover, the pharmacological treatments (antidepressants) have been reported for the treatment of depression [73].
6.4. GABA
γ-aminobutyric acid (GABA), a repressive neurotransmitter in CNS [74], that controls the anxiety and overactive fear circuits which are centered in the amygdala [75]. GABA is an amino acid that may help to improve mood [76]. Generally, GABA interacts with GABA-A and GABA-B receptors and mediates neuronal communication and functions and helps with feelings of anxiety, stress, and fear. It has been observed that the binding of GABA with GABA-A receptor is responsible for the permeability of negatively charged ions such as chlorides (Cl−) through the opening of ion channels [77,78]. The flow of chloride ions hyperpolarizes the membrane potential of the neurons and makes it less likely to fire an action potential.
Whereas the GABA-B receptors causes the opening of potassium ion channels upon activation at the synaptic cleft. These channels allow positively charged potassium ions (K+) to flow out the neurons, again making the neuron hyperpolarized and less likely to fire an action potential [79]. These neuronal fluctuation and transmission of ions can control the depression and anxiety symptoms. Several studies showed that increased GABA levels in the brain can help to decrease anxiety, stress, and depression [80] and there are multiple drugs effects as alcohol and benzodiazepines which increase activity at GABA receptor [81,82].
7. miRNAs as Key Players in Depression
Many miRNAs are highly expressed in brain tissues and may be implicated in the pathological changes of the central nervous system (CNS) in depression (Shi et al., 2021). miRs are short (consisting of 19–24 nucleotides) and single-stranded RNAs which are genomically encoded to perform their function in gene expression and in post-transcriptional regulation of target genes [83]. These are non-coding single RNA molecules (that produce no proteins). miRNAs are found naturally, produced endogenously in cells. They are found in plants, animals, and in some viruses [84]. Gene expression performs its function after transcription when messenger RNA (mRNA) is formed. Expression controls if the production of a given protein is increased or decreased, generally the protein production is usually decreased. MicroRNA binds to the untranslated genes of messenger RNA, and there are two ways of binding; perfect complementary binding, and imperfect complementary binding. When there is a perfect binding the mRNA will degrade, and no protein is formed and when there is incomplete binding then the translation is repressed [85]. The overview of different miRs in various brain regions have been depicted in Figure 5.
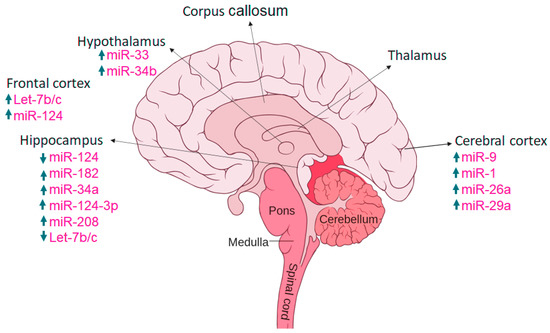
Figure 5.
Expression of miRs in various brain regions, with arrows up and down indicating increased and decreased expression.
7.1. miRNAs Control Neuronal Signaling Pathways
miRNAs are assumed to be a key players in the neuronal synaptic signaling [86,87]. The neurotransmitters such as glutamate, GABA, dopamine, and serotonin, are integrated into the presynaptic neurons, bundled into vesicles, and delivered into the post-synaptic neurons at synaptic cleft. This transportation of neuronal neurotransmitter is done by the encoded receptors present at the post synaptic neurons, encoded by mRNAs which are miRNA targets [88]. miR-153 regulates the synaptosomal-associated protein-25, (SNAP-25) [89] that mediates the calcium-initiated vesicle in combination with plasma layer and arrival of neurotransmitters in synaptic cleft [90]. Moreover, in model animal studies it was revealed that the overexpression of miR-153 represses the synaptic vesicle cycle at neuromuscular intersection whereas control expression results in antagonistic manner in the zebrafish. Furthermore, miR-153 also regulates the movement of zebrafish at undeveloped embryo stage which is probably the results of releasing the acetylcholine at neuromuscular intersections [90,91]. In another model animal study, it has been observed that miR-135a is directly involved in the regulation of early stress response. In the knockdown (KD) mouse study it was reported that miR-135a in amygdala induces an increase in anxiety-like behavior which was confirmed through spontaneous excitatory postsynaptic currents. Moreover, miR-135a also targets complexin-1 and complexin-2 (eukaryotic cytoplasmic neuronal proteins) which are significant for presynaptic vesicle combination [92].
The neuronal transportation across the presynaptic and postsynaptic neurons are impacted by different miRNAs through the calcium signaling at synaptic cleft. In another mice study, it was observed that miR-185 and miR-25 are depleted from mice to check their functional significance in brain. A couple of prior research showed that the deletion of miR-185 and miR-25 results in abnormally increase the level of sarco/endoplasmic reticulum Ca2+-ATPase (Serca2) which causes dysregulation of Ca2+ and glutamate in presynaptic terminals during the sustained neuronal activity that is required for synaptic plasticity at excitatory synapses. Therefore, serca2 levels are raised, which raises calcium levels in the presynaptic cytosol furthermore increasing synapse discharge [93,94].
A single miRNA might target numerous neurotransmitter receptors or subunits with restricting consequences for neuronal activity in the CNS [95]. miR-181a, is an smaller unit having great influence in the hippocampus-dependent memory formation [96]. Another report also showed that miR-181a targets both the α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) and the GABAAα-1 receptors, respectively and controls the mediating signaling pathways [97,98]. Overexpression of miR-181a in essential hippocampal neurons decreases the volume and thickness of dendritic spines, suggesting that miR181a may especially stop excitatory neurotransmission in neuronal cells [97]. Moreover, in vivo data also suggest that silencing of miR-181a produces neuroprotection against hippocampus neuron of cell apoptosis [99]. However, investigations also suggested that knowing a single target of a miRNA may not be adequate to predict the biological impacts of miRNA action on neuronal signaling [99].
The signaling impacts of numerous neurotransmitters are ended by reuptake of the neurotransmitter into the presynaptic transporters, and miRNAs might potentiate neurotransmission by restraining these transporters [100,101]. It has been observed that miR-16 targets SERT which ends serotonergic signaling and may acts as active player in the treatment of depression [102]. The specific serotonin reuptake inhibitor (SSRI) fluoxetine increases miR-16 and diminishes SERT expression in the raphe nuclei, and miR-16 overexpression copies the antidepressant impacts of fluoxetine in mouse models of depression. Then again, neurotransmission might be ended by enzymes that degrade the neurotransmission inside the synaptic split, and miRNAs that target these enzymes are anticipated to potentiate neurotransmission [102]. Similarly, miR-132 has a role in neuroplasticity function and may play an important role in depression [103]. miR-132 represses the formation of acetylcholinesterase (AChE) the catalyst (enzyme) that catalyzes the hydrolysis of acetylcholine at the neuromuscular intersection and in the CNS [104,105]. In mice study it has been observed that increased miR-132 levels in the hippocampus, decrease acetylcholinesterase levels, and impair execution of the hippocampus-dependent tasks. The disturbed expression level of miR-132 may cause neurological disturbance by regulating AChE [105]. Moreover, miRNAs can additionally target mRNAs that encode catalyst (enzyme) and cytoskeletal proteins that impact receptor surface expression, and degradation. For example, miR-125a regulates postsynaptic density protein 95 (PSD-95), a scaffolding protein that regulates the synaptic localization of many receptors, channels, and signaling proteins [106,107].
miR-146a-5p has significant role in neurological diseases and particularly acts as new potential therapeutic target for depression [108]. miR-146a-5p regulates the formation of microtubule-associated protein 1B (MAP1B) present in dendrites which regulates mGluR-induced AMPA receptor endocytosis and consequent long-term depression [109]. The overexpression of miR-146a-5p can block the mGluR-dependent AMPA receptor endocytosis and as a result it may be helpful in the depression. It has been observed that AMPA receptor endocytosis is increased without affecting GluA1 protein levels, and synaptic transmission is depressed by a MAP1B-dependent mechanism [108].
7.2. miRNAs and Growth Factor Signaling
Growth factors are group of proteins which control different metabolic processes through the activation and deactivation of downstream signaling pathways [110]. The clinical data showed that neurotrophic factor (BDNF), a neurotrophin growth factor involved in neuronal maturation, synapse formation and synaptic plasticity [111]. Moreover, the other vascular growth factors such as VEGF or VEGFA are involved in the possessing of neurotrophic and neuroprotective properties, respectively [112,113].
It has been observed that miRNAs straightforwardly target the mRNAs that encode growth factors [114]. The 3′UTR region of BDNF mRNA, which encodes BDNF, appears to be especially significant for regulating stimulus-induced BDNF formation in neurons [115]. A couple of miRNAs such as miR-26a and miR-26b target the 3′UTR of BDNF mRNA in HeLa cells and control the transcriptional activity of mRNA of BDNF [116]. Furthermore, miR-140 and miR-211 also play a similar function and bind with 3′UTR of BDNF in human astrocyte and regulate their inflammation-induced proliferation [117,118]. Another miRNA, miR-206 has good binding behavior with 3′UTR of mRNA of BDNF in mouse and represses BDNF protein formation in Neuro2A cells [119]. Another study shows that miR-149 inhibits the neuregulins (NRGs) signaling by targeting the mRNA that encodes the ErbB3 receptor [120,121]. These studies showed that miRNAs are key player in the regulation of growth factors which control the neurological behavior. Therefore, dysregulation of these miRNAs that target development factors may, add to neurological sickness and can provide a therapeutic target against brain related disorders.
The tropomyosin receptor kinase B (TrkB) acts as receptor for BDNF and its binding alters the neuronal excitability of ion channels and enhances post-synaptic glutamate receptor activation causing influx of Ca2+ ions post-synoptically [122]. The mouse study showed that miR-592 levels decrease in the hippocampus following ischemic injury, permitting the quick synthesis of its target p75NTR, which thus advances apoptotic signaling and neuronal death [123].
7.3. miRNAs in Intrinsic Neuronal Excitability
The excitability of neurons is the ability to generate a large, rapid change of membrane voltage in response to a very small stimulus [124]. miRNAs can regulate inborn neuronal volatility by targeting ion channels that are voltage-gated, rather than ligand-gated. The miR-129 reduces the expression of the shaker voltage-gated potassium channel Kv1.1 encoded by the KCNA1 gene, and miR-324 restrains the synthesis of a Shal-type potassium channel Kv4.2. Both voltage-gated potassium channels Kv1.1 and Kv4.2 open in reaction to membrane depolarization and permit influx of potassium particles [125,126], and the loss of either channel leads to increased susceptibility to seizure in mouse models [127,128]. Both in vitro and in vivo studies showed that inhibition of miR-324-5p protects against kainic acid-induced downregulation of Kv4.2 in neurons. Restraining miR-324-5p fails to defer seizure in potassium voltage-gated channel subfamily D member 2 (KCND2) knockout mice. There is evidence that miR-324-5p plays a role in epileptogenesis via targeting of potassium channel Kv4.2. Some other miRNAs modify seizure susceptibility in mice and rodents by changing gene expression, which makes them attractive candidates as novel treatment targets in epilepsy [129]. It has been shown that reduced expression of miR-101 during early development in mice induced neuronal sensitivity later in life [130].
7.4. Role of miRNAs in Intracellular Signaling
Ion channels proteins and receptors manage the neuronal signaling through membrane potential regulation [131]. miR-126 regulates the phosphoinositide-3-kinase (PI3K) downstream of insulin-like development factor 1 (IGF-1) by targeting the mRNAs that encode insulin receptor substrate 1 (IRS-1) and the PI3K administrative (regulatory) subunit p85β [132,133,134,135]. It has been observed that the hippocampal PI3K/AKT pathway mediates hydrogen sulfide (H2S) ameliorated depression and anxiety in diabetic rats by improving the hippocampal neurogenesis. Additionally, the inhibition of PI3K/AKT pathway by LY294002 exerted the improvement of hippocampal neurogenesis and the antidepressant and anxiolytic-like effects in the streptozotocin (STZ)-induced diabetic rats [136,137]. Another research study showed that, in neuroblastoma cell lines, the overexpression of miR-126 enhances IGF-1 functionality in cell expansion and increases sensitivity. The inhibition of miR-126 upgrades the defensive impacts of IGF-1 against neuronal cell death [135]. Furthermore, the overexpression of miR-126 in neurons diminishes the neuroprotective impacts of BDNF, nerve growth factor (NGF), IGF-1 and increases neuronal weakness to amyloid-beta (Aβ) to reduce neurotoxicity [138]. Besides, overexpression of miR-126 also decreases IGF-1 and induced Akt and p85β pathways [138]. These studies exhibited that miR-126 regulates various proteins downstream signaling pathways and directs neuronal responses to different growth factors. Another functional study showed that increased levels of miR-126 enhanced the functions of dopaminergic neurons signaling in cerebrum [139]. The miR-183 also focuses on mRNA which encodes mTOR and control translation [140]. The miRNA rise might contribute to motor neuron deficiencies in SMA, and stopes excess miRNA activities might have therapeutic utility. Additionally, miR-128 controls the downstream signaling of dopamine 1 receptors (Drd1) in motor neurons by targeting a few parts of the ERK1/2 pathway [141]. Moreover, loss of miR-128 in Drd1 positive neurons leads to motor hyperactivity and deadly seizures [118]. The ERK1/2 inhibitor rectifies ERK2 phosphorylation and motor hyperactivity in mice, showing a role of miR-128 in regulation of ERK1/2 signaling in controlling neuronal movement [142].
7.5. miRNAs and Neuronal Function
miRNAs play an essential role to regulate the genes which are involved in different stages of neurogenesis. There are many miRNAs involved in brain functioning some miRNAs are miR-132 which are involved in the formation of newborn neurons. miR-137 is used in proliferation and forming adult neural stem cells. miR-124 is the most abundant in mouse brain and involved in upregulation, differentiation and mature neurons. miR-9 is neuron-specific mainly expressed in the CNS that helps in cognition, learning, memory, and neuropsychiatric disorders [143]. miR-133b play important role in the maturation of dopaminergic neurons. Dopaminergic cells are the collection of neurons in CNS which can synthesize neurotransmitter dopamine which plays important role in multiple brain functions which include voluntary movement, behavior processes e.g., mood, addiction, stress. miR-18a is the negative regulator in neuronal cells, and it increases expression whereas, miR-16, miR-221, miR-204, miR-15b are highly expressed in distal axons. Recent evidence shows that members of the miR-34 family are the critical modulators for stress response involved in fear and anxiety-related behavior. The miR34c is upregulated in the central nucleus of the amygdala causing acute and chronic stress, therefore, could be used as antidepressant target [144] (Table 1) (Figure 6).
Figure 6.
The predicted three-dimensional (3D) structures of miRNAs involved in anxiety and depression.
7.6. miRNAs and Stress Response
Let us examine the role of miRNAs in response to different types of stress. Acute and chronic stress causes different changes in miRNA expression in brain areas [145]. Acute stress can cause an increase in expression of some selected miRNAs (such as miR-9, miR-26b, miR-29b, miR-30b, miR-30c, miR-30e, miR-125a, miR-126-3P, miR-129-3P, miR-207, miR-212, miR-351, miR-423, miR-487b, miR-494, miR-690, miR-691, miR-709, miR-711, and miR-7a-e) and induce a response in frontal cortex not in the hippocampus. Some of the miRNAs such as let7a, miR-9, miR-26a/b, miR-30b/c and miR-125a increase expression after acute stress. Physiological stress changes the expression of miRNAs in the central amygdala and hippocampus. Chronic stress causes more changes than acute stress, some miRNAs that were changed during acute and chronic stress are miR-132, let7a-1, miR-9-1, and miR124a-1. Prior studies showed that the overexpression of miR-124a is increased in the dentate gyrus and can exacerbate anxiety-like behavior target genes are BDNF [146].
miRNAs responses differently in stress-responsive brain regions. In the CA1 region, miR-376b and miR-208 shows increased expression, whereas, miR-9-1 depicts decreased expression in both acute and chronic stress conditions [147]. miRNAs are important in brain functioning they are involved in learning and memory processes and synaptic plasticity [148]. Moreover, some miRNAs are ubiquitously expressed e.g., let7b, miR-17-5P, and miR-21, respectively [149]. The expression patterns depend on the specific cell type and developmental stages [150]. miR-34a is brain and spinal cord specific, and miR-409-3p is involved in brain development in mice [151]. miR-9 and miR-124a are involved in neural lineage differentiation in embryonic stem cells derived culture [152].
7.7. miRNAs and Immune Response in Depression
miRs are also involved in the regulation of immunological responses including development, maturation, activation, functioning, and aging of various immune cells. Furthermore, miRNAs also exhibited highly specific expression patterns in organs associated with the immune system. miRNAs are also salient key players in both innate and adaptive immune responses [153]. Recent literature data suggested that miR-155, miR-146, and miR-223 exhibited significant role in the regulation of acute inflammatory response as well as in pathogen recognition. Multiple published data also reported that miRs are also involved in the adaptive immune responses which are characterized by activation and clonal expansion of T and B-cells. This activation and expansion lead to cytotoxic effector response and the production of antibodies in response to infections [154]. miRNA has been widely associated with modulating adaptive immunity by regulating the development, activation, survival, and proliferation of T- and B-cells [153]. Differential expression of miR-181, miR-17-92, miR-214, miR-146a, miR-155, let-7, miR-29, miR-125, and miR-216 has been observed in the signaling cascade downstream of T-cell activation [155]. Furthermore, higher expression of miR-150 is also observed in progenitor cells whereas mature B-cells represent its downregulation. Ectopic expression of miRNA-150 followed the premature downregulation of C-Myb, trigger apoptosis during the pro-B stage. Higher levels of miR-150 are necessary for the conversion of pre-B to mature B-cells (to downregulate C-Myb expression), guaranteeing normal B-cell development [156].
Moreover, there are multiple in vitro and bioinformatics studies which showed the significant role of miRs in depression and in suicidal attempts [157,158,159]. It has been observed that stress elicits a host of biological responses including neurochemical cascades in the hypothalamic-pituitary–adrenal (HPA) axis and immune reactions, which can subsequently alter neuronal connectivity and signaling as well as brain matter density [160,161,162,163,164,165]. There is no doubt about the involvement of miRs in the immune responses, however, it has been also shown that their abnormal expression in the immune system can be linked to multiple human diseases including inflammatory disorders, such as inflammatory bowel disease, and cancers [166]. miR-146a in turn inhibits expression of two components of the TLR4 signaling pathway, IL-1 receptor associated kinase and TNF receptor-associated factor-6 [167]. Thus, miR-146a functions as a negative feedback regulator of the TLR/NF-κB pathway. miR-155 and miR-146 expression is increased in macrophages in response to LPS stimulation, while miR-125b expression is decreased. miR-125b can target TNF-α mRNA, and a decrease in its expression leading to elevated TNF-α production and consequently increased inflammatory response [168].
7.8. miRNAs and Major Depressive Disorders
Major Depressive disorder (MDD) is psychiatric mood disorder and in severe conditions, this syndrome is accompanied by either delusion or hallucination [169,170]. Depression is a mood disorder in which a person feels sad every time, feels guilty, or has hypochondriasis [171]. Prior data showed that multiple miRNAs such as miR-1 42-5p/3p, miR-494, miR-376a*, miR-496, and miR-369-3p, miR-23b, miR-27b, miR-24-1*, miR-34b* and miR-34c, miR-17* and miR-20a are significantly downregulated in the MDD [172]. Moreover, miR-134 has stage-specific effects on cortical development [148,173,174]. miR-134′s has also significant involvement in synaptic development and plasticity regulation [175]. Exosomal miRNAs have been suggested to participate in the pathogenesis of neuropsychiatric diseases. The published data showed that exosomes from patients with major depression caused depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Therefore, exosomal miRNAs are promising targets for the diagnosis and treatment of depression related diseases [176]. Another study showed that, miR are good and promising therapeutic biomarkers and good targets for treatment of depression [177].

Table 1.
Role of micro RNAs in anxiety and depression.
Table 1.
Role of micro RNAs in anxiety and depression.
| miRNAs | Expression/Functions | References |
|---|---|---|
| miR-183 | upregulate expression in the amygdala during acute stress | [147] |
| miR-212 | pre miRNA upregulated expression in the medial prefrontal cortex maternal separation | [178] |
| miR-24a | decrease expression in the hippocampus due to treatment with lithium, sodium valproate | [179] |
| miR-488 | SNP associated with panic disorder (genetic variation). | [180] |
| miR-491 | SNP associated with panic disorder | [180] |
| miR-9 | upregulate expression in frontal cortex during acute stress | [145] |
| miR-9-1, 3 | pre miRNA upregulate expression in the medial prefrontal cortex | [178] |
| miR-18 | stress and depression | [181] |
| miR-1 | upregulate expression in the amygdala during chronic stress and downregulate expression in the hippocampus during acute stress | [147] |
| miR-26a/b | during acute stress upregulate expression in frontal cortex | [145] |
| miR-29a | during maternal separation upregulate expression in the medial prefrontal cortex | [178] |
| miR-208 | under acute or chronic stress upregulation of expression in the central amygdala region of hippocampus | [147] |
| miR-221 | decrease expression in the hippocampus | [179] |
| Let-7b/c | upregulate expression in frontal cortex during acute stress. Downregulate in the amygdala after acute and chronic stress; increase expression in the hippocampus | [145,147,179] |
| miR-124 | Upregulate expression medial prefrontal cortex during maternal separation | [178] |
| miR-124-1 | pre miRNA upregulate expression in the medial prefrontal cortex during maternal separation | [178] |
| miR-128a | decreases expression in hippocampal due to treatment of lithium and sodium | [179] |
| miR-128b | regulate the formation of dear extinction memory in the prefrontal cortex. | [182] |
| miR-132 | upregulate expression in the medial prefrontal cortex during maternal separation | [178] |
| miR-134 | form synaptic plasticity in hippocampus | [183] |
| miR-138-2 | SNPs associated with panic disorder | [179] |
| miR-144 | decrease expression in the hippocampus due to treatment of sodium, lithium valproate | [179] |
| miR-148a | SNPs associated with panic disorder | [180] |
8. Pharmacological and Therapeutical Treatments for Depression and Related Disorders
8.1. Involvment of Selective Serotonin Reuptake Inhibitors in Depression
Selective serotonin reuptake inhibitors (SSRIs) such as fluoxetine, sertraline, citalopram, escitalopram, and fluvoxamine etc., are commonly used antidepressants agents [184]. These inhibitors are used in both moderate to severe depression and are relatively safe and typically cause fewer side effects [185]. These SSRIs inhibit the reuptake of serotonin by blocking serotonin transporter which result in increasing levels of serotonin available to bind to postsynaptic receptors [186,187]. Another rat study showed that depression can be controlled through the inhibition of SERT protein via miR-16 and as a result it has inhibited the reuptake of 5-HT in the hippocampus and serum [188]. Withdrawal of SSRIs can result in temporary deficiency of synaptic serotonin which in turn may lead to unpleasant symptoms such as headache, nausea, vomiting, and sleep disturbance [189].
It has also been observed that some miRs can regulate bacterial growth and gene transcription while also modulating the gut microbiota composition, suggesting the importance of miRs in gut and brain health. However, there are different treatment and prevention strategies for neuropsychiatric diseases, such as physical exercise, diet, and probiotics, that can modulate the gut microbiota composition and miRNAs expressions. The recent findings of the potential roles of microbiota and miR on the neuropathology of depression and anxiety, and its potential as treatment strategies [190].
8.2. Norepinephrine Reuptake Inhibitors
Norepinephrine reuptake inhibitors (NRIs) are drugs that act as a reuptake inhibitor for the noradrenaline and epinephrine by blocking the action of the norepinephrine transporter (NET) at the synapses and between nerve cells [191]. Norepinephrine is a naturally occurring neurotransmitter, or brain chemical, in the CNS that increases alertness and reaction time. Moreover, norepinephrine also plays a significant role in a person’s mood. It has been observed that low levels of norepinephrine can result in physical and mental symptoms, such as anxiety, depression, changes in blood pressure and heart rate, hypoglycemia, migraine and sleeping problems. NRI drugs are typically prescribed to patients who would benefit from increased levels of norepinephrine. Therefore, the list of NRIs which can be used in depression related symptoms includes atomoxetine (Strattera) for attention-deficit hyperactivity disorder (ADHD) and bupropion (Wellbutrin, Forfivo and Aplenzin) used for major depressive disorder (MDD).
8.3. Tricyclic Antidepressants
Tricyclic antidepressants (TCAs) are named due to their core chemical structure with connected rings [192,193]. TCAs mechanism of action is not straightforward as like SNRIs. TCAs are primarily inhibiting the reuptake of both serotonin and norepinephrine by blocking both transporters [194]. Therefore, it has also been seen that TCAs: including amitriptyline, amoxapine, clomipramine, desipramine, doxepin, imipramine, maprotiline, nortriptyline, and protriptyline are involved in the blockade of other receptors and are thought to be responsible for their side effects more than their antidepressant activity [195]. TCAs are mainly used for depression due to their broad mechanism of action, they also proved to be beneficial in the treatment of other medical problems for example amitriptyline and nortriptyline have been used for migraine prevention as well as treatment of neuropathic pain, on the other hand TCAs such as doxepin have been used for insomnia [196,197]. TCAs block cardiac sodium channels and produce effects like antiarrhythmic agents such as quinidine this ultimately can lead to cardiac conduction abnormalities [198,199,200].
8.4. Monoamine Oxidase Inhibitors
Monoamine oxidase inhibitors (MAOIs) are antidepressants drugs commonly used in depression [201]. MOAs are the mitochondrial enzymes that degrade monoamines such as serotonin and norepinephrine [202]. Monoamine oxidase (MAO) possesses two types A and B which are distributed in the brain, gut, and liver tissues [203]. The MAO type A MAO-A preferentially metabolizes serotonin, norepinephrine and dopamine while MAO-B metabolizes dopamine, therefore the inhibition of MAO-A is thought to be responsible for antidepressant effects of MAOIs [204,205]. The primary mechanism of action of MAOIs is to inhibit the activity of MAO and preventing the breakdown of monoamine neurotransmitters and increase their availability. The MAOIs such as isocarboxazid, phenalgine, and tranylcypromine are irreversible inhibitors of both types A and B which in turn makes them effective for the treatment of depression. Another MAOI is selegiline which is a selective inhibitor for MAO-B and has been effective in reducing symptoms of Parkinson’s disease. MAOIs could be a good choice as the first or second-line antidepressants. But in practice, they are usually a very last choice, because MAOIs show not only a high incidence of drug-drug interaction but also drug-food interactions.
8.5. Atypical and Typical Antidepressants
Antidepressants are medications used to treat major depressive disorders, some anxiety disorders, some chronic pain conditions, and to help manage some addictions. Common side-effects of antidepressants include dry mouth, weight gain, dizziness, headaches, sexual dysfunction, and emotional blunting [206,207]. An atypical antidepressant is any antidepressant medication that acts in a manner that is different from that of most other antidepressants [208]. The most recommended atypical antidepressants are bupropion, mirtazapine, trazodone, nefazodone, vilazodone, and vortioxetine with slightly different mechanisms of action [209]. Bupropion is a weak norepinephrine and dopamine reuptake inhibitor used for depression and it is effective in reducing the nicotine cravings [210]. Mirtazapine is an α-2 receptor antagonist that blocks presynaptic α-2 receptor and increases noradrenergic and serotonergic neurotransmission. Moreover, mirtazapine also exhibits postsynaptic serotonin receptor blocking and antihistaminic effects [211,212]. Vilazodone is another antidepressant drug that has unique mechanism of action and partially stimulates serotonin receptors [213,214].
Another couple of drugs such as imipramine and trazodone are used for the treatment of generalized anxiety disorder (GAD) in controlled trials [215,216]. In other studies, it has been shown that imipramine is effective as chlordiazepoxide and reduces depression however, trazodone is less sedating but effective against GAD [217]. Venlafaxine is the first approved antidepressant drug for the treatment of GAD [218,219].
8.6. Lithium
Lithium is mood-stabilizing drug that has been used in medicine for a long time, initially it was prescribed for depression however, currently it is also being used for bipolar disorder [220,221]. However, it has also been observed that unfortunately lithium has a fairly narrow therapeutic index which means that minor changes in dose or its blood levels can lead to toxicity [222,223].
8.7. Pharmacological Interventions Used in Anxiety Treatments
Medications that are used in anxiety and various other disorders have been shown to modify levels of miRNAs in the brain, but miRNAs are not the original target [146]. It has been shown that long-term direction of various mood stabilizers led to change in the expression of miRNAs in rat’s hippocampus when compared with saline treatment [179]. Several miRNAs including miR-29a, let-7c, let-7b, mir-128a, mir24a, mir30c, mir34a, and mir221 show changes in expression levels in patients with chronic oral treatment with valproate (VPA) or lithium. These microRNAs show decreased expression in the hippocampus, but increased expression of mir-144. For treatment of some forms of anxiety, VPA has shown promise for beneficial treatment such as social anxiety disorder [224]. Investigation of miR-34a downstream target genes in hippocampus shows that lithium direction of miR-34a and VPA increases the expression of miR-34a target genes. The direction of mirR-34a precursor reduces the expression of metabotropic glutamate receptor 7 (GRM7) [225]. Studies show that various numbers of miRNAs are linked with related downstream effects of therapeutics drugs which may be commonly used in the treatment of anxiety and various other disorders. Antidepressant-like SSRIs are used as first-line treatment in various anxiety disorders [226]. It has been shown SSRIs increase the expression of miR-16 in raphe nucleus and decrease the expression in locus coeruleus in mice [102]. In raphe nucleus and locus coeruleus, the expression patterns are inversely related with the expression level of its target genes, which are serotonin transporters (SERT). Studies reveal that therapeutic effects of fluoxetine could be related to different expression levels of mir-16 in monoaminergic neurons which target SERT in locus coeruleus and raphe nucleus [102]. In a mouse model with PTSD, 1-month treatment of fluoxetine can reduce or decrease anxiety-related behavior and is related to long-term effects on the expression of microRNAs [227]. After 74 days the expression level of mir-1971 decreased in the prefrontal cortex in comparison to the control group. Studies have revealed that some other SSRIs show possible relation between their miRNA regulation and corresponding behavioral effects. Maternal separation and chronic stress increase anxiety and depression which are associated with decreased levels of mir-326 expression in striatum and nucleus accumbent [228]. In long-lasting treatment, mir-326 levels increased in both areas of the brain compared with those of non-stressed or saline-treated rats. Mir-135a targets the serotonergic system and is attached to the serotonin transporter and 5HT-1a receptor gene [144]. Overexpression of mir-135a in serotonergic neurons is related to decreased anxiety and depression-like behaviors and knockdown can increase anxiety and depression [144]. Enriched microRNAs in the brain can target the GRM4 gene which can decrease expression when treated with antidepressants [229]. It has been shown that miRNA1202 plays an important role in antidepressant action by modifying glutamatergic neurotransmission. Major depressive disorder is related to decreased expression of miRNA1202 in the prefrontal cortex and the blood [230]. Therapeutic medication targeting other neurotransmitters and hormonal systems different than the serotonergic system have been analyzed for the contribution of miRNAs candidates in their antidepressant activities. Treatment with antidepressant agomelatine, which can target melatonin receptor and tianeptine glutamatergic, adenosine, and narcotic system, produced standardized stress-induced changes in miR levels in the dentate gyrus including the expression levels of miRNA-181b, miR-9, and miR-411 among others [231,232]. Treatment with the corticotropin-releasing hormone receptor 1 (CRHR1 also known as CRF1) antagonist NBI-27914 reduced the anxiety-like behavior of mice in elevated plus-maze and open field tests. miR-34b targets the CRHR1 genes and it was shown that administration of agomir for mir-34b in the paraventricular nucleus in the hypothalamus results in decreased anxiety behavior [233]. To target GABAergic neurotransmission, which can cause changes in microRNA expression, GABA receptor agonist gaboxadol was directed, which led to decreased anxiety-like behavior and increased expression of mir33 in the dorsal hippocampus [146]. It has been shown that NMDA-agonist D-cycloserine (DCS) which acts like an agonist at low doses but has antagonistic features with high doses, can be used to treat anxiety-like disorders [234].
8.8. Mechanism of Action of Rapid-Acting Antidepressant Ketamine in the Medial Prefrontal Cortex
Ketamine generates a glutamate burst by disinhibiting GABAergic interneurons [235]. The tonic firing of these GABA interneurons is controlled by NMDA receptors, and the active, open channel state allows ketamine to enter and block channel function. The glutamate burst that follows activates AMPA receptors, resulting in depolarization and activation. BDNF is released and TrkB is stimulated because of the activation of voltage dependent Ca2+ channels [236]. This promotes mTORC1 signaling, increasing protein synthesis essential for survival, and development and creation of synapses (GluA1 and PSD95). BDNF is produced in situations where it is needed, the release is inhibited or neutralized by a mutant BDNF Val/Met knock-in mice or BDNF neutralizing antibody. When mTORC1 signaling is inhibited (e.g., by rapamycin infusion into the mPFC), synaptic and neuronal activity decreases. Ketamine’s behavioral effects are inhibited. Scopolamine also triggers a glutamate surge in GABA interneurons by blocking acetylcholine muscarinic M1 (Ach-M1) receptors [237]. Antagonists of glutamate metabotropic 2/3 receptors (mGluR2/3) also cause a rapid increase in glutamate levels (Figure 7). Blocking presynaptic autoreceptors, which inhibits the release of dopamine, has antidepressant effects [238]. Glutamate relapse into depression is linked to a loss of synapses in the mPFC [239]. Stress and endocrine (cortisol) and estrogen imbalances have been linked to the death of neurons. Inflammatory cytokines, metabolic, and cardiovascular diseases are all linked to inflammatory cytokines [240].
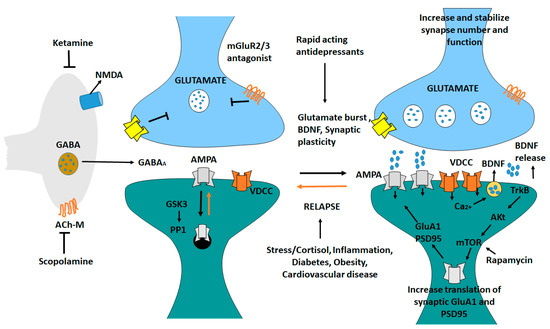
Figure 7.
Antidepressant ketamine pathway. Ketamine causes a burst of glutamate which stimulates AMPA receptors causing depolarization and activation of voltage dependent Ca2+ channels, leading to release of BDNF and stimulation of TrkB-Akt that activates mTORC1 signaling leading to increased synthesis of proteins required for synapse maturation and formation. In other conditions where BDNF release is blocked the synaptic actions of ketamine are blocked. Scopolamine also causes a glutamate burst via blockade of acetylcholine muscarinic M1 (ACh-M1) receptors on GABA interneurons [239].
8.9. Pharmacotherapy and Psychotherapy for Generalized Anxiety Disorder
The combination of pharmacotherapy and psychotherapy is used in the treatment of generalized anxiety disorder (GAD), which results in the reduction of symptoms, disability and improves health-related quality of life. Antidepressants, such as several benzodiazepines, buspirone, are used in the treatment of GAD. SSRIs and SNRIs are also used as first-line pharmacotherapies for generalized anxiety disorder [241]. Evidence is shown that SSRIs and SNRIs can be used for a treatment of anxiety disorder in children and adolescents [242]. Medication is provided to those children and adolescents only when the psychological approaches have failed. Control trials evaluated several psychotherapeutic techniques for GAD which include cognitive behavioral therapy, psychodynamic therapies, mindfulness-based therapies, relaxation therapy, with cognitive-behavioral therapy being the most successful [243]. Benzodiazepines are used for short-term relief of anxiety [244]. Evidence has shown that benzodiazepines are effective to reduce anxiety for a short period of time and there is no evidence that these drugs can work for a long period. As compared with other agents which were used to treat anxiety disorder, they are safe, rapid acting and have fewer side effects. Currently available benzodiazepines are effective for GAD. About 2/3 of patients experience improvement by using this drug within 1–2 weeks of treatment [215].
8.10. Obsessive-Compulsive Disorder and Diagnosis
The obsessive-compulsive disorder (OCD) is commonly chronic and long-lasting. In this disorder, a person has uncontrollable thoughts (obsession) [245,246]. Obsession means repetition of something in the mind which produces worry, and compulsive behavior in which the person repeats things, such as washing hands, cleaning home even if it is clean, etc. [247]. There are many types of neurotransmitters present in the brain, but serotonin is the most important neurotransmitter. When the levels of serotonin decrease in the body then the symptoms of obsessive-compulsive disorder increase. In our brain, there is a circuit which is the corticosteroid thalamic cortical circuit which is involved in connecting our lower body. Evidence is shown that if there is any dysfunction in this circuit then the symptoms of OCD are increased [248]. The combinatorial approach (medicines and psychotherapy) is important to cure these symptoms. Medicines which are selective serotonin reuptake inhibitors SSRIs are used to increase the level of serotonin. Some other medicines that can overcome anxiety for a short time are also used. The combination of medication and psychotherapy provides the best results [249].
8.11. Social Anxiety Disorder
This disorder, also known as social phobia it is a mental health condition [250]. It is an instant and continuous condition of fear when other people watch and judge the person. It is a chronic disorder. Onset occurs in children and adolescents (mid-teens) [251]. High-risk factors of social anxiety disorder are more prevalent in females as compared to males. Another factor is a family history of a social anxiety disorder (genetically or environmentally), shyness during childhood, behaviorally inhibited temperament, difficult unpleasant experiences during early childhood (child abuse or something that happened traumatically during childhood), and personality traits [252]. Any minor criticism is very seriously overreacted. These are associated frequently with other medical/psychiatric conditions such as major depressive disorder and substance abuse. Dysregulation of serotonin, or depletion of serotonin may increase the autonomic arousal by the increase of glutamate levels and increase of hypothalamic pituitary adrenal axis functioning which increases levels of stress hormones like activation of cortisol, and increased activity in the limbic fear circuit which increases activation of the amygdala [253]. The amygdala is the fear center in the brain. In adults fear or anxiety about one or more social situations include social interaction (conversation, meeting unfamiliar person), being observed (eating or drinking), performing in front of others (giving a speech). In children fear and anxiety occur in the situation of social interaction with other children but not with adults [254].
8.12. Social Anxiety Disorder and Medication
Usually, an individual with such disorder is fine with social interaction and shows only symptoms during performance situations. Treatment depends on social anxiety type, generally, we can use psychological treatments like cognitive behavioral therapy which could help in performance [255]. Pharmacologically we use selective serotonin reuptake inhibitors SSRIs (paroxetine, sertraline fluoxetine) serotonin-norepinephrine reuptake inhibitor SNRI (venlafaxine), or monoamine oxidase inhibitors MAOIs (phenalgine). In social anxiety disorder, performance type beta-blockers or benzodiazepines in an acute situation are used for short period (public speaking) [256].
8.13. Panic Disorder
Panic disorder is the type of anxiety disorder that causes panic attacks [257]. The person feels sudden terror even there is no real danger. The person feels losing control, with physical symptoms such as palpitations (an increase of heart rate), abdominal pain without any reason, nausea, intense fear of death, chest pain without any cardiac cause. Panic disorder is most common in students, it causes shortness of breath, trembling (shivering), sweating, and swearing [258]. For the patient with the acute condition, the benzodiazepine medication (BZD) is prescribed. This medicine is not for the chronic patient because BZD is highly addictive and has side effects. In chronic conditions, the selective serotonin reuptake inhibitor (SSRI) medication is prescribed together with psychotherapy (cognitive behavior therapy) which is highly effective for patient recovery. When the patient is facing a chronic condition increased amounts of panic attacks lead to depression, patient becomes agoraphobic being afraid of going into crowded areas, and having panic attacks in public. The frequent outcome of this disorder is substance abuse by taking alcohol, drugs and increased risk of suicide [259,260].
8.14. Bipolar Depression
Antidepressants can be divided into different classes such as selective serotonin reuptake inhibitors (SSRIs), serotonin/norepinephrine reuptake inhibitors (SNRIs), tricyclic antidepressants (TCAs), monoamine oxidase inhibitors (MAOIs) and atypical antidepressants [261]. Antidepressants differ in the way they work on presynaptic serotonergic neurons or serotonin-producing neurons, on the other hand presynaptic noradrenergic neurons or norepinephrine-producing neurons interact with corresponding postsynaptic neurons. The postsynaptic receptors of noradrenergic neurons are beta and alpha-1 while postsynaptic receptors of serotonergic neurons are serotonin 5-HT receptors; there are few subtypes of serotonin receptors present. Serotonin is synthesized from an amino acid tryptophan by serotonergic neurons and stored in vesicles [262,263]. On the other side norepinephrine is synthesized from an amino acid tyrosine by noradrenergic neurons and it is also stored in their vesicles for release. When serotonin and norepinephrine get released, they begin to stimulate the receptors and at the same time, they are transported from the synapse back to their neurons in a process called reuptake. Serotonin is reabsorbed by serotonin transporter (SERT) while norepinephrine is reabsorbed by norepinephrine transporter (NET) [264,265]. Once the serotonin and norepinephrine get reabsorbed back to their neurons they are partially repackaged into synaptic vesicles and partially broken down into inactive metabolites by an enzyme monoamine oxidase (MAO).
9. Conclusions and Future Prospectus
miRNAs have been shown to play a variety of roles in the development, progression, and treatment of depression and the related disorders. In this review, we had highlighted multiple miRNAs such as miR15a, miR17-92, miR34, miR-101, miR-124, miR-135, and miR-155 (Table 1) that may act as key players in treatment of depression through activation and deactivation of signaling pathways. The collected data indicate that, peripheral miRNAs levels are prone to dysregulation such as miR-320a and miR-335 are significantly downregulated, while miR-451a and miR-124-3p are significantly upregulated in the depression patients. Controlling the expression of miRNAs is considered a possible way to treat depression, as this interaction is believed to be highly relevant. Therefore, interaction between of miRs and gene encoded proteins has led to the identification of signaling pathways like neurotrophic signaling, mTOR signaling, and PI3k/AKT involved in depression. In future studies, miRs could be used as new biomarkers for the treatment of depression by controlling different signaling pathways.
Author Contributions
Conceptualization, M.H. and A.K., investigation, M.H., S.S. and A.A., resources, M.H., S.S. and A.A., writing—original draft preparation, M.H., S.S. and A.A., writing—review and editing, M.H. and A.K., project administration, M.H. and A.K., funding acquisition, A.K. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support from NSF grant DBI1661391, and NIH Grants R01 GM127701, and R01HG012117.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
M.H. acknowledges The Ohio State University for “President’s Postdoctoral Scholars Program (PPSP)” fellowship for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dias, B.G.; Banerjee, S.B.; Goodman, J.V.; Ressler, K.J. Towards new approaches to disorders of fear and anxiety. Curr. Opin. Neurobiol. 2013, 23, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Australian Bureau of Statistics. National Survey of Mental Health and Wellbeing: Summary of Results; Australian Bureau of Statistics: Canberra, Australia, 2007. [Google Scholar]
- Zinbarg, R.E.; Barlow, D.H. Structure of anxiety and the anxiety disorders: A hierarchical model. J. Abnorm. Psychol. 1996, 105, 181. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Wang, P.S. The descriptive epidemiology of commonly occurring mental disorders in the United States. Annu. Rev. Public Health 2008, 29, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Beebe, J.E. A nation at risk: Bullying among children & adolescents. In Youth at Risk: A Prevention Resource for Counselors, Teachers, & Parents; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 441–464. [Google Scholar]
- Asken, M.J.; Grossman, D.; Christensen, L.W. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. Arlington, VA: American Psychiatric Pub-lishing, 2013. Archibald, Herbert C., and Read D. Tuddenham.“Persistent Stress Reac-tion after Combat: A 20-Year Follow-Up.” Archives of General Psy. Therapy 2007, 45, 2317–2325. [Google Scholar]
- Hill, J.L.; Martinowich, K. Activity-dependent signaling: Influence on plasticity in circuits controlling fear-related behavior. Curr. Opin. Neurobiol. 2016, 36, 59–65. [Google Scholar] [CrossRef]
- Mayur, P.; Murat, A.; Malone, D.A., Jr.; Anand, A. Where in the brain is depression? Curr. Psychiatry Rep. 2012, 14, 634–642. [Google Scholar]
- Calhoon, G.G.; Tye, K.M. Resolving the neural circuits of anxiety. Nat. Neurosci. 2015, 18, 1394–1404. [Google Scholar] [CrossRef]
- Martin, E.I.; Ressler, K.J.; Binder, E.; Nemeroff, C.B. The neurobiology of anxiety disorders: Brain imaging, genetics, and psychoneuroendocrinology. Psychiatr. Clin. 2009, 32, 549–575. [Google Scholar] [CrossRef]
- MacQueen, G.M.; Yucel, K.; Taylor, V.H.; Macdonald, K.; Joffe, R. Posterior hippocampal volumes are associated with remission rates in patients with major depressive disorder. Biol. Psychiatry 2008, 64, 880–883. [Google Scholar] [CrossRef]
- Savitz, J.; Drevets, W.C. Bipolar and major depressive disorder: Neuroimaging the developmental-degenerative divide. Neurosci. Biobehav. Rev. 2009, 33, 699–771. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K. Synaptic dysfunction in depression: Potential therapeutic targets. Science 2012, 338, 68–72. [Google Scholar] [CrossRef]
- McEwen, B.S.; Eiland, L.; Hunter, R.G.; Miller, M.M. Stress and anxiety: Structural plasticity and epigenetic regulation as a consequence of stress. Neuropharmacology 2012, 62, 3–12. [Google Scholar] [CrossRef]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef]
- Sanacora, G.; Zarate, C.A.; Krystal, J.H.; Manji, H.K. Targeting the glutamatergic system to develop novel, improved therapeutics for mood disorders. Nat. Rev. Drug Discov. 2008, 7, 426–437. [Google Scholar] [CrossRef]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The stressed synapse: The impact of stress and glucocorticoids on glutamate transmission. Nat. Rev. Neurosci. 2012, 13, 22–37. [Google Scholar] [CrossRef]
- Hammen, C. Stress and depression. Annu. Rev. Clin. Psychol. 2005, 1, 293–319. [Google Scholar] [CrossRef]
- Sun, H.; Kennedy, P.J.; Nestler, E.J. Epigenetics of the depressed brain: Role of histone acetylation and methylation. Neuropsychopharmacology 2013, 38, 124–137. [Google Scholar] [CrossRef]
- Menke, A.; Binder, E.B. Epigenetic alterations in depression and antidepressant treatment. Dialogues Clin. Neurosci. 2014, 16, 395. [Google Scholar] [CrossRef]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Manji, H.K.; Drevets, W.C.; Charney, D.S. The cellular neurobiology of depression. Nat. Med. 2001, 7, 541–547. [Google Scholar] [CrossRef]
- Nemeroff, C.B.; Owens, M.J. Treatment of mood disorders. Nat. Neurosci. 2002, 5, 1068–1070. [Google Scholar] [CrossRef]
- Liu, R.-J.; Aghajanian, G.K. Stress blunts serotonin-and hypocretin-evoked EPSCs in prefrontal cortex: Role of corticosterone-mediated apical dendritic atrophy. Proc. Natl. Acad. Sci. USA 2008, 105, 359–364. [Google Scholar] [CrossRef]
- Magarin, A.; McEwen, B. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: Involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 1995, 69, 89–98. [Google Scholar] [CrossRef]
- Lowy, M.T.; Wittenberg, L.; Yamamoto, B.K. Effect of acute stress on hippocampal glutamate levels and spectrin proteolysis in young and aged rats. J. Neurochem. 1995, 65, 268–274. [Google Scholar] [CrossRef]
- Moghaddam, B.; Adams, B.; Verma, A.; Daly, D. Activation of glutamatergic neurotransmission by ketamine: A novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J. Neurosci. 1997, 17, 2921–2927. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Labonté, B.; Wen, X.L.; Turecki, G.; Meaney, M.J. Epigenetic mechanisms for the early environmental regulation of hippocampal glucocorticoid receptor gene expression in rodents and humans. Neuropsychopharmacology 2013, 38, 111–123. [Google Scholar] [CrossRef]
- Hosang, G.M.; Shiles, C.; Tansey, K.E.; McGuffin, P.; Uher, R. Interaction between stress and the BDNF Val66Met polymorphism in depression: A systematic review and meta-analysis. BMC Med. 2014, 12, 7. [Google Scholar] [CrossRef]
- Holtmaat, A.; Svoboda, K. Experience-dependent structural synaptic plasticity in the mammalian brain. Nat. Rev. Neurosci. 2009, 10, 647–658. [Google Scholar] [CrossRef]
- Jourdi, H.; Hsu, Y.-T.; Zhou, M.; Qin, Q.; Bi, X.; Baudry, M. Positive AMPA receptor modulation rapidly stimulates BDNF release and increases dendritic mRNA translation. J. Neurosci. 2009, 29, 8688–8697. [Google Scholar] [CrossRef]
- Bocchio-Chiavetto, L.; Bagnardi, V.; Zanardini, R.; Molteni, R.; Gabriela Nielsen, M.; Placentino, A.; Giovannini, C.; Rillosi, L.; Ventriglia, M.; Riva, M.A. Serum and plasma BDNF levels in major depression: A replication study and meta-analyses. World J. Biol. Psychiatry 2010, 11, 763–773. [Google Scholar] [CrossRef]
- Turner, C.A.; Akil, H.; Watson, S.J.; Evans, S.J. The fibroblast growth factor system and mood disorders. Biol. Psychiatry 2006, 59, 1128–1135. [Google Scholar] [CrossRef]
- Phillips, C. Brain-derived neurotrophic factor, depression, and physical activity: Making the neuroplastic connection. Neural Plast. 2017, 2017, 7260130. [Google Scholar] [CrossRef]
- Bueller, J.A.; Aftab, M.; Sen, S.; Gomez-Hassan, D.; Burmeister, M.; Zubieta, J.-K. BDNF Val66Met allele is associated with reduced hippocampal volume in healthy subjects. Biol. Psychiatry 2006, 59, 812–815. [Google Scholar] [CrossRef]
- Chen, H.; Pandey, G.N.; Dwivedi, Y. Hippocampal cell proliferation regulation by repeated stress and antidepressants. Neuroreport 2006, 17, 863–867. [Google Scholar] [CrossRef]
- Liu, R.-J.; Fuchikami, M.; Dwyer, J.M.; Lepack, A.E.; Duman, R.S.; Aghajanian, G.K. GSK-3 inhibition potentiates the synaptogenic and antidepressant-like effects of subthreshold doses of ketamine. Neuropsychopharmacology 2013, 38, 2268–2277. [Google Scholar] [CrossRef]
- Magarinos, A.; Li, C.; Gal Toth, J.; Bath, K.; Jing, D.; Lee, F.; McEwen, B. Effect of brain-derived neurotrophic factor haploinsufficiency on stress-induced remodeling of hippocampal neurons. Hippocampus 2011, 21, 253–264. [Google Scholar] [CrossRef]
- Autry, A.E.; Adachi, M.; Cheng, P.; Monteggia, L.M. Gender-specific impact of brain-derived neurotrophic factor signaling on stress-induced depression-like behavior. Biol. Psychiatry 2009, 66, 84–90. [Google Scholar] [CrossRef]
- Duman, C.H.; Schlesinger, L.; Kodama, M.; Russell, D.S.; Duman, R.S. A role for MAP kinase signaling in behavioral models of depression and antidepressant treatment. Biol. Psychiatry 2007, 61, 661–670. [Google Scholar] [CrossRef]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef]
- Minichiello, L. TrkB signalling pathways in LTP and learning. Nat. Rev. Neurosci. 2009, 10, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Watson, K.; Baar, K. mTOR and the health benefits of exercise. Semin. Cell Dev. Biol. 2014, 36, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J.M.; Duman, R.S. Activation of mammalian target of rapamycin and synaptogenesis: Role in the actions of rapid-acting antidepressants. Biol. Psychiatry 2013, 73, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Feyissa, A.M.; Chandran, A.; Stockmeier, C.A.; Karolewicz, B. Reduced levels of NR2A and NR2B subunits of NMDA receptor and PSD-95 in the prefrontal cortex in major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Jernigan, C.S.; Goswami, D.B.; Austin, M.C.; Iyo, A.H.; Chandran, A.; Stockmeier, C.A.; Karolewicz, B. The mTOR signaling pathway in the prefrontal cortex is compromised in major depressive disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1774–1779. [Google Scholar] [CrossRef]
- Li, N.; Lee, B.; Liu, R.-J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.-Y.; Aghajanian, G.; Duman, R.S. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 2010, 329, 959–964. [Google Scholar] [CrossRef]
- Voleti, B.; Navarria, A.; Liu, R.-J.; Banasr, M.; Li, N.; Terwilliger, R.; Sanacora, G.; Eid, T.; Aghajanian, G.; Duman, R.S. Scopolamine rapidly increases mammalian target of rapamycin complex 1 signaling, synaptogenesis, and antidepressant behavioral responses. Biol. Psychiatry 2013, 74, 742–749. [Google Scholar] [CrossRef]
- Ellisen, L.W.; Ramsayer, K.D.; Johannessen, C.M.; Yang, A.; Beppu, H.; Minda, K.; Oliner, J.D.; McKeon, F.; Haber, D.A. REDD1, a developmentally regulated transcriptional target of p63 and p53, links p63 to regulation of reactive oxygen species. Mol. Cell 2002, 10, 995–1005. [Google Scholar] [CrossRef]
- Sofer, A.; Lei, K.; Johannessen, C.M.; Ellisen, L.W. Regulation of mTOR and cell growth in response to energy stress by REDD1. Mol. Cell. Biol. 2005, 25, 5834–5845. [Google Scholar] [CrossRef]
- Shoshani, T.; Faerman, A.; Mett, I.; Zelin, E.; Tenne, T.; Gorodin, S.; Moshel, Y.; Elbaz, S.; Budanov, A.; Chajut, A. Identification of a novel hypoxia-inducible factor 1-responsive gene, RTP801, involved in apoptosis. Mol. Cell. Biol. 2002, 22, 2283–2293. [Google Scholar] [CrossRef]
- Katiyar, S.; Liu, E.; Knutzen, C.A.; Lang, E.S.; Lombardo, C.R.; Sankar, S.; Toth, J.I.; Petroski, M.D.; Ronai, Z.e.; Chiang, G.G. REDD1, an inhibitor of mTOR signalling, is regulated by the CUL4A–DDB1 ubiquitin ligase. EMBO Rep. 2009, 10, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Chandran, A.; Iyo, A.H.; Jernigan, C.S.; Legutko, B.; Austin, M.C.; Karolewicz, B. Reduced phosphorylation of the mTOR signaling pathway components in the amygdala of rats exposed to chronic stress. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 40, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, R.-J.; Dwyer, J.M.; Banasr, M.; Lee, B.; Son, H.; Li, X.-Y.; Aghajanian, G.; Duman, R.S. Glutamate N-methyl-D-aspartate receptor antagonists rapidly reverse behavioral and synaptic deficits caused by chronic stress exposure. Biol. Psychiatry 2011, 69, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.E.; Choi, D.C.; Ressler, K.J. Neuropeptide regulation of fear and anxiety: Implications of cholecystokinin, endogenous opioids, and neuropeptide Y. Physiol. Behav. 2012, 107, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Kalia, M. Neurobiological basis of depression: An update. Metabolism 2005, 54, 24–27. [Google Scholar] [CrossRef]
- Van Moffaert, M.; Dierick, M. Noradrenaline (norepinephrine) and depression. CNS Drugs 1999, 12, 293–305. [Google Scholar] [CrossRef]
- Chrousos, G.P. The stress response and immune function: Clinical implications. Ann. N. Y. Acad. Sci. 2000, 917, 38–67. [Google Scholar] [CrossRef]
- Southwick, S.M.; Bremner, J.D.; Rasmusson, A.; Morgan III, C.A.; Arnsten, A.; Charney, D.S. Role of norepinephrine in the pathophysiology and treatment of posttraumatic stress disorder. Biol. Psychiatry 1999, 46, 1192–1204. [Google Scholar] [CrossRef]
- Young, S.N. How to increase serotonin in the human brain without drugs. J. Psychiatry Neurosci. 2007, 32, 394–399. [Google Scholar]
- Deakin, J. The role of serotonin in panic, anxiety and depression. Int. Clin. Psychopharmacol. 1998, 13 (Suppl. 4), S1–S5. [Google Scholar] [CrossRef]
- Felten, D.L.; O’Banion, M.K.; Maida, M.E. Netter’s Atlas of Neuroscience; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Roth, B.L. Multiple serotonin receptors: Clinical and experimental aspects. Ann. Clin. Psychiatry 1994, 6, 67–78. [Google Scholar] [CrossRef]
- Roth, B.L.; Lopez, E.; Patel, S.; Kroeze, W.K. The multiplicity of serotonin receptors: Uselessly diverse molecules or an embarrassment of riches? Neuroscientist 2000, 6, 252–262. [Google Scholar] [CrossRef]
- Legangneux, E.; Mora, J.; Spreux-Varoquaux, O.; Thorin, I.; Herrou, M.; Alvado, G.; Gomeni, C. Cerebrospinal fluid biogenic amine metabolites, plasma-rich platelet serotonin and [3H] imipramine reuptake in the primary fibromyalgia syndrome. Rheumatology 2001, 40, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Kolachana, B.; Fera, F.; Goldman, D.; Egan, M.F.; Weinberger, D.R. Serotonin transporter genetic variation and the response of the human amygdala. Science 2002, 297, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in motivational control: Rewarding, aversive, and alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.M.; Geyer, H.; Milstein, M.J. Kaufman’s Clinical Neurology for Psychiatrists; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Wang, G.-J.; Volkow, N.D.; Thanos, P.K.; Fowler, J.S. Imaging of brain dopamine pathways: Implications for understanding obesity. J. Addict. Med. 2009, 3, 8. [Google Scholar] [CrossRef]
- Aso, Y.; Herb, A.; Ogueta, M.; Siwanowicz, I.; Templier, T.; Friedrich, A.B.; Ito, K.; Scholz, H.; Tanimoto, H. Three dopamine pathways induce aversive odor memories with different stability. PLoS Genet. 2012, 8, e1002768. [Google Scholar] [CrossRef]
- Tritsch, N.X.; Ding, J.B.; Sabatini, B.L. Dopaminergic neurons inhibit striatal output through non-canonical release of GABA. Nature 2012, 490, 262–266. [Google Scholar] [CrossRef]
- Brown, A.; Gershon, S. Dopamine and depression. J. Neural Transm. /Gen. Sect. JNT 1993, 91, 75–109. [Google Scholar] [CrossRef]
- Fritschy, J.M.; Panzanelli, P. GABAA receptors and plasticity of inhibitory neurotransmission in the central nervous system. Eur. J. Neurosci. 2014, 39, 1845–1865. [Google Scholar] [CrossRef]
- Kim, J.; Gorman, J. The psychobiology of anxiety. Clin. Neurosci. Res. 2005, 4, 335–347. [Google Scholar] [CrossRef]
- Kendell, S.F.; Krystal, J.H.; Sanacora, G. GABA and glutamate systems as therapeutic targets in depression and mood disorders. Expert Opin. Ther. Targets 2005, 9, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Nusser, Z.; Sieghart, W.; Somogyi, P. Segregation of different GABAA receptors to synaptic and extrasynaptic membranes of cerebellar granule cells. J. Neurosci. 1998, 18, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Skórzewska, A.; Lehner, M.; Wisłowska-Stanek, A.; Krząścik, P.; Ziemba, A.; Płaźnik, A. The effect of chronic administration of corticosterone on anxiety-and depression-like behavior and the expression of GABA-A receptor alpha-2 subunits in brain structures of low-and high-anxiety rats. Horm. Behav. 2014, 65, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, B.S.; Rogawski, M.A. Molecular targets for antiepileptic drug development. Neurotherapeutics 2007, 4, 18–61. [Google Scholar] [CrossRef] [PubMed]
- Lydiard, R.B. The role of GABA in anxiety disorders. J. Clin. Psychiatry 2003, 64, 21–27. [Google Scholar] [PubMed]
- Rudolph, U.; Knoflach, F. Beyond classical benzodiazepines: Novel therapeutic potential of GABAA receptor subtypes. Nat. Rev. Drug Discov. 2011, 10, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Nutt, D.J. Role of GABA in anxiety and depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar] [CrossRef]
- Hunsberger, J.G.; Austin, D.R.; Chen, G.; Manji, H.K. MicroRNAs in mental health: From biological underpinnings to potential therapies. Neuromolecular Med. 2009, 11, 173–182. [Google Scholar] [CrossRef][Green Version]
- Leucci, E.; Patella, F.; Waage, J.; Holmstrøm, K.; Lindow, M.; Porse, B.; Kauppinen, S.; Lund, A.H. microRNA-9 targets the long non-coding RNA MALAT1 for degradation in the nucleus. Sci. Rep. 2013, 3, 2535. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, A.; Patnala, R.; P Jadhav, S.; Eng-Ang, L.; Thameem Dheen, S. MicroRNAs: Key players in microglia and astrocyte mediated inflammation in CNS pathologies. Curr. Med. Chem. 2016, 23, 3528–3546. [Google Scholar] [CrossRef]
- Bicker, S.; Schratt, G. microRNAs: Tiny regulators of synapse function in development and disease. J. Cell. Mol. Med. 2008, 12, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Natera-Naranjo, O.; Aschrafi, A.; Gioio, A.E.; Kaplan, B.B. Identification and quantitative analyses of microRNAs located in the distal axons of sympathetic neurons. RNA 2010, 16, 1516–1529. [Google Scholar] [CrossRef]
- Antonucci, F.; Corradini, I.; Fossati, G.; Tomasoni, R.; Menna, E.; Matteoli, M. SNAP-25, a known presynaptic protein with emerging postsynaptic functions. Front. Synaptic Neurosci. 2016, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Thatcher, E.J.; Olena, A.F.; Cha, D.J.; Perdigoto, A.L.; Marshall, A.F.; Carter, B.D.; Broadie, K.; Patton, J.G. miR-153 regulates SNAP-25, synaptic transmission, and neuronal development. PLoS ONE 2013, 8, e57080. [Google Scholar] [CrossRef]
- Yan, M.-L.; Zhang, S.; Zhao, H.-M.; Xia, S.-N.; Jin, Z.; Xu, Y.; Yang, L.; Qu, Y.; Huang, S.-Y.; Duan, M.-J. MicroRNA-153 impairs presynaptic plasticity by blocking vesicle release following chronic brain hypoperfusion. Cell Commun. Signal. 2020, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Mannironi, C.; Biundo, A.; Rajendran, S.; De Vito, F.; Saba, L.; Caioli, S.; Zona, C.; Ciotti, T.; Caristi, S.; Perlas, E. miR-135a regulates synaptic transmission and anxiety-like behavior in amygdala. Mol. Neurobiol. 2018, 55, 3301–3315. [Google Scholar] [CrossRef]
- Earls, L.R.; Fricke, R.G.; Yu, J.; Berry, R.B.; Baldwin, L.T.; Zakharenko, S.S. Age-dependent microRNA control of synaptic plasticity in 22q11 deletion syndrome and schizophrenia. J. Neurosci. 2012, 32, 14132–14144. [Google Scholar] [CrossRef]
- Earls, L.R.; Zakharenko, S.S. A synaptic function approach to investigating complex psychiatric diseases. Neuroscientist 2014, 20, 257–271. [Google Scholar] [CrossRef]
- Higa, G.S.V.; de Sousa, E.; Walter, L.T.; Kinjo, E.R.; Resende, R.R.; Kihara, A.H. MicroRNAs in neuronal communication. Mol. Neurobiol. 2014, 49, 1309–1326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-f.; Chen, J.-C.; Zhang, J.; Xu, J.-G. miR-181a involves in the hippocampus-dependent memory formation via targeting PRKAA1. Sci. Rep. 2017, 7, 8480. [Google Scholar] [CrossRef] [PubMed]
- Saba, R.; Störchel, P.H.; Aksoy-Aksel, A.; Kepura, F.; Lippi, G.; Plant, T.D.; Schratt, G.M. Dopamine-regulated microRNA MiR-181a controls GluA2 surface expression in hippocampal neurons. Mol. Cell. Biol. 2012, 32, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, J.N.; Pochiraju, S.; Kannampalli, P.; Bruckert, M.; Addya, S.; Yadav, P.; Miranda, A.; Shaker, R.; Banerjee, B. MicroRNA-mediated GABAAα-1 receptor subunit down-regulation in adult spinal cord following neonatal cystitis-induced chronic visceral pain in rats. PAIN® 2013, 154, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhu, R.; Li, X. Silencing miR-181a produces neuroprotection against hippocampus neuron cell apoptosis post-status epilepticus in a rat model and in children with temporal lobe epilepsy. Genet. Mol. Res. 2016, 15, gmr.15017798. [Google Scholar] [CrossRef] [PubMed]
- Bliziotes, M.; Eshleman, A.; Zhang, X.-W.; Wiren, K. Neurotransmitter action in osteoblasts: Expression of a functional system for serotonin receptor activation and reuptake. Bone 2001, 29, 477–486. [Google Scholar] [CrossRef]
- Sangkuhl, K.; Klein, T.; Altman, R. Selective serotonin reuptake inhibitors (SSRI) pathway. Pharm. Genom. 2009, 19, 907. [Google Scholar] [CrossRef]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.-M.; Kellermann, O. miR-16 targets the serotonin transporter: A new facet for adaptive responses to antidepressants. Science 2010, 329, 1537–1541. [Google Scholar] [CrossRef]
- Zheng, Z.; Zeng, Y.; Huang, H.; Xu, F. MicroRNA-132 may play a role in coexistence of depression and cardiovascular disease: A hypothesis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2013, 19, 438. [Google Scholar]
- Shaked, I.; Meerson, A.; Wolf, Y.; Avni, R.; Greenberg, D.; Gilboa-Geffen, A.; Soreq, H. MicroRNA-132 potentiates cholinergic anti-inflammatory signaling by targeting acetylcholinesterase. Immunity 2009, 31, 965–973. [Google Scholar] [CrossRef]
- Shaltiel, G.; Hanan, M.; Wolf, Y.; Barbash, S.; Kovalev, E.; Shoham, S.; Soreq, H. Hippocampal microRNA-132 mediates stress-inducible cognitive deficits through its acetylcholinesterase target. Brain Struct. Funct. 2013, 218, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Yudowski, G.A.; Olsen, O.; Adesnik, H.; Marek, K.W.; Bredt, D.S. Acute inactivation of PSD-95 destabilizes AMPA receptors at hippocampal synapses. PLoS ONE 2013, 8, e53965. [Google Scholar] [CrossRef] [PubMed]
- Muddashetty, R.S.; Nalavadi, V.C.; Gross, C.; Yao, X.; Xing, L.; Laur, O.; Warren, S.T.; Bassell, G.J. Reversible inhibition of PSD-95 mRNA translation by miR-125a, FMRP phosphorylation, and mGluR signaling. Mol. Cell 2011, 42, 673–688. [Google Scholar] [CrossRef]
- Fan, C.; Li, Y.; Lan, T.; Wang, W.; Long, Y.; Yu, S.Y. Microglia secrete miR-146a-5p-containing exosomes to regulate neurogenesis in depression. Mol. Ther. 2022, 30, 1300–1314. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Shen, C.-K.J. Modulation of mGluR-dependent MAP1B translation and AMPA receptor endocytosis by microRNA miR-146a-5p. J. Neurosci. 2013, 33, 9013–9020. [Google Scholar] [CrossRef] [PubMed]
- Lotrich, F. Inflammatory cytokines, growth factors, and depression. Curr. Pharm. Des. 2012, 18, 5920–5935. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Nowacka, M.M.; Obuchowicz, E. Vascular endothelial growth factor (VEGF) and its role in the central nervous system: A new element in the neurotrophic hypothesis of antidepressant drug action. Neuropeptides 2012, 46, 1–10. [Google Scholar]
- Fournier, N.M.; Duman, R.S. Role of vascular endothelial growth factor in adult hippocampal neurogenesis: Implications for the pathophysiology and treatment of depression. Behav. Brain Res. 2012, 227, 440–449. [Google Scholar] [CrossRef]
- Blahna, M.T.; Hata, A. Regulation of miRNA biogenesis as an integrated component of growth factor signaling. Curr. Opin. Cell Biol. 2013, 25, 233–240. [Google Scholar]
- Lau, A.G.; Irier, H.A.; Gu, J.; Tian, D.; Ku, L.; Liu, G.; Xia, M.; Fritsch, B.; Zheng, J.Q.; Dingledine, R. Distinct 3′ UTRs differentially regulate activity-dependent translation of brain-derived neurotrophic factor (BDNF). Proc. Natl. Acad. Sci. USA 2010, 107, 15945–15950. [Google Scholar] [CrossRef] [PubMed]
- Caputo, V.; Sinibaldi, L.; Fiorentino, A.; Parisi, C.; Catalanotto, C.; Pasini, A.; Cogoni, C.; Pizzuti, A. Brain derived neurotrophic factor (BDNF) expression is regulated by microRNAs miR-26a and miR-26b allele-specific binding. PLoS ONE 2011, 6, e28656. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.-D.; He, X.-G.; Yang, F.-G.; Liu, M.-Q.; Wang, Y.-D.; Zhu, D.-X.; Zhang, G.-Z.; Ma, Z.-J.; Kang, X.-W. Research progress on the regulatory role of microRNAs in spinal cord injury. Regen. Med. 2021, 16, 465–476. [Google Scholar] [CrossRef]
- Thomas, K.T.; Gross, C.; Bassell, G.J. MicroRNAs sculpt neuronal communication in a tight balance that is lost in neurological disease. Front. Mol. Neurosci. 2018, 11, 455. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Chu, K.; Jung, K.H.; Kim, J.H.; Huh, J.Y.; Yoon, H.; Park, D.K.; Lim, J.Y.; Kim, J.M.; Jeon, D. miR-206 regulates brain-derived neurotrophic factor in Alzheimer disease model. Ann. Neurol. 2012, 72, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, A.; Bayerlová, M.; Strotbek, M.; Schmid, S.; Beissbarth, T.; Olayioye, M.A. A global microRNA screen identifies regulators of the ErbB receptor signaling network. Cell Commun. Signal. 2015, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A. Handbook of Biologically Active peptides; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Pradhan, J.; Noakes, P.G.; Bellingham, M.C. The role of altered BDNF/TrkB signaling in amyotrophic lateral sclerosis. Front. Cell. Neurosci. 2019, 13, 368. [Google Scholar] [CrossRef]
- Irmady, K.; Jackman, K.A.; Padow, V.A.; Shahani, N.; Martin, L.A.; Cerchietti, L.; Unsicker, K.; Iadecola, C.; Hempstead, B.L. Mir-592 regulates the induction and cell death-promoting activity of p75NTR in neuronal ischemic injury. J. Neurosci. 2014, 34, 3419–3428. [Google Scholar] [CrossRef]
- Salinas, F.V.; Malik, K.; Benzon, H.T. Local anesthetics for regional anesthesia and pain management. In Raj’s Practical Management of Pain; Elsevier Inc.: Amsterdam, The Netherlands, 2008; pp. 811–838. [Google Scholar]
- Raab-Graham, K.F.; Haddick, P.C.; Jan, Y.N.; Jan, L.Y. Activity-and mTOR-dependent suppression of Kv1. 1 channel mRNA translation in dendrites. Science 2006, 314, 144–148. [Google Scholar] [CrossRef]
- Gross, C.; Yao, X.; Engel, T.; Tiwari, D.; Xing, L.; Rowley, S.; Danielson, S.W.; Thomas, K.T.; Jimenez-Mateos, E.M.; Schroeder, L.M. MicroRNA-mediated downregulation of the potassium channel Kv4. 2 contributes to seizure onset. Cell Rep. 2016, 17, 37–45. [Google Scholar] [CrossRef]
- Rho, J.M.; Szot, P.; Tempel, B.L.; Schwartzkroin, P.A. Developmental seizure susceptibility of kv1. 1 potassium channel knockout mice. Dev. Neurosci. 1999, 21, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Barnwell, L.F.S.; Lugo, J.N.; Lee, W.L.; Willis, S.E.; Gertz, S.J.; Hrachovy, R.A.; Anderson, A.E. Kv4. 2 knockout mice demonstrate increased susceptibility to convulsant stimulation. Epilepsia 2009, 50, 1741–1751. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, D.; Peariso, K.; Gross, C. MicroRNA-induced silencing in epilepsy: Opportunities and challenges for clinical application. Dev. Dyn. 2018, 247, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Fernandes, C.C.; Ewell, L.A.; John, D.; Romoli, B.; Curia, G.; Taylor, S.R.; Frady, E.P.; Jensen, A.B.; Liu, J.C. MicroRNA-101 regulates multiple developmental programs to constrain excitation in adult neural networks. Neuron 2016, 92, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Petkov, G.V. Ion channels. In Pharmacology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 387–427. [Google Scholar]
- Guo, C.; Sah, J.F.; Beard, L.; Willson, J.K.; Markowitz, S.D.; Guda, K. The noncoding RNA, miR-126, suppresses the growth of neoplastic cells by targeting phosphatidylinositol 3-kinase signaling and is frequently lost in colon cancers. Genes Chromosomes Cancer 2008, 47, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Sessa, R.; Seano, G.; Di Blasio, L.; Gagliardi, P.A.; Isella, C.; Medico, E.; Cotelli, F.; Bussolino, F.; Primo, L. The miR-126 regulates angiopoietin-1 signaling and vessel maturation by targeting p85β. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2012, 1823, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, H.; Long, L.; Hui, L.; Chen, H.; Wang, X.; Shen, H.; Xu, W. miR-126 enhances the sensitivity of non-small cell lung cancer cells to anticancer agents by targeting vascular endothelial growth factor A. Acta Biochim. Biophys. Sin. 2012, 44, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Lee, Y.; McKenna, N.D.; Yi, M.; Simunovic, F.; Wang, Y.; Kong, B.; Rooney, R.J.; Seo, H.; Stephens, R.M. miR-126 contributes to Parkinson’s disease by dysregulating the insulin-like growth factor/phosphoinositide 3-kinase signaling. Neurobiol. Aging 2014, 35, 1712–1721. [Google Scholar] [CrossRef]
- Jiang, W.; Tang, Y.-Y.; Zhu, W.-W.; Li, C.; Zhang, P.; Li, R.-Q.; Chen, Y.-J.; Zou, W.; Tang, X.-Q. PI3K/AKT pathway mediates the antidepressant-and anxiolytic-like roles of hydrogen sulfide in streptozotocin-induced diabetic rats via promoting hippocampal neurogenesis. NeuroToxicology 2021, 85, 201–208. [Google Scholar] [CrossRef]
- Karege, F.; Perroud, N.; Burkhardt, S.; Fernandez, R.; Ballmann, E.; La Harpe, R.; Malafosse, A. Alterations in phosphatidylinositol 3-kinase activity and PTEN phosphatase in the prefrontal cortex of depressed suicide victims. Neuropsychobiology 2011, 63, 224–231. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Exportin 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [PubMed]
- Velu, V.K.; Ramesh, R.; Srinivasan, A.R. Circulating microRNAs as biomarkers in health and disease. J. Clin. Diagn. Res. JCDR 2012, 6, 1791. [Google Scholar] [CrossRef] [PubMed]
- Kye, M.J.; Niederst, E.D.; Wertz, M.H.; Gonçalves, I.d.C.G.; Akten, B.; Dover, K.Z.; Peters, M.; Riessland, M.; Neveu, P.; Wirth, B. SMN regulates axonal local translation via miR-183/mTOR pathway. Hum. Mol. Genet. 2014, 23, 6318–6331. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.L.; Plotkin, J.L.; Venø, M.T.; Von Schimmelmann, M.; Feinberg, P.; Mann, S.; Handler, A.; Kjems, J.; Surmeier, D.J.; O’Carroll, D. MicroRNA-128 governs neuronal excitability and motor behavior in mice. Science 2013, 342, 1254–1258. [Google Scholar] [CrossRef]
- Dwivedi, Y.; Zhang, H. Altered ERK1/2 signaling in the brain of learned helpless rats: Relevance in vulnerability to developing stress-induced depression. Neural Plast. 2016, 2016, 7383724. [Google Scholar] [CrossRef]
- Kuss, A.W.; Chen, W. MicroRNAs in brain function and disease. Curr. Neurol. Neurosci. Rep. 2008, 8, 190–197. [Google Scholar] [CrossRef]
- Issler, O.; Haramati, S.; Paul, E.D.; Maeno, H.; Navon, I.; Zwang, R.; Gil, S.; Mayberg, H.S.; Dunlop, B.W.; Menke, A. MicroRNA 135 is essential for chronic stress resiliency, antidepressant efficacy, and intact serotonergic activity. Neuron 2014, 83, 344–360. [Google Scholar] [CrossRef]
- Rinaldi, A.; Vincenti, S.; De Vito, F.; Bozzoni, I.; Oliverio, A.; Presutti, C.; Fragapane, P.; Mele, A. Stress induces region specific alterations in microRNAs expression in mice. Behav. Brain Res. 2010, 208, 265–269. [Google Scholar] [CrossRef]
- Murphy, C.P.; Singewald, N. Role of microRNAs in anxiety and anxiety-related disorders. In Behavioral Neurogenomics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 185–219. [Google Scholar]
- Meerson, A.; Cacheaux, L.; Goosens, K.A.; Sapolsky, R.M.; Soreq, H.; Kaufer, D. Changes in brain MicroRNAs contribute to cholinergic stress reactions. J. Mol. Neurosci. 2010, 40, 47–55. [Google Scholar] [CrossRef]
- Schratt, G.M.; Tuebing, F.; Nigh, E.A.; Kane, C.G.; Sabatini, M.E.; Kiebler, M.; Greenberg, M.E. A brain-specific microRNA regulates dendritic spine development. Nature 2006, 439, 283–289. [Google Scholar] [CrossRef]
- Tang, X.; Gal, J.; Zhuang, X.; Wang, W.; Zhu, H.; Tang, G. A simple array platform for microRNA analysis and its application in mouse tissues. RNA 2007, 13, 1803–1822. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.D.; Naldini, L. Exploiting and antagonizing microRNA regulation for therapeutic and experimental applications. Nat. Rev. Genet. 2009, 10, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Krichevsky, A.M.; King, K.S.; Donahue, C.P.; Khrapko, K.; Kosik, K.S. A microRNA array reveals extensive regulation of microRNAs during brain development. Rna 2003, 9, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Chandan, K.; Gupta, M.; Sarwat, M. Role of host and pathogen-derived microRNAs in immune regulation during infectious and inflammatory diseases. Front. Immunol. 2020, 10, 3081. [Google Scholar] [CrossRef] [PubMed]
- Contreras, J.; Rao, D. MicroRNAs in inflammation and immune responses. Leukemia 2012, 26, 404–413. [Google Scholar] [CrossRef]
- Wang, H.; Liu, L.; Chen, X.; Zhou, C.; Rao, X.; Li, W.; Li, W.; Liu, Y.; Fang, L.; Zhang, H. MicroRNA–Messenger RNA Regulatory Network Mediates Disrupted TH17 Cell Differentiation in Depression. Front. Psychiatry 2022, 13, 824209. [Google Scholar] [CrossRef]
- Tan, L.P.; Wang, M.; Robertus, J.-L.; Schakel, R.N.; Gibcus, J.H.; Diepstra, A.; Harms, G.; Peh, S.-C.; Reijmers, R.M.; Pals, S.T. miRNA profiling of B-cell subsets: Specific miRNA profile for germinal center B cells with variation between centroblasts and centrocytes. Lab. Investig. 2009, 89, 708–716. [Google Scholar] [CrossRef]
- Issler, O.; Chen, A. Determining the role of microRNAs in psychiatric disorders. Nat. Rev. Neurosci. 2015, 16, 201–212. [Google Scholar] [CrossRef]
- Homorogan, C.; Nitusca, D.; Seclaman, E.; Enatescu, V.; Marian, C. Uncovering the Roles of MicroRNAs in Major Depressive Disorder: From Candidate Diagnostic Biomarkers to Treatment Response Indicators. Life 2021, 11, 1073. [Google Scholar] [CrossRef]
- Ferrúa, C.P.; Giorgi, R.; da Rosa, L.C.; do Amaral, C.C.; Ghisleni, G.C.; Pinheiro, R.T.; Nedel, F. MicroRNAs expressed in depression and their associated pathways: A systematic review and a bioinformatics analysis. J. Chem. Neuroanat. 2019, 100, 101650. [Google Scholar] [CrossRef]
- Dwivedi, Y. MicroRNAs in depression and suicide: Recent insights and future perspectives. J. Affect. Disord. 2018, 240, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.; Dwivedi, Y. MicroRNA mediators of early life stress vulnerability to depression and suicidal behavior. Mol. Psychiatry 2020, 25, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bi, T.; Yang, S. Exosomal microRNA-150-5p from bone marrow mesenchymal stromal cells mitigates cerebral ischemia/reperfusion injury via targeting toll-like receptor 5. Bioengineered 2022, 13, 3029–3042. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhu, Y.; Chen, W.; Tang, Y. Emerging role of microRNAs in major depressive disorder and its implication on diagnosis and therapeutic response. J. Affect. Disord. 2021, 286, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-Y.; Xia, Q.-H.; Xia, Q.-R.; Zhang, X.-L.; Liang, J. MicroRNA-based biomarkers in the diagnosis and monitoring of therapeutic response in patients with depression. Neuropsychiatr. Dis. Treat. 2019, 15, 3583–3597. [Google Scholar] [CrossRef]
- Kuang, W.-H.; Dong, Z.-Q.; Tian, L.-T.; Li, J. MicroRNA-451a, microRNA-34a-5p, and microRNA-221-3p as predictors of response to antidepressant treatment. Braz. J. Med. Biol. Res. 2018, 51, e7212. [Google Scholar] [CrossRef] [PubMed]
- Raisch, J.; Darfeuille-Michaud, A.; Nguyen, H.T.T. Role of microRNAs in the immune system, inflammation and cancer. World J. Gastroenterol. WJG 2013, 19, 2985. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef]
- Tili, E.; Michaille, J.-J.; Cimino, A.; Costinean, S.; Dumitru, C.D.; Adair, B.; Fabbri, M.; Alder, H.; Liu, C.G.; Calin, G.A. Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-α stimulation and their possible roles in regulating the response to endotoxin shock. J. Immunol. 2007, 179, 5082–5089. [Google Scholar] [CrossRef]
- Wagner, G.S.; McClintock, S.M.; Rosenquist, P.B.; McCall, W.V. Major depressive disorder with psychotic features may lead to misdiagnosis of dementia: A case report and review of the literature. J. Psychiatr. Pract. 2011, 17, 432. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-P.; Liu, X.-L.; Chen, J.-J.; Cheng, K.; Bai, S.-J.; Zheng, P.; Zhou, C.-J.; Wang, W.; Wang, H.-Y.; Zhong, L.-M. Circulating microRNA 134 sheds light on the diagnosis of major depressive disorder. Transl. Psychiatry 2020, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.A.; Jelenchick, L.A.; Egan, K.G.; Cox, E.; Young, H.; Gannon, K.E.; Becker, T. Feeling bad on Facebook: Depression disclosures by college students on a social networking site. Depress. Anxiety 2011, 28, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y. Emerging role of microRNAs in major depressive disorder: Diagnosis and therapeutic implications. Dialogues Clin. Neurosci. 2014, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Gaughwin, P.; Ciesla, M.; Yang, H.; Lim, B.; Brundin, P. Stage-specific modulation of cortical neuronal development by Mmu-miR-134. Cereb. Cortex 2011, 21, 1857–1869. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, W.-Y.; Mao, Y.-W.; Gräff, J.; Guan, J.-S.; Pan, L.; Mak, G.; Kim, D.; Su, S.C.; Tsai, L.-H. A novel pathway regulates memory and plasticity via SIRT1 and miR-134. Nature 2010, 466, 1105–1109. [Google Scholar] [CrossRef]
- Yang, D.; Li, Q.; Fang, L.; Cheng, K.; Zhang, R.; Zheng, P.; Zhan, Q.; Qi, Z.; Zhong, S.; Xie, P. Reduced neurogenesis and pre-synaptic dysfunction in the olfactory bulb of a rat model of depression. Neuroscience 2011, 192, 609–618. [Google Scholar] [CrossRef]
- Wei, X.; Yi, X.; Lv, H.; Sui, X.; Lu, P.; Li, L.; An, Y.; Yang, Y.; Yi, H.; Chen, G. MicroRNA-377-3p released by mesenchymal stem cell exosomes ameliorates lipopolysaccharide-induced acute lung injury by targeting RPTOR to induce autophagy. Cell Death Dis. 2020, 11, 657. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Q.; Song, R.; Kong, Y.; Zhang, Z. Non-coding RNAs in depression: Promising diagnostic and therapeutic biomarkers. EBioMedicine 2021, 71, 103569. [Google Scholar] [CrossRef]
- Uchida, S.; Hara, K.; Kobayashi, A.; Funato, H.; Hobara, T.; Otsuki, K.; Yamagata, H.; McEwen, B.S.; Watanabe, Y. Early life stress enhances behavioral vulnerability to stress through the activation of REST4-mediated gene transcription in the medial prefrontal cortex of rodents. J. Neurosci. 2010, 30, 15007–15018. [Google Scholar] [CrossRef]
- Zhou, R.; Yuan, P.; Wang, Y.; Hunsberger, J.G.; Elkahloun, A.; Wei, Y.; Damschroder-Williams, P.; Du, J.; Chen, G.; Manji, H.K. Evidence for selective microRNAs and their effectors as common long-term targets for the actions of mood stabilizers. Neuropsychopharmacology 2009, 34, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Muiños-Gimeno, M.; Espinosa-Parrilla, Y.; Guidi, M.; Kagerbauer, B.; Sipilä, T.; Maron, E.; Pettai, K.; Kananen, L.; Navinés, R.; Martín-Santos, R. Human microRNAs miR-22, miR-138-2, miR-148a, and miR-488 are associated with panic disorder and regulate several anxiety candidate genes and related pathways. Biol. Psychiatry 2011, 69, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Uchida, S.; Nishida, A.; Hara, K.; Kamemoto, T.; Suetsugi, M.; Fujimoto, M.; Watanuki, T.; Wakabayashi, Y.; Otsuki, K.; McEwen, B.S. Characterization of the vulnerability to repeated stress in Fischer 344 rats: Possible involvement of microRNA-mediated down-regulation of the glucocorticoid receptor. Eur. J. Neurosci. 2008, 27, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Wei, W.; Coelho, C.M.; Li, X.; Baker-Andresen, D.; Dudley, K.; Ratnu, V.S.; Boskovic, Z.; Kobor, M.S.; Sun, Y.E. The brain-specific microRNA miR-128b regulates the formation of fear-extinction memory. Nat. Neurosci. 2011, 14, 1115–1117. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.; Olsen, L.; Lindow, M.; Jakobsen, K.D.; Ullum, H.; Jonsson, E.; Andreassen, O.A.; Djurovic, S.; Melle, I.; Agartz, I. Brain expressed microRNAs implicated in schizophrenia etiology. PLoS ONE 2007, 2, e873. [Google Scholar] [CrossRef]
- Laux, G. Serotonin Reuptake Inhibitors: Citalopram, Escitalopram, Fluoxetine, Fluvoxamine, Paroxetine, and Sertraline. In NeuroPsychopharmacotherapy; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–13. [Google Scholar]
- Preskorn, S.H.; Ross, R.; Stanga, C. Selective serotonin reuptake inhibitors. In Antidepressants: Past, Present and Future; Springer: Berlin/Heidelberg, Germany, 2004; pp. 241–262. [Google Scholar]
- Chu, A.; Wadhwa, R. Selective serotonin reuptake inhibitors. In StatPearls [Internet]; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Eldar-Lissai, A.; Cohen, J.T.; Meltzer-Brody, S.; Gerbasi, M.E.; Chertavian, E.; Hodgkins, P.; Bond, J.C.; Johnson, S.J. Cost-effectiveness of brexanolone versus selective serotonin reuptake inhibitors for the treatment of postpartum depression in the United States. J. Manag. Care Spec. Pharm. 2020, 26, 627–638. [Google Scholar] [CrossRef]
- Zhao, J.; Tian, H.; Song, H.; Wang, X.; Luo, T.; Ai, L.; Fang, Y.; Zhao, J.; Chao-Ke-Tu, S.; Li, Z. Effect of electroacupuncture on reuptake of serotonin via mirna-16 expression in a rat model of depression. Evid. Based Complement. Altern. Med. 2019, 2019, 7124318. [Google Scholar] [CrossRef]
- Delgado, P.L.; Charney, D.S.; Price, L.H.; Aghajanian, G.K.; Landis, H.; Heninger, G.R. Serotonin function and the mechanism of antidepressant action: Reversal of antidepressant-induced remission by rapid depletion of plasma tryptophan. Arch. Gen. Psychiatry 1990, 47, 411–418. [Google Scholar] [CrossRef]
- Rosa, J.M.; Formolo, D.A.; Yu, J.; Lee, T.H.; Yau, S.-y. The Role of MicroRNA and Microbiota in Depression and Anxiety. Front. Behav. Neurosci. 2022, 16, 828258. [Google Scholar] [CrossRef]
- Shams, Y. Serotonin norepinephrine re-uptake inhibitor (SNRI)-, selective norepinephrine reuptake inhibitor (S-NRI)-, and exogenously administered norepinephrine-induced takotsubo syndrome: Analysis of published cases. Int. J. Cardiol. 2017, 231, 228–233. [Google Scholar]
- Dharmshaktu, P.; Tayal, V.; Kalra, B.S. Efficacy of antidepressants as analgesics: A review. J. Clin. Pharmacol. 2012, 52, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhen, J.; Karpowich, N.K.; Goetz, R.M.; Law, C.J.; Reith, M.E.; Wang, D.-N. LeuT-desipramine structure reveals how antidepressants block neurotransmitter reuptake. Science 2007, 317, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Scully Cbe, C. Drug effects on salivary glands: Dry mouth. Oral Dis. 2003, 9, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Ornoy, A.; Weinstein-Fudim, L.; Ergaz, Z. Antidepressants, antipsychotics, and mood stabilizers in pregnancy: What do we know and how should we treat pregnant women with depression. Birth Defects Res. 2017, 109, 933–956. [Google Scholar] [CrossRef]
- Burch, R. Antidepressants for preventive treatment of migraine. Curr. Treat. Options Neurol. 2019, 21, 18. [Google Scholar] [CrossRef]
- Punay, N.C.; Couch, J.R. Antidepressants in the treatment of migraine headache. Curr. Pain Headache Rep. 2003, 7, 51–54. [Google Scholar] [CrossRef]
- Foianini, A.; Joseph Wiegand, T.; Benowitz, N. What is the role of lidocaine or phenytoin in tricyclic antidepressant-induced cardiotoxicity? Clin. Toxicol. 2010, 48, 325–330. [Google Scholar] [CrossRef]
- Stoudemire, A.; Atkinson, P. Use of cyclic antidepressants in patients with cardiac conduction disturbances. Gen. Hosp. Psychiatry 1988, 10, 389–397. [Google Scholar] [CrossRef]
- Yekehtaz, H.; Farokhnia, M.; Akhondzadeh, S. Cardiovascular considerations in antidepressant therapy: An evidence-based review. J. Tehran Univ. Heart Cent. 2013, 8, 169. [Google Scholar]
- Stahl, S.M.; Felker, A. Monoamine oxidase inhibitors: A modern guide to an unrequited class of antidepressants. CNS Spectr. 2008, 13, 855–871. [Google Scholar] [CrossRef]
- Mialet-Perez, J.; Santin, Y.; Parini, A. Monoamine oxidase-A, serotonin and norepinephrine: Synergistic players in cardiac physiology and pathology. J. Neural Transm. 2018, 125, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Bortolato, M.; Chen, K.; Shih, J.C. Monoamine oxidase inactivation: From pathophysiology to therapeutics. Adv. Drug Deliv. Rev. 2008, 60, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Riederer, P.; Müller, T. Use of monoamine oxidase inhibitors in chronic neurodegeneration. Expert Opin. Drug Metab. Toxicol. 2017, 13, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Kalgutkar, A.S.; Dalvie, D.K.; Castagnoli, N.; Taylor, T.J. Interactions of nitrogen-containing xenobiotics with monoamine oxidase (MAO) isozymes A and B: SAR studies on MAO substrates and inhibitors. Chem. Res. Toxicol. 2001, 14, 1139–1162. [Google Scholar] [CrossRef]
- Dell’Osso, B.; Palazzo, M.C.; Oldani, L.; Altamura, A.C. The noradrenergic action in antidepressant treatments: Pharmacological and clinical aspects. CNS Neurosci. Ther. 2011, 17, 723–732. [Google Scholar] [CrossRef]
- Campos, A.I.; Mulcahy, A.; Thorp, J.G.; Wray, N.R.; Byrne, E.M.; Lind, P.A.; Medland, S.E.; Martin, N.G.; Hickie, I.B.; Rentería, M.E. Understanding genetic risk factors for common side effects of antidepressant medications. Commun. Med. 2021, 1, 1–10. [Google Scholar] [CrossRef]
- Buckley, N.A.; Faunce, T.A. ‘Atypical’antidepressants in overdose. Drug Saf. 2003, 26, 539–551. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Sharma, M.S.; Brunoni, A.R.; Vieta, E.; Fava, G.A. The safety, tolerability and risks associated with the use of newer generation antidepressant drugs: A critical review of the literature. Psychother. Psychosom. 2016, 85, 270–288. [Google Scholar] [CrossRef]
- Shelton, R.C.; Papakostas, G.I. Augmentation of antidepressants with atypical antipsychotics for treatment-resistant major depressive disorder. Acta Psychiatr. Scand. 2008, 117, 253–259. [Google Scholar] [CrossRef]
- Arnone, D.; Horder, J.; Cowen, P.; Harmer, C. Early effects of mirtazapine on emotional processing. Psychopharmacology 2009, 203, 685–691. [Google Scholar] [CrossRef]
- Davis, R.; Wilde, M.I. Mirtazapine. Cns Drugs 1996, 5, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Sahli, Z.T.; Banerjee, P.; Tarazi, F.I. The preclinical and clinical effects of vilazodone for the treatment of major depressive disorder. Expert Opin. Drug Discov. 2016, 11, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-x.; Jin, Z.-l.; Zhang, L.-m.; Xue, R.; Xu, X.-d.; Zhao, N.; Qiu, Z.-k.; Wang, X.-w.; Zhang, Y.-z.; Yang, R.-f. Antidepressant-like activity of YL-0919: A novel combined selective serotonin reuptake inhibitor and 5-HT1A receptor agonist. PLoS ONE 2013, 8, e83271. [Google Scholar] [CrossRef] [PubMed]
- Rickels, K.; Downing, R.; Schweizer, E.; Hassman, H. Antidepressants for the treatment of generalized anxiety disorder: A placebo-controlled comparison of imipramine, trazodone, and diazepam. Arch. Gen. Psychiatry 1993, 50, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Fagiolini, A.; Comandini, A.; Dell’Osso, M.C.; Kasper, S. Rediscovering trazodone for the treatment of major depressive disorder. CNS Drugs 2012, 26, 1033–1049. [Google Scholar] [CrossRef]
- Hedges, D.W.; Reimherr, F.W.; Strong, R.E.; Halls, C.H.; Rust, C. An open trial of nefazodone in adult patients with generalized anxiety disorder. Psychopharmacol. Bull. 1996, 32, 671–676. [Google Scholar]
- Sheehan, D.V. Venlafaxine extended release (XR) in the treatment of generalized anxiety disorder. Prim. Care Companion CNS Disord. 1999, 2, 22150. [Google Scholar]
- Allgulander, C.; Hackett, D.; Salinas, E. Venlafaxine extended release (ER) in the treatment of generalised anxiety disorder: Twenty-four-week placebo-controlled dose-ranging study. Br. J. Psychiatry 2001, 179, 15–22. [Google Scholar] [CrossRef]
- Shao, L.; Sun, X.; Xu, L.; Young, L.T.; Wang, J.-F. Mood stabilizing drug lithium increases expression of endoplasmic reticulum stress proteins in primary cultured rat cerebral cortical cells. Life Sci. 2006, 78, 1317–1323. [Google Scholar] [CrossRef]
- Licht, R.W. Lithium: Still a major option in the management of bipolar disorder. CNS Neurosci. Ther. 2012, 18, 219–226. [Google Scholar] [CrossRef]
- Bauer, M.; Bschor, T.; Kunz, D.; Berghöfer, A.; Ströhle, A.; Müller-Oerlinghausen, B. Double-blind, placebo-controlled trial of the use of lithium to augment antidepressant medication in continuation treatment of unipolar major depression. Am. J. Psychiatry 2000, 157, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Barroilhet, S.A.; Ghaemi, S.N. When and how to use lithium. Acta Psychiatr. Scand. 2020, 142, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Van Ameringen, M.; Mancini, C.; Pipe, B.; Bennett, M. Optimizing treatment in social phobia: A review of treatment resistance. CNS Spectr. 2004, 9, 753–762. [Google Scholar] [PubMed]
- Kandaswamy, R.; McQuillin, A.; Curtis, D.; Gurling, H. Allelic association, DNA resequencing and copy number variation at the metabotropic glutamate receptor GRM7 gene locus in bipolar disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2014, 165, 365–372. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Anderson, I.M.; Nutt, D.J.; Allgulander, C.; Bandelow, B.; den Boer, J.A.; Christmas, D.M.; Davies, S.; Fineberg, N.; Lidbetter, N. Evidence-based pharmacological treatment of anxiety disorders, post-traumatic stress disorder and obsessive-compulsive disorder: A revision of the 2005 guidelines from the British Association for Psychopharmacology. J. Psychopharmacol. 2014, 28, 403–439. [Google Scholar] [CrossRef]
- Schmidt, U.; Herrmann, L.; Hagl, K.; Novak, B.; Huber, C.; Holsboer, F.; Wotjak, C.T.; Buell, D.R. Therapeutic action of fluoxetine is associated with a reduction in prefrontal cortical miR-1971 expression levels in a mouse model of posttraumatic stress disorder. Front. Psychiatry 2013, 4, 66. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Wang, L.; Bai, M.; Zhang, X.; Zhu, X. Dopamine receptor D2 and associated microRNAs are involved in stress susceptibility and resistance to escitalopram treatment. Int. J. Neuropsychopharmacol. 2015, 18, pyv025. [Google Scholar] [CrossRef]
- Cruceanu, C.; Lopez, J.P.; Tsai, W.-T.; Turecki, G. Dysregulation of the glutamatergic receptors after antidepressant treatment in human neural progenitor cells. Mol. Psychiatry 2017, 22, 1228–1229. [Google Scholar] [CrossRef][Green Version]
- Lopez, J.P.; Lim, R.; Cruceanu, C.; Crapper, L.; Fasano, C.; Labonte, B.; Maussion, G.; Yang, J.P.; Yerko, V.; Vigneault, E. miR-1202 is a primate-specific and brain-enriched microRNA involved in major depression and antidepressant treatment. Nat. Med. 2014, 20, 764–768. [Google Scholar]
- Dagyte, G.; Luiten, P.G.; De Jager, T.; Gabriel, C.; Mocaër, E.; Den Boer, J.A.; Van der Zee, E.A. Chronic stress and antidepressant agomelatine induce region-specific changes in synapsin I expression in the rat brain. J. Neurosci. Res. 2011, 89, 1646–1657. [Google Scholar]
- Bai, G.; Dunbar, M.; Miller, L.D.; Roy, B.; Soller, I.J.; Shelton, R.C.; Dwivedi, Y. microRNAs As Novel Players in Depression Pathogenesis and in the Mechanisms of Action of Fluoxetine and Other Antidepressants. In Pharmacology, Mechanisms of Action and Potential Side Effects; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2015; p. 147. [Google Scholar]
- Zhu, J.; Chen, Z.; Meng, Z.; Ju, M.; Zhang, M.; Wu, G.; Guo, H.; Tian, Z. Electroacupuncture alleviates surgical trauma-induced hypothalamus pituitary adrenal axis hyperactivity via microRNA-142. Front. Mol. Neurosci. 2017, 10, 308. [Google Scholar] [CrossRef] [PubMed]
- Popova, D. Neurophysiological Mechanisms of Plasticity Induced in Adult Brain; Helsingin Yliopisto: Helsinki, Finland, 2015. [Google Scholar]
- S Wohleb, E.; Gerhard, D.; Thomas, A.; S Duman, R. Molecular and cellular mechanisms of rapid-acting antidepressants ketamine and scopolamine. Curr. Neuropharmacol. 2017, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, S.; Bang, E.; Yue, W.; Hare, B.D.; Lepack, A.E.; Girgenti, M.J.; Duman, R.S. Activity-dependent brain-derived neurotrophic factor release is required for the rapid antidepressant actions of scopolamine. Biol. Psychiatry 2018, 83, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, S.; Hare, B.D.; Duman, R.S. Prefrontal cortex GABAergic deficits and circuit dysfunction in the pathophysiology and treatment of chronic stress and depression. Curr. Opin. Behav. Sci. 2017, 14, 1–8. [Google Scholar] [CrossRef]
- Duman, R.S. Pathophysiology of depression and innovative treatments: Remodeling glutamatergic synaptic connections. Dialogues Clin. Neurosci. 2014, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef]
- Calabro, P.; Chang, D.W.; Willerson, J.T.; Yeh, E.T. Release of C-reactive protein in response to inflammatory cytokines by human adipocytes: Linking obesity to vascular inflammation. J. Am. Coll. Cardiol. 2005, 46, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.A.; Rickels, K. Pharmacological treatment for generalized anxiety disorder in adults: An update. Expert Opin. Pharmacother. 2015, 16, 1669–1681. [Google Scholar] [CrossRef]
- Walter, H.J.; Bukstein, O.G.; Abright, A.R.; Keable, H.; Ramtekkar, U.; Ripperger-Suhler, J.; Rockhill, C. Clinical practice guideline for the assessment and treatment of children and adolescents with anxiety disorders. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 1107–1124. [Google Scholar] [CrossRef]
- Ballou, S.; Keefer, L. Psychological interventions for irritable bowel syndrome and inflammatory bowel diseases. Clin. Transl. Gastroenterol. 2017, 8, e214. [Google Scholar] [CrossRef]
- Craske, M.G.; Barlow, D.H. Mastery of Your Anxiety and Worry; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Hirschtritt, M.E.; Bloch, M.H.; Mathews, C.A. Obsessive-compulsive disorder: Advances in diagnosis and treatment. Jama 2017, 317, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.A.; Eisen, J.L. The epidemiology and differential diagnosis of obsessive compulsive disorder. J. Clin. Psychiatry 1994, 55, 5–10. [Google Scholar] [PubMed]
- De Silva, P.; Rachman, S. Obsessive-Compulsive Disorder: The Facts; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Saxena, S.; Brody, A.L.; Schwartz, J.M.; Baxter, L.R. Neuroimaging and frontal-subcortical circuitry in obsessive-compulsive disorder. Br. J. Psychiatry 1998, 173, 26–37. [Google Scholar] [CrossRef]
- Shephard, E.; Stern, E.R.; van den Heuvel, O.A.; Costa, D.L.; Batistuzzo, M.C.; Godoy, P.B.; Lopes, A.C.; Brunoni, A.R.; Hoexter, M.Q.; Shavitt, R.G. Toward a neurocircuit-based taxonomy to guide treatment of obsessive–compulsive disorder. Mol. Psychiatry 2021, 26, 4583–4604. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.B.; Liebowitz, M.R.; Lydiard, R.B.; Pitts, C.D.; Bushnell, W.; Gergel, I. Paroxetine treatment of generalized social phobia (social anxiety disorder): A randomized controlled trial. JAMA 1998, 280, 708–713. [Google Scholar] [CrossRef]
- National Collaborating Centre for Mental Health. Common mental health disorders. In Common Mental Health Disorders: Identification and Pathways to Care; British Psychological Society: Leicester, UK, 2011. [Google Scholar]
- Merikangas, K.; Lieb, R.; Wittchen, H.U.; Avenevoli, S. Family and high-risk studies of social anxiety disorder. Acta Psychiatr. Scand. 2003, 108, 28–37. [Google Scholar] [CrossRef]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress. Anxiety 2000, 12, 2–19. [Google Scholar] [CrossRef]
- Beesdo, K.; Knappe, S.; Pine, D.S. Anxiety and anxiety disorders in children and adolescents: Developmental issues and implications for DSM-V. Psychiatr. Clin. 2009, 32, 483–524. [Google Scholar] [CrossRef]
- Rodebaugh, T.L.; Holaway, R.M.; Heimberg, R.G. The treatment of social anxiety disorder. Clin. Psychol. Rev. 2004, 24, 883–908. [Google Scholar]
- Pelissolo, A.; Abou Kassm, S.; Delhay, L. Therapeutic strategies for social anxiety disorder: Where are we now? Expert Rev. Neurother. 2019, 19, 1179–1189. [Google Scholar]
- Wilhelm, F.H.; Trabert, W.; Roth, W.T. Physiologic instability in panic disorder and generalized anxiety disorder. Biol. Psychiatry 2001, 49, 596–605. [Google Scholar] [CrossRef]
- Brown, L.A.; LeBeau, R.; Liao, B.; Niles, A.N.; Glenn, D.; Craske, M.G. A comparison of the nature and correlates of panic attacks in the context of Panic Disorder and Social Anxiety Disorder. Psychiatry Res. 2016, 235, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Katon, W.J. Panic disorder. N. Engl. J. Med. 2006, 354, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Noyes, R., Jr. Suicide and panic disorder: A review. J. Affect. Disord. 1991, 22, 1–11. [Google Scholar] [CrossRef]
- Youdim, M.B.; Edmondson, D.; Tipton, K.F. The therapeutic potential of monoamine oxidase inhibitors. Nat. Rev. Neurosci. 2006, 7, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Kayabaşı, Y.; Güneş, B.; Erbaş, O. Serotonin Receptors and Depression. J. Exp. Basic Med. Sci. 2021, 2, 240–246. [Google Scholar]
- Lookingland, K.; Shannon, N.; Moore, K. Effects of tryptophan administration on the synthesis, storage and metabolism of serotonin in the hypothalamus of normal and raphe-stimulated rats. In Amino Acid Availability and Brain Function in Health and Disease; Springer: Berlin/Heidelberg, Germany, 1988; pp. 159–166. [Google Scholar]
- Feng, H.; Gao, K.; Chen, D.; Robison, A.J.; Ellsworth, E.; Wei, G.-W. Machine learning analysis of cocaine addiction informed by DAT, SERT, and NET-based interactome networks. arXiv 2022, arXiv:2201.00114. [Google Scholar] [CrossRef]
- Chang, J.P.-C.; Zamparelli, A.; Nettis, M.; Pariante, C. Antidepressant Drugs: Mechanisms of Action and Side Effects. In Encyclopedia of Behavioral Neuroscience, 2nd ed.; Sala, S.D., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2022; pp. 613–626. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).