
Decoding Strategies to Evade Immunoregulators Galectin-1, -3, and -9 and Their Ligands as Novel Therapeutics in Cancer Immunotherapy

Abstract
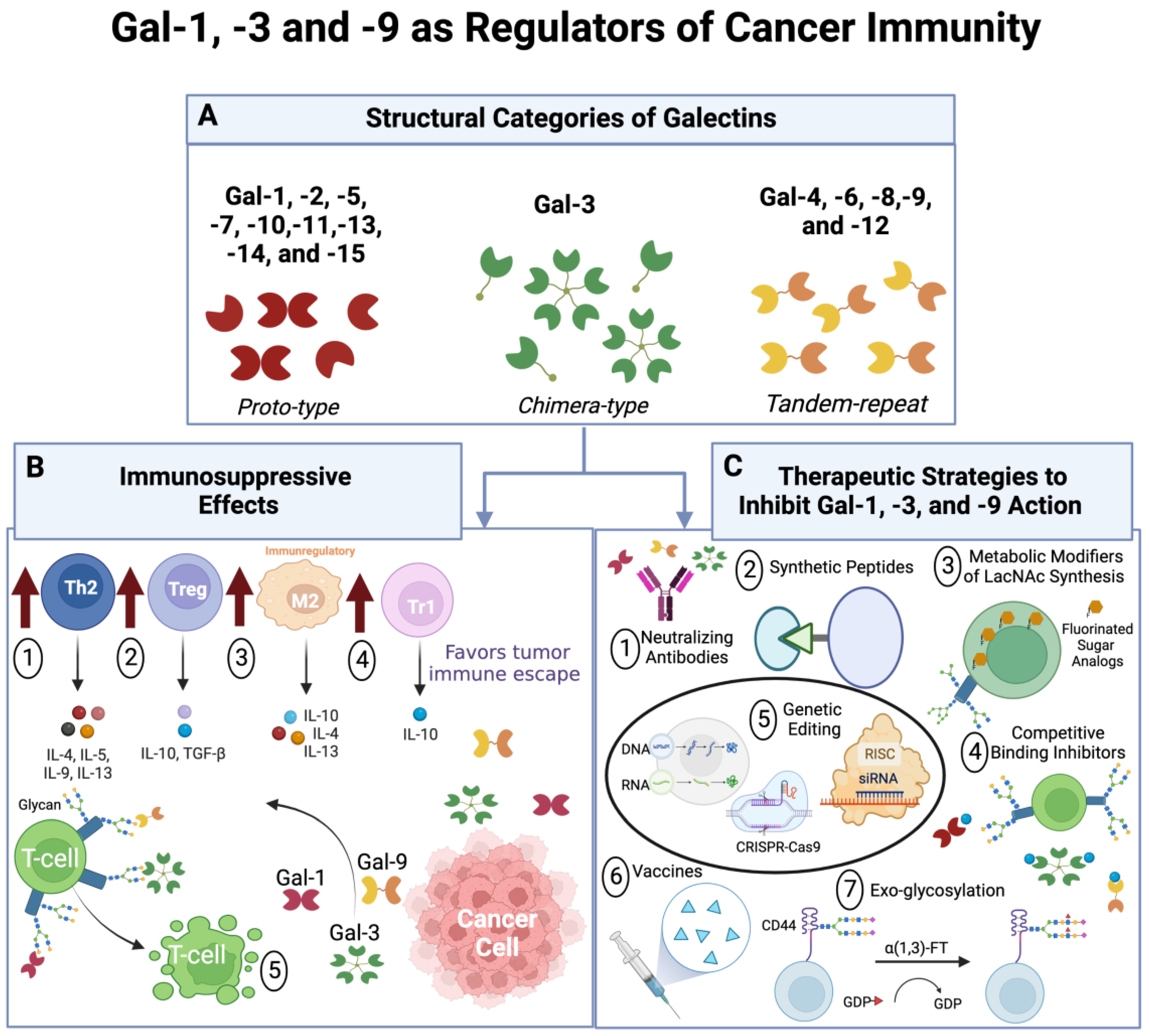
:1. Introduction
2. Molecular Structure of Gal-1, -3, and -9 and their Involvement in Cancer
2.1. Gal-1
2.2. Gal-3
2.3. Gal-9
3. Effects of Gal-1, -3, and -9 on Cancer Immunosurveillance
3.1. Regulation of T Cell Viability
3.2. Modulation of T Cell Differentiation and Effector Function
3.3. Regulation of Resistance against Cancer Immunotherapies
4. Strategies to Block Gal-1-, -3, and -9–Ligand Binding as Novel Anti-Cancer Therapies
4.1. Neutralizing Antibodies
4.2. Metabolic Modifiers of LacNAc Synthesis and Competitive Inhibitors of Galectin Binding
4.3. Vaccines
4.4. Gene Editing
4.5. Exo-Glycosylation
5. Cell-Based Anti-Cancer Therapies That Can Evade Gal-1, -3, and -9 Immunoregulation
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Toscano, M.A.; Allo, V.C.M.; Cutine, A.M.; Rabinovich, G.A.; Mariño, K.V. Untangling galectin-driven regulatory circuits in autoimmune inflammation. Trends Mol. Med. 2018, 24, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.N. Galectinomics: Finding themes in complexity. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2002, 1572, 209–231. [Google Scholar] [CrossRef]
- Kaltner, H.; Toegel, S.; Caballero, G.G.; Manning, J.C.; Ledeen, R.W.; Gabius, H.-J. Galectins: Their network and roles in immunity/tumor growth control. Histochem. Cell Biol. 2017, 147, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H.; Cooper, D.; Gitt, M.A.; Leffler, H. Galectins. Structure and function of a large family of animal lectins. J. Biol. Chem. 1994, 269, 20807–20810. [Google Scholar] [CrossRef]
- Cummings, R.D.; Liu, F.-T. Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; Chapter 33. Available online: https://www.ncbi.nlm.nih.gov/books/NBK1944/ (accessed on 11 November 2022).
- Liu, F.-T.; Patterson, R.J.; Wang, J.L. Intracellular functions of galectins. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2002, 1572, 263–273. [Google Scholar] [CrossRef]
- Elola, M.; Wolfenstein-Todel, C.; Troncoso, M.; Vasta, G.; Rabinovich, G. Galectins: Matricellular glycan-binding proteins linking cell adhesion, migration, and survival. Cell. Mol. Life Sci. 2007, 64, 1679–1700. [Google Scholar] [CrossRef]
- Thijssen, V.L.; Heusschen, R.; Caers, J.; Griffioen, A.W. Galectin expression in cancer diagnosis and prognosis: A systematic review. Biochim. Biophys. Acta 2015, 1855, 235–247. [Google Scholar] [CrossRef]
- Mendez-Huergo, S.P.; Blidner, A.G.; Rabinovich, G.A. Galectins: Emerging regulatory checkpoints linking tumor immunity and angiogenesis. Curr. Opin. Immunol. 2017, 45, 8–15. [Google Scholar] [CrossRef]
- Chou, F.C.; Chen, H.Y.; Kuo, C.C.; Sytwu, H.K. Role of Galectins in Tumors and in Clinical Immunotherapy. Int. J. Mol. Sci. 2018, 19, 430. [Google Scholar] [CrossRef] [Green Version]
- Chun, B.; Pucilowska, J.; Chang, S.; Kim, I.; Nikitin, B.; Koguchi, Y.; Redmond, W.L.; Bernard, B.; Rajamanickam, V.; Polaske, N.; et al. Changes in T-cell subsets and clonal repertoire during chemoimmunotherapy with pembrolizumab and paclitaxel or capecitabine for metastatic triple-negative breast cancer. J. Immunother. Cancer 2022, 10, e004033. [Google Scholar] [CrossRef]
- Yang, Y. Cancer immunotherapy: Harnessing the immune system to battle cancer. J. Clin. Investig. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahim, A.H.; Alalawi, Z.; Mirandola, L.; Rakhshanda, R.; Dahlbeck, S.; Nguyen, D.; Jenkins, M.; Grizzi, F.; Cobos, E.; Figueroa, J.A.; et al. Galectins in cancer: Carcinogenesis, diagnosis and therapy. Ann. Transl. Med. 2014, 2, 88. [Google Scholar] [CrossRef] [PubMed]
- Cedeno-Laurent, F.; Dimitroff, C.J. Galectin-1 research in T cell immunity: Past, present and future. Clin. Immunol. 2012, 142, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astorgues-Xerri, L.; Riveiro, M.E.; Tijeras-Raballand, A.; Serova, M.; Neuzillet, C.; Albert, S.; Raymond, E.; Faivre, S. Unraveling galectin-1 as a novel therapeutic target for cancer. Cancer Treat. Rev. 2014, 40, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Teichberg, V.I.; Silman, I.; Beitsch, D.D.; Resheff, G. A beta-D-galactoside binding protein from electric organ tissue of Electrophorus electricus. Proc. Natl. Acad. Sci. USA 1975, 72, 1383–1387. [Google Scholar] [CrossRef] [Green Version]
- Iida, A.; Ozaki, K.; Tanaka, T.; Nakamura, Y. Fine-scale SNP map of an 11-kb genomic region at 22q13. 1 containing the galectin-1 gene. J. Hum. Genet. 2005, 50, 42–45. [Google Scholar] [CrossRef] [Green Version]
- Kuroi, K.; Kamijo, M.; Ueki, M.; Niwa, Y.; Hiramatsu, H.; Nakabayashi, T. Time-resolved FTIR study on the structural switching of human galectin-1 by light-induced disulfide bond formation. Phys. Chem. Chem. Phys. 2020, 22, 1137–1144. [Google Scholar] [CrossRef]
- Inagaki, Y.; Sohma, Y.; Horie, H.; Nozawa, R.; Kadoya, T. Oxidized galectin-1 promotes axonal regeneration in peripheral nerves but does not possess lectin properties. Eur. J. Biochem. 2000, 267, 2955–2964. [Google Scholar] [CrossRef]
- Cummings, R.D.; Liu, F.T.; Rabinovich, G.A.; Stowell, S.R.; Vasta, G.R. Galectins. In Essentials of Glycobiology, 4th ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Mohnen, D., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022; pp. 491–504. [Google Scholar] [CrossRef]
- Giannini, S.; Lee-Sundlov, M.M.; Rivadeneyra, L.; Di Buduo, C.A.; Burns, R.; Lau, J.T.; Falet, H.; Balduini, A.; Hoffmeister, K.M. β4GALT1 controls β1 integrin function to govern thrombopoiesis and hematopoietic stem cell homeostasis. Nat. Commun. 2020, 11, 356. [Google Scholar] [CrossRef]
- Kimber, M.S. How to extend your (polylactosamine) antennae. J. Biol. Chem. 2021, 296, 100212. [Google Scholar] [CrossRef]
- Hao, Y.; Créquer-Grandhomme, A.; Javier, N.; Singh, A.; Chen, H.; Manzanillo, P.; Lo, M.-C.; Huang, X. Structures and mechanism of human glycosyltransferase β1,3-N-acetylglucosaminyltransferase 2 (B3GNT2), an important player in immune homeostasis. J. Biol. Chem. 2021, 296, 100042. [Google Scholar] [CrossRef] [PubMed]
- Sundblad, V.; Morosi, L.G.; Geffner, J.R.; Rabinovich, G.A. Galectin-1: A Jack-of-All-Trades in the Resolution of Acute and Chronic Inflammation. J. Immunol. 2017, 199, 3721–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovich, G.A.; Toscano, M.A. Turning’sweet’on immunity: Galectin–glycan interactions in immune tolerance and inflammation. Nat. Rev. Immunol. 2009, 9, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Galvan, M.; He, J.; Baum, L.G. The ST6Gal I Sialyltransferase selectively modifiesN-Glycans on CD45 to negatively regulate galectin-1-induced CD45 clustering, phosphatase modulation, and T cell death. J. Biol. Chem. 2003, 278, 7469–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, X.; Liu, Q.; Wu, J.; Wang, Y.; Dai, J.; Chen, D.; Zhou, Y.; Lian, Y. Galectin-1 promotes vasculogenic mimicry in gastric cancer by upregulating EMT signaling. J. Cancer 2019, 10, 6286. [Google Scholar] [CrossRef] [PubMed]
- Tinari, N.; Kuwabara, I.; Huflejt, M.E.; Shen, P.F.; Iacobelli, S.; Liu, F.T. Glycoprotein 90K/MAC-2BP interacts with galectin-1 and mediates galectin-1–induced cell aggregation. Int. J. Cancer 2001, 91, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hittelet, A.; Legendre, H.; Nagy, N.; Bronckart, Y.; Pector, J.C.; Salmon, I.; Yeaton, P.; Gabius, H.J.; Kiss, R.; Camby, I. Upregulation of galectins-1 and-3 in human colon cancer and their role in regulating cell migration. Int. J. Cancer 2003, 103, 370–379. [Google Scholar] [CrossRef]
- Ito, K.; Ralph, S.J. Inhibiting galectin-1 reduces murine lung metastasis with increased CD4+ and CD8+ T cells and reduced cancer cell adherence. Clin. Exp. Metastasis 2012, 29, 561–572. [Google Scholar] [CrossRef]
- Yazawa, E.M.; Geddes-Sweeney, J.E.; Cedeno-Laurent, F.; Walley, K.C.; Barthel, S.R.; Opperman, M.J.; Liang, J.; Lin, J.Y.; Schatton, T.; Laga, A.C. Melanoma cell galectin-1 ligands functionally correlate with malignant potential. J. Investig. Dermatol. 2015, 135, 1849–1862. [Google Scholar] [CrossRef]
- You, X.; Wu, J.; Wang, Y.; Liu, Q.; Cheng, Z.; Zhao, X.; Liu, G.; Huang, C.; Dai, J.; Zhou, Y.; et al. Galectin-1 promotes vasculogenic mimicry in gastric adenocarcinoma via the Hedgehog/GLI signaling pathway. Aging 2020, 12, 21837–21853. [Google Scholar] [CrossRef]
- Le Mercier, M.; Fortin, S.; Mathieu, V.; Roland, I.; Spiegl-Kreinecker, S.; Haibe-Kains, B.; Bontempi, G.; Decaestecker, C.; Berger, W.; Lefranc, F. Galectin 1 proangiogenic and promigratory effects in the Hs683 oligodendroglioma model are partly mediated through the control of BEX2 expression. Neoplasia 2009, 11, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Dalotto-Moreno, T.; Croci, D.O.; Cerliani, J.P.; Martinez-Allo, V.C.; Dergan-Dylon, S.; Méndez-Huergo, S.P.; Stupirski, J.C.; Mazal, D.; Osinaga, E.; Toscano, M.A. Targeting Galectin-1 Overcomes Breast Cancer-Associated Immunosuppression and Prevents Metastatic DiseaseTargeting Gal1 in Breast Cancer. Cancer Res. 2013, 73, 1107–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.-Y.; Jung, S.; Ryu, H.-H.; Jeong, Y.-I.; Jin, Y.-H.; Jin, S.-G.; Kim, I.-Y.; Kang, S.-S.; Kim, H.-S. Role of galectin-1 in migration and invasion of human glioblastoma multiforme cell lines. J. Neurosurg. 2008, 109, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Li, K.; Shen, Y.; Gao, P.; Dong, Z.; Cai, J.; Zhang, C.; Huang, X.; Tian, M.; Hu, Z. Galectin-1 induces hepatocellular carcinoma EMT and sorafenib resistance by activating FAK/PI3K/AKT signaling. Cell Death Dis. 2016, 7, e2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.-L.; Luo, H.-L.; Huang, C.-C.; Liu, T.-T.; Huang, E.-Y.; Sung, M.-T.; Lin, J.-J.; Chiang, P.-H.; Chen, Y.-T.; Kang, C.-H.; et al. Galectin-1 Overexpression Activates the FAK/PI3K/AKT/mTOR Pathway and Is Correlated with Upper Urinary Urothelial Carcinoma Progression and Survival. Cells 2020, 9, 806. [Google Scholar] [CrossRef] [Green Version]
- Paz, A.; Haklai, R.; Elad-Sfadia, G.; Ballan, E.; Kloog, Y. Galectin-1 binds oncogenic H-Ras to mediate Ras membrane anchorage and cell transformation. Oncogene 2001, 20, 7486–7493. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Zheng, Y.; Zhang, H.; Liu, Y.; Sun, H.; Zhang, P. Galectin-1 induces metastasis and epithelial-mesenchymal transition (EMT) in human ovarian cancer cells via activation of the MAPK JNK/p38 signalling pathway. Am. J. Transl. Res. 2019, 11, 3862. [Google Scholar]
- Ose, R.; Oharaa, O.; Nagase, T. Suppl 1: Galectin-1 and Galectin-3 Mediate Protocadherin-24-Dependent Membrane Localization of β-catenin in Colon Cancer Cell Line HCT116. Curr. Chem. Genom. 2012, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.K.; Springer, T.A. Mac-2, a novel 32,000 Mr mouse macrophage subpopulation-specific antigen defined by monoclonal antibodies. J. Immunol. 1982, 128, 1221–1228. [Google Scholar]
- Dumic, J.; Dabelic, S.; Flögel, M. Galectin-3: An open-ended story. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2006, 1760, 616–635. [Google Scholar] [CrossRef]
- Zhong, X.; Qian, X.; Chen, G.; Song, X. The role of galectin-3 in heart failure and cardiovascular disease. Clin. Exp. Pharmacol. Physiol. 2019, 46, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, J.; Kuno, A.; Hirabayashi, J.; Sato, S. Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J. Biol. Chem. 2007, 282, 1374–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bumba, L.; Laaf, D.; Spiwok, V.; Elling, L.; Křen, V.; Bojarová, P. Poly-N-Acetyllactosamine Neo-Glycoproteins as Nanomolar Ligands of Human Galectin-3: Binding Kinetics and Modeling. Int. J. Mol. Sci. 2018, 19, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haji-Ghassemi, O.; Gilbert, M.; Spence, J.; Schur, M.J.; Parker, M.J.; Jenkins, M.L.; Burke, J.E.; van Faassen, H.; Young, N.M.; Evans, S.V. Molecular basis for recognition of the cancer glycobiomarker, LacdiNAc (GalNAc [β1→ 4] GlcNAc), by Wisteria floribunda Agglutinin. J. Biol. Chem. 2016, 291, 24085–24095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, S.N.; Junqueira, M.S.; Francisco, G.; Vilanova, M.; Magalhães, A.; Baruffi, M.D.; Chammas, R.; Harris, A.L.; Reis, C.A.; Bernardes, E.S. O-glycan sialylation alters galectin-3 subcellular localization and decreases chemotherapy sensitivity in gastric cancer. Oncotarget 2016, 7, 83570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.I.; Stegmayr, J.; Grant, O.C.; Yang, Z.; Nilsson, U.J.; Boos, I.; Carlsson, M.C.; Woods, R.J.; Unverzagt, C.; Leffler, H.; et al. Galectin binding to cells and glycoproteins with genetically modified glycosylation reveals galectin-glycan specificities in a natural context. J. Biol. Chem. 2018, 293, 20249–20262. [Google Scholar] [CrossRef] [Green Version]
- Nangia-Makker, P.; Hogan, V.; Raz, A. Galectin-3 and cancer stemness. Glycobiology 2018, 28, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Girotti, M.R.; Salatino, M.; Dalotto-Moreno, T.; Rabinovich, G.A. Sweetening the hallmarks of cancer: Galectins as multifunctional mediators of tumor progression. J. Exp. Med. 2020, 217, e20182041. [Google Scholar] [CrossRef]
- Wu, K.-L.; Huang, E.-Y.; Jhu, E.-W.; Huang, Y.-H.; Su, W.-H.; Chuang, P.-C.; Yang, K.D. Overexpression of galectin-3 enhances migration of colon cancer cells related to activation of the K-Ras–Raf–Erk1/2 pathway. J. Gastroenterol. 2013, 48, 350–359. [Google Scholar] [CrossRef]
- Wang, L.P.; Chen, S.W.; Zhuang, S.M.; Li, H.; Song, M. Galectin-3 accelerates the progression of oral tongue squamous cell carcinoma via a Wnt/beta-catenin-dependent pathway. Pathol. Oncol. Res. 2013, 19, 461–474. [Google Scholar] [CrossRef]
- Kim, S.J.; Kang, H.G.; Kim, K.; Kim, H.; Zetterberg, F.; Park, Y.S.; Cho, H.S.; Hewitt, S.M.; Chung, J.Y.; Nilsson, U.J.; et al. Crosstalk between WNT and STAT3 is mediated by galectin-3 in tumor progression. Gastric Cancer 2021, 24, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Fortuna-Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S. Extracellular galectin-3 in tumor progression and metastasis. Front. Oncol. 2014, 4, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhad, M.; Rolig, A.S.; Redmond, W.L. The role of Galectin-3 in modulating tumor growth and immunosuppression within the tumor microenvironment. Oncoimmunology 2018, 7, e1434467. [Google Scholar] [CrossRef] [Green Version]
- Markowska, A.I.; Liu, F.-T.; Panjwani, N. Galectin-3 is an important mediator of VEGF-and bFGF-mediated angiogenic response. J. Exp. Med. 2010, 207, 1981–1993. [Google Scholar] [CrossRef] [Green Version]
- John, S.; Mishra, R. Galectin-9: From cell biology to complex disease dynamics. J. Biosci. 2016, 41, 507–534. [Google Scholar] [CrossRef] [PubMed]
- Wada, J.; Kanwar, Y.S. Identification and characterization of galectin-9, a novel β-galactoside-binding mammalian lectin. J. Biol. Chem. 1997, 272, 6078–6086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Türeci, Ö.; Schmitt, H.; Fadle, N.; Pfreundschuh, M.; Sahin, U. Molecular definition of a novel human galectin which is immunogenic in patients with Hodgkin’s disease. J. Biol. Chem. 1997, 272, 6416–6422. [Google Scholar] [CrossRef] [Green Version]
- Heusschen, R.; Griffioen, A.W.; Thijssen, V.L. Galectin-9 in tumor biology: A jack of multiple trades. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2013, 1836, 177–185. [Google Scholar] [CrossRef]
- Moure, M.J.; Gimeno, A.; Delgado, S.; Diercks, T.; Boons, G.J.; Jiménez-Barbero, J.; Arda, A. Selective 13C-Labels on Repeating Glycan Oligomers to Reveal Protein Binding Epitopes through NMR: Polylactosamine Binding to Galectins. Angew. Chem. Int. Ed. 2021, 60, 18777–18782. [Google Scholar] [CrossRef]
- Giovannone, N.; Liang, J.; Antonopoulos, A.; Geddes Sweeney, J.; King, S.L.; Pochebit, S.M.; Bhattacharyya, N.; Lee, G.S.; Dell, A.; Widlund, H.R.; et al. Galectin-9 suppresses B cell receptor signaling and is regulated by I-branching of N-glycans. Nat. Commun. 2018, 9, 3287. [Google Scholar] [CrossRef]
- Moar, P.; Tandon, R. Galectin-9 as a biomarker of disease severity. Cell. Immunol. 2021, 361, 104287. [Google Scholar] [CrossRef] [PubMed]
- Earl, L.A.; Bi, S.; Baum, L.G. Galectin multimerization and lattice formation are regulated by linker region structure. Glycobiology 2011, 21, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansour, A.A.; Raucci, F.; Sevim, M.; Saviano, A.; Begum, J.; Zhi, Z.; Pezhman, L.; Tull, S.; Maione, F.; Iqbal, A.J. Galectin-9 supports primary T cell transendothelial migration in a glycan and integrin dependent manner. Biomed. Pharmacother. 2022, 151, 113171. [Google Scholar] [CrossRef]
- Iqbal, A.J.; Krautter, F.; Blacksell, I.A.; Wright, R.D.; Austin-Williams, S.N.; Voisin, M.B.; Hussain, M.T.; Law, H.L.; Niki, T.; Hirashima, M. Galectin-9 mediates neutrophil capture and adhesion in a CD44 and β2 integrin-dependent manner. FASEB J. 2022, 36, e22065. [Google Scholar] [CrossRef]
- Chakraborty, A.; Staudinger, C.; King, S.L.; Erickson, F.C.; Lau, L.S.; Bernasconi, A.; Luscinskas, F.W.; Perlyn, C.; Dimitroff, C.J. Galectin-9 bridges human B cells to vascular endothelium while programming regulatory pathways. J. Autoimmun. 2021, 117, 102575. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Iwama, H.; Sakamoto, T.; Okura, R.; Kobayashi, K.; Takano, J.; Katsura, A.; Tatsuta, M.; Maeda, E.; Mimura, S. Galectin-9 suppresses the growth of hepatocellular carcinoma via apoptosis in vitro and in vivo. Int. J. Oncol. 2015, 46, 2419–2430. [Google Scholar] [CrossRef] [Green Version]
- Nobumoto, A.; Nagahara, K.; Oomizu, S.; Katoh, S.; Nishi, N.; Takeshita, K.; Niki, T.; Tominaga, A.; Yamauchi, A.; Hirashima, M. Galectin-9 suppresses tumor metastasis by blocking adhesion to endothelium and extracellular matrices. Glycobiology 2008, 18, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Enninga, E.A.L.; Chatzopoulos, K.; Butterfield, J.T.; Sutor, S.L.; Leontovich, A.A.; Nevala, W.K.; Flotte, T.J.; Markovic, S.N. CD206-positive myeloid cells bind galectin-9 and promote a tumor-supportive microenvironment. J. Pathol. 2018, 245, 468–477. [Google Scholar] [CrossRef]
- Yan, L.; Xiao, M.; Yuxin, M.; Yuxin, D.; Feng, J. A new emerging target in cancer immunotherapy: Galectin-9 (LGALS9). Genes Dis. 2022; in press. [Google Scholar]
- Swann, J.B.; Smyth, M.J. Immune surveillance of tumors. J. Clin. Investig. 2007, 117, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Gharib, K.; Han, A. Cancer immunosurveillance by T cells. Int. Rev. Cell Mol. Biol. 2019, 342, 149–173. [Google Scholar]
- Jhunjhunwala, S.; Hammer, C.; Delamarre, L. Antigen presentation in cancer: Insights into tumour immunogenicity and immune evasion. Nat. Rev. Cancer 2021, 21, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, K. Role of apoptosis resistance in immune evasion and metastasis of colorectal cancer. World J. Gastrointest. Oncol. 2010, 2, 399. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, A.; Lei, Q.; Zhang, Y. Tumor-intrinsic signaling pathways: Key roles in the regulation of the immunosuppressive tumor microenvironment. J. Hematol. Oncol. 2019, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Nüssing, S.; Trapani, J.A.; Parish, I.A. Revisiting T cell tolerance as a checkpoint target for cancer immunotherapy. Front. Immunol. 2020, 11, 589641. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.Y.; Li, Y.S.; Qiao, X.H.; Yang, J.W.; Guo, X.L. Targeting galectins in T cell-based immunotherapy within tumor microenvironment. Life Sci. 2021, 277, 119426. [Google Scholar] [CrossRef]
- Navarro, P.; Martínez-Bosch, N.; Blidner, A.G.; Rabinovich, G.A. Impact of galectins in resistance to anticancer therapies. Clin. Cancer Res. 2020, 26, 6086–6101. [Google Scholar] [CrossRef]
- Marrack, P.; Kappler, J. Control of T cell viability. Annu. Rev. Immunol. 2004, 22, 765. [Google Scholar] [CrossRef]
- Stillman, B.N.; Hsu, D.K.; Pang, M.; Brewer, C.F.; Johnson, P.; Liu, F.T.; Baum, L.G. Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J. Immunol. 2006, 176, 778–789. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef]
- Nagahara, K.; Arikawa, T.; Oomizu, S.; Kontani, K.; Nobumoto, A.; Tateno, H.; Watanabe, K.; Niki, T.; Katoh, S.; Miyake, M. Galectin-9 increases Tim-3+ dendritic cells and CD8+ T cells and enhances antitumor immunity via galectin-9-Tim-3 interactions. J. Immunol. 2008, 181, 7660–7669. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Sun, L.; Li, C.F.; Wang, Y.H.; Yao, J.; Li, H.; Yan, M.; Chang, W.C.; Hsu, J.M.; Cha, J.H.; et al. Galectin-9 interacts with PD-1 and TIM-3 to regulate T cell death and is a target for cancer immunotherapy. Nat. Commun. 2021, 12, 832. [Google Scholar] [CrossRef] [PubMed]
- Camby, I.; Le Mercier, M.; Lefranc, F.; Kiss, R. Galectin-1: A small protein with major functions. Glycobiology 2006, 16, 137R–157R. [Google Scholar] [CrossRef] [PubMed]
- Perillo, N.L.; Pace, K.E.; Seilhamer, J.J.; Baum, L.G. Apoptosis of T cells mediated by galectin-1. Nature 1995, 378, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Walzel, H.; Fahmi, A.A.; Eldesouky, M.A.; Abou-Eladab, E.F.; Waitz, G.; Brock, J.; Tiedge, M. Effects of N-glycan processing inhibitors on signaling events and induction of apoptosis in galectin-1-stimulated Jurkat T lymphocytes. Glycobiology 2006, 16, 1262–1271. [Google Scholar] [CrossRef]
- Bi, S.; Earl, L.A.; Jacobs, L.; Baum, L.G. Structural features of galectin-9 and galectin-1 that determine distinct T cell death pathways. J. Biol. Chem. 2008, 283, 12248–12258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Nguyen, J.T.; Evans, D.P.; Galvan, M.; Pace, K.E.; Leitenberg, D.; Bui, T.N.; Baum, L.G. CD45 modulates galectin-1-induced T cell death: Regulation by expression of core 2 O-glycans. J. Immunol. 2001, 167, 5697–5707. [Google Scholar] [CrossRef] [Green Version]
- Fukumori, T.; Takenaka, Y.; Yoshii, T.; Kim, H.-R.C.; Hogan, V.; Inohara, H.; Kagawa, S.; Raz, A. CD29 and CD7 mediate galectin-3-induced type II T-cell apoptosis. Cancer Res. 2003, 63, 8302–8311. [Google Scholar]
- Seyrek, K.; Richter, M.; Lavrik, I.N. Decoding the sweet regulation of apoptosis: The role of glycosylation and galectins in apoptotic signaling pathways. Cell Death Differ. 2019, 26, 981–993. [Google Scholar] [CrossRef]
- Elad-Sfadia, G.; Haklai, R.; Balan, E.; Kloog, Y. Galectin-3 augments K-Ras activation and triggers a Ras signal that attenuates ERK but not phosphoinositide 3-kinase activity. J. Biol. Chem. 2004, 279, 34922–34930. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Liu, L.; Zhao, Z.; Zhang, Z.; Guan, Y.; Cheng, H.; Zhou, Y.; Tai, G. The N-terminal tail coordinates with carbohydrate recognition domain to mediate galectin-3 induced apoptosis in T cells. Oncotarget 2017, 8, 49824–49838. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, G.; Alonso, C.; Sotomayor, C.; Durand, S.; Bocco, J.; Riera, C. Molecular mechanisms implicated in galectin-1-induced apoptosis: Activation of the AP-1 transcription factor and downregulation of Bcl-2. Cell Death Differ. 2000, 7, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Brandt, B.; Abou-Eladab, E.F.; Tiedge, M.; Walzel, H. Role of the JNK/c-Jun/AP-1 signaling pathway in galectin-1-induced T-cell death. Cell Death Dis. 2010, 1, e23. [Google Scholar] [CrossRef] [PubMed]
- Kashio, Y.; Nakamura, K.; Abedin, M.J.; Seki, M.; Nishi, N.; Yoshida, N.; Nakamura, T.; Hirashima, M. Galectin-9 induces apoptosis through the calcium-calpain-caspase-1 pathway. J. Immunol. 2003, 170, 3631–3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roudko, V.; Greenbaum, B.; Bhardwaj, N. Computational prediction and validation of tumor-associated neoantigens. Front. Immunol. 2020, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Csiszár, A.; Szentes, T.; Haraszti, B.; Balázs, A.; Petrányi, G.G.; Pócsik, É. The pattern of cytokine gene expression in human colorectal carcinoma. Pathol. Oncol. Res. 2004, 10, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.A.; Bianco, G.A.; Ilarregui, J.M.; Croci, D.O.; Correale, J.; Hernandez, J.D.; Zwirner, N.W.; Poirier, F.; Riley, E.M.; Baum, L.G. Differential glycosylation of TH1, TH2 and TH-17 effector cells selectively regulates susceptibility to cell death. Nat. Immunol. 2007, 8, 825–834. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, L. Sweetening the deal: Glycosylation and its clinical applications. J. Biomed. Sci. 2020, 9, 3–9. [Google Scholar] [CrossRef]
- Ilarregui, J.M.; Croci, D.O.; Bianco, G.A.; Toscano, M.A.; Salatino, M.; Vermeulen, M.E.; Geffner, J.R.; Rabinovich, G.A. Tolerogenic signals delivered by dendritic cells to T cells through a galectin-1-driven immunoregulatory circuit involving interleukin 27 and interleukin 10. Nat. Immunol. 2009, 10, 981–991. [Google Scholar] [CrossRef]
- Cedeno-Laurent, F.; Opperman, M.; Barthel, S.R.; Kuchroo, V.K.; Dimitroff, C.J. Galectin-1 triggers an immunoregulatory signature in Th cells functionally defined by IL-10 expression. J. Immunol. 2012, 188, 3127–3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juszczynski, P.; Ouyang, J.; Monti, S.; Rodig, S.J.; Takeyama, K.; Abramson, J.; Chen, W.; Kutok, J.L.; Rabinovich, G.A.; Shipp, M.A. The AP1-dependent secretion of galectin-1 by Reed Sternberg cells fosters immune privilege in classical Hodgkin lymphoma. Proc. Natl. Acad. Sci. USA 2007, 104, 13134–13139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Han, B.; Meng, X.; Duan, C.; Yang, C.; Wu, Z.; Magafurov, D.; Zhao, S.; Safin, S.; Jiang, C. Immunogenomic analysis reveals LGALS1 contributes to the immune heterogeneity and immunosuppression in glioma. Int. J. Cancer 2019, 145, 517–530. [Google Scholar] [CrossRef]
- Verschuere, T.; Toelen, J.; Maes, W.; Poirier, F.; Boon, L.; Tousseyn, T.; Mathivet, T.; Gerhardt, H.; Mathieu, V.; Kiss, R.; et al. Glioma-derived galectin-1 regulates innate and adaptive antitumor immunity. Int. J. Cancer 2014, 134, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Orozco, C.A.; Martinez-Bosch, N.; Guerrero, P.E.; Vinaixa, J.; Dalotto-Moreno, T.; Iglesias, M.; Moreno, M.; Djurec, M.; Poirier, F.; Gabius, H.-J. Targeting galectin-1 inhibits pancreatic cancer progression by modulating tumor–stroma crosstalk. Proc. Natl. Acad. Sci. USA 2018, 115, E3769–E3778. [Google Scholar] [CrossRef] [Green Version]
- Demotte, N.; Wieërs, G.; Van Der Smissen, P.; Moser, M.; Schmidt, C.; Thielemans, K.; Squifflet, J.-L.; Weynand, B.; Carrasco, J.; Lurquin, C. A Galectin-3 Ligand Corrects the Impaired Function of Human CD4 and CD8 Tumor-Infiltrating Lymphocytes and Favors Tumor Rejection in MiceGalectin-3 Ligand Corrects the Impaired Function of TIL. Cancer Res. 2010, 70, 7476–7488. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Alonso, M.; Hirsch, T.; Wildmann, C.; van der Bruggen, P. Galectin-3 captures interferon-gamma in the tumor matrix reducing chemokine gradient production and T-cell tumor infiltration. Nat. Commun. 2017, 8, 793. [Google Scholar] [CrossRef]
- Dardalhon, V.; Anderson, A.C.; Karman, J.; Apetoh, L.; Chandwaskar, R.; Lee, D.H.; Cornejo, M.; Nishi, N.; Yamauchi, A.; Quintana, F.J. Tim-3/galectin-9 pathway: Regulation of Th1 immunity through promotion of CD11b+ Ly-6G+ myeloid cells. J. Immunol. 2010, 185, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Thalhamer, T.; Franca, R.F.; Xiao, S.; Wang, C.; Hotta, C.; Zhu, C.; Hirashima, M.; Anderson, A.C.; Kuchroo, V.K. Galectin-9-CD44 interaction enhances stability and function of adaptive regulatory T cells. Immunity 2014, 41, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Burrell, R.A.; Swanton, C. Tumour heterogeneity and the evolution of polyclonal drug resistance. Mol. Oncol. 2014, 8, 1095–1111. [Google Scholar] [CrossRef]
- Kelderman, S.; Schumacher, T.N.; Haanen, J.B. Acquired and intrinsic resistance in cancer immunotherapy. Mol. Oncol. 2014, 8, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Lykken, J.M.; Horikawa, M.; Minard-Colin, V.; Kamata, M.; Miyagaki, T.; Poe, J.C.; Tedder, T.F. Galectin-1 drives lymphoma CD20 immunotherapy resistance: Validation of a preclinical system to identify resistance mechanisms. Blood 2016, 127, 1886–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambiar, D.K.; Aguilera, T.; Cao, H.; Kwok, S.; Kong, C.; Bloomstein, J.; Wang, Z.; Rangan, V.S.; Jiang, D.; von Eyben, R.; et al. Galectin-1-driven T cell exclusion in the tumor endothelium promotes immunotherapy resistance. J. Clin. Investig. 2019, 129, 5553–5567. [Google Scholar] [CrossRef]
- Yazdanifar, M.; Zhou, R.; Grover, P.; Williams, C.; Bose, M.; Moore, L.J.; Wu, S.-T.; Maher, J.; Dreau, D.; Mukherjee, P. Overcoming immunological resistance enhances the efficacy of a novel anti-tMUC1-CAR T cell treatment against pancreatic ductal adenocarcinoma. Cells 2019, 8, 1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Giobbie-Hurder, A.; Connolly, E.M.; Li, J.; Liao, X.; Severgnini, M.; Zhou, J.; Rodig, S.; Hodi, F.S. Anti-CTLA-4 based therapy elicits humoral immunity to galectin-3 in patients with metastatic melanoma. Oncoimmunology 2018, 7, e1440930. [Google Scholar] [CrossRef]
- Wu, X.; Li, J.; Connolly, E.M.; Liao, X.; Ouyang, J.; Giobbie-Hurder, A.; Lawrence, D.; McDermott, D.; Murphy, G.; Zhou, J.; et al. Combined Anti-VEGF and Anti-CTLA-4 Therapy Elicits Humoral Immunity to Galectin-1 Which Is Associated with Favorable Clinical Outcomes. Cancer Immunol. Res. 2017, 5, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Safety of GM-CT-01 with and without 5-Fluorouracil in Patients with Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/NCT00054977 (accessed on 11 November 2022).
- Study to Test the Benefit and Safety of GM-CT-01 in Combination with 5-FU to Treat Bile Duct and Gall Bladder Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00386516 (accessed on 11 November 2022).
- A New Agent GM-CT-01 in Combination with 5-FU, Avastin and Leucovorin in Subjects with Colorectal Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00388700 (accessed on 11 November 2022).
- Peptide Vaccinations Plus GM-CT-01 in Melanoma. Available online: https://clinicaltrials.gov/ct2/show/NCT01723813 (accessed on 11 November 2022).
- GCS-100LE in Combination with Etoposide and Dexamethasone in Relapsed or Refractory Diffuse Large B-Cell Lymphoma (GCS-100LE). Available online: https://clinicaltrials.gov/ct2/show/NCT00776802 (accessed on 11 November 2022).
- Study of the Safety of GCS-100 in Subjects with Chronic Lymphocytic Leukemia (PR-CS008). Available online: https://clinicaltrials.gov/ct2/show/NCT00514696 (accessed on 11 November 2022).
- Safety and Efficacy Study of GCS-100LE in the Treatment of Multiple Myeloma. Available online: https://clinicaltrials.gov/ct2/show/NCT00609817 (accessed on 11 November 2022).
- GR-MD-02 + Pembrolizumab versus Pembrolizumab Monotherapy in Melanoma and Squamous Cell Head and Neck Cancer Patients. Available online: https://clinicaltrials.gov/ct2/show/NCT04987996 (accessed on 11 November 2022).
- GR-MD-02 Plus Pembrolizumab in Melanoma, Non-Small Cell Lung Cancer, and Squamous Cell Head and Neck Cancer Catients. Available online: https://clinicaltrials.gov/ct2/show/NCT02575404 (accessed on 11 November 2022).
- A Study to Investigate the Safety and Efficacy of GB1211 (a Galectin-3 Inhibitor) in Combination with Atezolizumab in Patients with Non-Small Cell Lung Cancer (nsclc). Available online: https://clinicaltrials.gov/ct2/show/NCT05240131 (accessed on 11 November 2022).
- Study of Efficacy and Safety of MBG453 in Combination with Azacitidine in Subjects with Intermediate, High or Very High Risk Myelodysplastic Syndrome (Mds) as Per IPSS-R, or Chronic Myelomonocytic Leukemia-2 (CMML-2) (STIMULUS-MDS2). Available online: https://clinicaltrials.gov/ct2/show/NCT04266301 (accessed on 11 November 2022).
- A Study of MBG453 in Combination with Azacitidine and Venetoclax in AML Patients Unfit for Chemotherapy (STIMULUS-AML1). Available online: https://www.clinicaltrials.gov/ct2/show/NCT04150029 (accessed on 11 November 2022).
- Galectin Inhibitor (GR-MD-02) and Ipilimumab in Patients with Metastatic Melanoma. Available online: https://clinicaltrials.gov/ct2/show/NCT02117362 (accessed on 11 November 2022).
- Effect of Modified Citrus Pectin on PSA Kinetics in Biochemical Relapsed PC with Serial Increases in PSA. Available online: https://clinicaltrials.gov/ct2/show/NCT01681823 (accessed on 11 November 2022).
- LYT-200 Alone and in Combination with Chemotherapy or Anti-PD-1 in Patients with Metastatic Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/NCT04666688 (accessed on 11 November 2022).
- Phase I-Ib/II Study of MBG453 as Single Agent and in Combination with PDR001 in Patients with Advanced Malignancies. Available online: https://clinicaltrials.gov/ct2/show/NCT02608268 (accessed on 11 November 2022).
- Study of PDR001 and/or MBG453 in Combination with Decitabine in Patients with AML or High Risk MDS. Available online: https://clinicaltrials.gov/ct2/show/NCT03066648 (accessed on 11 November 2022).
- Trial of Anti-Tim-3 in Combination with Anti-PD-1 and SRS in Recurrent GBM. Available online: https://clinicaltrials.gov/ct2/show/NCT03961971 (accessed on 11 November 2022).
- Van Beijnum, J.R.; Thijssen, V.L.; Läppchen, T.; Wong, T.J.; Verel, I.; Engbersen, M.; Schulkens, I.A.; Rossin, R.; Grüll, H.; Griffioen, A.W. A key role for galectin-1 in sprouting angiogenesis revealed by novel rationally designed antibodies. Int. J. Cancer 2016, 139, 824–835. [Google Scholar] [CrossRef]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Ochi, A.; Heindel, D.W.; Lee, K.B.; Zambirinis, C.P.; Pandian, G.S.B.; Savadkar, S.; et al. Dectin 1 activation on macrophages by galectin 9 promotes pancreatic carcinoma and peritumoral immune tolerance. Nat. Med. 2017, 23, 556–567. [Google Scholar] [CrossRef]
- Sabatos-Peyton, C.; Longmire, T.; Baker, L.; Patel, N.; Wavreille, A.-S.; Verneret, M.; Jayaraman, P.; Jiang, X.; Schwartz, S.; Cremasco, V. 439 Dual modes of action for anti-TIM-3 antibody MBG453 in myelodysplastic syndromes (MDS) and acute myeloid leukemia (AML): Preclinical evidence for immune-mediated and anti-leukemic activity. BMJ Specialist J. 2020, 8. [Google Scholar]
- Zeidan, A.M.; Esteve, J.; Giagounidis, A.; Kim, H.-J.; Miyazaki, Y.; Platzbecker, U.; Schuh, A.C.; Sekeres, M.A.; Westermann, J.; Xiao, Z. The STIMULUS Program: Clinical Trials Evaluating Sabatolimab (MBG453) Combination Therapy in Patients (Pts) with Higher-Risk Myelodysplastic Syndromes (HR-MDS) or Acute Myeloid Leukemia (AML). Blood 2020, 136, 45–46. [Google Scholar] [CrossRef]
- Linclau, B.; Ardá, A.; Reichardt, N.C.; Sollogoub, M.; Unione, L.; Vincent, S.P.; Jiménez-Barbero, J. Fluorinated carbohydrates as chemical probes for molecular recognition studies. Current status and perspectives. Chem. Soc. Rev. 2020, 49, 3863–3888. [Google Scholar] [CrossRef] [PubMed]
- Barthel, S.R.; Antonopoulos, A.; Cedeno-Laurent, F.; Schaffer, L.; Hernandez, G.; Patil, S.A.; North, S.J.; Dell, A.; Matta, K.L.; Neelamegham, S.; et al. Peracetylated 4-fluoro-glucosamine reduces the content and repertoire of N- and O-glycans without direct incorporation. J. Biol. Chem. 2011, 286, 21717–21731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cedeno-Laurent, F.; Opperman, M.J.; Barthel, S.R.; Hays, D.; Schatton, T.; Zhan, Q.; He, X.; Matta, K.L.; Supko, J.G.; Frank, M.H. Metabolic inhibition of galectin-1-binding carbohydrates accentuates antitumor immunity. J. Investig. Dermatol. 2012, 132, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Rillahan, C.D.; Antonopoulos, A.; Lefort, C.T.; Sonon, R.; Azadi, P.; Ley, K.; Dell, A.; Haslam, S.M.; Paulson, J.C. Global metabolic inhibitors of sialyl-and fucosyltransferases remodel the glycome. Nat. Chem. Biol. 2012, 8, 661–668. [Google Scholar] [CrossRef]
- Bull, C.; Boltje, T.J.; Balneger, N.; Weischer, S.M.; Wassink, M.; van Gemst, J.J.; Bloemendal, V.R.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic Acid Blockade Suppresses Tumor Growth by Enhancing T-cell-Mediated Tumor Immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.M.; Correia, A.; Pereira, M.S.; Almeida, C.R.; Alves, I.; Pinto, V.; Catarino, T.A.; Mendes, N.; Leander, M.; Oliva-Teles, M.T. Metabolic control of T cell immune response through glycans in inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2018, 115, E4651–E4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirois, S.; Giguere, D.; Roy, R. A first QSAR model for galectin-3 glycomimetic inhibitors based on 3D docked structures. Med. Chem. 2006, 2, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.; Sanam, S.; Alvala, R.; Alvala, M. An updated patent review of galectin-1 and galectin-3 inhibitors and their potential therapeutic applications (2016-present). Expert Opin. Ther. Pat. 2021, 31, 709–721. [Google Scholar] [CrossRef]
- Kurfiřt, M.; Dračínský, M.; Červenková Šťastná, L.; Cuřínová, P.; Hamala, V.; Hovorková, M.; Bojarová, P.; Karban, J. Selectively Deoxyfluorinated N-Acetyllactosamine Analogues as 19F NMR Probes to Study Carbohydrate-Galectin Interactions. Chem.—A Eur. J. 2021, 27, 13040–13051. [Google Scholar] [CrossRef]
- Heine, V.; Hovorkova, M.; Vlachova, M.; Filipova, M.; Bumba, L.; Janouskova, O.; Hubalek, M.; Cvacka, J.; Petraskova, L.; Pelantova, H.; et al. Immunoprotective neo-glycoproteins: Chemoenzymatic synthesis of multivalent glycomimetics for inhibition of cancer-related galectin-3. Eur. J. Med. Chem. 2021, 220, 113500. [Google Scholar] [CrossRef]
- Wdowiak, K.; Francuz, T.; Gallego-Colon, E.; Ruiz-Agamez, N.; Kubeczko, M.; Grochoła, I.; Wojnar, J. Galectin Targeted Therapy in Oncology: Current Knowledge and Perspectives. Int. J. Mol. Sci. 2018, 19, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic cancer vaccines. Nat. Rev. Cancer 2021, 21, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, S.K.; Perez-Penco, M.; Hübbe, M.L.; Martinenaite, E.; Orebo Holmström, M.; Weis-Banke, S.E.; Grønne Dahlager Jørgensen, N.; Jørgensen, M.A.; Munir Ahmad, S.; Jensen, K.M.; et al. Peptide vaccination activating Galectin-3-specific T cells offers a novel means to target Galectin-3-expressing cells in the tumor microenvironment. Oncoimmunology 2022, 11, 2026020. [Google Scholar] [CrossRef] [PubMed]
- Femel, J.; van Hooren, L.; Herre, M.; Cedervall, J.; Saupe, F.; Huijbers, E.J.M.; Verboogen, D.R.J.; Reichel, M.; Thijssen, V.L.; Griffioen, A.W.; et al. Vaccination against galectin-1 promotes cytotoxic T-cell infiltration in melanoma and reduces tumor burden. Cancer Immunol. Immunother. 2022, 71, 2029–2040. [Google Scholar] [CrossRef]
- Liebscher, L.; Weissenborn, C.; Langwisch, S.; Gohlke, B.O.; Preissner, R.; Rabinovich, G.A.; Christiansen, N.; Christiansen, H.; Zenclussen, A.C.; Fest, S. A minigene DNA vaccine encoding peptide epitopes derived from Galectin-1 has protective antitumoral effects in a model of neuroblastoma. Cancer Lett. 2021, 509, 105–114. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Büll, C.; Chen, Y.-H.; Wandall, H.H.; Yang, Z.; Clausen, H. Genetic glycoengineering in mammalian cells. J. Biol. Chem. 2021, 296, 100448. [Google Scholar] [CrossRef]
- Rini, J.M.; Moremen, K.W.; Davis, B.G.; Esko, J.D. Glycosyltransferases and glycan-processing enzymes. In Essentials of Glycobiology [Internet], 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022. [Google Scholar]
- Costa, A.F.; Campos, D.; Reis, C.A.; Gomes, C. Targeting glycosylation: A new road for cancer drug discovery. Trends Cancer 2020, 6, 757–766. [Google Scholar] [CrossRef]
- Griffin, M.E.; Hsieh-Wilson, L.C. Glycan engineering for cell and developmental biology. Cell Chem. Biol. 2016, 23, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Stolfa, G.; Mondal, N.; Zhu, Y.; Yu, X.; Buffone, A.; Neelamegham, S. Using CRISPR-Cas9 to quantify the contributions of O-glycans, N-glycans and Glycosphingolipids to human leukocyte-endothelium adhesion. Sci. Rep. 2016, 6, 30392. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, S.; Halim, A.; Schulz, M.A.; Frodin, M.; Rahman, S.H.; Vester-Christensen, M.B.; Behrens, C.; Kristensen, C.; Vakhrushev, S.Y. Engineered CHO cells for production of diverse, homogeneous glycoproteins. Nat. Biotechnol. 2015, 33, 842–844. [Google Scholar] [CrossRef]
- Pereira, M.S.; Alves, I.; Vicente, M.; Campar, A.; Silva, M.C.; Padrão, N.A.; Pinto, V.; Fernandes, Â.; Dias, A.M.; Pinho, S.S. Glycans as Key Checkpoints of T Cell Activity and Function. Front. Immunol 2018, 9, 2754. [Google Scholar] [CrossRef] [PubMed]
- Giovannone, N.; Antonopoulos, A.; Liang, J.; Geddes Sweeney, J.; Kudelka, M.R.; King, S.L.; Lee, G.S.; Cummings, R.D.; Dell, A.; Barthel, S.R.; et al. Human B Cell Differentiation Is Characterized by Progressive Remodeling of O-Linked Glycans. Front. Immunol. 2018, 9, 2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, J.G.; Liang, J.; Antonopoulos, A.; Giovannone, N.; Kang, S.; Mondala, T.S.; Head, S.R.; King, S.L.; Tani, Y.; Brackett, D.; et al. Loss of GCNT2/I-branched glycans enhances melanoma growth and survival. Nat. Commun. 2018, 9, 3368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A.; Perez, M.; Carroll, J.D.; Antonopoulos, A.; Dell, A.; Ortega, L.; Mohammed, N.B.B.; Wells, M.; Staudinger, C.; Griswold, A.; et al. Hypoxia controls the glycome signature and galectin-8—Ligand axis to promote pro-tumorigenic properties of metastatic melanoma. J. Investig. Dermatol. 2022; in press. [Google Scholar] [CrossRef]
- Schaefer, K.; Webb, N.E.; Pang, M.; Hernandez-Davies, J.E.; Lee, K.P.; Gonzalez, P.; Douglass, M.V.; Lee, B.; Baum, L.G. Galectin-9 binds to O-glycans on protein disulfide isomerase. Glycobiology 2017, 27, 878–887. [Google Scholar] [CrossRef] [Green Version]
- Thiemann, S.; Man, J.H.; Chang, M.H.; Lee, B.; Baum, L.G. Galectin-1 regulates tissue exit of specific dendritic cell populations. J. Biol. Chem. 2015, 290, 22662–22677. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, V.L.; Postel, R.; Brandwijk, R.J.; Dings, R.P.; Nesmelova, I.; Satijn, S.; Verhofstad, N.; Nakabeppu, Y.; Baum, L.G.; Bakkers, J.; et al. Galectin-1 is essential in tumor angiogenesis and is a target for antiangiogenesis therapy. Proc. Natl. Acad. Sci. USA 2006, 103, 15975–15980. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.C.; Pang, M.; Hsu, D.K.; Liu, F.T.; de Vos, S.; Gascoyne, R.D.; Said, J.; Baum, L.G. Galectin-3 binds to CD45 on diffuse large B-cell lymphoma cells to regulate susceptibility to cell death. Blood 2012, 120, 4635–4644. [Google Scholar] [CrossRef] [Green Version]
- Videla-Richardson, G.A.; Morris-Hanon, O.; Torres, N.I.; Esquivel, M.I.; Vera, M.B.; Ripari, L.B.; Croci, D.O.; Sevlever, G.E.; Rabinovich, G.A. Galectins as Emerging Glyco-Checkpoints and Therapeutic Targets in Glioblastoma. Int. J. Mol. Sci. 2021, 23, 316. [Google Scholar] [CrossRef]
- Mondal, N.; Silva, M.; Castano, A.P.; Maus, M.V.; Sackstein, R. Glycoengineering of chimeric antigen receptor (CAR) T-cells to enforce E-selectin binding. J. Biol. Chem. 2019, 294, 18465–18474. [Google Scholar] [CrossRef]
- Sackstein, R. The first step in adoptive cell immunotherapeutics: Assuring cell delivery via glycoengineering. Front. Immunol. 2019, 9, 3084. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, M.; Liu, Z.; Zhang, L.; Felding, B.H.; Moremen, K.W.; Lauvau, G.; Abadier, M.; Ley, K.; Wu, P. A single-step chemoenzymatic reaction for the construction of antibody–cell conjugates. ACS Cent. Sci. 2018, 4, 1633–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackstein, R. Glycosyltransferase-programmed stereosubstitution (GPS) to create HCELL: Engineering a roadmap for cell migration. Immunol. Rev. 2009, 230, 51–74. [Google Scholar] [CrossRef] [Green Version]
- Sackstein, R.; Merzaban, J.S.; Cain, D.W.; Dagia, N.M.; Spencer, J.A.; Lin, C.P.; Wohlgemuth, R. Ex vivo glycan engineering of CD44 programs human multipotent mesenchymal stromal cell trafficking to bone. Nat. Med. 2008, 14, 181–187. [Google Scholar] [CrossRef]
- Sackstein, R. Glycoengineering of HCELL, the human bone marrow homing receptor: Sweetly programming cell migration. Ann. Biomed. Eng. 2012, 40, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Dimitroff, C.J.; Lee, J.Y.; Rafii, S.; Fuhlbrigge, R.C.; Sackstein, R. CD44 is a major E-selectin ligand on human hematopoietic progenitor cells. J. Cell Biol. 2001, 153, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Dykstra, B.; Lee, J.; Mortensen, L.J.; Yu, H.; Wu, Z.L.; Lin, C.P.; Rossi, D.J.; Sackstein, R. Glycoengineering of E-Selectin Ligands by Intracellular versus Extracellular Fucosylation Differentially Affects Osteotropism of Human Mesenchymal Stem Cells. Stem Cells 2016, 34, 2501–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauss, M.; Donia, M.; Harbst, K.; Andersen, R.; Mitra, S.; Rosengren, F.; Salim, M.; Vallon-Christersson, J.; Törngren, T.; Kvist, A.; et al. Mutational and putative neoantigen load predict clinical benefit of adoptive T cell therapy in melanoma. Nat. Commun. 2017, 8, 1738. [Google Scholar] [CrossRef] [Green Version]
- Verdegaal, E.M.; Visser, M.; Ramwadhdoebé, T.H.; van der Minne, C.E.; van Steijn, J.A.; Kapiteijn, E.; Haanen, J.B.; van der Burg, S.H.; Nortier, J.W.; Osanto, S. Successful treatment of metastatic melanoma by adoptive transfer of blood-derived polyclonal tumor-specific CD4+ and CD8+ T cells in combination with low-dose interferon-alpha. Cancer Immunol. Immunother. 2011, 60, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Melief, S.M.; Visser, M.; van der Burg, S.H.; Verdegaal, E.M.E. IDO and galectin-3 hamper the ex vivo generation of clinical grade tumor-specific T cells for adoptive cell therapy in metastatic melanoma. Cancer Immunol. Immunother. 2017, 66, 913–926. [Google Scholar] [CrossRef] [Green Version]
- Steentoft, C.; Migliorini, D.; King, T.R.; Mandel, U.; June, C.H.; Posey, A.D., Jr. Glycan-directed CAR-T cells. Glycobiology 2018, 28, 656–669. [Google Scholar] [CrossRef] [Green Version]
- Hombach, A.; Wieczarkowiecz, A.; Marquardt, T.; Heuser, C.; Usai, L.; Pohl, C.; Seliger, B.; Abken, H. Tumor-specific T cell activation by recombinant immunoreceptors: CD3ζ signaling and CD28 costimulation are simultaneously required for efficient IL-2 secretion and can be integrated into one combined CD28/CD3ζ signaling receptor molecule. J. Immunol. 2001, 167, 6123–6131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach, A.; Köhler, H.; Rappl, G.; Abken, H. Human CD4+ T cells lyse target cells via granzyme/perforin upon circumvention of MHC class II restriction by an antibody-like immunoreceptor. J. Immunol. 2006, 177, 5668–5675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaresan, P.R.; Manuri, P.R.; Albert, N.D.; Maiti, S.; Singh, H.; Mi, T.; Roszik, J.; Rabinovich, B.; Olivares, S.; Krishnamurthy, J. Bioengineering T cells to target carbohydrate to treat opportunistic fungal infection. Proc. Natl. Acad. Sci. USA 2014, 111, 10660–10665. [Google Scholar] [CrossRef] [PubMed]
- Davenport, A.J.; Jenkins, M.R.; Cross, R.S.; Yong, C.S.; Prince, H.M.; Ritchie, D.S.; Trapani, J.A.; Kershaw, M.H.; Darcy, P.K.; Neeson, P.J. CAR-T Cells Inflict Sequential Killing of Multiple Tumor Target CellsIndividual CAR-T Cells Kill Multiple Tumor Targets. Cancer Immunol. Res. 2015, 3, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Koehler, H.; Kofler, D.; Hombach, A.; Abken, H. CD28 Costimulation overcomes transforming growth factor-β–mediated repression of proliferation of redirected human CD4+ and CD8+ T Cells in an antitumor cell attack. Cancer Res. 2007, 67, 2265–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, S.; Martin, S. Mechanisms of granule-dependent killing. Cell Death Differ. 2008, 15, 251–262. [Google Scholar] [CrossRef] [Green Version]
- de Saint Basile, G.; Ménasché, G.; Fischer, A. Molecular mechanisms of biogenesis and exocytosis of cytotoxic granules. Nat. Rev. Immunol. 2010, 10, 568–579. [Google Scholar] [CrossRef]
- Porter, D.L.; Levine, B.L.; Kalos, M.; Bagg, A.; June, C.H. Chimeric antigen receptor–modified T cells in chronic lymphoid leukemia. N. Engl. J. Med. 2011, 365, 725–733. [Google Scholar] [CrossRef] [Green Version]
- Brentjens, R.J.; Curran, K.J. Novel cellular therapies for leukemia: CAR-modified T cells targeted to the CD19 antigen. Hematology 2012, 2012, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-C.S.; Lo, A.; Scholler, J.; Sun, J.; Majumdar, R.S.; Kapoor, V.; Antzis, M.; Cotner, C.E.; Johnson, L.A.; Durham, A.C. Targeting Fibroblast Activation Protein in Tumor Stroma with Chimeric Antigen Receptor T Cells Can Inhibit Tumor Growth and Augment Host Immunity without Severe ToxicityFAP-Redirected CAR T Cells. Cancer Immunol. Res. 2014, 2, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Scarfò, I.; Maus, M.V. Current approaches to increase CAR T cell potency in solid tumors: Targeting the tumor microenvironment. J. Immunother. Cancer 2017, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Feng, Z.; Ma, J.; Ling, S.; Cao, Y.; Gurung, B.; Wu, Y.; Katona, B.W.; O’Dwyer, K.P.; Siegel, D.L. Bispecific and split CAR T cells targeting CD13 and TIM3 eradicate acute myeloid leukemia. Blood 2020, 135, 713–723. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Galectin | Type of Inhibitor | Molecule | Cancer Model | Clinical Trial | Combination Therapy | Citation |
|---|---|---|---|---|---|---|
| Gal-1/Gal-3 | Allosteric. inhibitor | GM-CT-01 (Davanat®) | Colorectal, lung, breast, head and neck, and prostate cancers | NCT00054977 | 5-fluorouracil (Adrucil®) | [120] |
| Gal-3 | Allosteric inhibitor | GM-CT-01 (Davanat®) | Cancers of the bile duct and gallbladder | NCT00386516 | 5-fluorouracil (Adrucil®), folinic acid (Leucovorin), bevacizumab (Avastin®) | [121] |
| Gal-3 | Allosteric inhibitor | GM-CT-01 (Davanat®) | Colorectal cancer | NCT00388700 | 5-fluorouracil, folinic acid (Leucovorin), bevacizumab (Avastin®) | [122] |
| Gal-3 | Allosteric inhibitor | GM-CT-01 (Davanat®) | Metastatic melanoma | NCT01723813 | Tumor-specific peptide: MAGE-3. A1/or NA17.A2 | [123] |
| Gal-3 | Competitive inhibitor | GCS-100 Modified citrus pectin (MCP) | Diffuse large B-cell lymphoma | NCT00776802 | Etoposide (Vepesid®); dexamethasone (Maxidex) | [124] |
| Gal-3 | Competitive inhibitor | GCS-100 MCP | Chronic lymphocytic leukemia | NCT00514696 | [125] | |
| Gal-3 | Competitive inhibitor | GCS-100 MCP | Multiple myeloma | NCT00609817 | Bortezomib (VelcadeTM)/ dexamethasone (Maxidex) | [126] |
| Gal-3 | Competitive inhibitor | GR-MD-02 (Belapectin) | Metastatic melanoma, head and neck squamous cell carcinoma | NCT04987996 | Pembrolizumab (Keytruda®) | [127] |
| Gal-3 | Competitive inhibitor | GR-MD-02 (Belapectin) | Melanoma, non-small-cell lung cancer, and squamous cell head and neck cancer | NCT02575404 | Pembrolizumab (Keytruda®) | [128] |
| Gal-3 | Allosteric inhibitor | GB1211 | Non-small-cell lung cancer | NCT05240131 | Atezolizumab (Tecentriq®) | [129] |
| Gal-3 | Neutralizing antibody | MBG453 | Myelodysplastic syndrome or chronic myelomonocytic leukemia-2 | NCT04266301 | Azacitidine (VidazaTM) | [130] |
| Gal-3 | Neutralizing antibody | MBG453 | Acute myeloid leukemia | NCT04150029 | Azacitidine (Onureg®) and venetoclax (Venclexta®) | [131] |
| Gal-3 | Competitive inhibitor | GR-MD-02 (Belapectin) | Metastatic melanoma | NCT02117362 | Ipilimumab (Yervoy®) | [132] |
| Gal-3 | Competitive inhibitor | Modified Citrus Pectin (MCP, PectaSol-C) | Prostate cancer | NCT01681823 | 5-fluorouracil (Adrucil®) | [133] |
| Gal-9 | Neutralizing antibody | LYT-200 | Cholangiocarcinoma, colorectal cancer, pancreatic cancer | NCT04666688 | Anti-PD-1 mAb, gemcitabine (GemzarTM)/ nab-paclitaxel | [134] |
| Gal-9/Tim-3 | Neutralizing antibody | MBG453 (Sabatolimab) | Colon cancer and melanoma | NCT02608268 | PDR001 (Spartalizumab) | [135] |
| Gal-9/Tim-3 | Neutralizing antibody | MBG453 (Sabatolimab) | Acute myeloid leukemia | NCT04150029 | Azacitidine (Onureg®) and venetoclax | [131] |
| Gal-9/Tim-3 | Neutralizing antibody | MBG453 (Sabatolimab) | Myeloid leukemia, acute myelodysplastic syndromes, preleukemia, myelomonocytic leukemia | NCT03066648 | Decitabine (DacogenTM) or azacitidine (Onureg®) | [136] |
| Gal-9/Tim-3 | Neutralizing antibody | MBG453 (Sabatolimab) | Glioblastoma multiforme | NCT03961971 | - | [137] |
| Gal-9/Tim-3 | Neutralizing antibody | MBG453 (Sabatolimab) | Myelodysplastic syndrome or chronic myelomonocytic leukemia-2 | NCT04266301 | Azacitidine (Onureg®) | [130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, L.S.; Mohammed, N.B.B.; Dimitroff, C.J. Decoding Strategies to Evade Immunoregulators Galectin-1, -3, and -9 and Their Ligands as Novel Therapeutics in Cancer Immunotherapy. Int. J. Mol. Sci. 2022, 23, 15554. https://doi.org/10.3390/ijms232415554
Lau LS, Mohammed NBB, Dimitroff CJ. Decoding Strategies to Evade Immunoregulators Galectin-1, -3, and -9 and Their Ligands as Novel Therapeutics in Cancer Immunotherapy. International Journal of Molecular Sciences. 2022; 23(24):15554. https://doi.org/10.3390/ijms232415554
Chicago/Turabian StyleLau, Lee Seng, Norhan B. B. Mohammed, and Charles J. Dimitroff. 2022. "Decoding Strategies to Evade Immunoregulators Galectin-1, -3, and -9 and Their Ligands as Novel Therapeutics in Cancer Immunotherapy" International Journal of Molecular Sciences 23, no. 24: 15554. https://doi.org/10.3390/ijms232415554