Abstract
In this opinion article, we have analyzed the relevancy of a hypothesis which is based on the idea that in Arabidopsis thaliana jasmonic acid, a (JA)-mediated defense system against necrotrophic fungi is weakened when NO3− supply is high. Such a hypothesis is based on the fact that when NO3− supply is high, it induces an increase in the amount of bioactive ABA which induces the sequestration of the phosphatase ABI2 (PP2C) into the PYR/PYL/RCAR receptor. Consequently, the Ca sensors CBL1/9-CIPK23 are not dephosphorylated by ABI2, thus remaining able to phosphorylate targets such as AtNPF6.3 and AtKAT1, which are NO3− and K+ transporters, respectively. Therefore, the impact of phosphorylation on the regulation of these two transporters, could (1) reduce NO3− influx as in its phosphorylated state AtNPF6.3 shifts to low capacity state and (2) increase K+ influx, as in its phosphorylated state KAT1 becomes more active. It is also well known that in roots, K+ loading in the xylem and its transport to the shoot is activated in the presence of NO3−. As such, the enrichment of plant tissues in K+ can impair a jasmonic acid (JA) regulatory pathway and the induction of the corresponding biomarkers. The latter are known to be up-regulated under K+ deficiency and inhibited when K+ is resupplied. We therefore suggest that increased K+ uptake and tissue content induced by high NO3− supply modifies the JA regulatory pathway, resulting in a weakened JA-mediated plant’s defense system against necrotrophic fungi.
1. Introduction
1.1. The Dual Function of Nitrate as a Nutrient and Signaling Molecule
Nitrogen (N) is an essential inorganic nutrient to sustain plant development and growth. Unlike ammonium, when it occurs in soil, it is either adsorbed on clay particles or oxidized to nitrate by soil microorganisms. Nitrate (NO3−) is the main source of N available in the soil for plants. Probably because of its importance in mineral nutrition, plants have evolved two distinct NO3− transport and absorption systems in order to adapt to its availability in the soil: a high-affinity, low capacity, transporter system (HATS) that belongs to NRT2 (Nitrate Transporter 2) family and a low-affinity, high capacity, transporter system (LATS) that belongs to a family of proteins formerly named NRT1/PTR (Nitrate Transporter 1/Peptide Transporter), renamed NPF (NRT1/PTR Family) [1].
The function of NO3− as a signaling molecule has been studied in depth mostly in the model plant Arabidospsis thaliana, and notably in relation to the developmental plasticity of the root system architecture [2,3] that allows an optimal adaptation of the plant-to-soil N availability [2,4,5]. In particular, it has been shown that when there is a high supply of NO3−, lateral root (LR) elongation is inhibited right after their emergence from the primary root. It has been proposed that such an inhibitory mechanism is under the control of the tissue of NO3− content following the finding that in a nitrate reductase-deficient mutant, its systemic inhibitory effect on lateral root growth does not occur [5]. When high rates of NO3− were applied locally to a root system subjected to nitrate deficiency, LR growth was locally stimulated and oriented towards the source of nitrate. The localized effect of nitrate on LR growth was shown to be mediated by the nitrate transporter AtNPF6.3 (also known as NRT1.1/CHL1.5) [6]. It has been shown that under a high NO3− supply, AtNPF6.3-dependent auxin basipetal transport is inhibited by nitrate, leading to auxin accumulation in the LR tip, which stimulates LR emergence. Conversely, under a low nitrate supply, AtNPF6.3 transports auxin away from the LR tip, thus decreasing its content in the tip, and causing, therefore, the inhibition of LR outgrowth [7].
1.2. High-NO3− Supply Increases Biotic Stress Susceptibility
In comparison with the above-mentioned roles of nitrate in plant development, the role of this nutrient in plant response to pathogens is less known [8,9]. Still, on the basis of experimental and field experiments, it has been observed that overapplication of N fertilizer may affect plant–pathogen interaction, causing in some cases an enhanced disease susceptibility [10,11]. However, this view should be tempered as it appears that NO3− interference with plant defense machinery is more complex and specific to each pathosystem, as disease severity may be linked to either increasing or decreasing N fertilizer [12,13]. Therefore, considering N only from a nutritional angle, i.e., as a precursor of nutrients and defense molecules, seems unreliable [14].
Indeed, more recently, the idea of a signaling role of NO3− in plant health emerged and is supported by an increasing amount of evidence [8,9,10,15,16,17,18]. The studies suggest that NO3− may interfere as a signal molecule in the signaling pathways that lead in each pathosystem to the production of specific signaling molecules such as salicylic acid, or hormones such as ethylene and jasmonic acid (JA); in turn, these molecules trigger the expression of specific genes belonging to families such as pathogen related (PR) or the antifungal plant defensin family (PDF) [12,13,19,20,21]. In line with these considerations, one of the most relevant findings in our opinion was the discovery of an additional role for the high-affinity nitrate transporter of Arabidopsis thaliana AtNRT2.1 as being linked to plant defense against the bacterial pathogen Pseudomonas syringae pv tomato DC3000 (Pst) [15,22]. Authors found that a functional AtNRT2.1 antagonizes the priming of the plant’s defense against Pseudomonas syringae. Conversely, deletion mutant nrt2, in which the priming of salicylic acid signaling operated properly, showed a reduced susceptibility to the pathogen. Very interestingly, in nrt2, hormonal homeostasis was concomitantly affected with irregular function of JA and abscisic acid (ABA) pathways upon infection [15,22].
Studies on various pathosystems such as tomato/Oidium [23], Arabidopsis thaliana/Botrytis cinerea [8,9], and Arabidopsis thaliana/Alternaria brassisicola [24] report that high concentrations of NO3− in the growth medium increase plant susceptibility to necrotic fungi. Tests of pathogenicity following Arabidopsis thaliana inoculation with Botrytis cinerea carried out on plants grown on 0.5, 2, and 10 mM NO3− showed that leaf lesion areas (cm2) were larger in plants grown on 2 and 10 mM compared to those grown on 0.5 mM NO3− [9]. Furthermore, the lesion propagation rate (cm2/24 h) was positively correlated to NO3− concentration in the medium; for example, the propagation rate of the wild-type strain Bd90 was nil on leaves of plants grown on 0.5 mM NO3−, increasing to 0.1 at 2 mM NO3− and doubling to reach 0.2 at 10 mM NO3− [9]. Similarly, susceptibility of Arabidopsis thaliana to Alternaria brassisicola at the rosette stage was largely higher under nitrate (5 mM) compared to ammonium (5 mM) conditions [24]. The lesion area was tiny on leaves of ammonium-fed plants whereas it was almost 10 times bigger (ca. 0.7 mm2 at 7 days’ post-infection) in nitrate-fed plants [24].
After reviewing the literature, we attempted to decipher mechanisms through which NO3 supplied at high concentration would cause deleterious effects on plant health. For this aim, we focused on the well-studied model plant Arabidopsis thaliana and its interaction with necrotrophic fungi.
2. NO3− Uptake, Signaling, and Sensing Involves Calcineurin B-Like (CBL)-Interacting Protein Kinase (CIPK)
AtNPF6.3 was shown to interact with other molecules as a component of a macromolecular complex dedicated to nitrate sensing and signaling along with a protein kinase, CIPK23, calcium sensors (CBL1 and CBL9) [25], and ABI2, a phosphatase of the phosphatase 2C family [26]. CIPK23 belongs to a family of protein kinases (calcineurin B-like (CBL)-interacting protein kinase (CIPK)), encompassing 26 members in Arabidopsis thaliana [27]. Each CIPK specifically interacts with one or several of the 10 CBL calcium sensors to specifically decode calcium signals [28,29]. Specific roles for some CIPK–CBL pairs have been elucidated and several targets have been identified [30,31,32]. CBL1/9 interact with and activate CIPK23 that, in turn, phosphorylates AtNPF6.3, causing a decrease in its NO3− absorption capacity. The phosphatase ABI2, by dephosphorylating CIPK23 and the calcium sensor CBL1, counteracts their action on nitrate transport, signaling, and sensing by allowing AtNPF6.3 to remain in an unphosphorylated state [26].
3. High-NO3− Supply Induces an Increase in Bioactive ABA Content in Planta
Although deficiencies in essential mineral nutrients, e.g., N, phosphorus (P), and potassium (K), lead to common stress reactions such as an increase in reactive oxygen species, involving ABA can be different depending on the type of the nutrient [33]. Comparison of the changes in ABA concentration in Ricinus communis under mineral nutrients deficiencies showed contrasted results between P and N. The ABA concentration increased 29-fold in xylem sap under P limitation whereas it showed a 5-fold increase in the xylem with the increase in NO3− supply (see Figures 1 and 2 in [34]). Accordingly, ABA concentration increased in the leaves showing a linear positive correlation between ABA content and NO3− supply (Figure 1).
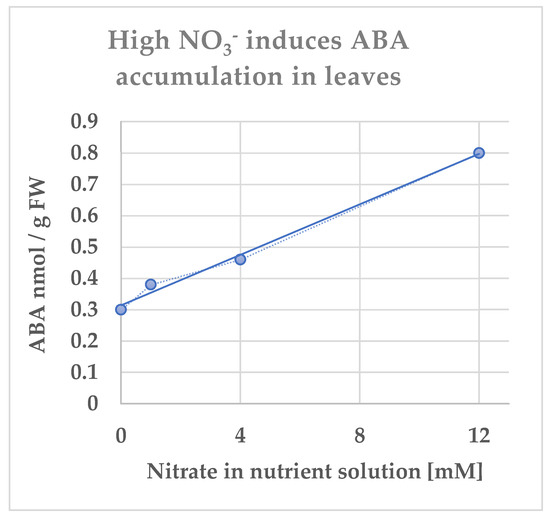
Figure 1.
Concentration of ABA per fresh weight of leaves of Ricinus communis grown under different nitrate concentrations 41 d after sowing. Data are from A. Peuke (Personal communication), see also Peuke et al. [34].
In Arabidopsis thaliana, ABA is mobilized for the local stimulation of lateral root elongation by patches of high NO3− [35]. Moreover, it has been shown that the effect of nitrate on ABA concentration is controlled by a signaling pathway that resulted in the release of bioactive ABA from the inactive conjugated form, ABA-glucose ester (ABA-GE), by the action of enzymes of the beta-glucosidase (BG) family [36]. This release catalyzed by the enzyme beta-Glucuronidase 1 primarily occurs in the root tips, which then allows it to transduce the NO3− mediated hormonal signal to other organs [36].
4. High-NO3− Regulates NO3− Uptake via an ABA-Induced Negative Feedback Loop
Soluble receptors of ABA have been thoroughly studied in Arabidopsis thaliana. They belong to a family of 14 proteins, named Pyrabactin Resistance/Pyrabactin-Like (PYR/PYL) or Regulatory Components of ABA Receptor (RCAR) and are referred to as PYR/PYL or PYR/PYL/RCAR. Mechanisms of ABA signaling pathway start with the structural changes in the PYR/PYL receptors induced by binding the phytohormone. Changes in the structure of the receptors allow them to sequester members of the clade A negative-regulating protein phosphatase 2Cs (PP2Cs) such as the phosphatase ABA insensitive (ABI2) [37,38]. Consequently, targets of ABI2 such as Ca2+ sensor/kinase complexes, CBL/CIPK, are not dephosphorylated by the phosphatase which affect their activity. Precise activation of CBL/CIPK complexes in order to phosphorylate their target proteins often requires CIPK autophosphorylation and CIPK-dependent phosphorylation of the Ca2+-sensor moiety in the associated CBL [39]. The interactions between CBL1, CIPK23, AtNPF6.3, and ABI2 were shown unequivocally in planta by bimolecular fluorescence complementation (BiFC) [26]. In vitro phosphorylation assays showed that CIPK23 autophosphorylation and CIPK23-dependent phosphorylation of CBL1 was dramatically lowered in the presence of ABI2, thus revealing that ABI2 effectively dephosphorylated CBL1 and CIPK23 (see Figure 4A in [26]).
In the presence of high exogenous NO3−, a gradual increase in bioactive ABA content in the root is triggered as mentioned above, and the latter is recognized by the receptor PYL/PYR, which after structural change sequesters the phosphatase ABI2. As a result, CBL1, CBL9, and CIPK23 complexes remain active and able to phosphorylate their targets, because these complexes cannot be dephosphorylated by ABI2.
Thus, ABA appears as a determinant in the regulation of NO3− uptake by NO3− itself through a phosphorylation process by recruiting the CBL1, CBL9 and CIPK23 complexes and actually acting as a signaling module responsible of the phosphorylation of AtNPF6.3, causing a decrease in its NO3− absorption capacity. This process has been demonstrated by Harris et al. [40] and has been described as a slow-acting negative feedback loop, activated by NO3− itself (Figure 2).
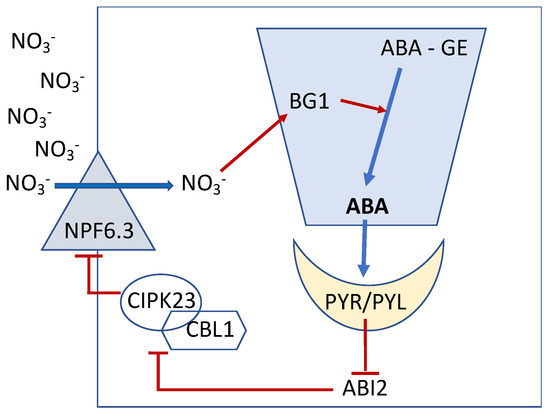
Figure 2.
Overview of NO3− signaling via the release of bioactive ABA in the Arabidopsis root tip. Nitrate is absorbed through the transporter and sensor AtNPF6.3 (AtNRT1.1). Under high levels of exogenous NO3−, this ion, once absorbed, rapidly stimulates the expression of the AtBG1 gene, encoding an ER-localized β-glucosidase, that cleaves the inactive ABA conjugate, ABA-glucose ester (ABA-GE), releasing bioactive ABA. The increase in ABA levels is a slow and gradual process during which ABA binds the intracellular PYR/PYL receptor, causing the inactivation of the ABA co-receptor, ABI2. Once ABI2 is inactivated, the CIPK23/CBL1 complex is free to phosphorylate AtNPF6.3, thus inhibiting its ability to transport NO3−. Schematic presentation modified from articles of Ondzighi–Assoume et al. and Harris [36,41].
5. ABA Functionally Links NO3− and K+ Uptake via the Action of CBL/CIPK
The signaling module constituted by CBL1, CBL9, and CIPK23 is also involved in the regulation of K+ uptake by regulating the cognate transporter, AtKAT1. This finding is supported by data obtained by omics-based techniques and molecular physiology approaches that have improved our understanding of the management of K+ acquisition by plants. At least 7 of the 9 members of the Shaker family of Arabidopsis were characterized in heterologous systems (e.g., Xenopus oocytes, insect and mammalian cell lines, and yeast) and found to be highly selective K+ channels with various rectification properties, i.e., inward, outward, and weakly inward. The functional data obtained in heterologous systems along with subcellular localization and characterization of a knockout mutant (akt1) showed that the inward rectifier AtAKT1 is the major K+-uptake system in the root, and is present in the root cortex, epidermis, and root hair [42]. In the same way as AtNPF6.3, AtAKT1 was found to be regulated by a calcium-dependent signaling pathway involving sensors of CBLs family and a target kinase [32,43,44]. A precise combination of a genetic approach (yeast two-hybrid) and electrophysiology (Xenopus oocyte patch-clamping) showed that CBL1 and CBL9 and their target kinase (CIPK23) form alternative complexes located at the plasma membrane where they jointly activate the AtAKT1 channel, thereby increasing K+ uptake capacity [43,45,46]. These experiments present evidence that CBL1/9-CIPK23 physically interact with AKT1 channel protein to switch on its activity by phosphorylation [47,48].
Interestingly, one can observe that when the availability of NO3− is high, AtNPF6.3 is negatively regulated with a decrease in the influx of NO3− via an ABA-mediated phosphorylation process, and concomitantly, AtKAT1 activity is upregulated leading to an increase in the influx of K+ (Figure 3). Moreover, it has been shown that the regulation of K+ loading in the xylem and its translocation to the shoots was also dependent on NO3− availability. More precisely, one could observe that the K+-mediated translocation to the shoot through the xylem-loading K+ channel SKOR (Stelar K+ Outward Rectifier) is stimulated in the presence of high amounts of NO3− [47,49]; consistently, expression of AtSKOR was upregulated by high NO3− (5 times higher under 10 mM compared to 1 mM) in roots of Arabidopsis Col-0 irrespective of the level of K+ supply (see Supplemental Figure S9 in [50]).
Furthermore, the link between these two ions is supported by the long-lasting observation that in most plant species K+ uptake from the soil is positively correlated with NO3− uptake [47,51,52]. This effect was explained at the whole plant physiology level taking into account charge balance in plant tissue, in particular xylem, where K+ would serve as counterion to compensate for the negative charge of NO3− [53,54,55]. This is very well illustrated by Engels and Marschner [56] who shown in maize that the K+ flux rate (µmol/h/g root) was 30% higher under nitrate compared to that under ammonium nutrition. Consequently, K+ absorption and accumulation would increase when NO3− availability and absorption increase.

Figure 3.
Schematic representation of functional links between NO3− and K+ uptake via the action of CBL/CIPK complex. Under high levels of exogenous NO3−, the CIPK23/CBL1 complex is set free to phosphorylate AtNPF6.3 and AtKAT1 thus leading to a decrease in NO3− influx and oppositely an increase in K+ influx. It is proposed that the increased K+ concentration inhibits JA-regulatory pathway of defense against pathogens. The latter is known to be up-regulated during K+ deficiency and inhibited upon K+ resupply [57,58,59,60,61].
Figure 3.
Schematic representation of functional links between NO3− and K+ uptake via the action of CBL/CIPK complex. Under high levels of exogenous NO3−, the CIPK23/CBL1 complex is set free to phosphorylate AtNPF6.3 and AtKAT1 thus leading to a decrease in NO3− influx and oppositely an increase in K+ influx. It is proposed that the increased K+ concentration inhibits JA-regulatory pathway of defense against pathogens. The latter is known to be up-regulated during K+ deficiency and inhibited upon K+ resupply [57,58,59,60,61].

6. Jasmonic Acid Links K+ Content and Defense against Necrotrophic Fungi
It has been proposed that necrotrophic pathogens induce JA-dependent defense in plants [62,63,64]. Among several K-dependent changes in metabolites of Arabidopsis thaliana, accumulation of indole and aliphatic glucosinolates appeared as a characteristic of K-deficient plants [57]. This finding is in agreement with transcriptome analyses in which expression of genes related to JA biosynthesis was enhanced in K-deficient Arabidopsis plants [58,59]. The gene encoding AtLOX2, which catalyzes the first committed step in JA biosynthesis [60,61], responded to low K prior to developmental symptoms, e.g., growth retardation and senescence, demonstrating that the induction of the JA pathway was not a secondary effect of stress symptoms [57]. Furthermore, levels of JA, as well as its precursors 12-oxo-phytodienoic acid (OPDA) and hydroxyl-12-oxo-octadecadienoic acids (HODs), were increased in K-deficient plants [57]. Expression of genes involved in JA signaling and response to biotic stress in particular defense genes dependent on the function of the JA receptor coronatine-insensitive 1 (COI1) was also boosted by K+ starvation [58,59].
It has been observed that a low NO3− supply to Arabidopsis mutant nrt1.5-5 caused K+ deficiency in the shoot as a result of an impairment of K+ loading in the xylem at the root level [50]. Interestingly, the survey by q-RT-PCR of the expression of 34 genes related to JA biosynthesis (e.g., AtLOX2), calcium signaling (e.g., ATCML41), defense (e.g., the JA-induced genes AtPDF1.2b, AtNATA1, and AtTPS4), secondary metabolism, and reactive oxygen species production were all more than twofold up-regulated in the mutant compared to the wild type (see Supplemental Figure S4 in [50]); 26 of the 34 tested genes were reported by Armengaud et al. [58] to be up-regulated by K+ starvation. Altogether, these results strengthen the idea that K+ status is an important player in plant response to necrotrophic fungi-induced disease by modulating JA synthesis and signaling pathways.
7. Conclusions
In the pathosystem Arabidopsis thaliana/Botrytis cinerea, only a small set of 182 genes exhibited an altered level of expression when NO3− is supplied to the plant [9]. Among those encoding proteins involved in plant defense, four genes, PME7, PR1, PR5, and PDF1.2a, were selected for a more detailed expression analysis. Interestingly, only the level of expression of PDF1.2a, a marker gene of the JA signaling pathway was fourfold higher in infected plants grown in the presence of low amounts of NO3−. These infected plants did not develop disease symptoms compared to the infected plants grown on high NO3− which developed visible symptoms of fungal infection (see Figure 4 in [9]). It is thus concluded that in infected plants when the supply of NO3− is high (2 or 10 mM), it counteracts the fungal-induced up-regulation of PDF1.2a, thus weakening plant defense against the fungi. Moreover, an NO3− supply above a certain threshold could contribute to plant tissue K+ enrichment which in turn is not favorable to the induction of the JA-dependent defense pathway (Figure 3). In line with this finding, the JA-dependent defense pathway is known to be up-regulated under K+ deficient conditions and inhibited when K+ is resupplied to the plant [57,65].
Author Contributions
A.L. wrote the original draft; A.L., B.H. and J.L. reviewed and edited the article. All authors have read and agreed to the published version of the manuscript.
Funding
Our research work is supported by University of Angers, INRAE, the French Region Pays de la Loire.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors are grateful to Andreas Peuke for providing unpublished data on ABA content of Ricinus communis under various nitrate regimes.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Léran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Forde, B.G. Regulation of Arabidopsis root development by nitrate availability. J. Exp. Bot. 2000, 51, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Rong, H.; Pilbeam, D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 2329–2338. [Google Scholar] [CrossRef] [PubMed]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, H.M. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef]
- Zhang, H.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Remans, T.; Nacry, P.; Pervent, M.; Filleur, S.; Diatloff, E.; Mounier, E.; Tillard, P.; Forde, B.G.; Gojon, A. The Arabidopsis NRT1.1 transporter participates in the signaling pathway triggering root colonization of nitrate-rich patches. Proc. Natl. Acad. Sci. USA 2006, 103, 19206–19211. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell. 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Fagard, M.; Launay, A.; Clément, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulié, M.C.; Masclaux-Daubresse, C. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef]
- Soulie, M.C.; Koka, S.M.; Floch, K.; Vancostenoble, B.; Barbe, D.; Daviere, A.; Soubigou-Taconnat, L.; Brunaud, V.; Poussereau, N.; Loisel, E.; et al. Plant nitrogen supply affects the Botrytis cinerea infection process and modulates known and novel virulence factors. Mol. Plant Pathol. 2020, 21, 1436–1450. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Simpson, C.; Kumari, A.; Gupta, A.K.; Gupta, K.J. Moving nitrogen to the centre of plant defence against pathogens. Ann. Bot. 2017, 119, 703–709. [Google Scholar] [CrossRef]
- Solomon, P.S.; Tan, K.C.; Oliver, R.P. The nutrient supply of pathogenic fungi; a fertile field for study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef]
- Huber, D.M.; Watson, R.D. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Richard-Molard, C.; Wuilleme, S.; Scheel, C.; Gresshoff, P.M.; Morot-Gaudry, J.F.; Limami, A.M. Nitrogen-induced changes in morphological development and bacterial susceptibility of belgian endive (Cichorium intybus L.) are genotype-dependent. Planta 1999, 209, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Camañes, G.; Pastor, V.; Cerezo, M.; García-Andrade, J.; Vicedo, B.; García-Agustín, P.; Flors, V. A deletion in NRT2.1 attenuates Pseudomonas syringae-induced hormonal perturbation, resulting in primed plant defenses. Plant Physiol. 2012, 158, 1054–1066. [Google Scholar] [CrossRef]
- Mur, L.A.; Sivakumaran, A.; Mandon, J.; Cristescu, S.M.; Harren, F.J.; Hebelstrup, K.H. Haemoglobin modulates salicylate and jasmonate/ethylene-mediated resistance mechanisms against pathogens. J. Exp. Bot. 2012, 63, 4375–4387. [Google Scholar] [CrossRef] [PubMed]
- Farjad, M.; Rigault, M.; Pateyron, S.; Martin-Magniette, M.L.; Krapp, A.; Meyer, C.; Fagard, M. Nitrogen Limitation Alters the Response of Specific Genes to Biotic Stress. Int. J. Mol. Sci. 2018, 19, 3364. [Google Scholar] [CrossRef] [PubMed]
- Farjad, M.; Clément, G.; Launay, A.; Jeridi, R.; Jolivet, S.; Citerne, S.; Rigault, M.; Soulie, M.C.; Dinant, S.; Fagard, M. Plant nitrate supply regulates Erwinia amylovora virulence gene expression in Arabidopsis. Mol. Plant Pathol. 2021, 22, 1332–1346. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Navarro, L.; Bari, R.; Jones, J.D. Pathological hormone imbalances. Curr. Opin. Plant Biol. 2007, 10, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Fagard, M.; Dellagi, A.; Roux, C.; Périno, C.; Rigault, M.; Boucher, V.; Shevchik, V.E.; Expert, D. Arabidopsis thaliana expresses multiple lines of defense to counterattack Erwinia chrysanthemi. Mol. PlantMicrobe Interact. 2007, 20, 794–805. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pathak, R.R.; Mandal, V.K.; Jangam, A.P.; Sharma, N.; Madan, B.; Jaiswal, D.K.; Raghuram, N. Heterotrimeric G-protein α subunit (RGA1) regulates tiller development, yield, cell wall, nitrogen response and biotic stress in rice. Sci. Rep. 2021, 11, 2323. [Google Scholar] [CrossRef]
- Camañes, G.; Pastor, V.; Cerezo, M.; García-Agustín, P.; Flors Herrero, V. A deletion in the nitrate high affinity transporter NRT2.1 alters metabolomic and transcriptomic responses to Pseudomonas syringae. Plant Signal. Behav. 2012, 7, 619–622. [Google Scholar] [CrossRef][Green Version]
- Hoffland, E.; Jeger, M.; vanBeusichem, M. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Barrit, T.; Porcher, A.; Cukier, C.; Satour, P.; Guillemette, T.; Limami, A.M.; Teulat, B.; Campion, C.; Planchet, E. Nitrogen nutrition modifies the susceptibility of Arabidopsis thaliana to the necrotrophic fungus, Alternaria brassicicola. Physiol. Plant 2022, 174, e13621. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Lin, S.H.; Hu, H.C.; Tsay, Y.F. CHL1 functions as a nitrate sensor in plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Edel, K.H.; Pervent, M.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Tillard, P.; Gojon, A.; Kudla, J.; Lacombe, B. Nitrate sensing and uptake in Arabidopsis are enhanced by ABI2, a phosphatase inactivated by the stress hormone abscisic acid. Sci. Signal. 2015, 8, ra43. [Google Scholar] [CrossRef] [PubMed]
- Weinl, S.; Kudla, J. The CBL-CIPK Ca(2+)-decoding signaling network: Function and perspectives. New Phytol. 2009, 184, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, V.; Ritz, O.; Linder, S.; Harter, K.; Kudla, J. The NAF domain defines a novel protein-protein interaction module conserved in Ca2+-regulated kinases. EMBO J. 2001, 20, 1051–1063. [Google Scholar] [CrossRef]
- Albrecht, V.; Weinl, S.; Blazevic, D.; D’Angelo, C.; Batistic, O.; Kolukisaoglu, U.; Bock, R.; Schulz, B.; Harter, K.; Kudla, J. The calcium sensor CBL1 integrates plant responses to abiotic stresses. Plant J. 2003, 36, 457–470. [Google Scholar] [CrossRef]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratgé-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.B.; et al. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta 2012, 1820, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Schachtman, D.P.; Shin, R. Nutrient sensing and signaling: NPKS. Annu. Rev. Plant Biol. 2007, 58, 47–69. [Google Scholar] [CrossRef] [PubMed]
- Peuke, A.D. ABA flow modelling in Ricinus communis exposed to salt stress and variable nutrition. J. Exp. Bot. 2016, 67, 5301–5311. [Google Scholar] [CrossRef]
- Signora, L.; De Smet, I.; Foyer, C.H.; Zhang, H. ABA plays a central role in mediating the regulatory effects of nitrate on root branching in Arabidopsis. Plant J. 2001, 28, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Ondzighi-Assoume, C.A.; Chakraborty, S.; Harris, J.M. Environmental Nitrate Stimulates Abscisic Acid Accumulation in Arabidopsis Root Tips by Releasing It from Inactive Stores. Plant Cell 2016, 28, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Joshi-Saha, A.; Valon, C.; Leung, J. A brand new START: Abscisic acid perception and transduction in the guard cell. Sci. Signal. 2011, 4, re4. [Google Scholar] [CrossRef]
- Joshi-Saha, A.; Valon, C.; Leung, J. Abscisic acid signal off the STARting block. Mol. Plant 2011, 4, 562–580. [Google Scholar] [CrossRef]
- Hashimoto, K.; Eckert, C.; Anschütz, U.; Scholz, M.; Held, K.; Waadt, R.; Reyer, A.; Hippler, M.; Becker, D.; Kudla, J. Phosphorylation of calcineurin B-like (CBL) calcium sensor proteins by their CBL-interacting protein kinases (CIPKs) is required for full activity of CBL-CIPK complexes toward their target proteins. J. Biol. Chem. 2012, 287, 7956–7968. [Google Scholar] [CrossRef]
- Harris, J.M.; Ondzighi-Assoume, C.A. Environmental nitrate signals through abscisic acid in the root tip. Plant Signal. Behav. 2017, 12, e1273303. [Google Scholar] [CrossRef]
- Harris, J.M. Abscisic Acid: Hidden Architect of Root System Structure. Plants 2015, 4, 548–572. [Google Scholar] [CrossRef] [PubMed]
- Véry, A.A.; Sentenac, H. Cation channels in the Arabidopsis plasma membrane. Trends Plant Sci. 2002, 7, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Lan, W.Z.; Kim, B.G.; Li, L.; Cheong, Y.H.; Pandey, G.K.; Lu, G.; Buchanan, B.B.; Luan, S. A protein phosphorylation/dephosphorylation network regulates a plant potassium channel. Proc. Natl. Acad. Sci. USA 2007, 104, 15959–15964. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kim, B.G.; Cheong, Y.H.; Pandey, G.K.; Luan, S. A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 12625–12630. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.H.; Pandey, G.K.; Grant, J.J.; Batistic, O.; Li, L.; Kim, B.G.; Lee, S.C.; Kudla, J.; Luan, S. Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. 2007, 52, 223–239. [Google Scholar] [CrossRef]
- Wang, X.P.; Chen, L.M.; Liu, W.X.; Shen, L.K.; Wang, F.L.; Zhou, Y.; Zhang, Z.; Wu, W.H.; Wang, Y. AtKC1 and CIPK23 Synergistically Modulate AKT1-Mediated Low-Potassium Stress Responses in Arabidopsis. Plant Physiol. 2016, 170, 2264–2277. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. Nitrogen transformations in modern agriculture and the role of biological nitrification inhibition. Nat. Plants 2017, 3, 17074. [Google Scholar] [CrossRef]
- Kellermeier, F.; Armengaud, P.; Seditas, T.J.; Danku, J.; Salt, D.E.; Amtmann, A. Analysis of the Root System Architecture of Arabidopsis Provides a Quantitative Readout of Crosstalk between Nutritional Signals. Plant Cell 2014, 26, 1480–1496. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Drechsler, N.; Zheng, Y.; Bohner, A.; Nobmann, B.; von Wirén, N.; Kunze, R.; Rausch, C. Nitrate-Dependent Control of Shoot K Homeostasis by the Nitrate Transporter1/Peptide Transporter Family Member NPF7.3/NRT1.5 and the Stelar K+ Outward Rectifier SKOR in Arabidopsis. Plant Physiol. 2015, 169, 2832–2847. [Google Scholar] [CrossRef]
- Raddatz, N.; Morales de Los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty Years from Transport to Signaling Networks. Plant Cell 2020, 32, 2094–2119. [Google Scholar] [CrossRef] [PubMed]
- Touraine, B.; Grignon, N.; Grignon, C. Charge Balance in NO3-Fed Soybean: Estimation of K and Carboxylate Recirculation. Plant Physiol. 1988, 88, 605–612. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Knight, A.H. Influence of the level of nitrate nutrition on ion uptake and assimilation, organic Acid accumulation, and cation-anion balance in whole tomato plants. Plant Physiol. 1977, 60, 349–353. [Google Scholar] [CrossRef]
- Engels, C.; Marschner, H. Adaptation of potassium translocation into the shoot of maize (Zea mays) to shoot demand: Evidence for xylem loading as a regulating step. Physiol. Plant. 1992, 86, 263–268. [Google Scholar] [CrossRef]
- Engels, C.; Marschner, H. Influence of the Form of Nitrogen Supply on Root Uptake and Translocation of Cations in the Xylem Exudate of Maize (Zea mays L.). J. Exp. Bot. 1993, 44, 1695–1701. [Google Scholar] [CrossRef]
- Troufflard, S.; Mullen, W.; Larson, T.R.; Graham, I.A.; Crozier, A.; Amtmann, A.; Armengaud, P. Potassium deficiency induces the biosynthesis of oxylipins and glucosinolates in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 172. [Google Scholar] [CrossRef]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef]
- Armengaud, P.; Breitling, R.; Amtmann, A. Coronatine-insensitive 1 (COI1) mediates transcriptional responses of Arabidopsis thaliana to external potassium supply. Mol. Plant 2010, 3, 390–405. [Google Scholar] [CrossRef]
- Delker, C.; Stenzel, I.; Hause, B.; Miersch, O.; Feussner, I.; Wasternack, C. Jasmonate biosynthesis in Arabidopsis thaliana--enzymes, products, regulation. Plant Biol. (Stuttg.) 2006, 8, 297–306. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Lyons, R. Intervention of Phytohormone Pathways by Pathogen Effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A.; Troufflard, S.; Armengaud, P. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant 2008, 133, 682–691. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).