
Hand-Foot-and-Mouth Disease-Associated Enterovirus and the Development of Multivalent HFMD Vaccines

Abstract
:1. Introduction
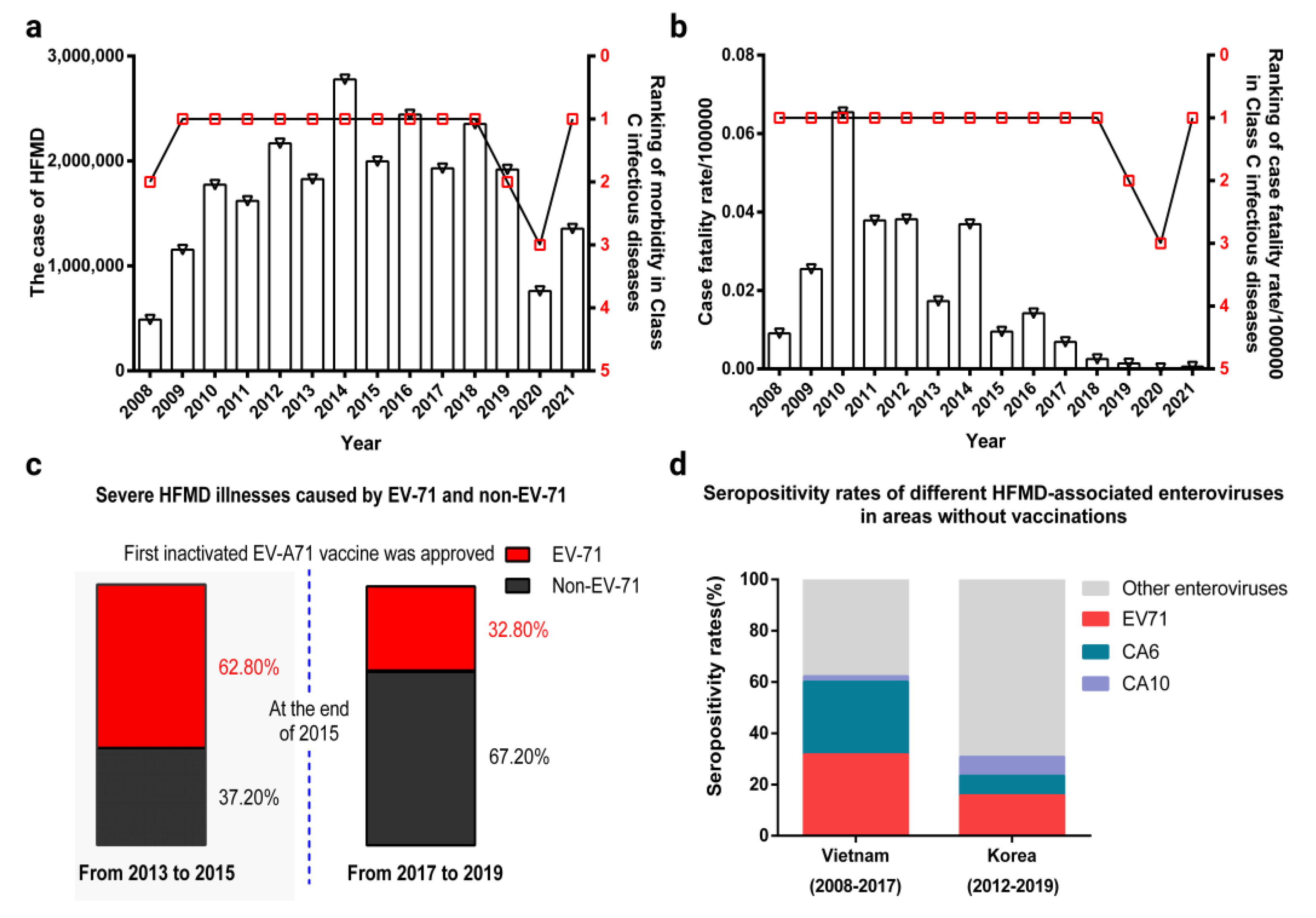
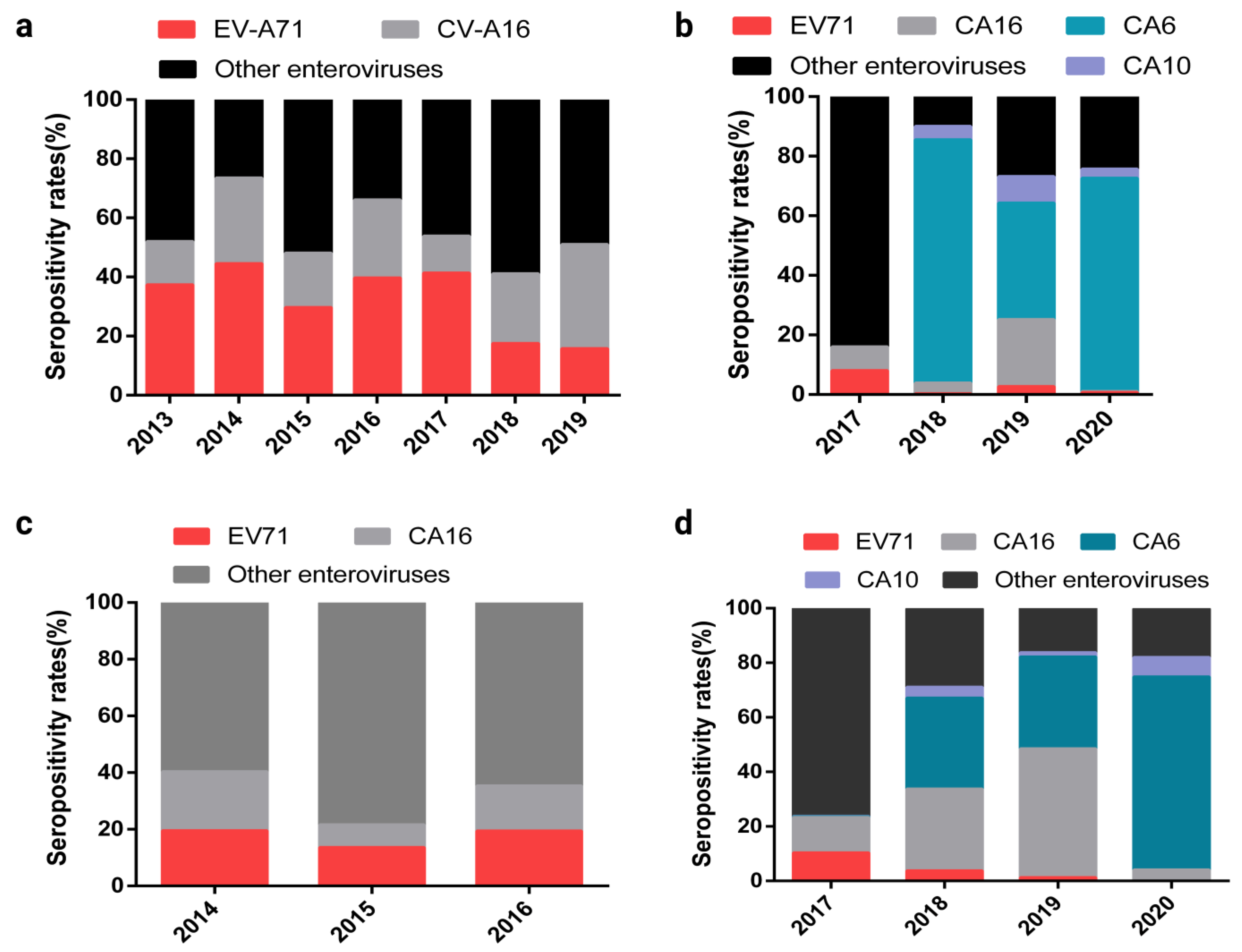
2. Changes in the Epidemiological Spectrum of HFMD-Associated Enteroviruses after the EV-A71 Inactivated Vaccine Was Approved for Marketing
3. Immunological Response to HFMD-Associated Enterovirus Infection
3.1. Native Immune Response Induced by a Viral Protein of HFMD-Associated Enterovirus
3.2. Adaptive Immune Response Induced by a Viral Protein of HFMD-Related Enterovirus
3.3. Immune Response to Dendritic Cells Infected with HFMD-Associated Enterovirus
4. The Immune Effect of the Experimental HFMD-Associated Enterovirus Multivalent Vaccine Reported in Recent Studies
4.1. EV-A71/CV-A16 Bivalent Vaccine
4.2. Bivalent CV-A6/CV-A10 Vaccine
4.3. EV-A71/CV-A6/CV-A10 Trivalent Vaccine
4.4. EV-A71/CV-A16/CV-A6/CV-A10 Tetravalent Vaccine
4.5. CVB Hexavalent Inactivated Vaccine
5. Discussion
5.1. Enhancing the Immune Protection of the HFMD Multivalent Vaccine by Inducing an Effective Innate Immune Response
5.2. Balancing Immunogenicity between Antigens to Reduce the Risk of Immune Interference
5.3. Inducing a Broad Spectrum of Protective Immune Response Coping Changes in the Epidemic Spectrum of HFMD-Associated Enterovirus
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ang, P.Y.; Chong, C.W.H.; Alonso, S. Viral determinants that drive Enterovirus-A71 fitness and virulence. Emerg. Microbes Infect. 2021, 10, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Koike, S. Adaptation and Virulence of Enterovirus-A71. Viruses 2021, 13, 1661. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Salas, E.; Francisco-Velilla, R.; Fernandez-Chamorro, J.; Lozano, G.; Diaz-Toledano, R. Picornavirus IRES elements: RNA structure and host protein interactions. Virus Res. 2015, 206, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Miragall, O.; de Quinto, S.L.; Martínez-Salas, E. Relevance of RNA structure for the activity of picornavirus IRES elements. Virus Res. 2009, 139, 172–182. [Google Scholar] [CrossRef]
- Jang, S.K.; Pestova, T.V.; Hellen, C.U.; Witherell, G.W.; Wimmer, E. Cap-independent translation of picornavirus RNAs: Structure and function of the internal ribosomal entry site. Enzyme 1990, 44, 292–309. [Google Scholar] [CrossRef]
- Francisco-Velilla, R.; Embarc-Buh, A.; Abellan, S.; Martinez-Salas, E. Picornavirus translation strategies. FEBS Open Bio 2022, 12, 1125–1141. [Google Scholar] [CrossRef]
- Aswathyraj, S.; Arunkumar, G.; Alidjinou, E.K.; Hober, D. Hand, foot and mouth disease (HFMD): Emerging epidemiology and the need for a vaccine strategy. Med. Microbiol. Immunol. 2016, 205, 397–407. [Google Scholar] [CrossRef]
- Bello, A.M.; Roshorm, Y.M. Recent progress and advances towards developing enterovirus 71 vaccines for effective protection against human hand, foot and mouth disease (HFMD). Biologicals 2022, 79, 1–9. [Google Scholar] [CrossRef]
- Esposito, S.; Principi, N. Hand, foot and mouth disease: Current knowledge on clinical manifestations, epidemiology, aetiology and prevention. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 391–398. [Google Scholar] [CrossRef]
- Légeret, C.; Furlano, R. Oral ulcers in children—A clinical narrative overview. Ital. J. Pediatr. 2021, 47, 144. [Google Scholar] [CrossRef]
- Nassef, C.; Ziemer, C.; Morrell, D.S. Hand-foot-and-mouth disease: A new look at a classic viral rash. Curr. Opin. Pediatr. 2015, 27, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Yi, E.-J.; Shin, Y.-J.; Kim, J.-H.; Kim, T.-G.; Chang, S.-Y. Enterovirus 71 infection and vaccines. Clin. Exp. Vaccine Res. 2017, 6, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Kinobe, R.; Wiyatno, A.; Artika, I.M.; Safari, D. Insight into the Enterovirus A71: A review. Rev. Med. Virol. 2022, 32, e2361. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Chong, P. Is a multivalent hand, foot, and mouth disease vaccine feasible? Hum. Vaccines Immunother. 2015, 11, 2688–2704. [Google Scholar] [CrossRef] [Green Version]
- Frydenberg, A.; Starr, M. Hand, foot and mouth disease. Aust. Fam. Physician 2003, 32, 594–595. [Google Scholar]
- Mirand, A.; Henquell, C.; Archimbaud, C.; Ughetto, S.; Antona, D.; Bailly, J.-L.; Peigue-Lafeuille, H. Outbreak of hand, foot and mouth disease/herpangina associated with coxsackievirus A6 and A10 infections in 2010, France: A large citywide, prospective observational study. Clin. Microbiol. Infect. 2012, 18, E110–E118. [Google Scholar] [CrossRef] [Green Version]
- Saguil, A.; Kane, S.; Lauters, R.; Mercado, M.G. Hand-Foot-and-Mouth Disease: Rapid Evidence Review. Am. Fam. Physician 2019, 100, 408–414. [Google Scholar]
- Nayak, G.; Bhuyan, S.K.; Bhuyan, R.; Sahu, A.; Kar, D.; Kuanar, A. Global emergence of Enterovirus 71: A systematic review. Beni Suef Univ. J. Basic Appl. Sci. 2022, 11, 78. [Google Scholar] [CrossRef]
- Alhazmi, A.; Nekoua, M.P.; Mercier, A.; Vergez, I.; Sane, F.; Alidjinou, E.K.; Hober, D. Combating coxsackievirus B infections. Rev. Med. Virol. 2022, e2406. [Google Scholar] [CrossRef]
- Di Prinzio, A.; Bastard, D.P.; Torre, A.C.; Mazzuoccolo, L.D. Hand, foot, and mouth disease in adults caused by Coxsackievirus B1-B6. An. Bras. Dermatol. 2022, 97, 321–325. [Google Scholar] [CrossRef]
- Sharma, V.; Goessling, L.S.; Brar, A.K.; Eghtesady, P. Multiple subtypes of coxsackievirus group B can cause congenital heart disease. Birth Defects Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jmii, H.; Fisson, S.; Aouni, M.; Jaidane, H. Type B coxsackieviruses and central nervous system disorders: Critical review of reported associations. Rev. Med. Virol. 2021, 31, e2191. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, L.; Zhang, C.; He, W.; Tan, Y.; Ning, C. The changes in the epidemiology of hand, foot, and mouth disease after the introduction of the EV-A71 vaccine. Vaccine 2021, 39, 3319–3323. [Google Scholar] [CrossRef] [PubMed]
- Hoa-Tran, T.N.; Dao AT, H.; Nguyen, A.T.; Kataoka, C.; Takemura, T.; Pham, C.H.; Vu, H.M.; Hong, T.T.T.; Ha, N.T.V.; Duong, T.N.; et al. Coxsackieviruses A6 and A16 associated with hand, foot, and mouth disease in Vietnam, 2008–2017: Essential information for rational vaccine design. Vaccine 2020, 38, 8273–8285. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Yoon, Y.; Lee, Y.-P.; Kim, H.-J.; Lee, D.-Y.; Lee, J.-W.; Hyeon, J.-Y.; Yoo, J.S.; Lee, S.; Kang, C.; et al. A Different Epidemiology of Enterovirus A and Enterovirus B Co-circulating in Korea, 2012–2019. J. Pediatric. Infect. Dis. Soc. 2021, 10, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Liu, F.; Qi, H.; Tu, W.; Ward, M.P.; Ren, M.; Zhao, Z.; Su, Q.; Huang, J.; Chen, X.; et al. Changing epidemiology of hand, foot, and mouth disease in China, 2013–2019: A population-based study. Lancet Reg. Health West. Pac. 2022, 20, 100370. [Google Scholar] [CrossRef]
- Duan, X.; Zhang, C.; Wang, X.; Ren, X.; Peng, H.; Tang, X.; Zhang, L.; Chen, Z.; Ye, Y.; Zheng, M.; et al. Molecular epidemiology and clinical features of hand, foot and mouth disease requiring hospitalization after the use of enterovirus A71 inactivated vaccine in chengdu, China, 2017–2022: A descriptive study. Emerg. Microbes Infect. 2022, 11, 2510–2519. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, Z.; Rao, Q.; Wang, X.; Wang, M.; Du, T.; Tang, J.; Long, S.; Zhang, J.; Luo, J.; et al. The epidemiological characteristics of enterovirus infection before and after the use of enterovirus 71 inactivated vaccine in Kunming, China. Emerg. Microbes Infect. 2021, 10, 619–628. [Google Scholar] [CrossRef]
- Wang, M.; Chen, T.; Peng, J.; Luo, Y.; Du, L.; Lu, Z.; He, J.; Liu, C.; Gan, Q.; Ma, W.; et al. The spatial-temporal distribution and etiological characteristics of hand-foot-and-mouth disease before and after EV-A71 vaccination in Kunming, China, 2017–2020. Sci. Rep. 2022, 12, 17028. [Google Scholar] [CrossRef]
- Doherty, P.C.; Turner, S.J. The virus-immunity ecosystem. Arch. Virol. Suppl. 2005, 17–32. [Google Scholar] [CrossRef]
- Baggen, J.; Thibaut, H.J.; Strating, J.; Van Kuppeveld, F.J.M. The life cycle of non-polio enteroviruses and how to target it. Nat. Rev. Microbiol. 2018, 16, 368–381. [Google Scholar] [CrossRef]
- Maarouf, M.; Rai, K.; Goraya, M.; Chen, J.-L. Immune Ecosystem of Virus-Infected Host Tissues. Int. J. Mol. Sci. 2018, 19, 1379. [Google Scholar] [CrossRef] [PubMed]
- Owino, C.; Chu, J.J.H. Recent advances on the role of host factors during non-poliovirus enteroviral infections. J. Biomed. Sci. 2019, 26, 47. [Google Scholar] [CrossRef] [Green Version]
- Thibaut, H.J.; De Palma, A.M.; Neyts, J. Combating enterovirus replication: State-of-the-art on antiviral research. Biochem. Pharmacol. 2012, 83, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Guy, C.; Bowie, A.G. Recent insights into innate immune nucleic acid sensing during viral infection. Curr. Opin. Immunol. 2022, 78, 102250. [Google Scholar] [CrossRef] [PubMed]
- Beachboard, D.C.; Horner, S.M. Innate immune evasion strategies of DNA and RNA viruses. Curr. Opin. Microbiol. 2016, 32, 113–119. [Google Scholar] [CrossRef]
- Damania, B.; Blackbourn, D.J. Innate barriers to viral infection. Future Microbiol. 2012, 7, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Aoshi, T.; Koyama, S.; Kobiyama, K.; Akira, S.; Ishii, K. Innate and adaptive immune responses to viral infection and vaccination. Curr. Opin. Virol. 2011, 1, 226–232. [Google Scholar] [CrossRef]
- Sadler, A.J.; Williams, B.R.G. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Aw-Yong, K.L.; NikNadia, N.M.N.; Tan, C.W.; Sam, I.; Chan, Y.F. Immune responses against enterovirus A71 infection: Implications for vaccine success. Rev. Med. Virol. 2019, 29, e2073. [Google Scholar] [CrossRef]
- Scutigliani, E.M.; Kikkert, M. Interaction of the innate immune system with positive-strand RNA virus replication organelles. Cytokine Growth Factor Rev. 2017, 37, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.; Bárcena, M. Multiscale Electron Microscopy for the Study of Viral Replication Organelles. Viruses 2021, 13, 197. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, O.H.; Svedin, E.; Kapell, S.; Nurminen, A.; Hytönen, V.P.; Flodström-Tullberg, M. Enteroviral proteases: Structure, host interactions and pathogenicity. Rev. Med. Virol. 2016, 26, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, A.; Motta, M.; Singh, S.; Kasniya, G.; Mane, S.S.; Cartaya, S.; Rahman, M.M.; Dudeja, P. Enteroviral Infections in Infants. Newborn 2022, 1, 297–305. [Google Scholar] [CrossRef]
- Tsu, B.V.; Fay, E.J.; Nguyen, K.T.; Corley, M.R.; Hosuru, B.; Dominguez, V.A.; Daugherty, M.D. Running With Scissors: Evolutionary Conflicts between Viral Proteases and the Host Immune System. Front. Immunol. 2021, 12, 769543. [Google Scholar] [CrossRef]
- Verburg, S.G.; Lelievre, R.M.; Westerveld, M.J.; Inkol, J.M.; Sun, Y.L.; Workenhe, S.T. Viral-mediated activation and inhibition of programmed cell death. PLoS Pathog. 2022, 18, e1010718. [Google Scholar] [CrossRef]
- Cox, J.A.; Hiscox, J.A.; Solomon, T.; Ooi, M.H.; Ng, L.F. Immunopathogenesis and Virus-Host Interactions of Enterovirus 71 in Patients with Hand, Foot and Mouth Disease. Front. Microbiol. 2017, 8, 2249. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; He, Y.; Chen, Y.; Kung, H.-F.; He, M.-L. Potent inhibition of human enterovirus 71 replication by type I interferon subtypes. Antivir. Ther. 2011, 16, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Park, N.; Schweers, N.J.; Gustin, K.E. Selective Removal of FG Repeat Domains from the Nuclear Pore Complex by Enterovirus 2A(pro). J. Virol. 2015, 89, 11069–11079. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Xi, X.; Lei, X.; Zhang, X.; Cui, S.; Wang, J.; Jin, Q.; Zhao, Z. Enterovirus 71 protease 2Apro targets MAVS to inhibit anti-viral type I interferon responses. PLoS Pathog. 2013, 9, e1003231. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.K.; Panda, S.; Sahu, B.P.; Sarangi, R. Activation of Host Cellular Signaling and Mechanism of Enterovirus 71 Viral Proteins Associated with Hand, Foot and Mouth Disease. Viruses 2022, 14, 2190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Li, H.; Zhang, Z.; Meng, J.; Mao, D.; Bai, B.; Lu, B.; Mao, P.; Hu, Q.; Wang, H. Enterovirus 71 2C protein inhibits TNF-α-mediated activation of NF-κB by suppressing IκB kinase β phosphorylation. J. Immunol. 2011, 187, 2202–2212. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Hou, H.; Wang, F.; Qiao, L.; Wang, X.; Yu, J.; Liu, W.; Sun, Z. ATP1B3: A virus-induced host factor against EV71 replication by up-regulating the production of type-I interferons. Virology 2016, 496, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.S.; Stobart, C.C.; Luo, H. Innate immune evasion mediated by picornaviral 3C protease: Possible lessons for coronaviral 3C-like protease? Rev. Med. Virol. 2021, 31, 1–22. [Google Scholar] [CrossRef]
- Wallace, H.L.; Russell, R.S. Promiscuous Inflammasomes: The False Dichotomy of RNA/DNA Virus-Induced Inflammasome Activation and Pyroptosis. Viruses 2022, 14, 2113. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xiao, F.; Wan, P.; Pan, P.; Zhang, Y.; Liu, F.; Wu, K.; Liu, Y.; Wu, J. Correction: EV71 3D Protein Binds with NLRP3 and Enhances the Assembly of Inflammasome Complex. PLoS Pathog. 2018, 14, e1006921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahand, J.S.; Salmaninejad, A.; Mollazadeh, S.; Zadeh, S.S.T.; Rezaee, M.; Sheida, A.H.; Sadoughi, F.; Dana, P.M.; Rafiyan, M.; Zamani, M.; et al. Virus, Exosome, and MicroRNA: New Insights into Autophagy. Adv. Exp. Med. Biol. 2022, 1401, 97–162. [Google Scholar]
- Sarry, M.; Vitour, D.; Zientara, S.; Kassimi, L.B.; Blaise-Boisseau, S. Foot-and-Mouth Disease Virus: Molecular Interplays with IFN Response and the Importance of the Model. Viruses 2022, 14, 2129. [Google Scholar] [CrossRef]
- Wang, S.-H.; Wang, A.; Liu, P.-P.; Zhang, W.-Y.; Du, J.; Xu, S.; Liu, G.-C.; Zheng, B.-S.; Huan, C.; Zhao, K.; et al. Divergent Pathogenic Properties of Circulating Coxsackievirus A6 Associated with Emerging Hand, Foot, and Mouth Disease. J. Virol. 2018, 92, e00303-18. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.; Su, J.; Wang, H.; Chang, J.; Wang, S.; Zheng, W.; Cai, Y.; Wei, W.; Gordy, J.T.; Markham, R.; et al. Disruption of MDA5-Mediated Innate Immune Responses by the 3C Proteins of Coxsackievirus A16, Coxsackievirus A6, and Enterovirus D68. J. Virol. 2017, 91, e00546-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yu, X.; Meng, X.; Huo, W.; Su, Y.; Liu, J.; Liu, Y.; Zhang, J.; Wang, S.; Yu, J. Coxsackievirus A6 Induces Necroptosis for Viral Production. Front. Microbiol. 2020, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.M.; Zhang, Y.L.; Bahreyni, A.; Luo, H.; Mohamud, Y. Coxsackievirus Protease 2A Targets Host Protease ATG4A to Impair Autophagy. Viruses 2022, 14, 2026. [Google Scholar] [CrossRef] [PubMed]
- Visser, L.J.; Langereis, M.A.; Rabouw, H.H.; Wahedi, M.; Muntjewerff, E.M.; de Groot, R.J.; van Kuppeveld, F.J.M. Essential Role of Enterovirus 2A Protease in Counteracting Stress Granule Formation and the Induction of Type I Interferon. J. Virol. 2019, 93, e00222-19. [Google Scholar] [CrossRef]
- Wessels, E.; Duijsings, D.; Lanke, K.H.W.; Melchers, W.J.G.; Jackson, C.; van Kuppeveld, F.J.M. Molecular determinants of the interaction between coxsackievirus protein 3A and guanine nucleotide exchange factor GBF1. J. Virol. 2007, 81, 5238–5245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Morosky, S.A.; Delorme-Axford, E.; Dybdahl-Sissoko, N.; Oberste, M.S.; Wang, T.; Coyne, C.B. The coxsackievirus B 3C protease cleaves MAVS and TRIF to attenuate host type I interferon and apoptotic signaling. PLoS Pathog. 2011, 7, e1001311. [Google Scholar] [CrossRef] [Green Version]
- Plevka, P.; Perera, R.; Cardosa, J.; Kuhn, R.J.; Rossmann, M.G. Crystal structure of human enterovirus 71. Science 2012, 336, 1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuente, J.O.; Lee, H.; Yoder, J.D.; Shingler, K.L.; Carnegie, M.S.; Yoder, J.L.; Ashley, R.E.; Makhov, A.M.; Conway, J.F.; Hafenstein, S. Structures of the procapsid and mature virion of enterovirus 71 strain 1095. J. Virol. 2013, 87, 7637–7645. [Google Scholar] [CrossRef] [Green Version]
- Thanongsaksrikul, J.; Srimanote, P.; Tongtawe, P.; Glab-Ampai, K.; Malik, A.A.; Supasorn, O.; Chiawwit, P.; Poovorawan, Y.; Chaicumpa, W. Identification and production of mouse scFv to specific epitope of enterovirus-71 virion protein-2 (VP2). Arch. Virol. 2018, 163, 1141–1152. [Google Scholar] [CrossRef]
- Anasir, M.I.; Poh, C.L. Advances in Antigenic Peptide-Based Vaccine and Neutralizing Antibodies against Viruses Causing Hand, Foot, and Mouth Disease. Int. J. Mol. Sci. 2019, 20, 1256. [Google Scholar] [CrossRef] [Green Version]
- Lui, Y.L.E.; Tan, T.L.; Woo, W.H.; Timms, P.; Hafner, L.M.; Tan, K.H.; Tan, E.L. Enterovirus71 (EV71) utilise host microRNAs to mediate host immune system enhancing survival during infection. PLoS ONE 2014, 9, e102997. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, L.; Wu, Z.; Tien, P. Differential interferon pathway gene expression patterns in Rhabdomyosarcoma cells during Enterovirus 71 or Coxsackievirus A16 infection. Biochem. Biophys. Res. Commun. 2014, 447, 550–555. [Google Scholar] [CrossRef]
- Pääbo, S.; Severinsson, L.; Andersson, M.; Martens, I.; Peterson, P.A.; Nilsson, T. Adenovirus proteins and MHC expression. Adv. Cancer Res. 1989, 52, 151–163. [Google Scholar] [PubMed]
- Marongiu, L.; Valache, M.; Facchini, F.A.; Granucci, F. How dendritic cells sense and respond to viral infections. Clin. Sci. Lond. 2021, 135, 2217–2242. [Google Scholar] [CrossRef] [PubMed]
- Vinjamuri, S.; Li, L.; Bouvier, M. SARS-CoV-2 ORF8: One protein, seemingly one structure, and many functions. Front. Immunol. 2022, 13, 1035559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Li, Y.; Huang, F.; Luo, B.; Yuan, Y.; Xia, B.; Ma, X.; Yang, T.; Yu, F.; et al. The ORF8 protein of SARS-CoV-2 mediates immune evasion through down-regulating MHC-I. Proc. Natl. Acad. Sci. USA 2021, 118, e2024202118. [Google Scholar] [CrossRef]
- Natarajan, V.; Simoneau, C.R.; Erickson, A.L.; Meyers, N.L.; Baron, J.L.; Cooper, S.; McDevitt, T.C.; Ott, M. Modelling T-cell immunity against hepatitis C virus with liver organoids in a microfluidic coculture system. Open Biol. 2022, 12, 210320. [Google Scholar] [CrossRef]
- Kang, W.; Sung, P.S.; Park, S.-H.; Yoon, S.; Chang, D.-Y.; Kim, S.; Han, K.H.; Kim, J.K.; Rehermann, B.; Chwae, Y.-J.; et al. Hepatitis C virus attenuates interferon-induced major histocompatibility complex class I expression and decreases CD8+ T cell effector functions. Gastroenterology 2014, 146, 1351–1360.e4. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Ou, J.-H.J. Hepatitis C virus and intracellular antiviral response. Curr. Opin. Virol. 2022, 52, 244–249. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Zhang, X.; Hu, Y.; Dong, C.; Liu, L.; Yang, E.; Che, Y.; Pu, J.; Wang, X.; et al. Pathologic and immunologic characteristics of coxsackievirus A16 infection in rhesus macaques. Virology 2017, 500, 198–208. [Google Scholar] [CrossRef]
- Fan, S.; Liao, Y.; Lian, Y.; Jiang, G.; Jiang, L.; Dong, C.; Yang, E.; Wang, L.; Xu, X.; Feng, M.; et al. Erratum: Author Correction: Role of innate lymphoid cells and dendritic cells in intradermal immunization of the enterovirus antigen. NPJ Vaccines 2019, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Liao, Y.; Jiang, G.; Jiang, L.; Wang, L.; Xu, X.; Feng, M.; Yang, E.; Zhang, Y.; Cui, W.; et al. Study of integrated protective immunity induced in rhesus macaques by the intradermal administration of a bivalent EV71-CA16 inactivated vaccine. Vaccine 2020, 38, 2034–2044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Dong, Z.; Wang, Q.; Carr, M.J.; Li, J.; Liu, T.; Li, D.; Shi, W. Characterization of an inactivated whole-virus bivalent vaccine that induces balanced protective immunity against coxsackievirus A6 and A10 in mice. Vaccine 2018, 36, 7095–7104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Chou, C.-T.; Lei, H.-Y.; Liu, C.-C.; Wang, S.-M.; Yan, J.-J.; Su, I.-J.; Wang, J.-R.; Yeh, T.-M.; Chen, S.-H.; et al. A mouse-adapted enterovirus 71 strain causes neurological disease in mice after oral infection. J. Virol. 2004, 78, 7916–7924. [Google Scholar] [CrossRef] [PubMed]
- Caine, E.A.; Fuchs, J.; Das, S.C.; Partidos, C.D.; Osorio, J.E. Efficacy of a Trivalent Hand, Foot, and Mouth Disease Vaccine against Enterovirus 71 and Coxsackieviruses A16 and A6 in Mice. Viruses 2015, 7, 5919–5932. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Dai, W.; Zhang, C.; Zhou, Y.; Xiong, P.; Wang, S.; Ye, X.; Liu, Q.; Zhou, D.; Huang, Z. A virus-like particle-based tetravalent vaccine for hand, foot, and mouth disease elicits broad and balanced protective immunity. Emerg. Microbes Infect. 2018, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Yu, C.-I.; Hu, Y.-C.; Tsai, T.-J.; Kuo, Y.-C.; Chi, W.-K.; Lin, A.-N.; Chiang, B.-L. Enterovirus type 71 neutralizing antibodies in the serum of macaque monkeys immunized with EV71 virus-like particles. Vaccine 2012, 30, 1305–1312. [Google Scholar] [CrossRef]
- Stone, V.M.; Hankaniemi, M.M.; Laitinen, O.H.; Sioofy-Khojine, A.B.; Lin, A.; Lozano, I.M.D.; Mazur, M.A.; Marjomäki, V.; Loré, K.; Hyöty, H.; et al. A hexavalent Coxsackievirus B vaccine is highly immunogenic and has a strong protective capacity in mice and nonhuman primates. Sci. Adv. 2020, 6, eaaz2433. [Google Scholar] [CrossRef]
- Stone, V.M.; Hankaniemi, M.M.; Svedin, E.; Sioofy-Khojine, A.; Oikarinen, S.; Hyöty, H.; Laitinen, O.H.; Hytönen, V.P.; Flodström-Tullberg, M. A Coxsackievirus B vaccine protects against virus-induced diabetes in an experimental mouse model of type 1 diabetes. Diabetologia 2018, 61, 476–481. [Google Scholar] [CrossRef] [Green Version]
- Ghazvini, K.; Keikha, M. Multivalent vaccines against new SARS-CoV-2 hybrid variants. Vacunas 2022. [Google Scholar] [CrossRef]
- Liu, C.-C.; Guo, M.-S.; Wu, S.-R.; Lin, H.-Y.; Yang, Y.-T.; Liu, W.-C.; Chow, Y.-H.; Shieh, D.-B.; Wang, J.-R.; Chong, P. Immunological and biochemical characterizations of coxsackievirus A6 and A10 viral particles. Antivir. Res. 2016, 129, 58–66. [Google Scholar] [CrossRef]
- Dickey, T.H.; Tang, W.K.; Butler, B.; Ouahes, T.; Orr-Gonzalez, S.; Salinas, N.D.; Lambert, L.E.; Tolia, N.H. Design of the SARS-CoV-2 RBD vaccine antigen improves neutralizing antibody response. Sci. Adv. 2022, 8, eabq8276. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Chou, A.-H.; Lien, S.-P.; Lin, H.-Y.; Liu, S.-J.; Chang, J.-Y.; Guo, M.-S.; Chow, Y.-H.; Yang, W.-S.; Chang, K.H.-W.; et al. Identification and characterization of a cross-neutralization epitope of Enterovirus 71. Vaccine 2011, 29, 4362–4372. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Functions | Innate Immune Response | Re | |
|---|---|---|---|---|
| EV-71 | 2A | Cleaves NLRP3 protein Cleaves IFN-α/β receptor 1(IFNAR1) Cleaves the nuclear pore glycoprotein protein 62 (Nup62) Slices mitochondrial antiviral signaling (MAVS) protein Reduces serine phosphorylation of signal transducers and activators of transcription 1 (STAT1) | Inhibits inflammasome activation Blocks IFN-induced Jak/STAT signaling Disrupts host nuclear transport pathways and alters nuclear permeability Inactivates the antiviral innate immune response of RIG-I Attenuates IFN-γ signaling | [46] [47] [48] [49] [50] |
| 2C | Interacts with the IPT domain of RelA(p65) Inhibits IKKβ activation | Reduces the formation of the predominant form of NF-κB(heterodimer p65/p50) Blocks NF-κB activation | [51] [52] | |
| 3A | Interacts with the human β3 subunit of Na+/K+-ATPase (ATP1B3) protein, | Enhances the production of type-I IFN | [53] | |
| 3C | Cleaves the TAK1/TAB1/TAB2/TAB3 Complex Cleaves gasdermin D and NLRP3 | Suppresses cytokine expression Inhibits cell pyroptosis and NLRP3 inflammasome activation | [54] [55] | |
| 3D | Binds to NLRP3 Decreases STAT1 expression | Activates NLRP3 inflammasome Attenuates IFN-γ signaling | [56] [32] | |
| CA16 | 2C | Blocks the fusion of autophagosomes with lysosomes and triggers autophagosome accumulation | Induces autophagy | [57] |
| 3C | Binds to MDA5 and inhibits its interaction with MAVS | Blocks MDA5-triggered type I IFN induction | [58] | |
| CA6 | 2C | Blocks the fusion of autophagosomes with lysosomes and triggers autophagosome accumulation | Induces autophagy | [59] |
| 3C | Binds to MDA5 and inhibits its interaction with MAVS | Blocks MDA5-triggered type I IFN induction | [60] | |
| 3D | Binds to RIPK3 | Induces RIPK3-dependent necroptosis | [61] | |
| CB3 | 2A | Cleaves host protease ATG4A | Impairs autophagy | [62,63] |
| 3A | Binds to GBF1 | interference with Arf1-mediated COP-I recruitment | [64] | |
| 3C | Cleaves MAVS and TRIF | inhibits both the type I IFN and apoptotic signaling | [65] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, Y.; Li, H.; Liu, L. Hand-Foot-and-Mouth Disease-Associated Enterovirus and the Development of Multivalent HFMD Vaccines. Int. J. Mol. Sci. 2023, 24, 169. https://doi.org/10.3390/ijms24010169
Zhang X, Zhang Y, Li H, Liu L. Hand-Foot-and-Mouth Disease-Associated Enterovirus and the Development of Multivalent HFMD Vaccines. International Journal of Molecular Sciences. 2023; 24(1):169. https://doi.org/10.3390/ijms24010169
Chicago/Turabian StyleZhang, Xinglong, Yifan Zhang, Heng Li, and Longding Liu. 2023. "Hand-Foot-and-Mouth Disease-Associated Enterovirus and the Development of Multivalent HFMD Vaccines" International Journal of Molecular Sciences 24, no. 1: 169. https://doi.org/10.3390/ijms24010169
APA StyleZhang, X., Zhang, Y., Li, H., & Liu, L. (2023). Hand-Foot-and-Mouth Disease-Associated Enterovirus and the Development of Multivalent HFMD Vaccines. International Journal of Molecular Sciences, 24(1), 169. https://doi.org/10.3390/ijms24010169