
AEC and AFMSC Transplantation Preserves Fertility of Experimentally Induced Rat Varicocele by Expressing Differential Regenerative Mechanisms

 ,
,  ,
,  ,
, 
 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Influence of VAR Treatment with hAECs and hAFMSCs on Rat Fertility Rate
2.2. Testicular Morphology in VAR Rat Model after hAECs and hAFMSCs Transplantation
2.3. Effect of hAECs and hAFMSCs Transplantation on VAR Testes ECS Gene Expression
2.4. Inflammatory Asset in Testes of VAR Rats Treated without or with hAECs and hAFMSCs
2.5. hAECs and hAFMSCs Engraftment and Their Contribution in the Host VAR Tissues
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Isolation and Characterization Phenotype of Human Amniotic Membrane Epithelial Cells (hAECs)
4.3. Isolation, Culture, and Characterization of Human Mesenchymal Stromal Cells from Amniotic Fluid (hAFMSCs)
4.4. hAECs and hAFMSCs Stain with the Red Fluorescent Cell Linker PKH26
4.5. Animals
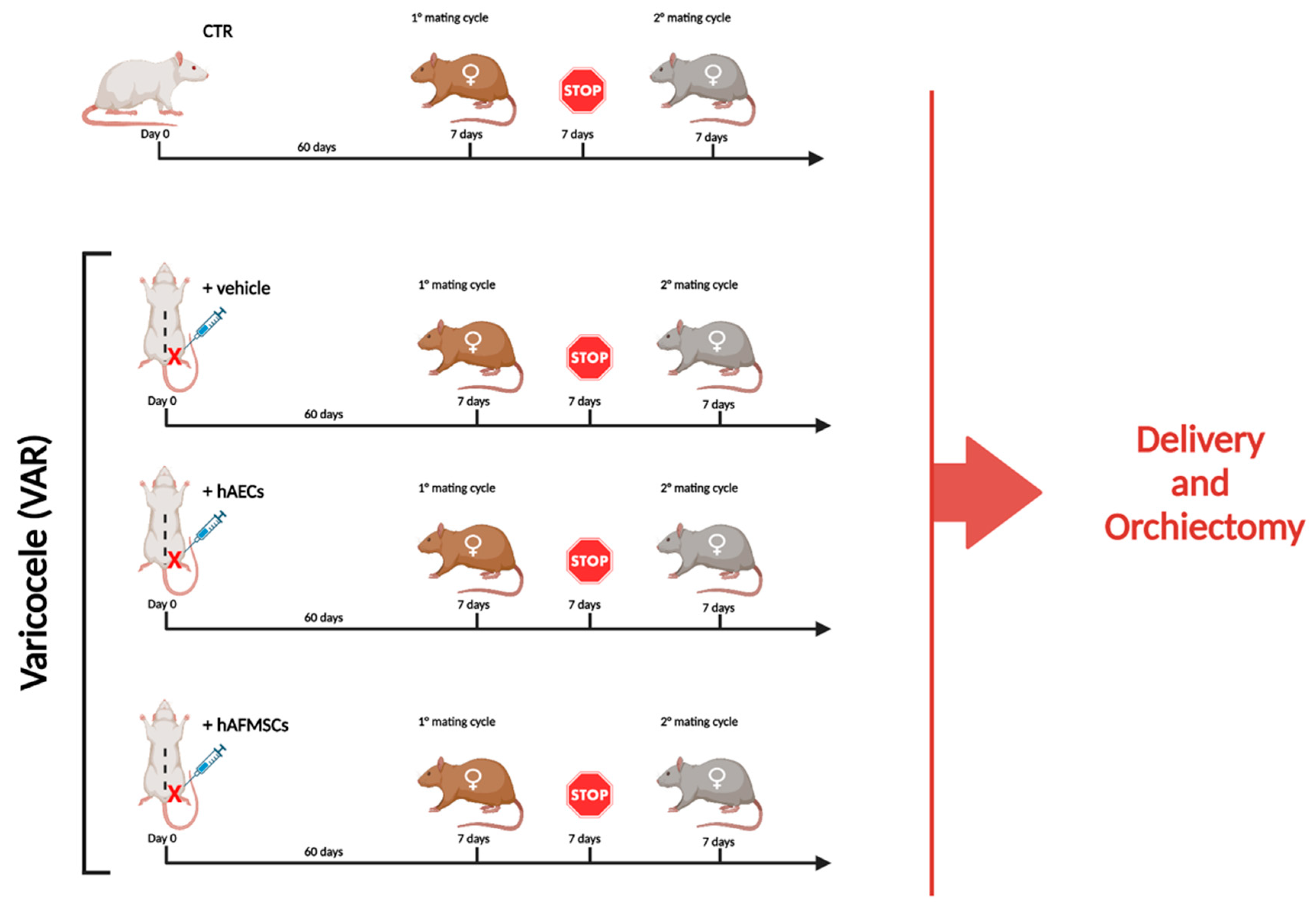
4.6. VAR Surgery Induction and Animal Groups Treatments
4.7. Histology and Immunohistochemistry Analyses
4.7.1. Haematoxylin-Eosin Staining
4.7.2. Immunohistochemistry (IHC)
4.7.3. Detection of hAECs and hAFMSCs in the Engrafted Testes
4.8. FISH Analysis for Human Species-Specific COT1 DNA Probe
4.9. Genomic DNA Extraction and PCR Amplification of Human MT-CYB Gene
4.10. Quantitative Real Time PCR
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sousa, A.P.; Santos-Pereira, J.; Freire, M.J.; Parada, B.; Almeida-Santos, T.; Bernardino, J.; Ramalho-Santos, J. Using Data Mining to Assist in Predicting Reproductive Outcomes Following Varicocele Embolization. J. Clin. Med. 2021, 10, 3503. [Google Scholar] [CrossRef] [PubMed]
- Bellastella, G.; Carotenuto, R.; Caiazzo, F.; Longo, M.; Cirillo, P.; Scappaticcio, L.; Carbone, C.; Arcaniolo, D.; Maiorino, M.I.; Esposito, K. Varicocele: An Endocrinological Perspective. Front. Reprod. Health 2022, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Perruzza, D.; Bernabò, N.; Rapino, C.; Valbonetti, L.; Falanga, I.; Russo, V.; Mauro, A.; Berardinelli, P.; Stuppia, L.; Maccarrone, M.; et al. Artificial Neural Network to Predict Varicocele Impact on Male Fertility through Testicular Endocannabinoid Gene Expression Profiles. Biomed. Res. Int. 2018, 2018, 3591086. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Punjani, N.; Lee, R.K.; Li, P.S.; Goldstein, M. Effect of Varicoceles on Spermatogenesis. Semin. Cell Dev. Biol. 2022, 121, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Bettella, A.; Rossato, M.; La Sala, G.; De Paoli, M.; Plebani, M. Inhibin B Plasma Concentrations in Oligozoospermic Subjects before and after Therapy with Follicle Stimulating Hormone. Hum. Reprod. 1999, 14, 906–912. [Google Scholar] [CrossRef]
- Pallotti, F.; Paoli, D.; Carlini, T.; Vestri, A.R.; Martino, G.; Lenzi, A.; Lombardo, F. Varicocele and Semen Quality: A Retrospective Case-Control Study of 4230 Patients from a Single Centre. J. Endocrinol. Investig. 2018, 41, 185–192. [Google Scholar] [CrossRef]
- Gomaa, M.D.; Motawaa, M.A.; Al-Nashar, A.M.; El-Sakka, A.I. Impact of Subinguinal Varicocelectomy on Serum Testosterone to Estradiol Ratio in Male Patients with Infertility. Urology 2018, 117, 70–77. [Google Scholar] [CrossRef]
- Barchi, M.; Innocenzi, E.; Giannattasio, T.; Dolci, S.; Rossi, P.; Grimaldi, P. Cannabinoid Receptors Signaling in the Development, Epigenetics, and Tumours of Male Germ Cells. Int. J. Mol. Sci. 2019, 21, 25. [Google Scholar] [CrossRef]
- Takeyama, M.; Honjoh, M.; Kodama, M.; Sakaguchi, H.; Koh, E.; Kondoh, N.; Fujioka, H.; Nakamura, M.; Namiki, M.; Okuyama, A.; et al. Testicular Steroids in Spermatic and Peripheral Veins after Single Injection of HCG in Patients with Varicocele. Arch. Androl. 1990, 24, 207–213. [Google Scholar] [CrossRef]
- Santi, D.; Crépieux, P.; Reiter, E.; Spaggiari, G.; Brigante, G.; Casarini, L.; Rochira, V.; Simoni, M. Follicle-Stimulating Hormone (FSH) Action on Spermatogenesis: A Focus on Physiological and Therapeutic Roles. J. Clin. Med. 2020, 9, 1014. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Su, Y.; Xu, J.; Hu, Z.; Zhao, K.; Liu, C.; Zhang, H. Varicocele-Mediated Male Infertility: From the Perspective of Testicular Immunity and Inflammation. Front. Immunol. 2021, 12, 729539. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, I. The Pathophysiological Role of Inflammatory Parameters in Patients with Varicocele. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 6572–6577. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, A.; Hedger, M.P. Immunological, Paracrine and Endocrine Aspects of Testicular Immune Privilege. Mol. Cell. Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Indumathy, S.; Pueschl, D.; Klein, B.; Fietz, D.; Bergmann, M.; Schuppe, H.C.; Da Silva, N.; Loveland, B.E.; Hickey, M.J.; Hedger, M.P.; et al. Testicular Immune Cell Populations and Macrophage Polarisation in Adult Male Mice and the Influence of Altered Activin A Levels. J. Reprod. Immunol. 2020, 142, 103204. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Thompson, L.A.; Dufour, J.M. Sertoli Cells—Immunological Sentinels of Spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Washburn, R.L.; Hibler, T.; Kaur, G.; Dufour, J.M. Sertoli Cell Immune Regulation: A Double-Edged Sword. Front. Immunol. 2022, 13, 913502. [Google Scholar] [CrossRef]
- Bhushan, S.; Theas, M.S.; Guazzone, V.A.; Jacobo, P.; Wang, M.; Fijak, M.; Meinhardt, A.; Lustig, L. Immune Cell Subtypes and Their Function in the Testis. Front. Immunol. 2020, 11, 583304. [Google Scholar] [CrossRef]
- Lysiak, J.J. The Role of Tumor Necrosis Factor-Alpha and Interleukin-1 in the Mammalian Testis and Their Involvement in Testicular Torsion and Autoimmune Orchitis. Reprod. Biol. Endocrinol. 2004, 2, 9. [Google Scholar] [CrossRef]
- Mazhari, S.; Razi, M.; Sadrkhanlou, R. Silymarin and Celecoxib Ameliorate Experimental Varicocele-Induced Pathogenesis: Evidences for Oxidative Stress and Inflammation Inhibition. Int. Urol. Nephrol. 2018, 50, 1039–1052. [Google Scholar] [CrossRef]
- Battista, N.; Meccariello, R.; Cobellis, G.; Fasano, S.; Di Tommaso, M.; Pirazzi, V.; Konje, J.C.; Pierantoni, R.; Maccarrone, M. The Role of Endocannabinoids in Gonadal Function and Fertility along the Evolutionary Axis. Mol. Cell. Endocrinol. 2012, 355, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids and Reproductive Events in Health and Disease. Handb. Exp. Pharmacol. 2015, 231, 341–365. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Barboni, B.; Paradisi, A.; Bernabò, N.N.; Gasperi, V.; Pistilli, M.G.; Fezza, F.; Lucidi, P.; Mattioli, M. Characterization of the Endocannabinoid System in Boar Spermatozoa and Implications for Sperm Capacitation and Acrosome Reaction. J. Cell Sci. 2005, 118, 4393–4404. [Google Scholar] [CrossRef]
- Bernabò, N.; Barboni, B.; Maccarrone, M. The Biological Networks in Studying Cell Signal Transduction Complexity: The Examples of Sperm Capacitation and of Endocannabinoid System. Comput. Struct. Biotechnol. J. 2014, 11, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ramal-Sanchez, M.; Bernabò, N.; Valbonetti, L.; Cimini, C.; Taraschi, A.; Capacchietti, G.; Machado-Simoes, J.; Barboni, B. Role and Modulation of TRPV1 in Mammalian Spermatozoa: An Updated Review. Int. J. Mol. Sci. 2021, 22, 4306. [Google Scholar] [CrossRef]
- Salahshouri, S.; Akbarian, F.; Tavalaee, M.; Seifati, S.M.; Nasr-Esfahani, M.H. Expression of TRPV1 as A Heat Sensitive Voltage-Dependent Ion Channel and Oxidative Stress in Sperm Samples of Infertile Men with Varicocele: A Case-Control Study. Cell J. 2022, 24, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.M.; Rapino, C.; Di Tommaso, M.; Pucci, M.; Battista, N.; Paro, R.; Simon, L.; Lutton, D.; Maccarrone, M. Differences in the Endocannabinoid System of Sperm from Fertile and Infertile Men. PLoS ONE 2012, 7, e47704. [Google Scholar] [CrossRef]
- Park, S.G.; Yon, J.M.; Lin, C.; Gwon, L.W.; Lee, J.G.; Baek, I.J.; Lee, B.J.; Yun, Y.W.; Nam, S.Y. Capsaicin Attenuates Spermatogenic Cell Death Induced by Scrotal Hyperthermia through Its Antioxidative and Anti-Apoptotic Activities. Andrologia 2017, 49, e12656. [Google Scholar] [CrossRef]
- Saha, S.; Roy, P.; Corbitt, C.; Kakar, S.S. Application of Stem Cell Therapy for Infertility. Cells 2021, 10, 1613. [Google Scholar] [CrossRef]
- Siregar, S.; Noegroho, B.S.; Adriansjah, R.; Mustafa, A.; Bonar, A. The Effect of Intratesticular Injection of Human Adipose-Derived Mesenchymal Cell on Testicular Oxidative Stress and Spermatogenesis Process in the Varicocele Rat Model. Res. Rep. Urol. 2021, 13, 759–765. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Berardinelli, P.; Mauro, A.; Valbonetti, L.; Sanyal, H.; Canciello, A.; Greco, L.; Muttini, A.; Gatta, V.; et al. Placental Stem Cells from Domestic Animals: Translational Potential and Clinical Relevance. Cell Transplant. 2018, 27, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Miki, T. Stem Cell Characteristics and the Therapeutic Potential of Amniotic Epithelial Cells. Am. J. Reprod. Immunol. 2018, 80, e13003. [Google Scholar] [CrossRef] [PubMed]
- Pianta, S.; Magatti, M.; Vertua, E.; Bonassi Signoroni, P.; Muradore, I.; Nuzzo, A.M.; Rolfo, A.; Silini, A.; Quaglia, F.; Todros, T.; et al. Amniotic Mesenchymal Cells from Pre-Eclamptic Placentae Maintain Immunomodulatory Features as Healthy Controls. J. Cell. Mol. Med. 2016, 20, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Soncini, M.; Evangelista, M.; Schmidt, D. Amniotic Membrane and Amniotic Fluid-Derived Cells: Potential Tools for Regenerative Medicine? Regen. Med. 2009, 4, 275–291. [Google Scholar] [CrossRef]
- Insausti, C.L.; Blanquer, M.; García-Hernández, A.M.; Castellanos, G.; Moraleda, J.M. Amniotic Membrane-Derived Stem Cells: Immunomodulatory Properties and Potential Clinical Application. Stem Cells Cloning 2014, 7, 53–63. [Google Scholar] [CrossRef]
- Naeem, A.; Gupta, N.; Naeem, U.; Elrayess, M.A.; Albanese, C. Amniotic Stem Cells as a Source of Regenerative Medicine to Treat Female Infertility. Hum. Cell 2023, 36, 15–25. [Google Scholar] [CrossRef]
- Turner, T.T. The Study of Varicocele through the Use of Animal Models. Hum. Reprod. Update 2001, 7, 78–84. [Google Scholar] [CrossRef]
- Rapino, C.; Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids as Biomarkers of Human Reproduction. Hum. Reprod. Update 2014, 20, 501–516. [Google Scholar] [CrossRef]
- Sayed, T.S.; Balasinor, N.H.; Nishi, K. Diverse Role of Endocannabinoid System in Mammalian Male Reproduction. Life Sci. 2021, 286, 120035. [Google Scholar] [CrossRef]
- Grimaldi, P.; Di Giacomo, D.; Geremia, R. The Endocannabinoid System and Spermatogenesis. Front. Endocrinol. 2013, 4, 192. [Google Scholar] [CrossRef]
- Francavilla, F.; Battista, N.; Barbonetti, A.; Vassallo, M.R.C.; Rapino, C.; Antonangelo, C.; Pasquariello, N.; Catanzaro, G.; Barboni, B.; Maccarrone, M. Characterization of the Endocannabinoid System in Human Spermatozoa and Involvement of Transient Receptor Potential Vanilloid 1 Receptor in Their Fertilizing Ability. Endocrinology 2009, 150, 4692–4700. [Google Scholar] [CrossRef] [PubMed]
- Bovolin, P.; Cottone, E.; Pomatto, V.; Fasano, S.; Pierantoni, R.; Cobellis, G.; Meccariello, R. Endocannabinoids Are Involved in Male Vertebrate Reproduction: Regulatory Mechanisms at Central and Gonadal Level. Front. Endocrinol. 2014, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, P.; Orlando, P.; Di Siena, S.; Lolicato, F.; Petrosino, S.; Bisogno, T.; Geremia, R.; De Petrocellis, L.; Di Marzo, V. The Endocannabinoid System and Pivotal Role of the CB2 Receptor in Mouse Spermatogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 11131–11136. [Google Scholar] [CrossRef] [PubMed]
- Cobellis, G.; Ricci, G.; Cacciola, G.; Orlando, P.; Petrosino, S.; Cascio, M.G.; Bisogno, T.; De Petrocellis, L.; Chioccarelli, T.; Altucci, L.; et al. A Gradient of 2-Arachidonoylglycerol Regulates Mouse Epididymal Sperm Cell Start-Up. Biol. Reprod. 2010, 82, 451–458. [Google Scholar] [CrossRef]
- Klein, T.W.; Newton, C.; Larsen, K.; Chou, J.; Perkins, I.; Lu, L.; Nong, L.; Friedman, H. Cannabinoid Receptors and T Helper Cells. J. Neuroimmunol. 2004, 147, 91–94. [Google Scholar] [CrossRef]
- Tanasescu, R.; Constantinescu, C.S. Cannabinoids and the Immune System: An Overview. Immunobiology 2010, 215, 588–597. [Google Scholar] [CrossRef]
- Grabner, G.F.; Eichmann, T.O.; Wagner, B.; Gao, Y.; Farzi, A.; Taschler, U.; Radner, F.P.W.; Schweiger, M.; Lass, A.; Holzer, P.; et al. Deletion of Monoglyceride Lipase in Astrocytes Attenuates Lipopolysaccharide-Induced Neuroinflammation. J. Biol. Chem. 2016, 291, 913–923. [Google Scholar] [CrossRef]
- Kozak, K.R.; Marnett, L.J. Oxidative Metabolism of Endocannabinoids. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 211–220. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Marnett, L.J. Non-Redundant Functions of Cyclooxygenases: Oxygenation of Endocannabinoids. J. Biol. Chem. 2008, 283, 8065–8069. [Google Scholar] [CrossRef]
- Piro, J.R.; Suidan, G.L.; Quan, J.; Pi, Y.Q.; O’Neill, S.M.; Ilardi, M.; Pozdnyakov, N.; Lanz, T.A.; Xi, H.; Bell, R.D.; et al. Inhibition of 2-AG Hydrolysis Differentially Regulates Blood Brain Barrier Permeability after Injury. J. Neuroinflamm. 2018, 15, 142. [Google Scholar] [CrossRef]
- Kohnz, R.A.; Nomura, D.K. Chemical Approaches to Therapeutically Target the Metabolism and Signaling of the Endocannabinoid 2-AG and Eicosanoids. Chem. Soc. Rev. 2014, 43, 6859–6869. [Google Scholar] [CrossRef] [PubMed]
- Barboni, B.; Russo, V.; Gatta, V.; Bernabò, N.; Berardinelli, P.; Mauro, A.; Martelli, A.; Valbonetti, L.; Muttini, A.; Di Giacinto, O.; et al. Therapeutic Potential of HAECs for Early Achilles Tendon Defect Repair through Regeneration. J. Tissue Eng. Regen. Med. 2018, 12, e1594–e1608. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, A.; Wang, M.; Schulz, C.; Bhushan, S. Microenvironmental Signals Govern the Cellular Identity of Testicular Macrophages. J. Leukoc. Biol. 2018, 104, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Winnall, W.R.; Muir, J.A.; Hedger, M.P. Rat Resident Testicular Macrophages Have an Alternatively Activated Phenotype and Constitutively Produce Interleukin-10 in Vitro. J. Leukoc. Biol. 2011, 90, 133–143. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Manuelpillai, U.; Lourensz, D.; Vaghjiani, V.; Tchongue, J.; Lacey, D.; Tee, J.Y.; Murthi, P.; Chan, J.; Hodge, A.; Sievert, W. Human Amniotic Epithelial Cell Transplantation Induces Markers of Alternative Macrophage Activation and Reduces Established Hepatic Fibrosis. PLoS ONE 2012, 7, e38631. [Google Scholar] [CrossRef]
- Magatti, M.; Vertua, E.; Cargnoni, A.; Silini, A.; Parolini, O. The Immunomodulatory Properties of Amniotic Cells: The Two Sides of the Coin. Cell Transplant. 2018, 27, 31–44. [Google Scholar] [CrossRef]
- Ye, L.; Li, X.; Li, L.; Chen, H.; Ge, R.S. Insights into the Development of the Adult Leydig Cell Lineage from Stem Leydig Cells. Front. Physiol. 2017, 8, 430. [Google Scholar] [CrossRef]
- Dentification of a Specific Sertoli Cell Marker, Sox9, for Use in Transplantation—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/12428738/ (accessed on 14 April 2023).
- Wen, Y.; Ma, X.; Wang, X.; Wang, F.; Dong, J.; Wu, Y.; Lv, C.; Liu, K.; Zhang, Y.; Zhang, Z.; et al. HnRNPU in Sertoli Cells Cooperates with WT1 and Is Essential for Testicular Development by Modulating Transcriptional Factors Sox8/9. Theranostics 2021, 11, 10030–10046. [Google Scholar] [CrossRef]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 1438. [Google Scholar] [CrossRef]
- Miki, T.; Marongiu, F.; Dorko, K.; Ellis, E.C.S.; Strom, S.C. Isolation of Amniotic Epithelial Stem Cells. Curr. Protoc. Stem Cell Biol. 2010, 12, 1E.3.1–1E.3.10. [Google Scholar] [CrossRef] [PubMed]
- Magatti, M.; Caruso, M.; De Munari, S.; Vertua, E.; De, D.; Manuelpillai, U.; Parolini, O. Human Amniotic Membrane-Derived Mesenchymal and Epithelial Cells Exert Different Effects on Monocyte-Derived Dendritic Cell Differentiation and Function. Cell Transplant. 2015, 24, 1733–1752. [Google Scholar] [CrossRef]
- Antonucci, I.; Iezzi, I.; Morizio, E.; Mastrangelo, F.; Pantalone, A.; Mattioli-Belmonte, M.; Gigante, A.; Salini, V.; Calabrese, G.; Tetè, S.; et al. Isolation of Osteogenic Progenitors from Human Amniotic Fluid Using a Single Step Culture Protocol. BMC Biotechnol. 2009, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Pipino, C.; Pierdomenico, L.; Di Tomo, P.; Di Giuseppe, F.; Cianci, E.; D’Alimonte, I.; Morabito, C.; Centurione, L.; Antonucci, I.; Mariggiò, M.; et al. Molecular and Phenotypic Characterization of Human Amniotic Fluid-Derived Cells: A Morphological and Proteomic Approach. Stem Cells Dev. 2015, 24, 1415–1428. [Google Scholar] [CrossRef]
- Johnsen, S.G. Testicular Biopsy Score Count—A Method for Registration of Spermatogenesis in Human Testes: Normal Values and Results in 335 Hypogonadal Males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Barboni, B.; Russo, V.; Curini, V.; Mauro, A.; Martelli, A.; Muttini, A.; Bernabò, N.; Valbonetti, L.; Marchisio, M.; Di Giacinto, O.; et al. Achilles Tendon Regeneration Can Be Improved by Amniotic Epithelial Cell Allotransplantation. Cell Transplant. 2012, 21, 2377–2395. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A.; Russo, V.; Di Marcantonio, L.; Berardinelli, P.; Martelli, A.; Muttini, A.; Mattioli, M.; Barboni, B. M1 and M2 Macrophage Recruitment during Tendon Regeneration Induced by Amniotic Epithelial Cell Allotransplantation in Ovine. Res. Vet. Sci. 2016, 105, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Saleh Jaweesh, M.; Hammadeh, M.E.; Dahadhah, F.W.; Al Zoubi, M.S.; Amor, H. Association between the Single Nucleotide Variants of the Mitochondrial Cytochrome B Gene (MT-CYB) and the Male Infertility. Mol. Biol. Rep. 2022, 49, 3609. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Localization | Function | hAECs T0 | hAECs-Treated Group | hAFMSCs T0 | hAFMSCs-Treated Group |
|---|---|---|---|---|---|---|
| beta 2 Microglobulin | Cell membrane | Component of the class I major histocompatibility complex | Negative | Negative | Negative | Negative |
| human CYP11A1 | Mitochondrion membrane | Leydig cells marker | Positive | Positive | Positive | Negative |
| SOX9 | Nucleus | Sertoli cells marker | Negative | Negative | Positive | Positive |
| SOHLH1 | Nucleus/Cytoplasm | Spermatogonia | Negative | Negative | Negative | Negative |
| NGN3 | Nucleus | Early spermatogonia | Negative | Negative | Negative | Negative |
| Primary Antibody | Dilution | Secondary Antibody | Dilution |
|---|---|---|---|
| Anti-human beta Microglobulin (ab181727) Abcam, Cambridge, UK | 1:250 | Rabbit Anti-Mouse IgG-Alexa Fluor® 488 (ab150125) Abcam, Cambridge, UK | 1:500 |
| Anti-human CYP11A1 (ab75497) Abcam, Cambridge, UK | 1:200 | Goat Anti-Rabbit IgG-Alexa Fluor® 488 (Abcam; ab150077) Abcam, Cambridge, UK | 1:200 |
| Anti-human SOX9 (ab182579) Abcam, Cambridge, UK | 1:500 | Rabbit Anti-Mouse IgG-Alexa Fluor® 488 (Abcam; ab150125) | 1:500 |
| Anti-human SOHLH1 (LS-C161387) LS Bio, Seattle, WA | 1:200 | Goat Anti-Rabbit IgG-Alexa Fluor® 488 (Abcam; ab150077) Abcam, Cambridge, UK | 1:200 |
| Ant-human NGN3 (ab38548) Abcam, Cambridge, UK | 1:500 | Goat Anti-Rabbit IgG-Alexa Fluor® 488 (Abcam; ab150077) Abcam, Cambridge, UK | 1:200 |
| Anti-CD86 antibody (ab119857) Abcam, Cambridge, UK | 1:50 | Rabbit Anti-Mouse IgG-Alexa Fluor® 488 (ab150125) Abcam, Cambridge, UK | 1:500 |
| Anti-CD206 antibody (ab64693) Abcam, Cambridge, UK | 1:50 | Goat Anti-Rabbit IgG-Alexa Fluor® 488 (Abcam; ab150077) Abcam, Cambridge, UK | 1:200 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| NAPE-PLD | 5′-TGTCCCGGGTTCCAAAGAGGAGC-3′ | 5′-ACCATCAGCGTCGCGTGTCC-3′ |
| FAAH | 5′-ATGGAAGTCCTCCAAGAGC-3′ | 5′-TAGAGCTTTCAGGCATAGCG-3′ |
| DAGLα | 5′-ATTCTCTCCTTCCTCCTGC-3′ | 5′-ATTTGGGCTTGGTGCTTCG-3′ |
| MAGL | 5′-ATGTTGAAGAGGCTGGACATGC-3′ | 5′-ATGCAGATTCCGGATTGGC-3′ |
| CB1 | 5′-TTCCACCGTAAAGACAGCCC-3′ | 5′-TCCACATCAGGCAAAAGGCC-3′ |
| CB2 | 5′-TTGACCGATACCTATGTCTGTGC-3′ | 5′-TGCTTTCCAGAGGACATACCC-3′ |
| TRPV1 | 5′-ATTGAACGGCGGAACATGACG-3 | 5′-ATCTCTTCCAGCTTCAGCG-3′ |
| CD86 | 5′ AAGACATGTGTAACCTGCACC 3′ | 5′ ACAGAACCGACTTTTTCCGGT 3′ |
| CD206 | 5′ AACTTCATCTGCCAGCGACA 3′ | 5′ CGTGCCTCTTTCCAGGTCTT 3′ |
| IL10 | 5′ CCTGCTCTTACTGGCTGGAG 3′ | 5′ TGTTGTCCAGCTGGTCCTTC 3′ |
| IL12 | 5′ CCGGTCCAGCATGTGTCAAT 3′ | 5′ CTTGGCAGGTCCAGAGACTG 3′ |
| GAPDH | 5′-AGACAGCCGCATCTTCTTGT-3′ | 5′-CTTGCCGTGGGTAGAGTCAT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peserico, A.; Barboni, B.; Russo, V.; Nardinocchi, D.; Turriani, M.; Cimini, C.; Bernabò, N.; Parolini, O.; Silini, A.R.; Antonucci, I.; et al. AEC and AFMSC Transplantation Preserves Fertility of Experimentally Induced Rat Varicocele by Expressing Differential Regenerative Mechanisms. Int. J. Mol. Sci. 2023, 24, 8737. https://doi.org/10.3390/ijms24108737
Peserico A, Barboni B, Russo V, Nardinocchi D, Turriani M, Cimini C, Bernabò N, Parolini O, Silini AR, Antonucci I, et al. AEC and AFMSC Transplantation Preserves Fertility of Experimentally Induced Rat Varicocele by Expressing Differential Regenerative Mechanisms. International Journal of Molecular Sciences. 2023; 24(10):8737. https://doi.org/10.3390/ijms24108737
Chicago/Turabian StylePeserico, Alessia, Barbara Barboni, Valentina Russo, Delia Nardinocchi, Maura Turriani, Costanza Cimini, Nicola Bernabò, Ornella Parolini, Antonietta Rosa Silini, Ivana Antonucci, and et al. 2023. "AEC and AFMSC Transplantation Preserves Fertility of Experimentally Induced Rat Varicocele by Expressing Differential Regenerative Mechanisms" International Journal of Molecular Sciences 24, no. 10: 8737. https://doi.org/10.3390/ijms24108737
APA StylePeserico, A., Barboni, B., Russo, V., Nardinocchi, D., Turriani, M., Cimini, C., Bernabò, N., Parolini, O., Silini, A. R., Antonucci, I., Stuppia, L., Berardinelli, P., Falanga, I., Perruzza, D., Valbonetti, L., & Mauro, A. (2023). AEC and AFMSC Transplantation Preserves Fertility of Experimentally Induced Rat Varicocele by Expressing Differential Regenerative Mechanisms. International Journal of Molecular Sciences, 24(10), 8737. https://doi.org/10.3390/ijms24108737