
Targeting the Inflammatory Hallmarks of Obesity-Associated Osteoarthritis: Towards Nutraceutical-Oriented Preventive and Complementary Therapeutic Strategies Based on n-3 Polyunsaturated Fatty Acids
 , , ,
, , ,
Abstract
:
1. Introduction
Literature Search Strategy
2. Pathogenesis of ObOA: From a Biomechanics-Centered Theoretical Paradigm toward a Key Role of Inflammation
2.1. Role of Impaired Loading and Biomechanics in Joint Degeneration in ObOA
2.2. Role of Inflammation in Joint Degeneration in ObOA
3. State-of-the-Art of n-3 PUFAs’ Role in Counteracting Inflammation: Toward Evidence-Based Medicine for Their Use in ObOA
3.1. FAs Metabolism: Focus on Inflammation
3.2. Effect of n-3 PUFAs in ObOA: Focus on Anti-Inflammatory and Pro-Resolving Mediators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | In Vitro Model | Treatment | Main Effects | Specific Outcomes | Ref. |
|---|---|---|---|---|---|
| DHA vs. palmitic acid (PA, SFA), oleic acid (OLA, MUFA) and linoleic acid (LA, n-6 PUFAs) | SW1353 chondrosarcoma cells ± LPS (1000 ng/mL) | 6.25, 25, 100 μM | Reduced pyroptosis-dependent inflammatory response due to inhibition of TLR4/NF-κB and NLRP3/caspase-1/GSDMD signaling. | DHA, OLA
| [60] |
| DHA | SW1353 chondrosarcoma ± IL-1β (5–100 ng/mL) | 3.125, 6.25, 12.5, 25, 50 µg/mL | Reduced inflammatory-dependent catabolic response by inhibition of p38 MAPK-dependent signaling. |
| [106] |
| DHA | Human chondrocytes ± TNF-α (50 ng/mL) | 25 μM for 48 h | Reduced inflammatory and catabolic response through downregulating Wnt/β-catenin and NF-κB signaling pathways. |
| [65] |
| DHA | Murine chondrocytes ± IL-1β (10 ng/mL) | 5, 10, 25 μM, pre-treatment for 24 h | Reduced inflammatory and catabolic response by inhibition of NF-κB p65 and β-catenin by Malat-1. |
| [111] |
| EPA/DHA/ALA | Bovine chondrocytes ± IL-1α (10 ng/mL) | 2.5, 5, 10, 20, 30 μg/mL for 8 h | Reduced inflammatory and catabolic response, efficiency EPA > DHA > ALA (modulation of more targets, modulation at low concentration). | EPA:
| [112] |
| EPA/DHA vs. AA | Canine chondrocytes ± IL-1β (10 ng/mL) | 10 μM for 8 days | Reduced inflammatory and catabolic response, efficiency EPA > DHA AA positively modulates some markers of inflammation. | EPA:
| [120] |
| EPA/DHA | Bovine cartilage explants ± IL-1β (10 ng/mL) | 0.1, 1, 10 μM EPA and/or DHA for 5 days | Reduce cytokine-induced articular cartilage degradation. Efficiency EPA > DHA at long term. |
| [113] |
| Green-lipped mussel (GLM) (abundant in DHA) | Human OA chondrocytes ± IL-1β (20 ng/mL) | 10, 100, 250 μg/mL | Reduced inflammatory response and necroptosis. |
| [115] |
| EPA | Normal human knee chondrocytes ± SNP (1 mM) | 10, 30, 50 µg/mL for 8 h | Reduced inflammatory-dependent catabolic response and apoptosis by inhibition of MAPK signaling. |
| [114] |
| RvD1 (DHA metabolite) | Human OA chondrocytes ± IL-1β (1 ng/mL) | 0–10 μM | Reduced inflammation by inactivation of NF-κB/p65, p38/MAPK and JNK1/2. |
| [117] |
| LA (n-6 PUFA) vs. ALA (n-3 PUFA) | Chondrocytes at high density ± IL-1β (100 pg/mL) | LA/ALA (1:1, 2:1, 4:1, 6:1, 8:1, 10:1) total amount 50 μg/mL for 1 h | Anti-catabolic effect, most effective ratio was 1:1, and 10:1 was not effective. |
| [121] |
| PDX | Rat chondrocytes ± IL-1β (10 ng/mL) | Pretreatment, 0.5, 1, 2, 4 μM | Inhibited inflammatory responses through the activation of AMPK and inhibition of NF-κB signaling pathway. |
| [118] |
| Conjugated linoleic acids + AA or EPA linoleic acid (LA) + AA or EPA | Human OA chondrocytes | 10 μM | Anti-inflammatory. |
| [131] |
| DHA | Murine bone marrow mesenchymal stromal cells during chondrogenesis ± IL-1β (10 ng/mL) | 25 μM, pre-treatment for 21 days | Rescued IL-1β-impaired chondrogenesis by NF-κB signaling inhibition by Malat-1. |
| [111] |
| DHA | 293 T cells ± TNFα (10 ng/mL) | 5, 10, 25 μM, for 24 h | Downregulated Wnt/β-catenin and NF-κB signaling. | Luciferase activities of reporter vector harbouring Wnt/β-catenin (TOPFlash) and NF-κB response element (NF-κB RE) showed a declining gradient. | [111] |
| 13- and 16-H- DHEA and DHEA (endocannabinoid from DHA) | RAW264.7 macrophage ± LPS (1 µg/mL) | 2.5–5 μM | Anti-inflammatory effects less pronounced compared to DHEA. | DHEA
| [127] |
| 13S,14S-epoxy- DHA (precursor of MaR-1) | Human macrophages (M1 and M2 subsets) | 10 nM | Reduced inflammation and switching from M1 to M2 phenotype. |
| [128] |
| DHA/EPA | Equine synoviocytes ± IL-1β (5 ng/mL) | 25–50 μM for 24 h | Reduced inflammatory and catabolic response due to increased integration within cell membranes and production of oxylipids (specialized pro-resolving mediators). | DHA
| [108] |
| RvD-1, -2, MaR-1, PDX | Equine synovial fibroblasts ± IL-1β (5 ng/mL) | Pre-treatment with 25 μM and 50 μM EPA and DHA | Reduced inflammatory and catabolic response due to increased integration within cells membrane. |
| [108] |
| RvD1 | Human OA fibroblast-like synoviocytes (FLs) | 20, 50, 100, 200 nM | Reduced inflammatory and catabolic response due to Hippo-YAP signaling pathway activation. |
| [123] |
| MaR-1 (DHA metabolite) | Rat FLSs ± IL-1β (10 ng/mL) | Pretreatment, 10, 100, 1000 nM, 1 h | Anti-inflammatory and anti-catabolic effect by stimulation of PI3k/Akt pathway and inhibition of NF-κB p65 pathway. |
| [124] |
| RvD1/RvE1/MaR1 | OA synovial fibroblast ± TNF-α (10 ng/mL) | Pretreatment, 100 nM | No anti-inflammatory effect. |
| [122] |
4. Current and Novel Perspectives of Dietary Interventions in ObOA: Strengths and Weaknesses of the Potential Use of PUFAs
4.1. Clinical Evidence of n-3PUFA in OA: Insights for Better ObOA Therapies
| Molecule Tested | Study Types | Patient Data | Treatments/ Follow-Up (F.up) | Main Effects | Ref. |
|---|---|---|---|---|---|
| FA intake | Prospective study. | N = 2092 participants with radiographic knee OA. | Followed at yearly intervals up to 48 months. Questionnaire for food intake. | Significant positive relationships between total fat and SFA with joint space width loss were observed. MUFA, PUFA and a higher ratio of PUFA to SFA were associated with a reduced joint space width loss. | [182] |
| Fasting plasma phospholipid n-6 (AA) and n-3 PUFAs (EPA and DHA) with synovitis | Multicenter Osteoarthritis Study (MOST). | N = 472 patients with knee OA (50% women). Mean age = 60 year. BMI = 30 (1° grade of Ob). | n-3 PUFAs. n-6 PUFAs. | Multivariable logistic regression showed the following:
| [165] |
| Fish oil (FO) (DHA + EPA) | Randomized, double-blind clinical study. | N = 152 older adults between 50 and 80 years. BMI 25–40 kg/m2 | Group 1—FO 2000 mg/day DHA + 400 mg/day EPA). Group 2—CUR curcumin (160 mg/day). Group 3—FO + CUR. |
| [164] |
| Cod liver oil (EPA) | A double-blind, placebo- RCT. | N = 26; Female, n = 21; Age range = 52–85 years. | Group 1—EPA oil (10 mL/d EPA) and ibuprofen (1200 mg/d). Group 2—placebo (oil of undescribed content) and ibuprofen. F.up = 6 months. |
| [166] |
| Cod liver oil (EPA + DHA) + NSAIDs | A double-blind, placebo- RCT. | N = 86; female, n = 60; Age range = 49–87 years. | Group 1—cod liver oil (10 mL of oil containing 786 mg EPA) + NSAIDs. Group 2—Placebo (10 mL olive oil) + NSAIDs. F.up = 24 weeks. |
| [167] |
| GLM (high proportion of EPA and DHA + low presence of several minor lipid components) | A double-blind, placebo- RCT. | N = 80; Female, n = 44; age = 66.4 ± 10 years. Pain rated > 30 mm in the last week on 100 mm VAS. | Group 1—GLM extract (600 mg/d). Group 2—Placebo (600 mg/d corn oil). F.-up = 0, 6, 12 and 15 wks. |
| [168] |
| Lyprinol® (a lipid extract of GLM rich in EPA and DHA) | A double-blind, placebo- RCT. | N = 80 patients with knee OA female, n = 69 knee pain, radiographic evidence of osteophytes. | GLM group—four capsules of Lyprinol®/day. Placebo group—olive oil in the same number of capsules. F.up = 6 months. Revision at week 0, 2, 4, 8, 12, 18 and 24. |
| [169] |
| GLM extract | Non-blinded randomized clinical trial. | N = 38 patients with knee OA. | Group 1—GLM extract (3000 mg/day). Group 2—glucosamine (3000 mg/day). Treatment for 12 weeks. |
| [172] |
| Phytalgic® (fish oil rich in n-3 PUFA+ n-6 PUFAs+ vitamin E, Urtica dioica) | Randomized double-blind parallel-groups clinical trial. | N = 81 patients with OA of the knee or hip using NSAIDs and/or analgesics regularly. Female, n = 55; Mean age = 57.5; Age range = 28–84 years) F.up = 3 months. | Group 1—Phytalgic® (n = 41). Group 2—placebo (n = 40). |
| [173] |
| EPA+ DHA | A randomized, double-blind, multicenter trial. Trial registration number ACTRN 12607000415404. | N = 202 patients with knee OA. Female, n = 100; Mean age = 61 ± 10 years, Participants were >40 years with clinical knee and VAS > 20 mm, No indication of BMI. | Group 1—hHigh-dose fish oil (4.5 g EPA + DHA per day) (59% women) 15 mL/day. Group 2—low-dose fish oil (0.45 g EPA + DHA per day) (40% women). |
| [174] |
| Neptune Krill Oil (NKOTM) EPA (20:5 n-3) + DHA (22:6 n-3) + antioxidants (e.g., astaxanthin, etc.) | A randomized, double-blind, placebo-controlled study. | N = 90 patients with cardiovascular disease and/or rheumatoid arthritis and/or OA and high levels of CRP (>1.0 mg/dL). | Group 1—treatment with NKO™ (300 mg daily). Group 2—placebo 30 days of treatment. |
| [175] |
| Krill oil (EPA (20:5 n-3) + DHA (22:6 n-3)) | Randomized, double-blind, parallel-group, placebo-controlled trial. | N = 50 patients with mild knee pain (no severe pain). | Group 1—treatment with 2 g/day. Group 2—placebo. F.up = 30 days. |
| [176] |
| Krill oil (EPA (20:5 n-3) + DHA (22:6 n-3)) | Multicenter, randomized, double-blind, placebo-controlled clinical trial. | N = 260 patients with clinical knee OA, significant knee pain and effusion-synovitis. | Group 1—treatment of 2 g/day. Group 2—placebo. F.up = 6 months. |
| [177] |
| Krill oil (EPA (20:5 n-3) + DHA (22:6 n-3))] | Multicenter, randomized, double-blind, placebo-controlled trial | N = 235; Female, n = 129; Mean age = 55.9 ± 6.8 yrs; BMI > 18.5 to <35 kg/m2; Mild-to-moderate knee OA. | Group 1—4 g/d of a commercially available krill oil supplement daily (0.60 g EPA/d, 0.28 DHA/d, 0.45 mg astaxanthin/d). Group 2—placebo (4 g/d mixed vegetable oil). F.up = 6 months. |
| [178] |
| Combination of glucosamine sulfate + EPA DHA | RCT, a double-blind study. | N = 177 patients with moderate-to-severe hip or knee OA. Mean age 62 y; mean BMI = 29; 63% women. | Group 1—glucosamine sulfate + EPA DHA. Group 2—glucosamine sulfate alone. |
| [179] |
| EPA + l-serine | Randomized, double-blind, placebo-controlled, parallel-group study. | N = 120 participants aged ≥ 20 y (36 men and 84 women: mean ± SD age = 40.8 ± 10.9 year. | Group 1—oral administration of 549 mg l-serine+ 149 mg/daily EPA. Group 2—placebo group. 8 wk dosing and 4 wk post-treatment observation. |
| [180] |
| Resolvins D1, D2 and 17-HDHA, DHA | OA case-control cohort. | N = 62 individuals affected with radiographic knee OA (Kellgren–Lawrence grade of 2 or higher). 52 individuals without radiographic or clinical symptoms of OA. | No treatment gas chromatography |
| [181] |
4.2. Nutritional Recommendations for ObOA Patients
4.3. PUFA-Based Approaches for OA: From Current Evidence to New Perspectives for Intra-Articular Administration
| Molecule | In Vivo Model | Dose/Delivery Route | Main Effects | Specific Outcomes | Indications for Preventive-Therapeutic Strategies for ObOA | Ref. |
|---|---|---|---|---|---|---|
| RvD1 | ObOA model: DMM model C57Bl/J6 mice + HFD (45 kcal% fat) vs. control diet (10 kcal% fat). | 20 ng/μL (one week before and on weeks 1 and 6 after OA induction). IA injection | Protective role of IA injection of the pro-resolving RvD1 in modulating macrophage phenotype to counteract inflammation. |
| The potential of targeting macrophage phenotypes to prevent OA aggravation. | [161] |
| RvD1-loaded nano-liposomal formulation (Lipo-RvD1) | Post-traumatic OA model: DMM model in male C57BL/6 mice. | Liposomes (~1 mg per joint in a total volume of 10 μL). IA injection. | Lipo-RvD1 formulation could be a therapeutic candidate thanks to its anti-inflammatory and analgesic properties. |
| The potential of targeting macrophage phenotypes to prevent OA aggravation with analgesic effects. | [222] |
| RvD1 | ObOA model: DMM model + HFD. | RvD1 encapsulated in liposomes (lipo-RvD1). | Improved joint health following the treatment with the lipo- than the free RvD1 treatment. |
| The potential of lipo-RvD1 as an anti-OA agent. | [234] |
| MaR-1 | Inflammatory model of OA: MIA model in Sprague–Dawley rats. | 10 ng MaR-1 + 50 µL sterile saline (two treatments per week for 4 weeks). IA injection. | Chondroprotective effects in mitigating OA progression. |
| The potential of promoting cartilage repair. | [124] |
| Gelatin hydrogels with EPA | Post-traumatic OA: DMM model in mice. | Group 1—SHAM. Group 2—DMM. Group 3—DMM + corn oil. Group 4—EPA-I (DMM + corn oil and EPA). Group 5—control (DMM + gelatin hydrogels). Group 6—EPA-G (DMM + gelatin hydrogels containing EPA). IA injection. | Hydrogel incorporating EPA was more effective in attenuating the inflammatory effects underlying the progression of OA. | Gelatin hydrogels containing EPA were more potent compared with a single EPA injection through:
| IA administration of controlled-release EPA can be a new therapeutic approach to target inflammatory and catabolic markers also in patients with ObOA. | [235] |
| Seed oil (DSO) in niosomes | Carrageenan-induced paw oedema in rats. | 0.5 g/kg DSO pure extract a day. i.p. injection. | Controlled release and therapeutic effective level of DSO niosomes in mitigating OA progression |
| Nanoparticles as a targeted delivery system can be a valuable tool for ObOA. | [236] |
| Cel-MEs@MNs (microemulsion-incorporated dissolving microneedle co-loading celecoxib and α-linolenic acid) | OA model. | Transdermal injection vs. oral administration of celecoxib and α-linolenic acid. | Synergistic anti-inflammation and potent transdermal delivery, |
| Microemulsion with improved transdermal injection potency holds great potential in the solubilization of water-insoluble drugs. | [240] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| 9-HODE | 9-hydroxy octadecadienoic acid |
| AA | arachidonic acid |
| ACLT | anterior cruciate ligament transection |
| ADAMTs | aggrecanases |
| AEA | ethanolamine |
| ALA | α-linoleic acid |
| AMPK | AMP-activated protein kinase |
| AP | ascorbyl palmitate |
| BMI | body mass index |
| BMD | bone mineral density |
| CB | cannabinoid receptor |
| ChemR23 | chemoattractant receptor 23 |
| Coll II | collagen II |
| COX | cyclooxygenase |
| CRP | c-reactive protein |
| CUR | curcumin |
| DAMPs | damage-associated molecular patterns |
| DGLA | dihomo-y-linolenic acid |
| DHA | docosahexaenoic acid |
| DHEA | docosahexaenoyl ethanolamide |
| DHETs | di-hydroxy-eicosatrienoic acids |
| DMM | destabilization of the medial meniscus |
| DSO | date seed oil |
| ECM | extracellular matrix |
| EEs | ethyl esters |
| EETs | epoxyeicosatrienoic acids |
| EP4 | prostaglandin EP4 receptor |
| EPA | eicosapentaenoic acid |
| EPEA | ethanolamide |
| FAAH | fatty acid amide hydrolase |
| FAs | fatty acid |
| FPR2/ALX | formyl peptide receptor 2 |
| GAG | glycosaminoglycans |
| GI | gastrointestinal |
| GLA | gamma-linolenic acid |
| GLM | green-lipped mussel |
| GPR120 | G-protein coupled receptor 120 |
| GSDMD | gansdermin D |
| HA | hyaluronan |
| HETEs | 5-hydroxyeicosatetraenoic acid |
| HFD | high fat diet |
| IA | intra-articular |
| IFP | infrapatellar fat pad |
| MAPK | mitogen-activated phosphokinase |
| IL | interleukin |
| IL-1RN | interleukin 1 receptor antagonist |
| iNOS | inducible nitric oxide synthase |
| KL | kellegren/lawrence |
| LA | linoleic acid |
| lncRNAs | long non-coding RNA |
| LO | lipoxygenase |
| LPS | lipopolysaccaride |
| l-Ser | l-serine |
| M2 | macrophage |
| MAG | monoglycerides |
| Malat-1 | metastasis-associated lung adenocarcinoma transcript 1 |
| Mar-1 | maresins |
| MFD | medium fat diet |
| MIA | monosodium iodoacetate injections |
| MLKL | mixed lineage kinase domain-like protein |
| MMPs | metalloproteinases |
| MNX | medial meniscal transection |
| mTOR | mammalian target of rapamycin |
| MUFAs | monounsaturated fatty acid |
| NF-κB | nuclear factor-κB |
| NLRP | NOD-like receptor protein |
| NMDA | n-methyl-d-aspartate |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| OA | osteoarthritis |
| Ob | obesity |
| ObOA | obesity-associated osteoarthritis |
| OLA | oleic acid |
| PA | palmitic acid |
| PAMPs | pathogen-associated moleculars patterns |
| PDX | protectin-D |
| PGE2 | prostaglandin-E2 |
| PGD2 | prostaglandin D2 |
| PGH2 | prostaglandin H2 |
| PGI2 | prostaglandin I2 |
| PGF2α | prostaglandin F2α |
| PKC | protein kinase C |
| PLs | phospholipid species |
| PO | peanut oil |
| PPRs | pattern recognition receptors |
| PUFAs | n-3 polyunsaturated fatty acids |
| RANKL | receptor activator of NF-kappaB ligand |
| RCT | randomized controlled trials |
| RIP | receptor interacting protein kinase 1 |
| ROS | reactive oxygen species |
| RUNX-2 | Runt-related transcription factor 2 |
| RvD | D-series resolvins |
| RvE | E-series resolvins |
| SF | synovial fluid |
| SFAs | saturated fatty acid |
| SNP | sodium nitroprusside |
| SO | soybean oil |
| SPMs | specialized pro-resolving mediators |
| TD2 | type II diabetes |
| TGs | triglicerides |
| TG | transgenic |
| THA | tetracosahexaenoic acid |
| THC | Δ9-tetrahydrocannabinol |
| TIMP | tissue inhibitors of metalloproteinases |
| TLR | toll-like receptor |
| TNF | tumour necrosis factor |
| TRAP | tartrate-resistant acid phosphatase |
| TRPV1 | transient receptor potential vanilloid 1 |
| WOMAC | Western Ontario and McMaster Universities Arthritis Index |
References
- Oliveria, S.A.; Felson, D.; Cirillo, P.A.; Reed, J.I.; Walker, A.M. Body weight, body mass index, and incident symptomatic osteoarthritis of the hand, hip, and knee. Epidemiology 1999, 10, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Lou, C.; Cahue, S.; Dunlop, D.D. The mechanism of the effect of obesity in knee osteoarthritis: The mediating role of malalignment. Arthritis Rheum. 2000, 43, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, C. Body mass index and risk of knee osteoarthritis: Systematic review and meta-analysis of prospective studies. BMJ Open 2015, 5, e007568. [Google Scholar] [CrossRef] [PubMed]
- Johnson, V.L.; Hunter, D.J. The epidemiology of osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2014, 28, 5–15. [Google Scholar] [CrossRef]
- Coggon, D.; Reading, I.; Croft, P.; McLaren, M.; Barrett, D.; Cooper, C. Knee osteoarthritis and obesity. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 622–627. [Google Scholar] [CrossRef]
- Guilak, F. Biomechanical factors in osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2011, 25, 815–823. [Google Scholar] [CrossRef]
- Scotece, M.; Conde, J.; Lopez, V.; Lago, F.; Pino, J.; Gómez-Reino, J.J.; Gualillo, O. Adiponectin and leptin: New targets in inflammation. Basic Clin. Pharmacol. Toxicol. 2014, 114, 97–102. [Google Scholar] [CrossRef]
- Azamar-Llamas, D.; Hernandez-Molina, G.; Ramos-Avalos, B.; Furuzawa-Carballeda, J. Adipokine Contribution to the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2017, 2017, 5468023. [Google Scholar] [CrossRef]
- Poonpet, T.; Honsawek, S. Adipokines: Biomarkers for osteoarthritis? World J. Orthop. 2014, 5, 319–327. [Google Scholar] [CrossRef]
- Lippiello, L.; Walsh, T.; Fienhold, M. The association of lipid abnormalities with tissue pathology in human osteoarthritic articular cartilage. Metabolism 1991, 40, 571–576. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef]
- Liput, K.P.; Lepczyński, A.; Ogłuszka, M.; Nawrocka, A.; Poławska, E.; Grzesiak, A.; Ślaska, B.; Pareek, C.S.; Czarnik, U.; Pierzchała, M. Effects of Dietary n-3 and n-6 Polyunsaturated Fatty Acids in Inflammation and Cancerogenesis. Int. J. Mol. Sci. 2021, 22, 6965. [Google Scholar] [CrossRef]
- Abshirini, M.; Ilesanmi-Oyelere, B.L.; Kruger, M.C. Potential modulatory mechanisms of action by long-chain polyunsaturated fatty acids on bone cell and chondrocyte metabolism. Prog. Lipid Res. 2021, 83, 101113. [Google Scholar] [CrossRef]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef]
- Wu, C.L.; Kimmerling, K.A.; Little, D.; Guilak, F. Serum and synovial fluid lipidomic profiles predict obesity-associated osteoarthritis, synovitis, and wound repair. Sci. Rep. 2017, 7, 44315. [Google Scholar] [CrossRef]
- Micallef, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 Polyunsaturated Fatty Acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Gil, A. Polyunsaturated fatty acids and inflammatory diseases. Biomed. Pharmacother. 2002, 56, 388–396. [Google Scholar] [CrossRef]
- Oppedisano, F.; Bulotta, R.M.; Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Ilari, S.; Serra, M.; Muscoli, C.; Gratteri, S.; et al. The Role of Nutraceuticals in Osteoarthritis Prevention and Treatment: Focus on n-3 PUFAs. Oxid. Med. Cell. Longev. 2021, 2021, 4878562. [Google Scholar] [CrossRef]
- AlAli, M.; Alqubaisy, M.; Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Molouki, A.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. Nutraceuticals: Transformation of Conventional Foods into Health Promoters/Disease Preventers and Safety Considerations. Molecules 2021, 26, 2540. [Google Scholar] [CrossRef]
- Votava, L.; Schwartz, A.G.; Harasymowicz, N.S.; Wu, C.-L.; Guilak, F. Effects of dietary fatty acid content on humeral cartilage and bone structure in a mouse model of diet-induced obesity. J. Orthop. Res. 2019, 37, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pora, B.L.R.; Dong, K.; Hasjim, J. Health benefits of docosahexaenoic acid and its bioavailability: A review. Food Sci. Nutr. 2021, 9, 5229–5243. [Google Scholar] [CrossRef] [PubMed]
- Parratte, S.; Pesenti, S.; Argenson, J.N. Obesity in orthopedics and trauma surgery. Orthop. Traumatol. Surg. Res. 2014, 100, S91–S97. [Google Scholar] [CrossRef] [PubMed]
- King, L.K.; March, L.; Anandacoomarasamy, A. Obesity & osteoarthritis. Indian J. Med. Res. 2013, 138, 185–193. [Google Scholar] [PubMed]
- Felson, D.T.; Anderson, J.J.; Naimark, A.; Walker, A.M.; Meenan, R.F. Obesity and knee osteoarthritis. The Framingham Study. Ann. Intern. Med. 1988, 109, 18–24. [Google Scholar] [CrossRef]
- Koonce, R.C.; Bravman, J.T. Obesity and osteoarthritis: More than just wear and tear. J. Am. Acad. Orthop. Surg. 2013, 21, 161–169. [Google Scholar]
- Thijssen, E.; van Caam, A.; van der Kraan, P.M. Obesity and osteoarthritis, more than just wear and tear: Pivotal roles for inflamed adipose tissue and dyslipidaemia in obesity-induced osteoarthritis. Rheumatology 2015, 54, 588–600. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, J.J.Y.; Li, G.; Yuan, J.; Ebert, J.R.; Li, H.; Papadimitriou, J.; Wang, Q.; Wood, D.; Jones, C.W.; et al. Pathogenesis and clinical management of obesity-related knee osteoarthritis: Impact of mechanical loading. J. Orthop. Translat. 2020, 24, 66–75. [Google Scholar] [CrossRef]
- Darling, E.M.; Athanasiou, K.A. Biomechanical strategies for articular cartilage regeneration. Ann. Biomed. Eng. 2003, 31, 1114–1124. [Google Scholar] [CrossRef]
- Rustenburg, C.M.; Emanuel, K.; Peeters, M.; Lems, W.F.; Vergroesen, P.-P.A.; Smit, T.H. Osteoarthritis and intervertebral disc degeneration: Quite different, quite similar. JOR Spine 2018, 1, e1033. [Google Scholar] [CrossRef]
- Ravalli, S.; Szychlinska, M.A.; Lauretta, G.; Musumeci, G. New Insights on Mechanical Stimulation of Mesenchymal Stem Cells for Cartilage Regeneration. Appl. Sci. 2020, 10, 2927. [Google Scholar] [CrossRef]
- Karsdal, M.A.; Madsen, S.H.; Christiansen, C.; Henriksen, K.; Fosang, A.J.; Sondergaard, B.C. Cartilage degradation is fully reversible in the presence of aggrecanase but not matrix metalloproteinase activity. Arthritis Res. Ther. 2008, 10, R63. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sport. Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Sarma, A.V.; Powell, G.L.; LaBerge, M. Phospholipid composition of articular cartilage boundary lubricant. J. Orthop. Res. 2001, 19, 671–676. [Google Scholar] [CrossRef]
- Kosonen, J.P.; Eskelinen, A.S.A.; Orozco, G.A.; Nieminen, P.; Anderson, D.D.; Grodzinsky, A.J.; Korhonen, R.K.; Tanska, P. Injury-related cell death and proteoglycan loss in articular cartilage: Numerical model combining necrosis, reactive oxygen species, and inflammatory cytokines. PLoS Comput. Biol. 2023, 19, e1010337. [Google Scholar] [CrossRef]
- Pratta, M.A.; Yao, W.; Decicco, C.; Tortorella, M.D.; Liu, R.-Q.; Copeland, R.A.; Magolda, R.; Newton, R.C.; Trzaskos, J.M.; Arner, E.C. Aggrecan protects cartilage collagen from proteolytic cleavage. J. Biol. Chem. 2003, 278, 45539–45545. [Google Scholar] [CrossRef]
- Radin, E.L.; Paul, I.L.; Rose, R.M. Role of mechanical factors in pathogenesis of primary osteoarthritis. Lancet 1972, 1, 519–522. [Google Scholar] [CrossRef]
- Kanthawang, T.; Bodden, J.; Joseph, G.B.; Lane, N.E.; Nevitt, M.; McCulloch, C.E.; Link, T.M. Obese and overweight individuals have greater knee synovial inflammation and associated structural and cartilage compositional degeneration: Data from the osteoarthritis initiative. Skeletal. Radiol. 2021, 50, 217–229. [Google Scholar] [CrossRef]
- Oliviero, F.; Nigro, A.L.; Bernardi, D.; Giunco, S.; Baldo, G.; Scanu, A.; Sfriso, P.; Ramonda, R.; Plebani, M.; Punzi, L. A comparative study of serum and synovial fluid lipoprotein levels in patients with various arthritides. Clin. Chim. Acta 2012, 413, 303–307. [Google Scholar] [CrossRef]
- Kosinska, M.K.; Ludwig, T.E.; Liebisch, G.; Zhang, R.; Siebert, H.-C.; Wilhelm, J.; Kaesser, U.; Dettmeyer, R.B.; Klein, H.; Ishaque, B.; et al. Articular Joint Lubricants during Osteoarthritis and Rheumatoid Arthritis Display Altered Levels and Molecular Species. PLoS ONE 2015, 10, e0125192. [Google Scholar] [CrossRef]
- Hui, A.Y.; McCarty, W.J.; Masuda, K.; Firestein, G.S.; Sah, R.L. A systems biology approach to synovial joint lubrication in health, injury, and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2012, 4, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Medvedeva, E.V.; Grebenik, E.A.; Gornostaeva, S.N.; Telpuhov, V.I.; Lychagin, A.V.; Timashev, P.S.; Chagin, A.S. Repair of Damaged Articular Cartilage: Current Approaches and Future Directions. Int. J. Mol. Sci. 2018, 19, 2366. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Adams, J.; Leddy, H.A.; McNulty, A.L.; O’conor, C.J.; Guilak, F. The mechanobiology of articular cartilage: Bearing the burden of osteoarthritis. Curr. Rheumatol. Rep. 2014, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Dahl, L.B.; Dahl, I.M.; Engstrom-Laurent, A.; Granath, K. Concentration and molecular weight of sodium hyaluronate in synovial fluid from patients with rheumatoid arthritis and other arthropathies. Ann. Rheum. Dis. 1985, 44, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, H.; Altmann, S. Rheology and hyaluronic acid in inflammatory joint effusions (author's transl). Wien Klin. Wochenschr. 1977, 89, 716–720. [Google Scholar]
- More, S.; Kotiya, A.; Ghosh, S.; Spyrou, L.; Sarris, I. Rheological properties of synovial fluid due to viscosupplements: A review for osteoarthritis remedy. Comput. Methods Programs Biomed. 2020, 196, 105644. [Google Scholar] [CrossRef]
- Pejovic, M.; Stankovic, A.; Mitrovic, D.R. Determination of the apparent synovial permeability in the knee joint of patients suffering from osteoarthritis and rheumatoid arthritis. Br. J. Rheumatol. 1995, 34, 520–524. [Google Scholar] [CrossRef]
- Schaffler, A.; Ehling, A.; Neumann, E.; Herfarth, H.; Tarner, I.; Scholmerich, J.; Muller-Ladern, U.; Gay, S. Adipocytokines in synovial fluid. JAMA 2003, 290, 1709–1710. [Google Scholar]
- Collins, K.H.; Lenz, K.L.; Pollitt, E.N.; Ferguson, D.; Hutson, I.; Springer, L.E.; Oestreich, A.K.; Tang, R.; Choi, Y.-R.; Meyer, G.A.; et al. Adipose tissue is a critical regulator of osteoarthritis. Proc. Natl. Acad. Sci. USA 2021, 118, e2021096118. [Google Scholar] [CrossRef]
- Grotle, M.; Hagen, K.B.; Natvig, B.; A Dahl, F.; Kvien, T.K. Obesity and osteoarthritis in knee, hip and/or hand: An epidemiological study in the general population with 10 years follow-up. BMC Musculoskelet. Disord. 2008, 9, 132. [Google Scholar] [CrossRef]
- Urban, H.; Little, C.B. The role of fat and inflammation in the pathogenesis and management of osteoarthritis. Rheumatology 2018, 57, iv10–iv21. [Google Scholar] [CrossRef]
- Clockaerts, S.; Bastiaansen-Jenniskens, Y.; Runhaar, J.; Van Osch, G.; Van Offel, J.; Verhaar, J.; De Clerck, L.; Somville, J. The infrapatellar fat pad should be considered as an active osteoarthritic joint tissue: A narrative review. Osteoarthr. Cartil. 2010, 18, 876–882. [Google Scholar] [CrossRef]
- Klein-Wieringa, I.R.; Kloppenburg, M.; Bastiaansen-Jenniskens, Y.M.; Yusuf, E.; Kwekkeboom, J.C.; El-Bannoudi, H.; Nelissen, R.G.H.H.; Zuurmond, A.; Stojanovic-Susulic, V.; Van Osch, G.J.V.M.; et al. The infrapatellar fat pad of patients with osteoarthritis has an inflammatory phenotype. Ann. Rheum. Dis. 2011, 70, 851–857. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Pearson, M.J.; Herndler-Brandstetter, D.; Tariq, M.A.; Nicholson, T.A.; Philp, A.M.; Smith, H.L.; Davis, E.T.; Jones, S.W.; Lord, J.M. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci. Rep. 2017, 7, 3451. [Google Scholar] [CrossRef]
- Sekar, S.; Shafie, S.R.; Prasadam, I.; Crawford, R.; Panchal, S.K.; Brown, L.; Xiao, Y. Saturated fatty acids induce development of both metabolic syndrome and osteoarthritis in rats. Sci. Rep. 2017, 7, 46457. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Sibille, K.T.; King, C.; Garrett, T.J.; Glover, T.L.; Zhang, H.; Chen, H.; Reddy, D.; Goodin, B.R.; Sotolongo, A.; Petrov, M.E.; et al. Omega-6: Omega-3 PUFA Ratio, Pain, Functioning, and Distress in Adults With Knee Pain. Clin. J. Pain 2018, 34, 182–189. [Google Scholar] [CrossRef]
- Valdes, A.M.; Ravipati, S.; Pousinis, P.; Menni, C.; Mangino, M.; Abhishek, A.; Chapman, V.; Barrett, D.A.; Doherty, M. Omega-6 oxylipins generated by soluble epoxide hydrolase are associated with knee osteoarthritis. J. Lipid Res. 2018, 59, 1763–1770. [Google Scholar] [CrossRef]
- Jin, X.; Dong, X.; Sun, Y.; Liu, Z.; Liu, L.; Gu, H. Dietary Fatty Acid Regulation of the NLRP3 Inflammasome via the TLR4/NF-kappaB Signaling Pathway Affects Chondrocyte Pyroptosis. Oxid. Med. Cell. Longev. 2022, 2022, 3711371. [Google Scholar] [CrossRef]
- Rogero, M.M.; Calder, P.C. Obesity, Inflammation, Toll-Like Receptor 4 and Fatty Acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzoglou, E.; Lopes, E.B.P.; Fu, Y.; Herron, J.C.; Flaming, J.M.; Donovan, E.L.; Hu, Y.; Filiberti, A.; Griffin, T.M.; Humphrey, M.B. TLR4 Promotes and DAP12 Limits Obesity-Induced Osteoarthritis in Aged Female Mice. JBMR Plus 2019, 3, e10079. [Google Scholar] [CrossRef] [PubMed]
- Drummer, C., IV; Saaoud, F.; Jhala, N.C.; Cueto, R.; Sun, Y.; Xu, K.; Shao, Y.; Lu, Y.; Shen, H.; Yang, L.; et al. Caspase-11 promotes high-fat diet-induced NAFLD by increasing glycolysis, OXPHOS, and pyroptosis in macrophages. Front. Immunol. 2023, 14, 1113883. [Google Scholar] [CrossRef] [PubMed]
- Miao, P.; Ruiqing, T.; Yanrong, L.; Zhuwen, S.; Huan, Y.; Qiong, W.; Yongnian, L.; Chao, S. Pyroptosis: A possible link between obesity-related inflammation and inflammatory diseases. J. Cell Physiol. 2022, 237, 1245–1265. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, D.; Ho, K.W.; Lin, S.; Suen, W.C.-W.; Zhang, H.; Zha, Z.; Li, G.; Leung, P.S. GPR120 is an important inflammatory regulator in the development of osteoarthritis. Arthritis Res. Ther. 2018, 20, 163. [Google Scholar] [CrossRef]
- Trayhurn, P.; Denyer, G. Mining microarray datasets in nutrition: Expression of the GPR120 (n-3 fatty acid receptor/sensor) gene is down-regulated in human adipocytes by macrophage secretions. J. Nutr. Sci. 2012, 1, e3. [Google Scholar] [CrossRef]
- Valdes, A.M. Metabolic syndrome and osteoarthritis pain: Common molecular mechanisms and potential therapeutic implications. Osteoarthr. Cartil. 2020, 28, 7–9. [Google Scholar] [CrossRef]
- Stevens, R.M.; Ervin, J.; Nezzer, J.; Nieves, Y.; Guedes, K.; Burges, R.; Hanson, P.D.; Campbell, J.N. Randomized, Double-Blind, Placebo-Controlled Trial of Intraarticular Trans-Capsaicin for Pain Associated With Osteoarthritis of the Knee. Arthritis Rheumatol. 2019, 71, 1524–1533. [Google Scholar] [CrossRef]
- Lv, Z.; Xu, X.; Sun, Z.; Shi, D. TRPV1 alleviates osteoarthritis by inhibiting M1 macrophage polarization via Ca(2+)/CaMKII/Nrf2 signaling pathway. Cell Death Dis. 2021, 12, 504. [Google Scholar] [CrossRef]
- Liu, D.Y.; Zhu, Z.M.; Tepel, M. The Role of Transient Receptor Potential Channels in Metabolic Syndrome. Hypertens. Res. 2008, 31, 1989–1995. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yan Liu, D.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yang, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J.; et al. Activation of transient receptor potential vanilloid type-1 channel prevents adipogenesis and obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef]
- Baskaran, P.; Nazminia, K.; Frantz, J.; O’neal, J.; Thyagarajan, B. Mice lacking endogenous TRPV1 express reduced levels of thermogenic proteins and are susceptible to diet-induced obesity and metabolic dysfunction. FEBS Lett. 2021, 595, 1768–1781. [Google Scholar] [CrossRef]
- Kennedy, A.; Martinez, K.; Chuang, C.-C.; LaPoint, K.; McIntosh, M. Saturated fatty acid-mediated inflammation and insulin resistance in adipose tissue: Mechanisms of action and implications. J. Nutr. 2009, 139, 1–4. [Google Scholar] [CrossRef]
- Bondeson, J.; Blom, A.B.; Wainwright, S.; Hughes, C.; Caterson, B.; Berg, W.B.V.D. The role of synovial macrophages and macrophage-produced mediators in driving inflammatory and destructive responses in osteoarthritis. Arthritis Rheum. 2010, 62, 647–657. [Google Scholar] [CrossRef]
- Yao, Z.; Qi, W.; Zhang, H.; Zhang, Z.; Liu, L.; Shao, Y.; Zeng, H.; Jin, J.; Pan, H.; Guo, X.; et al. Down-regulated GAS6 impairs synovial macrophage efferocytosis andpromotes obesity-associated osteoarthritis. Elife 2023, 12, e83069. [Google Scholar] [CrossRef]
- Sun, A.R.; Panchal, S.K.; Friis, T.; Sekar, S.; Crawford, R.; Brown, L.; Xiao, Y.; Prasadam, I. Obesity-associated metabolic syndrome spontaneously induces infiltration of pro-inflammatory macrophage in synovium and promotes osteoarthritis. PLoS ONE 2017, 12, e0183693. [Google Scholar] [CrossRef]
- Warmink, K.; Kozijn, A.E.; Bobeldijk, I.; Stoop, R.; Weinans, H.; Korthagen, N.M. High-fat feeding primes the mouse knee joint to develop osteoarthritis and pathologic infrapatellar fat pad changes after surgically induced injury. Osteoarthr. Cartil. 2020, 28, 593–602. [Google Scholar] [CrossRef]
- Nedunchezhiyan, U.; Varughese, I.; Sun, A.R.; Wu, X.; Crawford, R.; Prasadam, I. Obesity, Inflammation, and Immune System in Osteoarthritis. Front. Immunol. 2022, 13, 907750. [Google Scholar] [CrossRef]
- Jasinski-Bergner, S.; Radetzki, A.-L.; Jahn, J.; Wohlrab, D.; Kielstein, H. Impact of the body mass index on perioperative immunological disturbances in patients with hip and knee arthroplasty. J. Orthop. Surg. Res. 2017, 12, 58. [Google Scholar] [CrossRef]
- Rosenberg, J.H.; Rai, V.; Dilisio, M.F.; Sekundiak, T.D.; Agrawal, D.K. Increased expression of damage-associated molecular patterns (DAMPs) in osteoarthritis of human knee joint compared to hip joint. Mol. Cell Biochem. 2017, 436, 59–69. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.; Harte, A.L.; Aletrari, M.O.; Daghri, N.M.; Al Disi, D.; Tripathi, G.; McTernan, P.G. Does endotoxaemia contribute to osteoarthritis in obese patients? Clin. Sci. 2012, 123, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.A.; Holscher, H.D. Microbiome-Mediated Effects of the Mediterranean Diet on Inflammation. Adv. Nutr. 2018, 9, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Janczy, A.; Szymanski, M.; Stankiewicz, M.; Kaska, L.; Waleron, K.; Stelmanska, E.; Sledzinski, T.; Mika, A. Increased Amount of Polyunsaturated Fatty Acids in the Intestinal Contents of Patients with Morbid Obesity. Obes. Surg. 2023, 33, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Szychlinska, M.A.; Di Rosa, M.; Castorina, A.; Mobasheri, A.; Musumeci, G. A correlation between intestinal microbiota dysbiosis and osteoarthritis. Heliyon 2019, 5, e01134. [Google Scholar] [CrossRef] [PubMed]
- Schott, E.M.; Farnsworth, C.W.; Grier, A.; Lillis, J.A.; Soniwala, S.; Dadourian, G.H.; Bell, R.D.; Doolittle, M.L.; Villani, D.A.; Awad, H.; et al. Targeting the gut microbiome to treat the osteoarthritis of obesity. JCI Insight 2018, 3, e95997. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ding, W.; Wang, H.; Dai, L.; Zong, W.; Wang, Y.; Bi, J.; Han, W.; Dong, G. Gut microbiota and obesity-associated osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1257–1265. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Kim, K.-A.; Gu, W.; Lee, I.-A.; Joh, E.-H.; Kim, D.-H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Loef, M.; Schoones, J.W.; Kloppenburg, M.; Ioan-Facsinay, A. Fatty acids and osteoarthritis: Different types, different effects. Jt. Bone Spine 2019, 86, 451–458. [Google Scholar] [CrossRef]
- Van de Vyver, A.; Clockaerts, S.; van de Lest, C.H.A.; Wei, W.; Verhaar, J.; Van Osch, G.J.V.M.; Bastiaansen-Jenniskens, Y.M. Synovial Fluid Fatty Acid Profiles Differ between Osteoarthritis and Healthy Patients. Cartilage 2020, 11, 473–478. [Google Scholar] [CrossRef]
- Guillou, H.; Zadravec, D.; Martin, P.G.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef]
- Hughes, C.L.; Dhiman, T.R. Dietary compounds in relation to dietary diversity and human health. J. Med. Food 2002, 5, 51–68. [Google Scholar] [CrossRef]
- Das, U.N. “Cell Membrane Theory of Senescence” and the Role of Bioactive Lipids in Aging, and Aging Associated Diseases and Their Therapeutic Implications. Biomolecules 2021, 11, 241. [Google Scholar] [CrossRef]
- Blake, D.R.; Robson, P.; Ho, M.; Jubb, R.W.; McCabe, C.S. Preliminary assessment of the efficacy, tolerability and safety of a cannabis-based medicine (Sativex) in the treatment of pain caused by rheumatoid arthritis. Rheumatology 2006, 45, 50–52. [Google Scholar] [CrossRef]
- Philpott, H.T.; O'Brien, M.; McDougall, J.J. Attenuation of early phase inflammation by cannabidiol prevents pain and nerve damage in rat osteoarthritis. Pain 2017, 158, 2442–2451. [Google Scholar] [CrossRef]
- Tan, L.; Harper, L.R.; Armstrong, A.; Carlson, C.S.; Yammani, R.R. Dietary saturated fatty acid palmitate promotes cartilage lesions and activates the unfolded protein response pathway in mouse knee joints. PLoS ONE 2021, 16, e0247237. [Google Scholar] [CrossRef]
- Alvarez-Garcia, O.; Rogers, N.H.; Smith, R.G.; Lotz, M.K. Palmitate has proapoptotic and proinflammatory effects on articular cartilage and synergizes with interleukin-1. Arthritis Rheumatol. 2014, 66, 1779–1788. [Google Scholar] [CrossRef]
- Miao, H.; Chen, L.; Hao, L.; Zhang, X.; Chen, Y.; Ruan, Z.; Liang, H. Stearic acid induces proinflammatory cytokine production partly through activation of lactate-HIF1 alpha pathway in chondrocytes. Sci. Rep. 2015, 5, 13092. [Google Scholar] [CrossRef]
- Pillon, N.J.; Chan, K.L.; Zhang, S.; Mejdani, M.; Jacobson, M.R.; Ducos, A.; Bilan, P.J.; Niu, W.; Klip, A. Saturated fatty acids activate caspase-4/5 in human monocytes, triggering IL-1beta and IL-18 release. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E825–E835. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, H.; Qi, W.; Zhang, Y.; Li, J.; Li, Z.; Lin, Y.; Bai, X.; Liu, X.; Chen, X.; et al. Nicotine promotes atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis. Cell Death Dis. 2018, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, C.B.; Hwang, S.H.; Yang, J.; Wan, D.; Wagner, K.; Morisseau, C.; Li, D.; Schmidt, W.K.; Hammock, B.D. Pharmaceutical Effects of Inhibiting the Soluble Epoxide Hydrolase in Canine Osteoarthritis. Front. Pharmacol. 2019, 10, 533. [Google Scholar] [CrossRef] [PubMed]
- Adkins, Y.; Soulika, A.M.; Mackey, B.; Kelley, D.S. Docosahexaenoic acid (22:6n-3) Ameliorated the Onset and Severity of Experimental Autoimmune Encephalomyelitis in Mice. Lipids 2019, 54, 13–23. [Google Scholar] [CrossRef]
- Navarini, L.; Afeltra, A.; Gallo Afflitto, G.; Margiotta, D.P.E. Polyunsaturated fatty acids: Any role in rheumatoid arthritis? Lipids Health Dis. 2017, 16, 197. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, A.; Ma, L.; Yu, H.; Zhang, L.; Meng, H.; Cui, Y.; Yu, F.; Yang, B. Docosahexenoic acid treatment ameliorates cartilage degeneration via a p38 MAPK-dependent mechanism. Int. J. Mol. Med. 2016, 37, 1542–1550. [Google Scholar] [CrossRef]
- Flower, R.J.; Perretti, M. Controlling inflammation: A fat chance? J. Exp. Med. 2005, 201, 671–674. [Google Scholar] [CrossRef]
- Caron, J.P.; Gandy, J.C.; Brown, J.L.; Sordillo, L.M. Omega-3 fatty acids and docosahexaenoic acid oxymetabolites modulate the inflammatory response of equine recombinant interleukin1beta-stimulated equine synoviocytes. Prostaglandins Other Lipid Mediat. 2019, 142, 1–8. [Google Scholar] [CrossRef]
- Bartolotti, I.; Roseti, L.; Petretta, M.; Grigolo, B.; Desando, G. A Roadmap of In Vitro Models in Osteoarthritis: A Focus on Their Biological Relevance in Regenerative Medicine. J. Clin. Med. 2021, 10, 1920. [Google Scholar] [CrossRef]
- Bednarczyk, E. Chondrocytes In Vitro Systems Allowing Study of OA. Int. J. Mol. Sci. 2022, 23, 10308. [Google Scholar] [CrossRef]
- Feng, L.; Yang, Z.; Li, Y.; Hou, N.; Yang, B.; Lu, X.; Bai, S.; Wang, M.; Zhang, X.; Wang, H.; et al. Malat1 attenuated the rescuing effects of docosahexaenoic acid on osteoarthritis treatment via repressing its chondroprotective and chondrogenesis activities. Biomed. Pharmacother. 2022, 154, 113608. [Google Scholar] [CrossRef]
- Zainal, Z.; Longman, A.; Hurst, S.; Duggan, K.; Caterson, B.; Hughes, C.; Harwood, J. Relative efficacies of omega-3 polyunsaturated fatty acids in reducing expression of key proteins in a model system for studying osteoarthritis. Osteoarthr. Cartil. 2009, 17, 896–905. [Google Scholar] [CrossRef]
- Wann, A.K.; Mistry, J.; Blain, E.J.; Michael-Titus, A.T.; Knight, M.M. Eicosapentaenoic acid and docosahexaenoic acid reduce interleukin-1beta-mediated cartilage degradation. Arthritis Res. Ther. 2010, 12, R207. [Google Scholar] [CrossRef]
- Sakata, S.; Hayashi, S.; Fujishiro, T.; Kawakita, K.; Kanzaki, N.; Hashimoto, S.; Iwasa, K.; Chinzei, N.; Kihara, S.; Haneda, M.; et al. Oxidative stress-induced apoptosis and matrix loss of chondrocytes is inhibited by eicosapentaenoic acid. J. Orthop. Res. 2015, 33, 359–365. [Google Scholar] [CrossRef]
- Jhun, J.; Na, H.S.; Cho, K.-H.; Kim, J.; Moon, Y.-M.; Lee, S.Y.; Lee, J.S.; Lee, A.R.; Kim, S.J.; Cho, M.-L.; et al. A green-lipped mussel reduces pain behavior and chondrocyte inflammation and attenuated experimental osteoarthritis progression. PLoS ONE 2021, 16, e0259130. [Google Scholar] [CrossRef]
- Jeon, J.; Noh, H.-J.; Lee, H.; Park, H.-H.; Ha, Y.-J.; Park, S.H.; Lee, H.; Kim, S.-J.; Kang, H.C.; Eyun, S.-I.; et al. TRIM24-RIP3 axis perturbation accelerates osteoarthritis pathogenesis. Ann. Rheum. Dis. 2020, 79, 1635–1643. [Google Scholar] [CrossRef]
- Benabdoune, H.; Rondon, E.-P.; Shi, Q.; Fernandes, J.; Ranger, P.; Fahmi, H.; Benderdour, M. The role of resolvin D1 in the regulation of inflammatory and catabolic mediators in osteoarthritis. Inflamm. Res. 2016, 65, 635–645. [Google Scholar] [CrossRef]
- Piao, S.; Du, W.; Wei, Y.; Yang, Y.; Feng, X.; Bai, L. Protectin DX attenuates IL-1beta-induced inflammation via the AMPK/NF-kappaB pathway in chondrocytes and ameliorates osteoarthritis progression in a rat model. Int. Immunopharmacol. 2020, 78, 106043. [Google Scholar] [CrossRef]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef]
- Adler, N.; Schoeniger, A.; Fuhrmann, H. Polyunsaturated fatty acids influence inflammatory markers in a cellular model for canine osteoarthritis. J. Anim. Physiol. Anim. Nutr. 2018, 102, e623–e632. [Google Scholar] [CrossRef]
- Yu, H.; Li, Y.; Ma, L.; Meng, H.; Bai, X.; Fan, Z.; Yu, F.; Guo, A. A low ratio of n-6/n-3 polyunsaturated fatty acids suppresses matrix metalloproteinase 13 expression and reduces adjuvant-induced arthritis in rats. Nutr. Res. 2015, 35, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Sueda, Y.; Okazaki, R.; Funaki, Y.; Hasegawa, Y.; Ishikawa, H.; Hirayama, Y.; Inui, G.; Harada, T.; Takata, M.; Morita, M.; et al. Specialized Pro-Resolving Mediators Do Not Inhibit the Synthesis of Inflammatory Mediators Induced by Tumor Necrosis Factor-alpha in Synovial Fibroblasts. Yonago Acta Med. 2022, 65, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Jiang, W.; Wang, X.; Du, S.; Qi, J.; Jia, Q.; Song, H. Resolvin D1 inhibits the proliferation of osteoarthritis fibroblast-like synoviocytes through the Hippo-YAP signaling pathway. BMC Musculoskelet. Disord. 2022, 23, 149. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Feng, X.; Zhang, H.; Wei, Y.; Yang, Y.; Tian, Y.; Bai, L. Maresin-1 suppresses IL-1beta-induced MMP-13 secretion by activating the PI3K/AKT pathway and inhibiting the NF-kappaB pathway in synovioblasts of an osteoarthritis rat model with treadmill exercise. Connect. Tissue Res. 2021, 62, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Li, S.-J.; Liu, R.; Zhan, J.-F.; Tan, C.; Fang, Y.-F.; Chen, Y.; Yu, B. Inhibition of YAP with siRNA prevents cartilage degradation and ameliorates osteoarthritis development. J. Mol. Med. 2019, 97, 103–114. [Google Scholar] [CrossRef]
- Liu, X.; Liu, L.; Zhao, J.; Wang, H.; Li, Y. Mechanotransduction regulates inflammation responses of epicardial adipocytes in cardiovascular diseases. Front. Endocrinol. 2022, 13, 1080383. [Google Scholar] [CrossRef]
- de Bus, I.; van Krimpen, S.; Hooiveld, G.J.; Boekschoten, M.V.; Poland, M.; Witkamp, R.F.; Albada, B.; Balvers, M.G.J. Immunomodulating effects of 13- and 16-hydroxylated docosahexaenoyl ethanolamide in LPS stimulated RAW264.7 macrophages. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158908. [Google Scholar] [CrossRef]
- Dalli, J.; Zhu, M.; Vlasenko, N.A.; Deng, B.; Haeggström, J.Z.; Petasis, N.A.; Serhan, C.N. The novel 13S,14S-epoxy-maresin is converted by human macrophages to maresin 1 (MaR1), inhibits leukotriene A(4) hydrolase (LTA(4)H), and shifts macrophage phenotype. FASEB J. 2013, 27, 2573–2583. [Google Scholar] [CrossRef]
- Liu, Y.; Kongsuphol, P.; Chiam, S.Y.; Zhang, Q.X.; Gourikutty, S.B.N.; Saha, S.; Biswas, S.K.; Ramadan, Q. Adipose-on-a-chip: A dynamic microphysiological in vitro model of the human adipose for immune-metabolic analysis in type II diabetes. Lab Chip 2019, 19, 241–253. [Google Scholar] [CrossRef]
- Saini, A.; Sharples, A.P.; Al-Shanti, N.; Stewart, C.E. Omega-3 fatty acid EPA improves regenerative capacity of mouse skeletal muscle cells exposed to saturated fat and inflammation. Biogerontology 2017, 18, 109–129. [Google Scholar] [CrossRef]
- Shen, C.-L.; Dunn, D.M.; Henry, J.H.; Li, Y.; Watkins, B.A. Decreased production of inflammatory mediators in human osteoarthritic chondrocytes by conjugated linoleic acids. Lipids 2004, 39, 161–166. [Google Scholar] [CrossRef]
- Mustonen, A.M.; Kakela, R.; Finnila, M.A.J.; Sawatsky, A.; Korhonen, R.K.; Saarakkala, S.; Herzog, W.; Paakkonen, T.; Nieminen, P. Anterior cruciate ligament transection alters the n-3/n-6 fatty acid balance in the lapine infrapatellar fat pad. Lipids Health Dis. 2019, 18, 67. [Google Scholar] [CrossRef]
- Suleiman, J.B.; Mohamed, M.; Bakar, A.B.A. A systematic review on different models of inducing obesity in animals: Advantages and limitations. J. Adv. Vet. Anim. Res. 2020, 7, 103–114. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, L.; Yan, Z.; Li, Z.; Fu, M.; Xue, C.; Wang, J. A low proportion n-6/n-3 PUFA diet supplemented with Antarctic krill (Euphausia superba) oil protects against osteoarthritis by attenuating inflammation in ovariectomized mice. Food Funct. 2021, 12, 6766–6779. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liao, J.K. A mouse model of diet-induced obesity and insulin resistance. Methods Mol. Biol. 2012, 821, 421–433. [Google Scholar]
- Buettner, R.; Scholmerich, J.; Bollheimer, L.C. High-fat diets: Modeling the metabolic disorders of human obesity in rodents. Obesity 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Griffin, T.M.; Huebner, J.L.; Kraus, V.B.; Yan, Z.; Guilak, F. Induction of osteoarthritis and metabolic inflammation by a very high-fat diet in mice: Effects of short-term exercise. Arthritis Rheum. 2012, 64, 443–453. [Google Scholar] [CrossRef]
- Sun, A.R.; Wu, X.; Liu, B.; Chen, Y.; Armitage, C.W.; Kollipara, A.; Crawford, R.; Beagley, K.W.; Mao, X.; Xiao, Y.; et al. Pro-resolving lipid mediator ameliorates obesity induced osteoarthritis by regulating synovial macrophage polarisation. Sci. Rep. 2019, 9, 426. [Google Scholar] [CrossRef]
- Hahn, A.; Batushansky, A.; Rawle, R.; Lopes, E.P.; June, R.; Griffin, T. Effects of long-term exercise and a high-fat diet on synovial fluid metabolomics and joint structural phenotypes in mice: An integrated network analysis. Osteoarthr. Cartil. 2021, 29, 1549–1563. [Google Scholar] [CrossRef]
- Portela, N.D.; Galván, C.; Sanmarco, L.M.; Bergero, G.; Aoki, M.P.; Cano, R.C.; Pesoa, S.A. Omega-3-Supplemented Fat Diet Drives Immune Metabolic Response in Visceral Adipose Tissue by Modulating Gut Microbiota in a Mouse Model of Obesity. Nutrients 2023, 15, 1404. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-L.; Jain, D.; McNeill, J.N.; Little, D.; Anderson, J.A.; Huebner, J.L.; Kraus, V.B.; Rodriguiz, R.M.; Wetsel, W.C.; Guilak, F. Dietary fatty acid content regulates wound repair and the pathogenesis of osteoarthritis following joint injury. Ann. Rheum. Dis. 2015, 74, 2076–2083. [Google Scholar] [CrossRef] [PubMed]
- Kimmerling, K.A.; Oswald, S.J.; Huebner, J.L.; Little, D.; Kraus, V.B.; Kang, J.X.; Wu, C.L.; Guilak, F. Transgenic conversion of omega-6 to omega-3 polyunsaturated fatty acids via fat-1 reduces the severity of post-traumatic osteoarthritis. Arthritis Res. Ther. 2020, 22, 83. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.J.; Wang, L.; Jin, D.D.; Zhang, Z.M.; Chen, T.Y.; Jia, C.H.; Wang, Y.; Zhen, X.C.; Huang, B.; Yan, B.; et al. Enhancement of the synthesis of n-3 PUFAs in fat-1 transgenic mice inhibits mTORC1 signalling and delays surgically induced osteoarthritis in comparison with wild-type mice. Ann. Rheum. Dis. 2014, 73, 1719–1727. [Google Scholar] [CrossRef]
- Cai, A.; Hutchison, E.; Hudson, J.; Kawashima, Y.; Komori, N.; Singh, A.; Brush, R.; Anderson, R.; Sonntag, W.; Matsumoto, H.; et al. Metabolic enrichment of omega-3 polyunsaturated fatty acids does not reduce the onset of idiopathic knee osteoarthritis in mice. Osteoarthr. Cartil. 2014, 22, 1301–1309. [Google Scholar] [CrossRef]
- Griffin, T.M.; Huebner, J.L.; Kraus, V.B.; Guilak, F. Extreme obesity due to impaired leptin signaling in mice does not cause knee osteoarthritis. Arthritis Rheum. 2009, 60, 2935–2944. [Google Scholar] [CrossRef]
- Pinel, A.; Pitois, E.; Rigaudiere, J.-P.; Jouve, C.; De Saint-Vincent, S.; Laillet, B.; Montaurier, C.; Huertas, A.; Morio, B.; Capel, F. EPA prevents fat mass expansion and metabolic disturbances in mice fed with a Western diet. J. Lipid Res. 2016, 57, 1382–1397. [Google Scholar] [CrossRef]
- Knott, L.; Avery, N.C.; Hollander, A.P.; Tarlton, J.F. Regulation of osteoarthritis by omega-3 (n-3) polyunsaturated fatty acids in a naturally occurring model of disease. Osteoarthr. Cartil. 2011, 19, 1150–1157. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, W.; Zhong, Z.; Yu, H.; Zhang, P.; Shen, H. Docosahexaenoic acid inhibits bone remodeling and vessel formation in the osteochondral unit in a rat model. Biomed. Pharmacother. 2019, 114, 108811. [Google Scholar] [CrossRef]
- Mehler, S.J.; May, L.R.; King, C.; Harris, W.S.; Shah, Z. A prospective, randomized, double blind, placebo-controlled evaluation of the effects of eicosapentaenoic acid and docosahexaenoic acid on the clinical signs and erythrocyte membrane polyunsaturated fatty acid concentrations in dogs with osteoarthritis. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 1–7. [Google Scholar] [CrossRef]
- Christmann, U.; Hancock, C.L.; Poole, C.M.; Emery, A.L.; Poovey, J.R.; Hagg, C.; Mattson, E.A.; Scarborough, J.J.; Christopher, J.S.; Dixon, A.T.; et al. Dynamics of DHA and EPA supplementation: Incorporation into equine plasma, synovial fluid, and surfactant glycerophosphocholines. Metabolomics 2021, 17, 41. [Google Scholar] [CrossRef]
- Huang, J.; Burston, J.J.; Li, L.; Ashraf, S.; Mapp, P.I.; Bennett, A.J.; Ravipati, S.; Pousinis, P.; Barrett, D.A.; Scammell, B.E.; et al. Targeting the D Series Resolvin Receptor System for the Treatment of Osteoarthritis Pain. Arthritis Rheumatol. 2017, 69, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Meesawatsom, P.; Burston, J.; Hathway, G.; Bennett, A.; Chapman, V. Inhibitory effects of aspirin-triggered resolvin D1 on spinal nociceptive processing in rat pain models. J. Neuroinflammation 2016, 13, 233. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.C.; Orlando, R.M.; Turchetti-Maia, R.M.; de Francischi, J.N. Comparative effects of the omega3 polyunsaturated fatty acid derivatives resolvins E1 and D1 and protectin DX in models of inflammation and pain. J. Inflamm. Res. 2017, 10, 119–133. [Google Scholar] [CrossRef]
- Fattori, V.; Zaninelli, T.H.; Ferraz, C.R.; Brasil-Silva, L.; Borghi, S.M.; Cunha, J.M.; Chichorro, J.G.; Casagrande, R.; Verri, W.A. Maresin 2 is an analgesic specialized pro-resolution lipid mediator in mice by inhibiting neutrophil and monocyte recruitment, nociceptor neuron TRPV1 and TRPA1 activation, and CGRP release. Neuropharmacology 2022, 216, 109189. [Google Scholar] [CrossRef]
- Zhang, T.; Dai, Y.; Zhang, L.; Tian, Y.; Li, Z.; Wang, J. Effects of Edible Oils with Different n-6/n-3 PUFA Ratios on Articular Cartilage Degeneration via Regulating the NF-kappaB Signaling Pathway. J. Agric. Food Chem. 2020, 68, 12641–12650. [Google Scholar] [CrossRef]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef]
- Kang, J.X. Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 263–267. [Google Scholar] [CrossRef]
- Li, J.; Li, F.R.; Wei, D.; Jia, W.; Kang, J.X.; Stefanovic-Racic, M.; Dai, Y.; Zhao, A.Z. Endogenous omega-3 polyunsaturated fatty acid production confers resistance to obesity, dyslipidemia, and diabetes in mice. Mol. Endocrinol. 2014, 28, 1316–1328. [Google Scholar] [CrossRef]
- Suriano, F.; Vieira-Silva, S.; Falony, G.; Roumain, M.; Paquot, A.; Pelicaen, R.; Régnier, M.; Delzenne, N.M.; Raes, J.; Muccioli, G.G.; et al. Novel insights into the genetically obese (ob/ob) and diabetic (db/db) mice: Two sides of the same coin. Microbiome 2021, 9, 147. [Google Scholar] [CrossRef]
- Gotoh, N.; Nagao, K.; Ishida, H.; Nakamitsu, K.; Yoshinaga, K.; Nagai, T.; Beppu, F.; Yoshinaga-Kiriake, A.; Watanabe, H.; Yanagita, T. Metabolism of Natural Highly Unsaturated Fatty Acid, Tetracosahexaenoic Acid (24:6n-3), in C57BL/KsJ-db/db Mice. J. Oleo Sci. 2018, 67, 1597–1607. [Google Scholar] [CrossRef]
- Nakamoto, K.; Nishinaka, T.; Mankura, M.; Fujita-Hamabe, W.; Tokuyama, S. Antinociceptive effects of docosahexaenoic acid against various pain stimuli in mice. Biol. Pharm. Bull. 2010, 33, 1070–1072. [Google Scholar] [CrossRef] [PubMed]
- Bliddal, H.; Leeds, A.R.; Christensen, R. Osteoarthritis, obesity and weight loss: Evidence, hypotheses and horizons—A scoping review. Obes. Rev. 2014, 15, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Courties, A.; Sellam, J.; Berenbaum, F. Metabolic syndrome-associated osteoarthritis. Curr. Opin. Rheumatol. 2017, 29, 214–222. [Google Scholar] [CrossRef]
- Kuszewski, J.C.; Wong, R.H.X.; Howe, P.R.C. Fish oil supplementation reduces osteoarthritis-specific pain in older adults with overweight/obesity. Rheumatol. Adv. Pract. 2020, 4, rkaa036. [Google Scholar] [CrossRef]
- Baker, K.; Matthan, N.; Lichtenstein, A.; Niu, J.; Guermazi, A.; Roemer, F.; Grainger, A.; Nevitt, M.; Clancy, M.; Lewis, C.; et al. Association of plasma n-6 and n-3 polyunsaturated fatty acids with synovitis in the knee: The MOST study. Osteoarthr. Cartil. 2012, 20, 382–387. [Google Scholar] [CrossRef]
- Stammers, T.; Sibbald, B.; Freeling, P. Fish oil in osteoarthritis. Lancet 1989, 2, 503. [Google Scholar] [CrossRef]
- Stammers, T.; Sibbald, B.; Freeling, P. Efficacy of cod liver oil as an adjunct to non-steroidal anti-inflammatory drug treatment in the management of osteoarthritis in general practice. Ann. Rheum. Dis. 1992, 51, 128–129. [Google Scholar] [CrossRef]
- Stebbings, S.; Gray, A.; Schneiders, A.G.; Sansom, A. A randomized double-blind placebo-controlled trial to investigate the effectiveness and safety of a novel green-lipped mussel extract -BioLex(R) -for managing pain in moderate to severe osteoarthritis of the hip and knee. BMC Complement. Altern. Med. 2017, 17, 416. [Google Scholar] [CrossRef]
- Lau, C.S.; Chiu PK, Y.; Chu EM, Y.; Cheng IY, W.; Tang, W.M.; Man RY, K.; Halpern, G.M. Treatment of knee osteoarthritis with Lyprinol®, lipid extract of the green-lipped mussel—A double-blind placebo-controlled study. Prog. Nutr. 2004, 6, 17–31. [Google Scholar]
- McPhee, S.; Hodges, L.; Wright, P.; Wynne, P.; Kalafatis, N.; Harney, D.; Macrides, T. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 346–356. [Google Scholar] [CrossRef]
- Wakimoto, T.; Kondo, H.; Nii, H.; Kimura, K.; Egami, Y.; Oka, Y.; Yoshida, M.; Kida, E.; Ye, Y.; Akahoshi, S.; et al. Furan fatty acid as an anti-inflammatory component from the green-lipped mussel Perna canaliculus. Proc. Natl. Acad. Sci. USA 2011, 108, 17533–17537. [Google Scholar] [CrossRef] [PubMed]
- Coulson, S.; Butt, H.; Vecchio, P.; Gramotnev, H.; Vitetta, L. Green-lipped mussel extract (Perna canaliculus) and glucosamine sulphate in patients with knee osteoarthritis: Therapeutic efficacy and effects on gastrointestinal microbiota profiles. Inflammopharmacology 2013, 21, 79–90. [Google Scholar] [CrossRef]
- Jacquet, A.; Girodet, P.O.; Pariente, A.; Forest, K.; Mallet, L.; Moore, N. Phytalgic, a food supplement, vs placebo in patients with osteoarthritis of the knee or hip: A randomised double-blind placebo-controlled clinical trial. Arthritis Res. Ther. 2009, 11, R192. [Google Scholar] [CrossRef]
- Hill, C.L.; March, L.M.; Aitken, D.; Lester, S.E.; Battersby, R.; Hynes, K.; Fedorova, T.; Proudman, S.M.; James, M.; Cleland, L.G.; et al. Fish oil in knee osteoarthritis: A randomised clinical trial of low dose versus high dose. Ann. Rheum. Dis. 2016, 75, 23–29. [Google Scholar] [CrossRef]
- Deutsch, L. Evaluation of the effect of Neptune Krill Oil on chronic inflammation and arthritic symptoms. J. Am. Coll. Nutr. 2007, 26, 39–48. [Google Scholar] [CrossRef]
- Suzuki, Y.; Fukushima, M.; Sakuraba, K.; Sawaki, K.; Sekigawa, K. Krill Oil Improves Mild Knee Joint Pain: A Randomized Control Trial. PLoS ONE 2016, 11, e0162769. [Google Scholar] [CrossRef]
- Mohammad, N.S.; Nazli, R.; Zafar, H.; Fatima, S. Effects of lipid based Multiple Micronutrients Supplement on the birth outcome of underweight pre-eclamptic women: A randomized clinical trial. Pak. J. Med. Sci. 2022, 38, 219–226. [Google Scholar]
- Stonehouse, W.; Benassi-Evans, B.; Bednarz, J.; Vincent, A.D.; Hall, S.; Hill, C.L. Krill oil improved osteoarthritic knee pain in adults with mild to moderate knee osteoarthritis: A 6-month multicenter, randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2022, 116, 672–685. [Google Scholar] [CrossRef]
- Gruenwald, J.; Petzold, E.; Busch, R.; Petzold, H.-P.; Graubaum, H.-J. Effect of glucosamine sulfate with or without omega-3 fatty acids in patients with osteoarthritis. Adv. Ther. 2009, 26, 858–871. [Google Scholar] [CrossRef]
- Sasahara, I.; Yamamoto, A.; Takeshita, M.; Suga, Y.; Suzuki, K.; Nishikata, N.; Takada, M.; Hashimoto, M.; Mine, T.; Kobuna, Y.; et al. l-Serine and EPA Relieve Chronic Low-Back and Knee Pain in Adults: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Nutr. 2020, 150, 2278–2286. [Google Scholar] [CrossRef]
- Valdes, A.M.; Ravipati, S.; Menni, C.; Abhishek, A.; Metrustry, S.; Harris, J.; Nessa, A.; Williams, F.M.K.; Spector, T.D.; Doherty, M.; et al. Association of the resolvin precursor 17-HDHA, but not D- or E-series resolvins, with heat pain sensitivity and osteoarthritis pain in humans. Sci. Rep. 2017, 7, 10748. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Driban, J.B.; Xu, C.; Lapane, K.L.; McAlindon, T.E.; Eaton, C.B. Dietary Fat Intake and Radiographic Progression of Knee Osteoarthritis: Data From the Osteoarthritis Initiative. Arthritis Care Res. 2017, 69, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Garvey, W.T.; Mechanick, J.I.; Brett, E.M.; Garber, A.J.; Hurley, D.L.; Jastreboff, A.M.; Nadolsky, K.; Pessah-Pollack, R.; Plodkowski, R.; Garvey, W.T.; et al. American Association of Clinical Endocrinologists and American College of Endocrinology Comprehensive Clinical Practice Guidelines for Medical Care of Patients with Obesity. Endocr. Prac. 2016, 22, 842–884. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.; Bartels, E.M.; Astrup, A.; Bliddal, H. Effect of weight reduction in obese patients diagnosed with knee osteoarthritis: A systematic review and meta-analysis. Ann. Rheum. Dis. 2007, 66, 433–439. [Google Scholar] [CrossRef]
- Basedow, M.; Runciman, W.B.; March, L.; Esterman, A. Australians with osteoarthritis; the use of and beliefs about complementary and alternative medicines. Complement. Ther. Clin. Pract. 2014, 20, 237–242. [Google Scholar] [CrossRef]
- Messina, O.D.; Vidal Wilman, M.; Vidal Neira, L.F. Nutrition, osteoarthritis and cartilage metabolism. Aging Clin. Exp. Res. 2019, 31, 807–813. [Google Scholar] [CrossRef]
- Lopez, H.L. Nutritional interventions to prevent and treat osteoarthritis. Part II: Focus on micronutrients and supportive nutraceuticals. PMR 2012, 4, S155–S168. [Google Scholar] [CrossRef]
- Liu, X.; Machado, G.C.; Eyles, J.P.; Ravi, V.; Hunter, D.J. Dietary supplements for treating osteoarthritis: A systematic review and meta-analysis. Br. J. Sport. Med. 2018, 52, 167–175. [Google Scholar] [CrossRef]
- Senftleber, N.K.; Nielsen, S.M.; Andersen, J.R.; Bliddal, H.; Tarp, S.; Lauritzen, L.; Furst, D.E.; Suarez-Almazor, M.E.; Lyddiatt, A.; Christensen, R. Marine Oil Supplements for Arthritis Pain: A Systematic Review and Meta-Analysis of Randomized Trials. Nutrients 2017, 9, 42. [Google Scholar] [CrossRef]
- Veselinovic, M.; Vasiljevic, D.; Vucic, V.; Arsic, A.; Petrovic, S.; Tomic-Lucic, A.; Savic, M.; Zivanovic, S.; Stojic, V.; Jakovljevic, V. Clinical Benefits of n-3 PUFA and ɤ-Linolenic Acid in Patients with Rheumatoid Arthritis. Nutrients 2017, 9, 325. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Hoteit, M.; Zoghbi, E.; Rady, A.; Shankiti, I.; Al-Jawaldeh, A. Fatty Acids Quality in Middle Eastern Traditional Dishes, Arabic Sweets and Market Foods Frequently Consumed in Lebanon. Nutrients 2021, 13, 2462. [Google Scholar] [CrossRef]
- Harika, R.K.; Eilander, A.; Alssema, M.; Osendarp, S.J.; Zock, P.L. Intake of fatty acids in general populations worldwide does not meet dietary recommendations to prevent coronary heart disease: A systematic review of data from 40 countries. Ann. Nutr. Metab. 2013, 63, 229–238. [Google Scholar] [CrossRef]
- Liu, T.; Zhao, Y.; Wang, X.; Li, X.; Yan, Y. A novel oriented immobilized lipase on magnetic nanoparticles in reverse micelles system and its application in the enrichment of polyunsaturated fatty acids. Bioresour. Technol. 2013, 132, 99–102. [Google Scholar] [CrossRef]
- Santos, H.O.; Price, J.C.; Bueno, A.A. Beyond Fish Oil Supplementation: The Effects of Alternative Plant Sources of Omega-3 Polyunsaturated Fatty Acids upon Lipid Indexes and Cardiometabolic Biomarkers-An Overview. Nutrients 2020, 12, 3159. [Google Scholar] [CrossRef]
- Alsharari, Z.D.; Leander, K.; Sjögren, P.; Carlsson, A.; Cederholm, T.; de Faire, U.; Hellenius, M.-L.; Marklund, M.; Risérus, U. Association between carbohydrate intake and fatty acids in the de novo lipogenic pathway in serum phospholipids and adipose tissue in a population of Swedish men. Eur. J. Nutr. 2020, 59, 2089–2097. [Google Scholar] [CrossRef]
- Liu, K.; Wang, B.; Zhou, R.; Lang, H.D.; Ran, L.; Wang, J.; Li, L.; Kang, C.; Zhu, X.H.; Zhang, Q.Y.; et al. Effect of combined use of a low-carbohydrate, high-protein diet with omega-3 polyunsaturated fatty acid supplementation on glycemic control in newly diagnosed type 2 diabetes: A randomized, double-blind, parallel-controlled trial. Am. J. Clin. Nutr. 2018, 108, 256–265. [Google Scholar] [CrossRef]
- Pesta, D.H.; Samuel, V.T. A high-protein diet for reducing body fat: Mechanisms and possible caveats. Nutr. Metab. 2014, 11, 53. [Google Scholar] [CrossRef]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke: From the American Association of Neurological Surgeons (AANS), American Society of Neuroradiology (ASNR), Cardiovascular and Interventional Radiology Society of Europe (CIRSE), Canadian Interventional Radiology Association (CIRA), Congress of Neurological Surgeons (CNS), European Society of Minimally Invasive Neurological Therapy (ESMINT), European Society of Neuroradiology (ESNR), European Stroke Organization (ESO), Society for Cardiovascular Angiography and Interventions (SCAI), Society of Interventional Radiology (SIR), Society of NeuroInterventional Surgery (SNIS), and World Stroke Organization (WSO). J. Vasc. Interv. Radiol. 2018, 29, 441–453. [Google Scholar] [PubMed]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Eseberri, I.; Trepiana, J.; Léniz, A.; Gómez-García, I.; Carr-Ugarte, H.; González, M.; Portillo, M.P. Variability in the Beneficial Effects of Phenolic Compounds: A Review. Nutrients 2022, 14, 1925. [Google Scholar] [CrossRef] [PubMed]
- Cormier, H.; Rudkowska, I.; Paradis, A.-M.; Thifault, E.; Garneau, V.; Lemieux, S.; Couture, P.; Vohl, M.-C. Association between polymorphisms in the fatty acid desaturase gene cluster and the plasma triacylglycerol response to an n-3 PUFA supplementation. Nutrients 2012, 4, 1026–1041. [Google Scholar] [CrossRef]
- Sparkes, C.; Sinclair, A.J.; Gibson, R.A.; Else, P.L.; Meyer, B.J. High Variability in Erythrocyte, Plasma and Whole Blood EPA and DHA Levels in Response to Supplementation. Nutrients 2020, 12, 1017. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Eckel, R.H.; Stafford, J.M. Sex differences in lipid and lipoprotein metabolism. Mol. Metab. 2018, 15, 45–55. [Google Scholar] [CrossRef]
- Saini, R.K.; Prasad, P.; Sreedhar, R.V.; Akhilender Naidu, K.; Shang, X.; Keum, Y.S. Omega-3 Polyunsaturated Fatty Acids (PUFAs): Emerging Plant and Microbial Sources, Oxidative Stability, Bioavailability, and Health Benefits-A Review. Antioxidants 2021, 10, 1627. [Google Scholar] [CrossRef]
- Costa, L.G. Contaminants in fish: Risk-benefit considerations. Arh. Hig. Rada Toksikol. 2007, 58, 367–374. [Google Scholar] [CrossRef]
- Storelli, M.M.; Stuffler, R.G.; Marcotrigiano, G.O. Total and methylmercury residues in tuna-fish from the Mediterranean sea. Food Addit. Contam. 2002, 19, 715–720. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Adults Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients 2021, 14, 29. [Google Scholar] [CrossRef]
- Burdge, G.C. Metabolism of alpha-linolenic acid in humans. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 161–168. [Google Scholar] [CrossRef]
- Chevalier, L.; Vachon, A.; Plourde, M. Pharmacokinetics of Supplemental Omega-3 Fatty Acids Esterified in Monoglycerides, Ethyl Esters, or Triglycerides in Adults in a Randomized Crossover Trial. J. Nutr. 2021, 151, 1111–1118. [Google Scholar] [CrossRef]
- Shen, Y.; Lu, T.; Liu, X.-Y.; Zhao, M.-T.; Yin, F.-W.; Rakariyatham, K.; Zhou, D.-Y. Improving the oxidative stability and lengthening the shelf life of DHA algae oil with composite antioxidants. Food Chem. 2020, 313, 126139. [Google Scholar] [CrossRef]
- Tou, J.C.; Jaczynski, J.; Chen, Y.C. Krill for human consumption: Nutritional value and potential health benefits. Nutr. Rev. 2007, 65, 63–77. [Google Scholar] [CrossRef]
- Ulven, S.M.; Kirkhus, B.; Lamglait, A.; Basu, S.; Elind, E.; Haider, T.; Berge, K.; Vik, H.; Pedersen, J.I. Metabolic effects of krill oil are essentially similar to those of fish oil but at lower dose of EPA and DHA, in healthy volunteers. Lipids 2011, 46, 37–46. [Google Scholar] [CrossRef]
- Ierna, M.; Kerr, A.; Scales, H.; Berge, K.; Griinari, M. Supplementation of diet with krill oil protects against experimental rheumatoid arthritis. BMC Musculoskelet. Disord. 2010, 11, 136. [Google Scholar] [CrossRef]
- Nain, C.W.; Berdal, G.; Thao, P.T.P.; Mignolet, E.; Buchet, M.; Page, M.; Larondelle, Y. Green Tea Extract Enhances the Oxidative Stability of DHA-Rich Oil. Antioxidants 2021, 10, 982. [Google Scholar] [CrossRef]
- Juric, S.; Juric, M.; Siddique, M.A.; Fathi, M. Vegetable Oils Rich in Polyunsaturated Fatty Acids: Nanoencapsulation Methods and Stability Enhancement. Food Rev. Int. 2022, 38, 32–69. [Google Scholar] [CrossRef]
- Wallace, J.; McCabe, A.; Robson, P.; Keogh, M.; Murray, C.; Kelly, P.; Márquez-Ruiz, G.; McGlynn, H.; Gilmore, W.; Strain, J. Bioavailability of n-3 polyunsaturated fatty acids (PUFA) in foods enriched with microencapsulated fish oil. Ann. Nutr. Metab. 2000, 44, 157–162. [Google Scholar] [CrossRef]
- Helal, N.A.; Eassa, H.A.; Amer, A.M.; Eltokhy, M.A.; Edafiogho, I.; Nounou, M.I. Nutraceuticals' Novel Formulations: The Good, the Bad, the Unknown and Patents Involved. Recent Pat. Drug Deliv. Formul. 2019, 13, 105–156. [Google Scholar] [CrossRef] [PubMed]
- Dravid, A.A.; Dhanabalan, K.M.; Agarwal, S.; Agarwal, R. Resolvin D1-loaded nanoliposomes promote M2 macrophage polarization and are effective in the treatment of osteoarthritis. Bioeng. Transl. Med. 2022, 7, e10281. [Google Scholar] [CrossRef] [PubMed]
- Charles, A.L.; Abdillah, A.A.; Saraswati, Y.R.; Sridhar, K.; Balderamos, C.; Masithah, E.D.; Alamsjah, M.A. Characterization of freeze-dried microencapsulation tuna fish oil with arrowroot starch and maltodextrin. Food Hydrocoll. 2021, 112, 106281. [Google Scholar] [CrossRef]
- Cassano, R.; Mellace, S.; Marrelli, M.; Conforti, F.; Trombino, S. alpha-Tocopheryl linolenate solid lipid nanoparticles for the encapsulation, protection, and release of the omega-3 polyunsaturated fatty acid: In vitro anti-melanoma activity evaluation. Colloids Surf. B Biointerfaces 2017, 151, 128–133. [Google Scholar] [CrossRef]
- Hur, S.J.; Kim, D.H.; Chun, S.C.; Lee, S.K.; Keum, Y.S. Effects of biopolymer encapsulation on trans fatty acid digestibility in an in vitro human digestion system. Food Funct. 2013, 4, 1827–1834. [Google Scholar] [CrossRef]
- Hur, S.J.; Kim, D.H.; Chun, S.C.; Lee, S.K. Effects of dietary conjugated linoleic acid and biopolymer encapsulation on lipid metabolism in mice. Int. J. Mol. Sci. 2013, 14, 6848–6862. [Google Scholar] [CrossRef]
- Torres-Giner, S.; Martinez-Abad, A.; Ocio, M.J.; Lagaron, J.M. Stabilization of a nutraceutical omega-3 fatty acid by encapsulation in ultrathin electrosprayed zein prolamine. J. Food. Sci. 2010, 75, N69–N79. [Google Scholar] [CrossRef]
- Perez, A.A.; Sponton, O.E.; Andermatten, R.B.; Rubiolo, A.C.; Santiago, L.G. Biopolymer nanoparticles designed for polyunsaturated fatty acid vehiculization: Protein-polysaccharide ratio study. Food Chem. 2015, 188, 543–550. [Google Scholar] [CrossRef]
- Zhang, Z.; Decker, E.A.; McClements, D.J. Encapsulation, protection, and release of polyunsaturated lipids using biopolymer-based hydrogel particles. Food Res. Int. 2014, 64, 520–526. [Google Scholar] [CrossRef]
- Han, W.; Chai, X.; Liu, Y.; Xu, Y.; Tan, C.-P. Crystal network structure and stability of beeswax-based oleogels with different polyunsaturated fatty acid oils. Food Chem. 2022, 381, 131745. [Google Scholar] [CrossRef]
- Miranda, J.; Brazil, J.C.; Morris, A.H.; Parkos, C.A.; Quiros, M.; Nusrat, A. Maresin-2 promotes mucosal repair and has therapeutic potential when encapsulated in thermostable nanoparticles. Proc. Natl. Acad. Sci. USA 2023, 120, e2218162120. [Google Scholar] [CrossRef]
- Cassano, R.; Serini, S.; Curcio, F.; Trombino, S.; Calviello, G. Preparation and Study of Solid Lipid Nanoparticles Based on Curcumin, Resveratrol and Capsaicin Containing Linolenic Acid. Pharmaceutics 2022, 14, 1593. [Google Scholar] [CrossRef]
- Ibrahim, D.; Arisha, A.H.; Khater, S.I.; Gad, W.M.; Hassan, Z.; Abou-Khadra, S.H.; Mohamed, D.I.; Ahmed Ismail, T.; Gad, S.A.; Eid, S.A.M.; et al. Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants 2022, 11, 1523. [Google Scholar] [CrossRef]
- Dravid, A.A.; Dhanabalan, K.M.; Naskar, S.; Vashistha, A.; Agarwal, S.; Padhan, B.; Dewani, M.; Agarwal, R. Sustained release resolvin D1 liposomes are effective in the treatment of osteoarthritis in obese mice. J. Biomed. Mater. Res. A 2023, 111, 765–777. [Google Scholar] [CrossRef]
- Tsubosaka, M.; Kihara, S.; Hayashi, S.; Nagata, J.; Kuwahara, T.; Fujita, M.; Kikuchi, K.; Takashima, Y.; Kamenaga, T.; Kuroda, Y.; et al. Gelatin hydrogels with eicosapentaenoic acid can prevent osteoarthritis progression in vivo in a mouse model. J. Orthop. Res. 2020, 38, 2157–2169. [Google Scholar] [CrossRef]
- Soliman, M.S.; Abd-Allah, F.I.; Hussain, T.; Saeed, N.M.; El-Sawy, H.S. Date seed oil loaded niosomes: Development, optimization and anti-inflammatory effect evaluation on rats. Drug Dev. Ind. Pharm. 2018, 44, 1185–1197. [Google Scholar] [CrossRef]
- Kheilnezhad, B.; Hadjizadeh, A. Factors Affecting the Penetration of Niosome into the Skin, Their Laboratory Measurements and Dependency to the Niosome Composition: A Review. Curr. Drug Deliv. 2021, 18, 555–569. [Google Scholar] [CrossRef]
- Fu, Y.; Lei, J.; Zhuang, Y.; Zhang, K.; Lu, D. Overexpression of HMGB1 A-box reduced IL-1beta-induced MMP expression and the production of inflammatory mediators in human chondrocytes. Exp. Cell Res. 2016, 349, 184–190. [Google Scholar] [CrossRef]
- Shang, H.; Younas, A.; Zhang, N. Recent advances on transdermal delivery systems for the treatment of arthritic injuries: From classical treatment to nanomedicines. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2022, 14, e1778. [Google Scholar] [CrossRef]
- Li, J.; Tian, X.; Wang, K.; Jia, Y.; Ma, F. Transdermal delivery of celecoxib and alpha-linolenic acid from microemulsion-incorporated dissolving microneedles for enhanced osteoarthritis therapy. J. Drug Target 2023, 31, 206–216. [Google Scholar] [CrossRef]
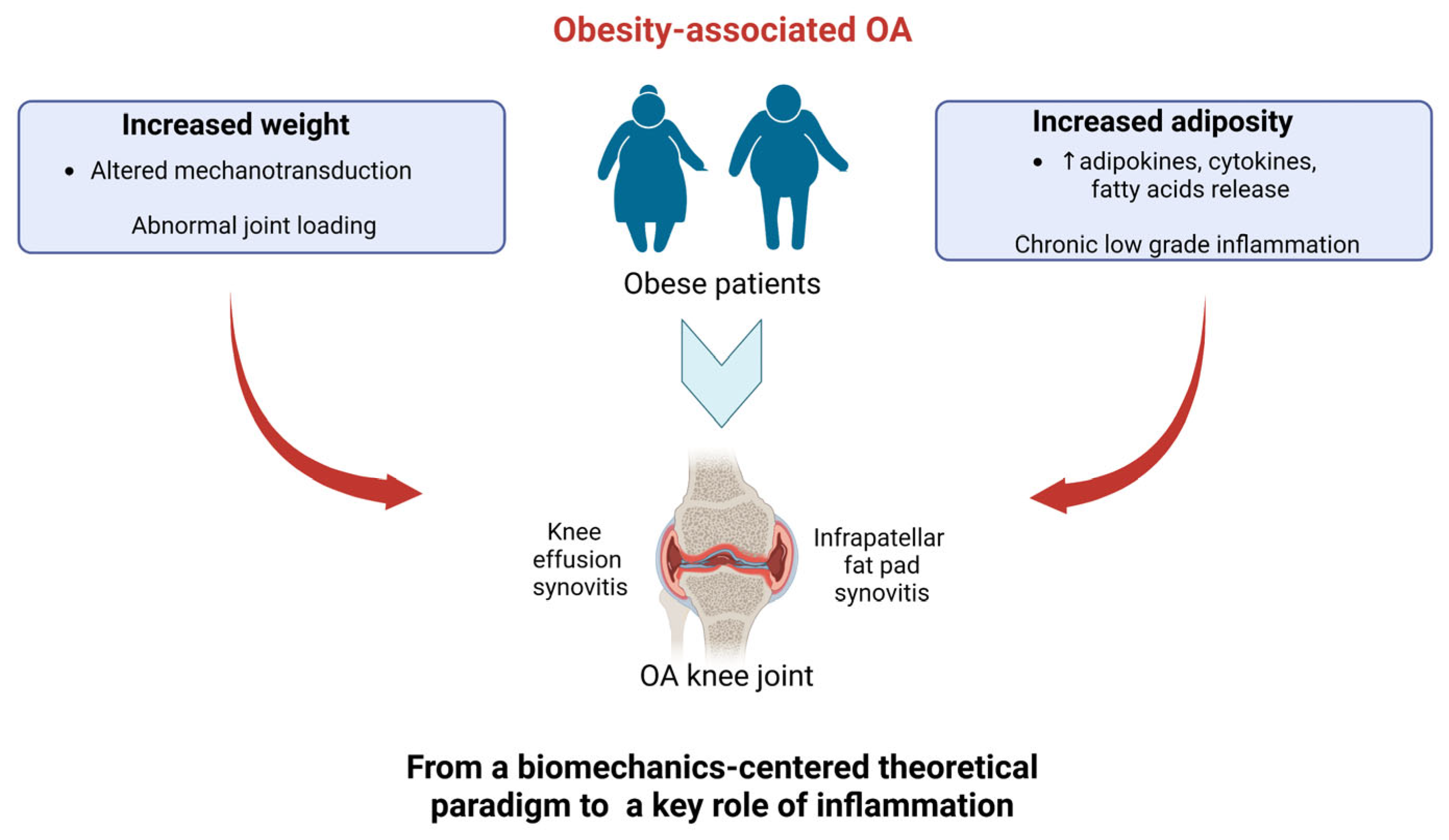




| Molecule | In Vivo Model | Dose/Delivery Route | Main Effects | Specific Outcomes | Indications for Preventive-Therapeutic Strategies for ObOA | Ref. |
|---|---|---|---|---|---|---|
| N/A | Post-traumatic model of OA: ACLT model in rabbits | N/A | Early-stage OA affected FA composition towards a pro-inflammatory phenotype. |
| Importance of preventing n-3/n-6 PUFAs imbalance/restoring n-3/n-6 PUFAs balance. | [132] |
| HFD (60% kcal fat) | Ob model: male C57BL/6J mice + HFD |
| HFD caused moderate OA. |
| Avoiding HFD to prevent OA onset or delay OA aggravation. | [139] |
| MFD | Ob model: male C57BL/6J (B6) mice + MFD |
| MFD promoted changes in immune metabolism and altered gut microbiota composition. |
| Fat diet content above 11% induces metabolic changes. | [140] |
| HFD (60% kcal fat) | Ob model: male C57BL/6J mice + HFD |
Dietary supplementation | HFD promoted OA onset. Moderate exercise improved glucose tolerance without reducing body fat or cytokine levels. | HFD promoted OA onset by:
| Avoid HFD for preventing OA onset or delaying OA aggravation. Recommend aerobic exercise in promoting joint health independently of weight loss. | [137] |
| PUFAs diet | ObOA model: DMM model in male mice + HFD (60% kcal fat) |
| Protective role of dietary supplementation with n-3 PUFAs in mitigating OA changes was observed. |
| Shifting the composition of Fas in the diet towards a low intake of n-6 PUFAs and SFAs and a high intake of n-3 PUFAs for mitigating OA. | [15] |
| PUFAs diet | ObOA model: DMM model in male mice + HFD |
| Dietary FA content modulated OA severity; small amounts of n-3 PUFAs could mitigate OA while independently increasing OA severity. |
| Shifting of FA composition in diet toward a low intake of n-6 PUFAs and SFAs and a high intake of n-3 PUFAs for mitigating OA. | [141] |
| HFD rich in n-6 PUFAs or control diet | ObOA model: DMM model + HFD in male and female fat-1 transgenic mice (encoding n-3 PUFAs desaturase) |
| Circulating FA composition and metabolic inflammation rather than “mechanical” factors were major risk factors for ObOA. |
| Potential genetic use of ω-3 FA desaturase to reduce ObOA. | [142] |
| Standard diet | Obese model of OA: DMM model in fat-1 transgenic (TG) mice vs. Wild-type mice |
| Protective role of desaturase in mitigating OA, probably through inhibition of mTORC1, and the promotion of autophagy and cell survival in cartilage chondrocytes was observed. |
| Potential genetic use of n-3 PUFAs desaturase to reduce ObOA. | [143] |
| Diet supplemented with 10% safflower oil and with 23% kcal fat, with an n-6:n-3 ratio of 274 | Spontaneous OA model in fat-1 TG mice |
| Cartilage degeneration and osteophyte formation was developed at levels comparable to WT mice. |
| Lower efficacy of n-3 PUFAs desaturase in spontaneous OA than post-traumatic OA models. | [144] |
| No diet | Ob models: leptin-deficient (ob/ob) and leptin-receptor-deficient (db/db) female mice No OA induction | N/A | Impaired leptin signaling significantly altered subchondral bone morphology without altering knee OA. |
| Leptin signaling is key to inducing systemic inflammation. | [145] |
| ALA, EPA, DHA | Ob models: - C57BL/6J mice + HFD (236 g/kg fat); - leptin-deficient (ob/ob) + HFD (236 g/kg fat) | C57BL/6J 16 weeks ob/ob mice for 6 weeks | Supplementation with EPA, but not ALA and DHA, could preserve glucose homeostasis in an obesogenic environment and limit fat mass accumulation. | HFD:
| EPA is more effective in targeting specific Ob features. | [146] |
| n-3 PUFAs | Spontaneous model of OA (OA guinea pig) |
| Chondroprotective effects were observed. |
| Relevance of a diet rich in n-3 PUFAs to counteract cartilage degradation. | [147] |
| GLM abundant in DHA | MIA-induced OA model in male Wistar rats | GLM (100–300 mg/kg) versus celecoxib (50 mg/kg) Oral administration (3 days from MIA injection) | Chondroprotective properties and a reduction in catabolic, inflammatory and necroptotic markers were observed. |
| Potential candidate in targeting inflammation and necroptosis. | [115] |
| PDX (DHA metabolite) | MIA-induced OA model in Sprague–Dawley rats | 10 µg/kg (every 3 days) Intraperitoneal injections | Chondroprotective and anti-inflammatory effects were observed. |
| Potential tool to target inflammatory hallmarks. | [118] |
| DHA | Post-traumatic OA: ACLT-induced OA model in male Sprague–Dawley rats | 1 mg/kg (two months) Injection in tail vein | Promotion of bone remodelling and cartilage reduction were observed. |
| Potential of targeting catabolic markers. | [148] |
| Antarctic krill oil (Euphausia superba) (rich in EPA and DHA) | DMM-induced OA in osteoporotic (ovariectomy) mice | Diet supplementation | Chondroprotection and reduction in inflammation were observed. |
| Potential of targeting inflammatory markers. | [134] |
| Triglyceride n-3 oil (rich in DHA + EPA) | Naturally occurring OA in dogs Prospective, randomized, double-blind, placebo-controlled clinical trial. | 69 mg EPA + DHA/kg/day (84 days) Diet supplementation | Improvement in clinical markers of OA was observed. |
| Potential of reducing systemic inflammation. | [149] |
| EPA and DHA | OA horses | Diet supplementation | Increased storage pools of n-3 PUFAs in SF and anti-inflammatory effect |
| Potential to improve the resolution of inflammation. | [150] |
| 17®-HDoHE (RvD2 precursor) | MIA model of OA + MNX model of OA | 1 ng/μL (every day from 14 to 28 days) Intra-peritoneal administration | Long-term inhibitory effects on nociceptive signaling. |
| Potential to exert analgesic potential. | [151] |
| Aspirin-triggered RvD1 (AT-RvD1) RvD1 isomer induced by aspirin and more resistant to enzymatic degradation than RvD1 | Carrageenan-induced inflammatory or MIA-induced OA in male Sprague–Dawley rats | 15 ng in 50 μL PBS (carrageenan model) 15 ng and 150 ng in 50 μL PBS (MIA model) Spinal treatment | The selective target of inflammation drives spinal hyperexcitability in nociceptive pathways (analgesic potential) |
| Potential to exert analgesic potential. | [152] |
| RvE1/RvD1/PDX | Rat paws inflamed by carrageenan or histamine, 5-hydroxytryptamine, substance P or prostaglandin E2 | 20 RvE1, 100 RvD1,100 μg PDX mL−1 versus standard anti-inflammatory drugs (INDO, celecoxib and dexamethasone) Injection in the hind paws (10 min before the stimuli) | Analgesic and anti-inflammatory effects |
| Rvs, as analgesic agents, may be a better therapeutic agent than NSAIDs. | [153] |
| MaR2 | Lipopolysaccharide (LPS)-induced mechanical hyperalgesia capsaicin (TRPV1 agonist) or AITC (TRPA1 agonist). | 3, 10 or 30 ng | Analgesic effect |
| Potential analgesic effects. | [154] |
| Linseed oil (LO), soybean oil (SO) and peanut oil (PO) n-6/n-3 PUFA ratios: 1:3.85 (LO), 9.15:1 (SO) and 372.73:1 (PO) | DMM OA murine model | 12 weeks Oral supplementation | Edible oils with low n-6/n-3 PUFAs exert an anti-inflammatory effect by inhibiting the NFκB pathway. |
| Anti-inflammatory potential of a low-n-6/n-3-PUFA diet. | [155] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambari, L.; Cellamare, A.; Grassi, F.; Grigolo, B.; Panciera, A.; Ruffilli, A.; Faldini, C.; Desando, G. Targeting the Inflammatory Hallmarks of Obesity-Associated Osteoarthritis: Towards Nutraceutical-Oriented Preventive and Complementary Therapeutic Strategies Based on n-3 Polyunsaturated Fatty Acids. Int. J. Mol. Sci. 2023, 24, 9340. https://doi.org/10.3390/ijms24119340
Gambari L, Cellamare A, Grassi F, Grigolo B, Panciera A, Ruffilli A, Faldini C, Desando G. Targeting the Inflammatory Hallmarks of Obesity-Associated Osteoarthritis: Towards Nutraceutical-Oriented Preventive and Complementary Therapeutic Strategies Based on n-3 Polyunsaturated Fatty Acids. International Journal of Molecular Sciences. 2023; 24(11):9340. https://doi.org/10.3390/ijms24119340
Chicago/Turabian StyleGambari, Laura, Antonella Cellamare, Francesco Grassi, Brunella Grigolo, Alessandro Panciera, Alberto Ruffilli, Cesare Faldini, and Giovanna Desando. 2023. "Targeting the Inflammatory Hallmarks of Obesity-Associated Osteoarthritis: Towards Nutraceutical-Oriented Preventive and Complementary Therapeutic Strategies Based on n-3 Polyunsaturated Fatty Acids" International Journal of Molecular Sciences 24, no. 11: 9340. https://doi.org/10.3390/ijms24119340