


Enzymatic Hydrolysis Optimization for Preparation of Sea Cucumber (Holothuria scabra) Hydrolysate with an Antiproliferative Effect on the HepG2 Liver Cancer Cell Line and Antioxidant Properties


 , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of Sea Cucumber
2.2. Optimization of Sea Cucumber Hydrolysis Conditions
Response Surface Model Generation
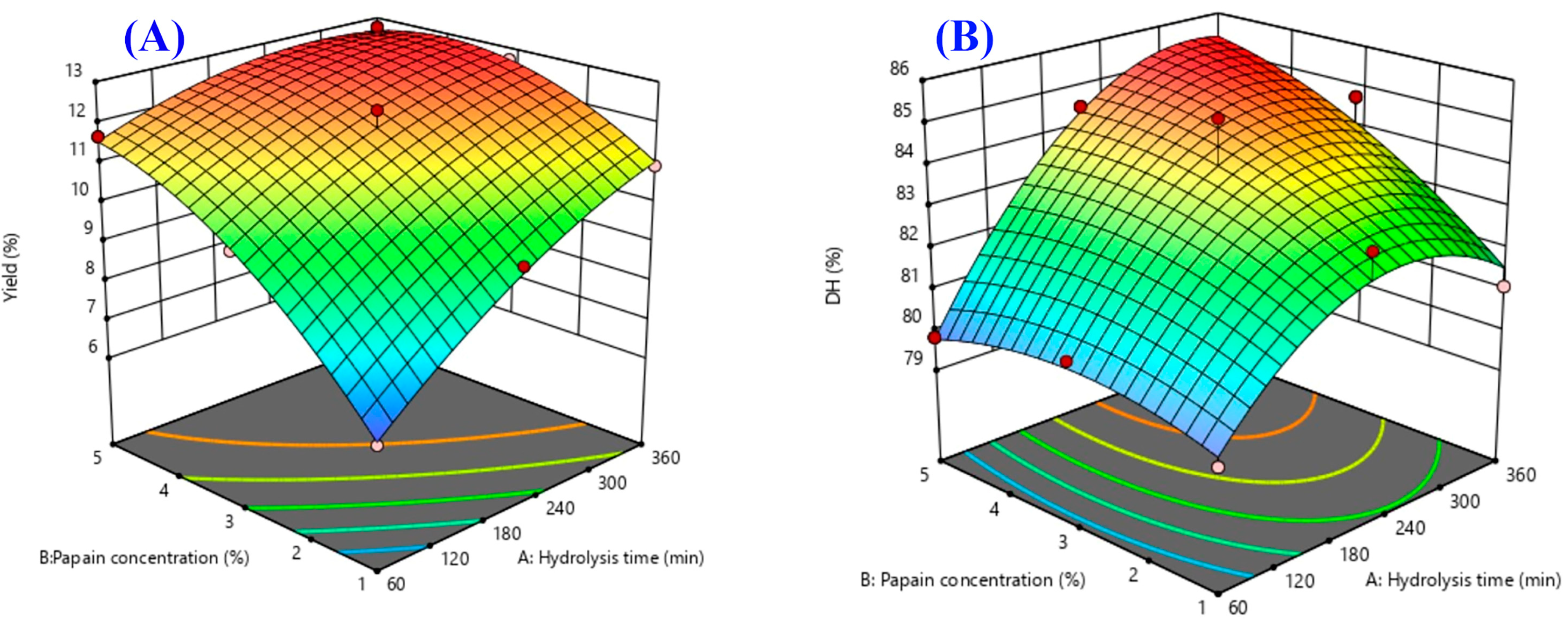
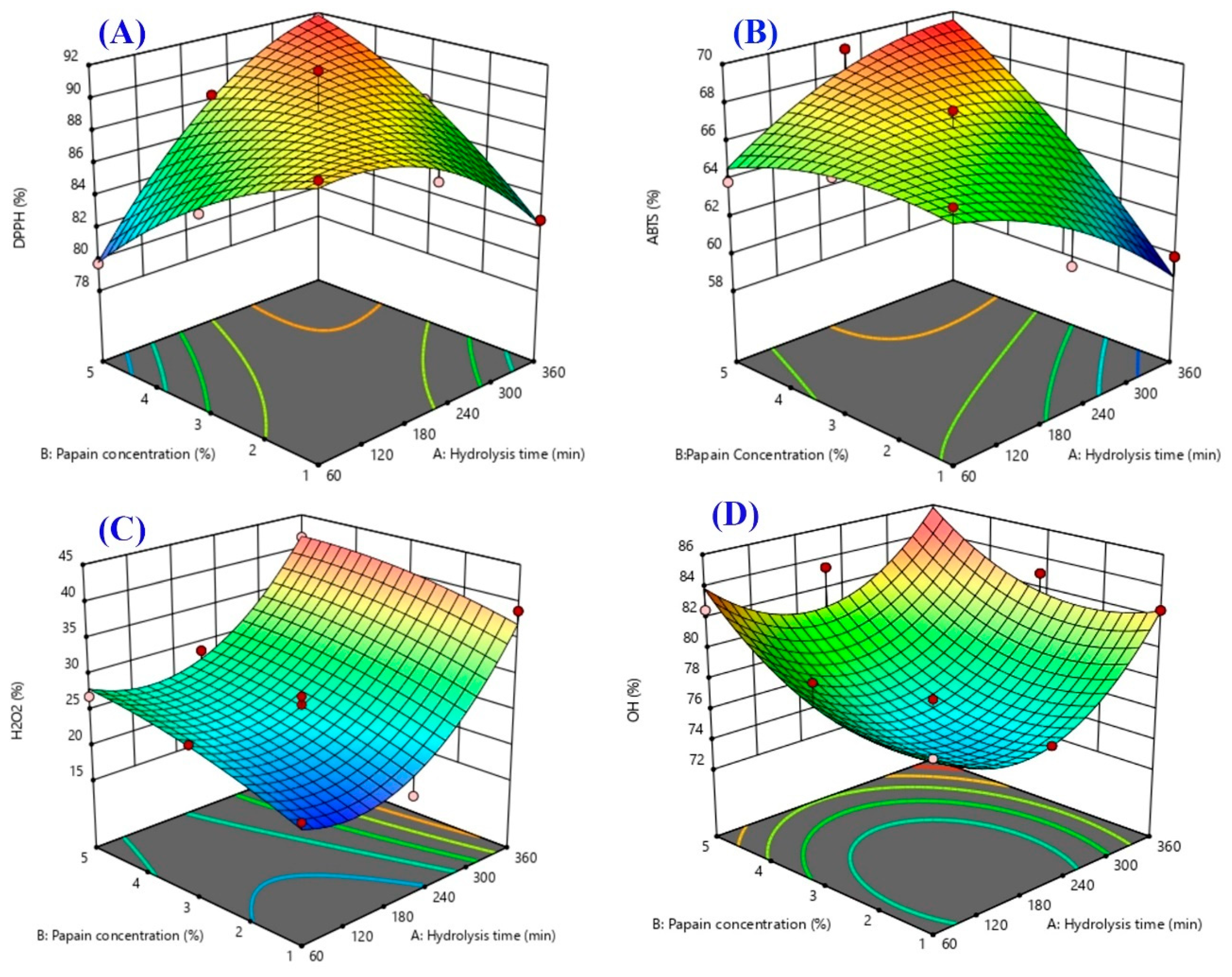
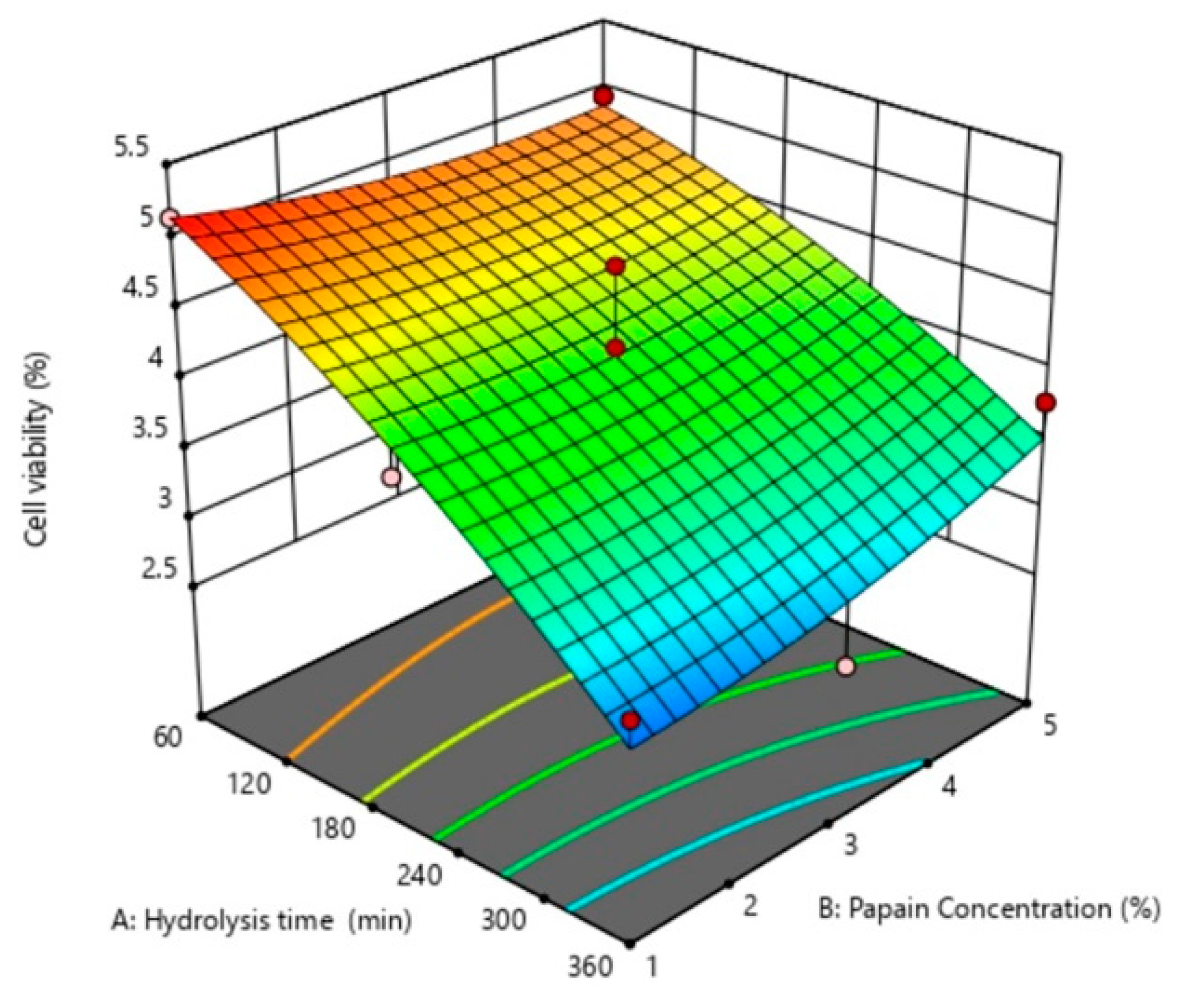
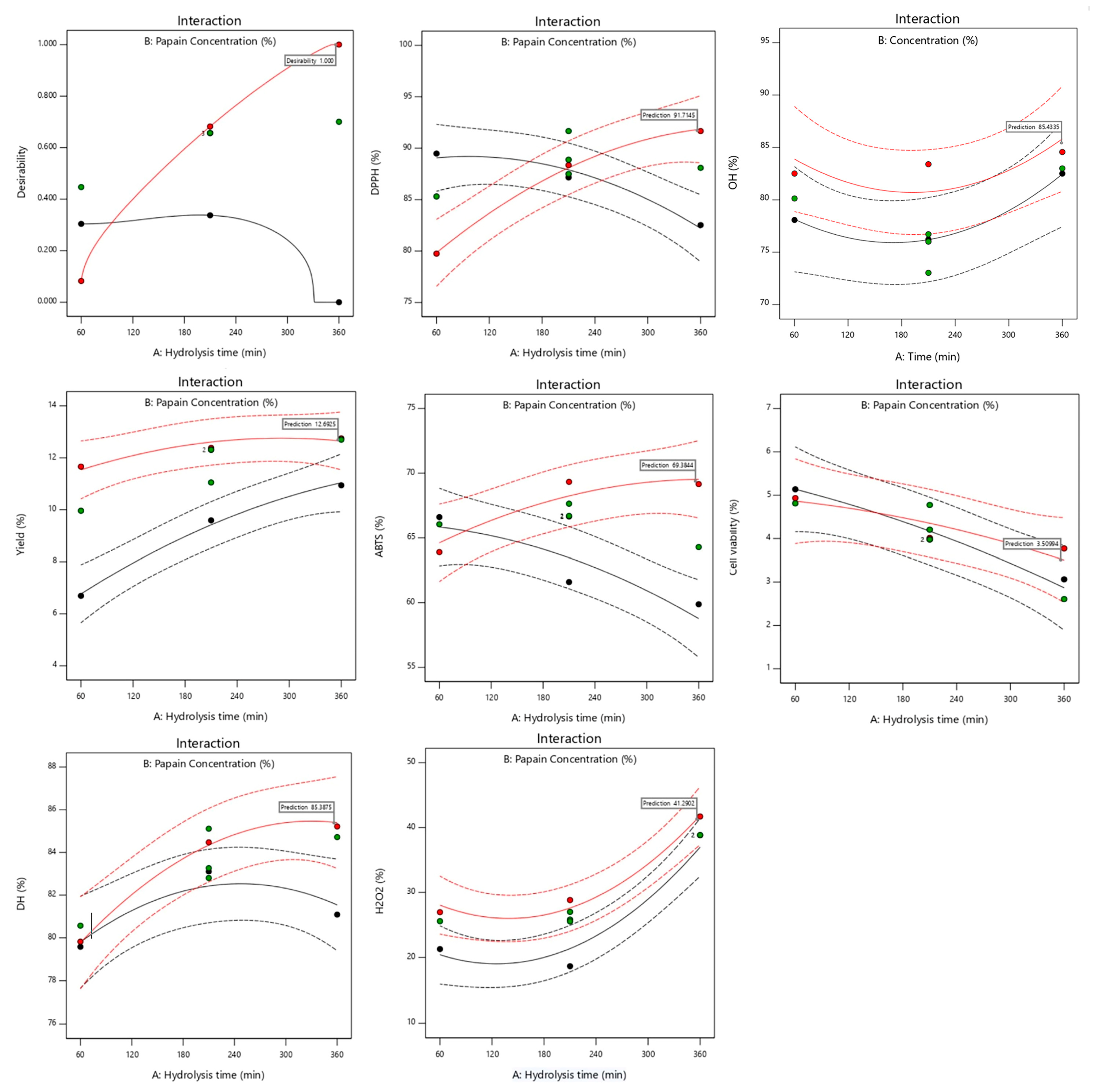
2.3. Response Surface Plots
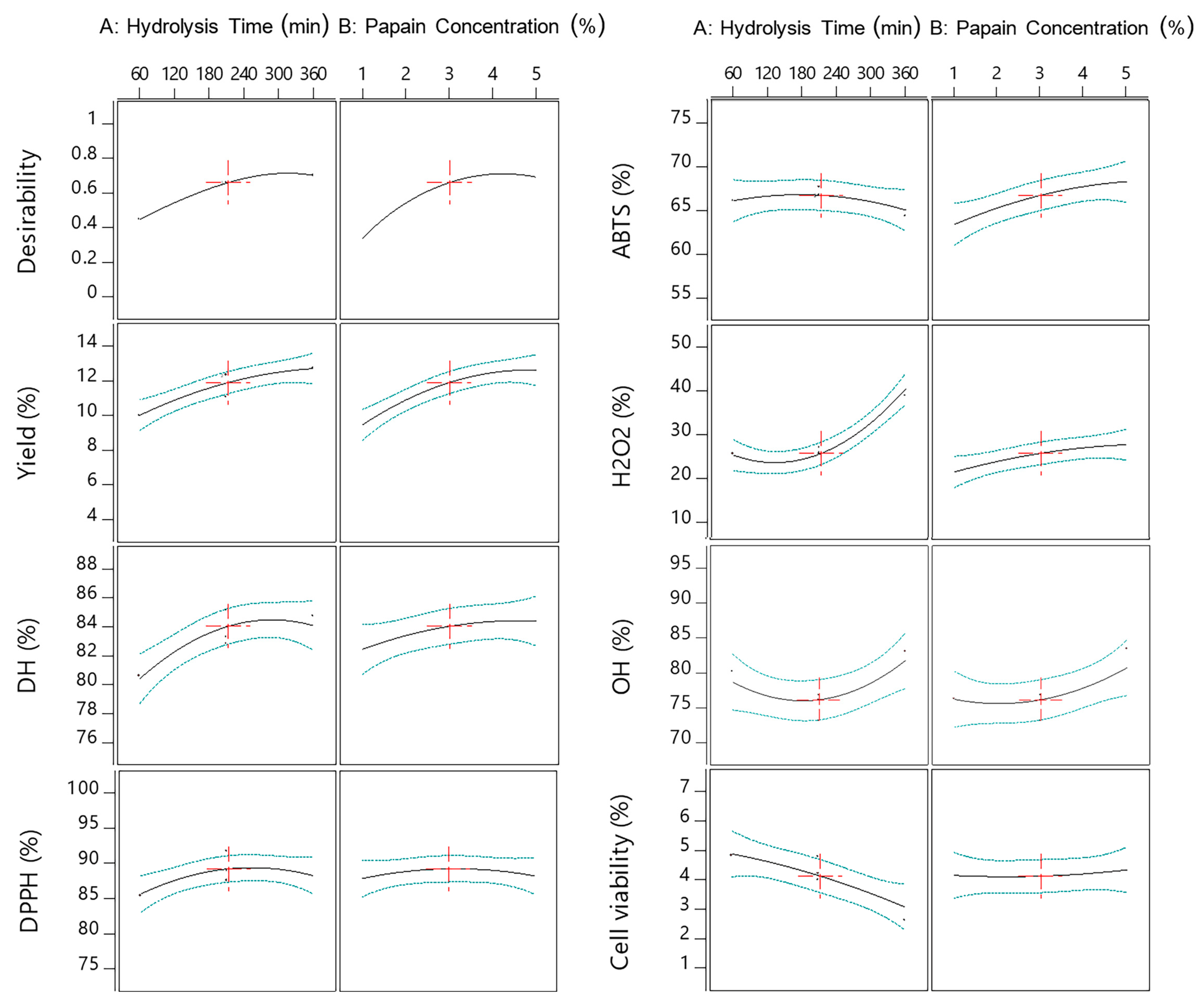
2.4. Optimization of Multiple Responses and Model Validation
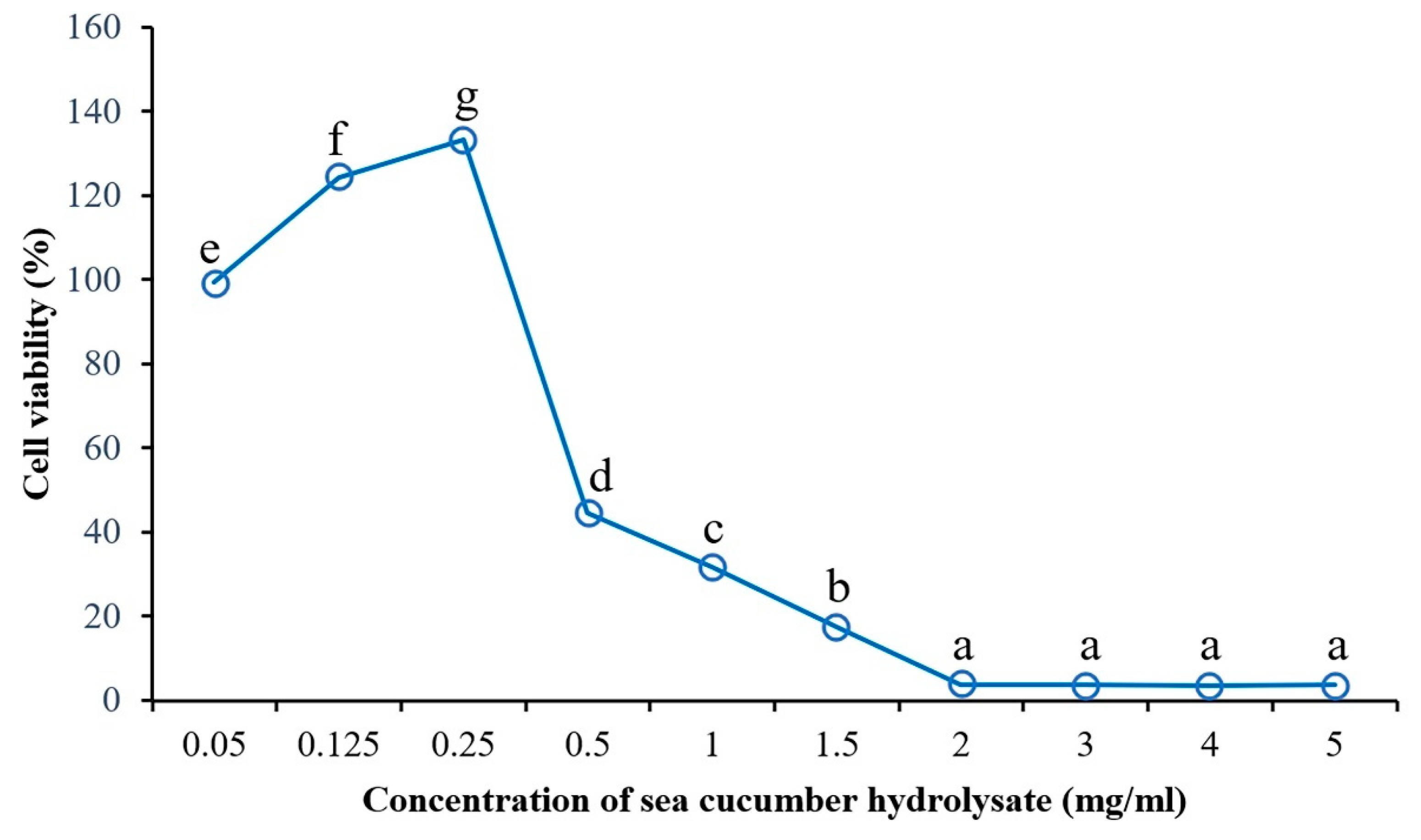
2.5. The Effect of the Sea Cucumber Hydrolysate Concentration on Anticancer Activity
3. Materials and Methods
3.1. Raw Materials and Preparations
3.2. Chemicals and Enzymes
3.3. Enzymatic Hydrolysis of Sea Cucumber
3.4. Analyses
3.4.1. Chemical Composition of Sea Cucumber
3.4.2. Yield
3.4.3. Degree of Hydrolysis (DH)
3.4.4. DPPH Radical Scavenging Activity
3.4.5. ABTS Radical Scavenging Activity
3.4.6. Hydrogen Peroxide (H2O2) Scavenging Activity
3.4.7. Hydroxyl Radical Scavenging Activity
3.4.8. Antiproliferative Activity in the HepG2 Liver Cancer Cell Line
3.5. The Effect of the Sea Cucumber Hydrolysate Concentration on the Antiproliferative Activity of the HepG2 Liver Cancer Cell Line
3.6. Experimental Design and Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Dabbagh, B.; Elhaty, I.A.; Hrout, A.A.; Sakkaf, R.A.; El-Awady, R.; Ashraf, S.S.; Amin, A. Antioxidant and anticancer activities of Trigonella foenum-graecum, Cassia acutifolia and Rhazya stricta. BMC Complement. Altern. Med. 2018, 18, 240. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubzadeh, Z.; Ghadikolaii, F.P.; Kaboosi, H.; Safari, R.; Fattahi, E. Antioxidant activity and anticancer effect of bioactive peptides from rainbow trout (Oncorhynchus mykiss) skin hydrolysate. Int. J. Pept. Res. Ther. 2020, 26, 625–632. [Google Scholar] [CrossRef]
- Hasani, K.; Ariaii, P.; Ahmadi, M. Antimicrobial, antioxidant and anti-cancer properties of protein hydrolysates from Indian mackerel (Rastrelliger kanagurta) waste prepared using commercial enzyme. Int. J. Pept. Res. Ther. 2022, 28, 86. [Google Scholar] [CrossRef]
- Ghanbari, R.; Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Angiotensin-I converting enzyme (ACE) inhibitory and antioxidant activities of sea cucumber (Actinopyga lecanora) hydrolysates. Int. J. Mol. Sci. 2015, 16, 28870–28885. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, T.; Ding, G.F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.J.; Ramasamy, P.; Wahid, M.E.A.; Vairappan, C.S. Anticancer activity and mediation of apoptosis in human HL-60 leukaemia cells by edible sea cucumber (Holothuria edulis) extract. Food Chem. 2013, 139, 326–331. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant protein-derived antioxidant peptides: Isolation, identification, mechanism of action and application in food systems: A review. LWT 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Doungapai, C.; Siriwoharn, T.; Malila, Y.; Autsavapromporn, N.; Makkhun, S.; Yarnpakdee, S.; Jantanasakulwong, K.; Regenstein, J.M.; Wangtueai, S. UV-B Protective and antioxidant activities of protein hydrolysate from sea cucumber (Holothuria scabra) using enzymatic hydrolysis. Front. Mar. Sci. 2022, 9, 892255. [Google Scholar] [CrossRef]
- Yan, M.; Tao, H.; Qin, S. Effect of enzyme type on the antioxidant activities and functional properties of enzymatic hydrolysates from sea cucumber (Cucumaria frondosa) viscera. J. Aquat. Food Prod. Technol. 2016, 25, 940–952. [Google Scholar] [CrossRef]
- Li, M.; Gao, Y.; Qi, Y.X.; Song, Z.Y.; Li, Z.B.; Lin, Y.T.; Zhao, Q.C. Assessment of the nutritional value of cultured sea cucumber Apostichopus japonicus. J. Aquat. Food Prod. Technol. 2021, 30, 868–879. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, R.; Wen, Z. Bioactive compounds and biological functions of sea cucumbers as potential functional foods. J. Funct. Foods 2018, 49, 73–84. [Google Scholar] [CrossRef]
- Wangtueai, S.; Phimolsiripol, Y.; Vichasilp, C.; Regenstein, J.M.; Schöenlechner, R. Optimization of gluten-free functional noodles formulation enriched with fish gelatin hydrolysates. LWT-Food. Sci. Technol. 2020, 133, 109977. [Google Scholar] [CrossRef]
- Upata, M.; Siriwoharn, T.; Makkhun, S.; Yarnpakdee, S.; Regenstein, J.M.; Wangtueai, S. Tyrosinase inhibitory and antioxidant activity of enzymatic protein hydrolysate from jellyfish (Lobnema smithii). Foods 2022, 11, 615. [Google Scholar] [CrossRef] [PubMed]
- Mongkonkamthorn, N.; Malila, Y.; Regenstein, J.M.; Wangtueai, S. Enzymatic hydrolysis optimization for preparation of tuna dark meat hydrolysate with antioxidant and angiotensin I-converting enzyme (ACE) inhibitory activities. J. Aquat. Food Prod. Technol. 2021, 30, 1090–1108. [Google Scholar] [CrossRef]
- Halim, N.R.A.; Sarbon, N.M. A response surface approach on hydrolysis condition of eel (Monopterus sp.) protein hydrolysate with antioxidant activity. Int. Food Res. J. 2017, 24, 1081–1093. [Google Scholar]
- Dai, Z.Y.; Zhang, Y.P.; Zhang, H.; Lu, Y.B. Preparation and characterization of mussel (Mytilus edulis) protein hydrolysates with angiotensin-I-converting enzyme (ACE) inhibitory activity by enzymatic hydrolysis. J. Food Biochem. 2010, 36, 66–74. [Google Scholar] [CrossRef]
- Mongkonkamthorn, N.; Malila, Y.; Yarnpakdee, S.; Makkhun, S.; Regenstein, J.M.; Wangtueai, S. Production of protein hydrolysate containing antioxidant and angiotensin-I-converting enzyme (ACE) inhibitory activities from tuna (Katsuwonus pelamis) blood. Processes 2020, 8, 1518. [Google Scholar] [CrossRef]
- Hsu, K.C.; Li-Chan, E.C.Y.; Jao, C.L. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chem. 2011, 126, 617–622. [Google Scholar] [CrossRef]
- Greyling, N.; Bordoloi, A.; Goosen, N.J. Optimising enzymatic conditions of monkfish (Lophius vomerinus) heads hydrolysis towards potential waste biomass valorisation. Biomass Conv. Bioref. 2021, 11, 2711–2722. [Google Scholar] [CrossRef]
- Silva, V.M.; Park, K.J.; Hubinger, M.D. Optimization of the enzymatic hydrolysis of mussel meat. J. Food Sci. 2010, 75, 36–42. [Google Scholar] [CrossRef]
- Song, L.; Li, T.; Yu, R.; Yan, C.; Ren, S.; Zhao, Y. Antioxidant activities of hydrolysates of Arca subcrenata prepared with three proteases. Mar. Drugs 2008, 6, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Park, S.Y.; Han, E.J.; Kim, H.S.; Kang, D.S.; Je, J.Y.; Ahn, C.B.; Ahn, G. Isolation of an antioxidant peptide from krill protein hydrolysates as a novel agent with potential hepatoprotective effects. J. Funct. Foods 2020, 67, 103889. [Google Scholar] [CrossRef]
- Fang, X.; Xie, N.; Chen, X.; Yu, H.; Chen, J. Optimization of antioxidant hydrolysate production from flying squid muscle protein using response surface methodology. Food Bioprod. Process. 2012, 90, 676–682. [Google Scholar] [CrossRef]
- Centenaro, G.S.; Centenaro, M.S.; Hernandez, C.P. Antioxidant activity of protein hydrolysates of fish and chicken bones. Adv. J. Food Sci. Technol. 2011, 3, 280–288. [Google Scholar]
- Latorres, J.M.; Rios, D.G.; Saggiomo, G.; Wasielesky, W.; Prentice-Hernandez, C. Functional and antioxidant properties of protein hydrolysates obtained from white shrimp (Litopenaeus vannamei). J. Food Sci. Technol. 2018, 55, 721–729. [Google Scholar] [CrossRef]
- Suresh, P.V. Protein hydrolysate from duck egg white by Flavourzyme® digestion: Process optimisation by model design approach and evaluation of antioxidant capacity and characteristic properties. LWT-Food. Sci. Technol. 2022, 156, 113018. [Google Scholar]
- Je, J.Y.; Lee, K.H.; Lee, M.H.; Ahn, C.B. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food. Res. Int. 2009, 42, 1266–1272. [Google Scholar] [CrossRef]
- Karnjanapratum, S.; Benjakul, S. Antioxidative gelatin hydrolysate from unicorn leatherjacket skin as affected by prior autolysis. Int. Aquat. Res. 2015, 7, 101–114. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Zhao, X.; Li, B.F. Optimization of antioxidant activity by response surface methodology in hydrolysates of jellyfish (Rhopilema esculentum) umbrella collagen. J. Zhejiang Univ. Sci. B 2009, 10, 572–579. [Google Scholar] [CrossRef]
- Ishak, N.H.; Sarbon, N.M. Optimization of the enzymatic hydrolysis conditions of waste from shortfin scad (Decapterus Macrosoma) for the production of angiotensin I-converting enzyme (ACE) inhibitory peptide using response surface methodology. Food Res. Int. 2017, 24, 1735. [Google Scholar]
- Halim, N.R.A.; Azlan, A.; Yusof, H.M.; Sarbon, N.M. Antioxidant and anticancer activities of enzymatic eel (Monopterus sp.) protein hydrolysate as influenced by different molecular weight. Biocatal. Agric. Biotechnol. 2018, 16, 10–16. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.G.; Li, R.F.; Li, Y.; Cui, Z.C.; Duan, L.X.; Lu, F. Gecko crude peptides induce apoptosis in human liver carcinoma cells in vitro and exert antitumor activity in a mouse ascites H22 xenograft model. J. Biomed. Biotechnol. 2012, 2012, 743573. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, Proximate Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Jumeri and Kim, S.M. Anticancer and anticancer activities of enzymatic hydrolysates of solitary tunicate (Styela clava). Food Sci. Biotechnol. 2011, 20, 1075–1085. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Sivagami, G.; Balasubramanian, T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed. Prev. Nutr. 2014, 4, 343–353. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Moisture (%) | Protein (%) | Fat (%) | Ash (%) |
|---|---|---|---|---|
| Dried sea cucumber | 17.22 ± 0.47 | 65.75 ± 0.78 | 1.41 ± 0.08 | 13.98 ± 0.03 |
| Sea cucumber Slurry | 82.69 ± 0.10 | 12.40 ± 1.3 | 1.24 ± 0.10 | 2.03 ± 0.09 |
| Experimental Units | Factors | Responses | |||||||
|---|---|---|---|---|---|---|---|---|---|
| X1 | X2 | Yield (%) | DH (%) | DPPH (%) | ABTS (%) | H2O2 (%) | OH· (%) | Cell Viability (%) | |
| (min) | (%) | ||||||||
| 1 | 60 | 1 | 6.69 a ± 1.06 | 79.6 a ± 0.44 | 89.5 bcd ± 0.84 | 66.6 bcde ± 3.14 | 21.3 ab ± 1.53 | 78.1 bc ± 3.77 | 5.14 d ± 0.55 |
| 2 | 360 | 1 | 10.9 bc ± 1.98 | 81.1 ab ± 1.74 | 82.5 abc ± 1.12 | 59.9 a ± 0.74 | 38.8 d ± 0.71 | 82.5 c ± 1.66 | 3.06 b ± 0.42 |
| 3 | 60 | 5 | 11.7 cd ± 0.23 | 79.8 a ± 0.34 | 79.8 a ± 1.97 | 63.9 abc ± 1.20 | 27.0 c ± 2.68 | 82.5 c ± 2.33 | 4.93 d ± 0.46 |
| 4 | 360 | 5 | 12.8 e ± 0.04 | 85.2 bc ± 0.95 | 91.7 cd ± 1.68 | 69.2 e ± 2.54 | 41.7 d ± 2.96 | 84.6 bc ± 4.01 | 3.77 bc ± 0.49 |
| 5 | 60 | 3 | 9.96 b ± 0.31 | 80.6 bc ± 1.99 | 85.3 ab ± 1.12 | 66.1 abcd ± 1.94 | 25.6 c ± 0.73 | 80.1 bc ± 1.39 | 4.81 cd ± 0.45 |
| 6 | 360 | 3 | 12.7 e ± 0.03 | 84.7 c ± 2.17 | 88.1 bcd ± 3.25 | 64.3 cde ± 0.85 | 38.9 d ± 0.85 | 83.0 bc ± 1.90 | 2.61 a ± 0.42 |
| 7 | 210 | 1 | 9.59 b ± 0.35 | 83.1 bc ± 0.97 | 87.2 bcd ± 3.08 | 61.6 ab ± 2.55 | 18.7 a ± 14.84 | 76.2 b ± 3.15 | 3.98 cd ± 0.62 |
| 8 | 210 | 5 | 12.4 de ± 0.23 | 84.5 bc ± 2.16 | 88.4 bcd ± 1.68 | 69.3 de ± 0.92 | 28.8 c ± 1.06 | 83.4 bc ± 0.92 | 4.01 bc ± 0.85 |
| 9 | 210 | 3 | 12.3 de ± 0.13 | 83.3 bc ± 3.81 | 91.7 d ± 0.56 | 67.6 de ± 0.10 | 25.8 c ± 1.40 | 73.0 a ± 7.59 | 4.77 cd ± 0.16 |
| 10 | 210 | 3 | 11.1 bc ± 0.78 | 85.1 bc ± 0.93 | 87.5 bcd ± 1.40 | 66.7 bcde ± 1.57 | 25.6 bc ± 1.92 | 76.7 bc ± 4.55 | 3.98 bc ± 0.53 |
| 11 | 210 | 3 | 12.3 de ± 2.29 | 82.8 ab ± 1.80 | 88.9 bcd ± 0.69 | 66.7 bcde ± 2.12 | 27.0 c ± 1.92 | 76.0 b ± 3.98 | 4.20 bc ± 0.24 |
| Responses | Quadratic Polynomial Model | R2 | p-Value for Models | p-Value for Lack of Fit |
|---|---|---|---|---|
| Yield (%) | Y1 = 11.9 + 1.35X1 + 1.59X2 − −0.79X1X2 − 0.52X12 − 0.86X22 | 0.9643 | 0.0013 | 0.9654 |
| DH (%) | Y2 = 84.0 + 1.84X1 + 0.96X2 + 0.97X1X2 − 1.76X12 − 0.61X22 | 0.9023 | 0.0146 | 0.8619 |
| DPPH (%) | Y3 = 89.2 + 1.29X1 − 0.10X2 + 4.72X1X2 − 2.26X12 − 1.20X22 | 0.9225 | 0.0083 | 0.9633 |
| ABTS (%) | Y4 = 66.8 − 0.54X1 + 2.38X2 + 2.99X1X2 − 1.17X12 − 0.88X22 | 0.9023 | 0.0145 | 0.1068 |
| H2O2 (%) | Y5 = 25.6 − 7.58X1 + 3.13X2 − 0.70X1X2 + 7.33X12 − 1.14X22 | 0.9661 | 0.0011 | 0.0925 |
| OH· (%) | Y6 = 76.2 + 1.56X1X + 0.28X2 − 0.59X1X2 + 4.10X12 + 2.34X22 | 0.8354 | 0.0495 | 0.5691 |
| Cell viability (%) | Y7 = 4.14 − 0.91X1 − 0.09X2 + 0.23X1X2 − 0.17X12 + 0.12X22 | 0.8525 | 0.0384 | 0.4981 |
| Responses | Code | Intercept | X1 | p-Value | X2 | p-Value | X1X2 | p-Value |
|---|---|---|---|---|---|---|---|---|
| Yield (%) | Y1 | 11.872 | 1.346 | 0.001 | 1.594 | 0.001 | −0.789 | 0.023 |
| DH (%) | Y2 | 83.997 | 1.841 | 0.005 | 0.957 | 0.055 | 0.971 | 0.093 |
| DPPH scavenging activity (%) | Y3 | 89.188 | 1.291 | 0.077 | 0.099 | 0.871 | 4.715 | 0.001 |
| ABTS scavenging activity (%) | Y4 | 66.728 | −0.539 | 0.359 | 2.384 | 0.007 | 2.994 | 0.006 |
| H2O2 scavenging activity (%) | Y5 | 25.636 | 7.584 | 0.0001 | 3.125 | 0.011 | −0.697 | 0.507 |
| OH· scavenging activity (%) | Y6 | 76.137 | 1.557 | 0.043 | 2.278 | 0.050 | −0.593 | 0.612 |
| Cell viability (%) | Y7 | 4.142 | −0.906 | 0.004 | 0.091 | 0.625 | 0.228 | 0.335 |
| Response | Parameters | Predicated Value | Real Value | Composite Desirability | ||||
|---|---|---|---|---|---|---|---|---|
| Goal | Lower | Upper | Weight | Importance | ||||
| X1 | is in range | 60 | 360 | 1 | 3 | 360 | 360 | 0.92 |
| X2 | is in range | 1 | 5 | 1 | 3 | 4.3 | 4.3 | |
| Yield (%) | maximize | 6.69 | 12.75 | 1 | 3 | 12.87 | 12.10 ± 0.54 | |
| DH (%) | maximize | 79.58 | 85.22 | 1 | 3 | 85.07 | 74.52 ± 1.06 | |
| DPPH (%) | maximize | 79.75 | 91.66 | 1 | 3 | 90.83 | 89.74 ± 1.51 | |
| ABTS (%) | maximize | 59.88 | 69.33 | 1 | 3 | 68.12 | 74.92 ± 0.81 | |
| H2O2 (%) | maximize | 18.66 | 41.7 | 1 | 3 | 41.64 | 39.42 ± 1.74 | |
| OH· (%) | maximize | 73.03 | 84.56 | 1 | 3 | 84.3 | 88.71 ± 1.49 | |
| Cell viability (%) | minimize | 2.61 | 5.14 | 1 | 3 | 3.32 | 9.89 ± 1.37 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saiwong, S.; Autsavapromporn, N.; Siriwoharn, T.; Techapun, C.; Wangtueai, S. Enzymatic Hydrolysis Optimization for Preparation of Sea Cucumber (Holothuria scabra) Hydrolysate with an Antiproliferative Effect on the HepG2 Liver Cancer Cell Line and Antioxidant Properties. Int. J. Mol. Sci. 2023, 24, 9491. https://doi.org/10.3390/ijms24119491
Saiwong S, Autsavapromporn N, Siriwoharn T, Techapun C, Wangtueai S. Enzymatic Hydrolysis Optimization for Preparation of Sea Cucumber (Holothuria scabra) Hydrolysate with an Antiproliferative Effect on the HepG2 Liver Cancer Cell Line and Antioxidant Properties. International Journal of Molecular Sciences. 2023; 24(11):9491. https://doi.org/10.3390/ijms24119491
Chicago/Turabian StyleSaiwong, Supansa, Narongchai Autsavapromporn, Thanyaporn Siriwoharn, Charin Techapun, and Sutee Wangtueai. 2023. "Enzymatic Hydrolysis Optimization for Preparation of Sea Cucumber (Holothuria scabra) Hydrolysate with an Antiproliferative Effect on the HepG2 Liver Cancer Cell Line and Antioxidant Properties" International Journal of Molecular Sciences 24, no. 11: 9491. https://doi.org/10.3390/ijms24119491
APA StyleSaiwong, S., Autsavapromporn, N., Siriwoharn, T., Techapun, C., & Wangtueai, S. (2023). Enzymatic Hydrolysis Optimization for Preparation of Sea Cucumber (Holothuria scabra) Hydrolysate with an Antiproliferative Effect on the HepG2 Liver Cancer Cell Line and Antioxidant Properties. International Journal of Molecular Sciences, 24(11), 9491. https://doi.org/10.3390/ijms24119491