Abstract
Enzymes have been used in the food processing industry for many years. However, the use of native enzymes is not conducive to high activity, efficiency, range of substrates, and adaptability to harsh food processing conditions. The advent of enzyme engineering approaches such as rational design, directed evolution, and semi-rational design provided much-needed impetus for tailor-made enzymes with improved or novel catalytic properties. Production of designer enzymes became further refined with the emergence of synthetic biology and gene editing techniques and a plethora of other tools such as artificial intelligence, and computational and bioinformatics analyses which have paved the way for what is referred to as precision fermentation for the production of these designer enzymes more efficiently. With all the technologies available, the bottleneck is now in the scale-up production of these enzymes. There is generally a lack of accessibility thereof of large-scale capabilities and know-how. This review is aimed at highlighting these various enzyme-engineering strategies and the associated scale-up challenges, including safety concerns surrounding genetically modified microorganisms and the use of cell-free systems to circumvent this issue. The use of solid-state fermentation (SSF) is also addressed as a potentially low-cost production system, amenable to customization and employing inexpensive feedstocks as substrate.
1. Introduction
In recent years, advances in synthetic biology have led to the rapid synthesis of bioengineered enzymes with applications in the food industry. Prior to modern molecular tools for enzyme engineering, classical enzyme modification strategies included mutagenesis and directed evolution, and extensive laborious screening to identify desired enzyme activity and functionality. While such approaches are still relevant and used, they have been largely superseded by high throughput gene editing technologies and combinations of synthetic biology for using microbial cells as microbial cell factories for more sustainable healthy food production. The bioengineering of food enzymes via fermentative processes has received particular attention, as it is considered a clean-label processing aid by the food industry.
Fermentation entails metabolic processes by which bacteria, yeast, fungi, or algae use carbohydrate sources to effect changes in substrates to be used for consumption, food additives, or become the source of food or supplements themselves (Figure 1). Many of the changes occurring during fermentation contribute to alterations in flavor, texture, and shelf-life extension, besides health-beneficial effects. However, fermentation technology, for the rapid production of designer molecules ensuing from synthetic biology has gained prominence in recent years. Touted as one of the emerging technology breakthroughs of the fourth industrial revolution, synthetic biology including other associated tools involving AI, bioinformatics, systems biology, and computational biology have paved the way for what is now known as precision fermentation [1]. It is essentially perceived as an advanced fermentation system due to the precise production of specific molecules under very controlled manufacturing processes to maximize the yield of desired products and minimize cost [2]. Unlike traditional fermentation, which includes products such as kimchi, yogurt, and tempeh, precision fermentation relies on more intensive control and manufacturing processes and investment in more specialized technology. Biomass fermentation, on the other hand, involves the cultivation of microorganisms capable of high-yield biomass with high nutritional contents, if targeted for food ingredient usage.
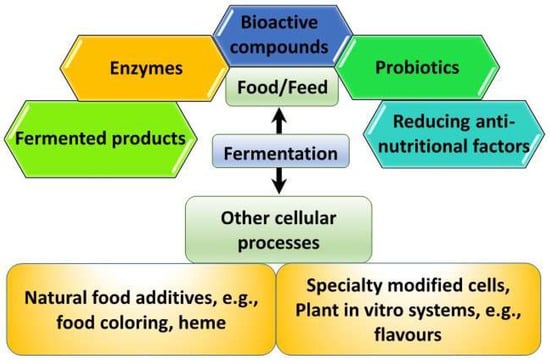
Figure 1.
Fermentation-enabled food valorization potential.
Considering the importance of enzymes in the food industry and the significant roles synthetic biology and precision fermentation are and will be playing in this sector in the foreseeable future, the objective of this review is to highlight the state of bioengineered enzymes in the food industry. Despite its impact on the food industry, enzyme-engineering technology is not without its challenges, and it is important to put those into perspective, especially with regard to safety and scale-up production. With the debate around the use of genetically modified microorganisms (GMO) still ongoing, we discuss gene editing which is likely to assuage some of those safety concerns. More importantly, we put into perspective the use of cell-free systems (CFS) for engineered enzyme production to circumvent the perceived GMO-associated risks. Due to the high costs associated with the manufacture of engineered enzymes by submerged liquid fermentation, be it conventional or precision, we also suggest a rethink of the current strategy of precision fermentation and start focusing on a solid-state precision fermentation (SSPF) strategy, since solid-state fermentation is low-cost, amenable to customization and easily adaptable to the use of inexpensive feedstocks.
2. Strategies and Challenges for the Development of Engineered Enzymes in the Food Industry
The ubiquitous nature of enzymes and their important roles in the alteration of food components cannot be overemphasized due to their multifunctional properties for the improvement of product quality and stability [3]. The major bottleneck in the industrial use of naturally occurring enzymes is generally associated with their low adaptability and stability under harsh food processing conditions (e.g., high/low pH and temperatures) [4,5]. To overcome these limitations, enzyme engineering strategies (Figure 2) have been developed to design new enzymes with improved stability, specificity, and activity while increasing biocatalysis and reducing energy consumption [6,7].
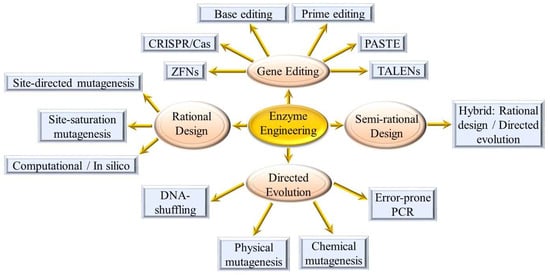
Figure 2.
Enzyme engineering strategies.
Directed evolution and rational design are the two well-established engineering approaches to designing new enzymes with tailor-made biocatalytic properties [8,9]. The selection of the appropriate approach relies on the level of knowledge and understanding of the target enzyme structure and function [10]. The selected best variant is then generally expressed in bacteria (e.g., Escherichia coli, Bacillus, and lactic acid bacteria), filamentous fungi (e.g., Aspergillus), and yeasts (e.g., Pichia pastoris) and then screened for functionality [11,12]. Engineered enzymes such as amylases, xylanases, lipases, proteases, cellulases, and pectinases are utilized in a wide range of food applications such as bakery, dairy, brewery, and confectionary (Table 1). With the advent of advanced gene editing methodologies, further opportunities for enzyme engineering were created. To put these different approaches into perspective, a brief overview is provided in the subsequent sections.

Table 1.
Examples of engineered enzymes and their effects in food applications.
2.1. Directed Evolution
Directed evolution relies on creating mutant libraries via iterative random mutagenesis (using PCR techniques, chemical mutagenesis, UV irradiation, or DNA-shuffling techniques) [12,34,35]. Systematic screening and evaluation processes are required to identify the enzymes having the desired properties [36]. For this approach, knowledge about enzyme structure is not necessary. However, the experimental burden of making and screening a high number of random mutants is highly resource and time intensive [37]. Advances in de novo computational design such as machine learning have accelerated directed evolution by screening in silico combinatorial libraries of mutations to predict the function from the sequence and thus leading to the generation of new enzymes with optimized functions [38,39,40].
For instance, directed evolution coupled with a high-throughput robotic screen was employed to overcome the limitations of naturally obtained amylases such as low pH and high temperature. Using error-prone PCR, novel α-amylase (Novamyl) from Bacillus sp. (TS-25) and Bacillus licheniformis showed increased thermostability and acid pH tolerance [8]. When added to bread dough, these engineered enzymes improved fresh bread quality and delayed staling (by reducing hardness, improving elasticity, and maintaining organoleptic features) [12]. Site-directed evolution also improved the thermostability and acid resistance of α-amylase from Rhizopus oryzae resulting in increased starch hydrolysis (1.61-fold) [9]. In silico approach resulted in α-amylase from Bacillus cereus GL96 with high thermostability (70 °C) and stability over a range of pH from 4 to 11 [13]. Bacillus amyloliquefaciens xylanase (reBaxA50) made by error-prone touchdown PCR increased the catalytic efficiency and the stability under thermal and extreme pH treatment compared to the wild-type [14]. Tailored xylo-oligosaccharides having prebiotic benefits were obtained through the hydrolysis of xylan-rich sources using engineered xylanase [15]. Using classical error-prone PCR, engineered Pseudomonas fluorescens and Penicillium cyclopium lipase were produced with enhanced thermostability and alkali stability which expanded their uses as catalysts in bakery and dairy [16,17]. A new β-galactosidase obtained using error-prone PCR had improved specific activity for lactose hydrolysis in milk processing [18]. Liu et al. [19] developed Bacillus alcalophilus alkaline protease using error-prone PCR with high cold adaptation suitable for cold-temperature food processing. A new transglutaminase enzyme from Streptomyces mobaraensis using site-directed mutagenesis also showed improved thermostability and specific activity for use in different applications such as bakery [20].
2.2. Rational Design
Rational design requires in-depth knowledge of the target enzyme properties (structure, catalytic mechanism, active site, and their distribution to function) [41]. To produce enzymes with desired properties, specific DNA regions encoding specific amino acids (identified using structural analysis and computational modeling) related to the desired activity/functionality are replaced, inserted, or deleted [42]. Site-directed mutagenesis is the most commonly used rational design approach, and it is based on the substitution of a specific amino acid to design enzymes with improved functionality [43,44]. For instance, a serine peptidase from Pseudomonas aeruginosa resulted in improved thermal stability and catalytic efficiency compared to the wild type [21]. Xylanase generated using site-directed mutagenesis enhanced substrate specificity owing to the production of tailor-made xylo-oligosaccharides [22]. In bread making, the use of engineered xylanase improved the performance of wheat flour using a lower dosage compared to the wild-type and resulted in increased bread volumes [45]. Recombinant β-glucanase from Bacillus showed enhanced catalytic efficiency, halostability, and thermostability [23,24]. Modified Candida rugosa lipase isozymes LIP1 showed high catalytic efficiency to produce fatty acid esters and diglycerides, which can be used as food emulsifiers [25].
Site-saturation mutagenesis is another rational method based on the substitution of a specific amino acid with the other 19 possible amino acids. De novo approaches are gaining increased interest due to their ability to create novel enzymes displaying diverse functions that do not occur naturally [46,47]. Computer-aided enzyme simulation enabled to redesign of Cel9A-68 cellulase from Thermobifida fusca with improved cellulase activity. Such features could be extremely relevant for cellulose degradation in the brewery sector [26]. A lipase from P. aeruginosa PAO1 was recently computationally “reverse-engineered” using proline mutations. This mutation enabled the creation of new variants with increased activity and stability to be used for flavor development in dairy products [27,48]. Several studies focused on upgrading xylanase activity due to its relevance in bread making. GH11 xylanase from Neocallimastix patriciarum had improved thermostability and kinetic efficiency [28]. In a recent study, GH11 xylanase from Bacillus sp. strain (T82A) was selected out of 576 strains as the most thermostable variant [29]. GH11 mutated sites from Aspergillus niger had the highest catalytic activity and thermostability due to improved binding affinity of enzyme and substrate [30]. Artificial intelligence tools such as protein “Hallucination”, ProteinMPNN, DenseCPD, and Unsupervised Learning Methods (ULM) could further boost the implementation of de novo features in tailored enzymes to fit conventional and emerging food applications [49,50,51].
2.3. Semi-Rational Design
Semi-rational design is a hybrid approach combining the benefits of rational design and directed evolution [52,53]. This approach uses existing libraries (like rational design) to select a specific promising sequence, but the specific amino acid related to the targeted function is unknown [54,55]. The mutation of single amino acids or a combination of them generates new mutants to be screened and evaluated like in directed evolution to identify the desirable variant in the library [54]. Smart libraries created by semi-rational design contributed to the improvement of the catalytic activity of type II ASNase from Bacillus licheniformis [31]. Type II ASNase can be used to mitigate acrylamide in fried potato products, bakery products, and coffee without affecting the appearance, quality, and taste of the product [56]. However, the low thermal stability of the wild type limits its use. Recently, structural alignment and molecular dynamic simulation demonstrated that new mutants made using semi-rational design had high catalytic efficiency, structural stability, and strong substrate binding [31]. Semi-rational mutagenesis and low-throughput screening also resulted in a double mutant (L341I_Q345S) of sucrose phosphorylase with a selectivity of 95% for kojibiose [32]. Kojibiose has several health benefits including delayed glucose release and a boost in SCFA production. In addition, it can be used as a sweetener in confectionery products [57]. Using a semi-rational design, a new mutant of cellobiose 2-epimerase with improved thermostability and catalytic efficiency was used in the production of lactose-based prebiotics [33].
2.4. Gene Editing
Molecular engineering of enzymes for improved functionality and activity is certainly a more tailored approach compared to screening for mutant cell lines or by directed evolution. However, the advent of more precise genome editing tools (Figure 2) has enabled faster generation of altered gene functions. The conventional restriction enzyme-based recombinant technology has largely been superseded by newer genome-editing tools such as zinc finger nucleases (ZFNs) [58], transcription activator-like effector nucleases (TALENs) [59], Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas9) systems [60], base editing [61], prime editing [62] and Programmable Addition via Site-specific Targeting Elements (PASTE) [63]. With such a repertoire of techniques, the choice of approach to use is dependent on the resources and expertise available. In recent years, a number of gene editing service providers have also emerged, thus allowing for faster results, and circumventing the need for less well-equipped laboratories to invest extensively in establishing such technologies.
While initially much of the efforts were directed towards achieving high levels of gene expression through multiple integrations [64], strong promoters, or inducible promoters [65], the focus has shifted to the gene editing techniques mentioned above to improve functionality and activity. For example, to disrupt proteases and enhance the activities of heterologous expression of pullulanase in Bacillus subtilis strain WS5, CRISPR/Cas9 was used [66]. CRISPR/Cas9 has also been used in fungi due to their high efficiency for enzyme production. To prevent Non-Homologous End Joining (NHEJ) DNA in Penicillium subrubescens and generate a strain capable only of homologous recombination when used as a parental strain for site-specific recombination, CRISPR/Cas9 was used to inactivate the Ku70 gene responsible for NHEJ repair system [67]. CRISPR has also been used to engineer microbes for the production of high-value products such as squalene, which is illegally extracted from shark liver or recovered at low yields from plants. Park et al. [68] were able to engineer Corynebacterium glutamicum using CRISPR interference to produce squalene from glucose by altering multiple key enzymes in the methylerythritol 4-phosphate (MEP) pathway. Heterologously expressed genes, followed by CRISPR/Cas9 editing have also been done to increase enzyme activity. For example, the trehalase enzyme, which catalyzes the conversion of one trehalose molecule to two glucose molecules, was transferred to Aspergillus niger from Myceliophthora sepedonium by homologous recombination and exhibited trehalase activity of 406.44 U/mL [69]. However, upon adopting a multi-copy knock-in expression strategy using CRISPR/Cas9 editing, trehalase expression in Aspergillus niger increased about five-fold compared to the homologously recombined wild-type trehalase gene.
The gene editing technologies are certainly promising in advancing metabolic circuitry alterations for flavor, texture, nutritional, and health beneficial value enhancement. The recent market release of the CRISPR-edited gamma-aminobutyric acid (GABA)-enriched tomato is the first such product in Japan [70]. GABA is promoted as a health supplement and was produced due to the increased activity of the enzyme glutamic acid decarboxylase to convert glutamate to GABA upon suppression of the calmodulin-binding domain gene. Prevention of food losses due to enzymatic browning has also been investigated using gene editing. Enzymatic browning is due to the enzyme polyphenol oxidase (PPO), which catalyzes the oxidation of phenolic compounds to dark-colored quinones. In potato tubers, edited PPO genes led to a 69% reduction in PPO activity and a 73% reduction in enzymatic browning [71]. While these gene-edited crop plants provide powerful new approaches for targeted enzyme modification for in situ expression, the acceptance of these gene-edited crops are still subject to debate. In the meantime, to circumvent these potential roadblocks, engineering enzymes using synthetic biology and gene editing is generally regarded as safe (GRAS) microorganisms for the food industry remains the best strategy.
2.5. Safety Challenges
One of the main safety aspects of developing engineered enzymes relies on ensuring that the used microorganism must qualify as a Qualified Presumption of Safety (QPS) organism and be able to produce a specific food enzyme at a constant level [72]. Even the use of GRAS microorganisms is not without its perceived safety risk over contamination and allergenicity potential. Thus, risk assessment of the safety of the genetic modification includes the evaluation of the source of the enzyme, the introduced sequences, the characteristics of the parental and recipient microorganisms, and the genetic modification process (e.g., the absence of antibiotic resistance genes).
For enzyme engineering, the alteration of amino acid sequences does not typically induce variability in the catalytic site or the identity of the enzyme. Nevertheless, the change of amino acid sequences may increase their allergenic potential. It was reported that genetically engineered enzymes could elicit immediate-type sensitization [73,74]. This urges regular surveillance of commercial enzymes through the development of specific IgE assays. For enzyme manufacturing, protein-engineered enzymes are produced using the same processes as the wild-type, and thus have similar process risks as the wild-type [75]. For the host microorganism, there is a list of strains such as Bacillus subtilis, Bacillus amyloliquefaciens, Bacillus licheniformis, Aspergillus niger, and Aspergillus oryzae with a long history of safe food uses and published in reports and opinions prepared by international organizations such as FAO/WHO, FDA and the European Food Safety Authority (EFSA) [76]. Safety measures and risk assessment procedures are continuously updated by established organizations to ensure the safety of newly engineered enzymes. Genome-editing tools such as CRISPR/Cas9 can precisely and safely target specific changes in microorganisms without introducing exogenous genetic elements [77]. Although the use of CRISPR is legal in the USA, the EU still applies the GMO Directive to the genome-edited organisms. Recently, EFSA concluded in their opinion that the risk assessment methodology and the existing guidelines in the GMO legislation are sufficient to prove the safety of genome-edited organisms [78]. However, the current GMO regulation was developed for organisms produced using established methods of genetic modification and not gene editing. Thus, suitable documentation and guidelines are still needed in the EU to regulate this sector [79].
2.6. Cell-Free Systems to Circumvent GMO Concerns
With the abundance of targets for modification and more recently, the availability of artificial intelligence-based target identification and selection for modification, bioinformatics, systems biology, and computational biology tools [2], there is no limit to the expanse of the gene-editing technology for improving food quality. Irrespective of the debates surrounding GMO safety risk assessments and evolving regulatory frameworks, gene editing for enzyme functionality improvement will continue to be significant in the foreseeable future. In fact, it is likely that cell-free systems will become an alternative to GMOs, thus eliminating the risks and controversies surrounding such microorganisms [80,81]. With the fourth wave of biocatalysis heralded by the advent of advances in molecular biology, synthetic biology, sequencing technologies, gene editing, bioinformatics, and computational biology [82,83], it was the logical next step for considering cell-free systems (CFS) for engineered enzyme production. CFS, is in itself not new, as it has been reported over six decades ago [84], and the availability of a repertoire of platform technologies has provided renewed interest in its potential applications [85]. Although still at an early stage, progress is being made to subsequently develop scalable production and manufacturing platforms. CFS essentially involves the production of a protein in a matrix of cell lysates, crude extracts, or synthetic cocktails in the presence of the transcriptional and translational components outside of an intact cell [86]. Mimicking the biological process outside of the cell offers advantages such as more control and monitoring of reactions, efficient protein folding, and post-translational modifications [87]. Furthermore, as mentioned above, safety risks associated with GMOs are eliminated.
While it is unlikely that CFS will supplant current food enzyme production approaches through conventional or precision fermentation, CFS may offer advantages in certain specific situations in the production of secondary metabolites, proteins, or even vitamins. For example, myo-inositol (Vitamin B8) was produced using a four-enzyme pathway system supplemented with two other enzymes for the conversion of starch into myo-inositol [88]. This non-fermentative production of myo-inositol was successfully demonstrated at a 20,000 L scale, although the enzymes themselves were produced via fermentation [88]. Of note in this study was also the demonstration of myo-inositol production using thermophilic and hyperthermophilic enzymes, which enabled the reactions to be conducted at high (70 °C) temperatures, thus preventing any stability or reaction kinetics hindrance. Using a similar strategy with hyperthermophilic enzymes, sucrose [89] and D-xylose [90] were used as substrates instead of starch for the production of myo-inositol. More recently, pigmented metabolites such as lycopene, indigoidine, betanin, and betaxanthins, which have applications in the food, cosmetic, textile, and pharmaceutical industries, were synthesized by co-expression of three enzymes in the presence of tobacco BY-2 cell suspension lysates [91]. The fact that these are visually discernible metabolites enables monitoring their accumulation during synthesis. The scale-up production of these metabolites is yet to be demonstrated. However, the production of cell lysates from the tobacco BY-2 cell suspension cultures can adequately be performed in large-scale bioreactors just as the large-scale quantities of the enzymes needed to catalyze the reaction.
3. Engineered Enzymes for Improved Plant-Based Beverages and Meat Alternatives
When discussing engineered enzymes in the food industry, it is important to consider their applications in the plant-based beverage and meat alternative context as well. Recent trends indicate an exponential growth in the use of plant proteins. This shift in preference by consumers for more plant-based diets stems from health, sustainability, and/or ethical considerations. However, several plant proteins have nutritional (lacking essential amino acids and low digestibility), functional (e.g., low solubility), and organoleptic (off-flavor and bitter taste) limitations [92]. Thus, there is a need to design enzymes to address the limitations of plant-based ingredients in the food industry due to their increasing demand. The development of new enzymes for debittering can be through engineering new proteases able to cleave plant proteins during extraction at specific sites, thus, limiting the release of low molecular weight peptides containing hydrophobic amino acid. Engineered enzymes could also serve to generate specific plant-based peptides with antioxidant, immunomodulatory, antihypertensive, and antimicrobial activities [93]. The yield and the purity of plant proteins are also closely related to the enzymes used during the extraction. In starch biorefinery, where engineered enzymes such as xylanase, amylase, and pectinase are used to hydrolyze starch, most of the proteins are released as a result of this process [94,95]. Another critical point is plant protein functionality. Engineered enzymes could serve as post-processing aids to improve the solubility of plant proteins and therefore their functionality. Custom-made modifications could expand the uses of plant proteins and further boost their markets beyond soy and pea proteins. For instance, proteins with allergenic and toxic propensities such as in faba and lupin could be reduced through engineered enzymes capable of degrading such proteins to obtain safer and more functional proteins able to compete in the protein market.
The development of engineered enzymes to produce high-quality plant-based beverages is also gaining interest. Engineered α-amylase can catalyze the hydrolysis of starch to obtain simpler and more digestible sugars, thus creating a natural sweetness. These enzymes could modulate the viscosity to obtain plant-based milk with improved texture. Engineered proteases could contribute to hydrolyzing proteins to increase their bioavailability and thus their digestibility. This will boost the nutritional quality of milk alternatives having lower nutritional quality compared to animal-based products. In cheese making, 80% of the currently used rennet is genetically engineered chymosin [96]. These recombinant milk-clotting enzymes have also served in creating plant-based cheeses. For instance, recombinant cardosin can produce large quantities of proteases to be used in making vegan cheese [97]. Precision fermentation has enabled the production of protein, enzymes, fats, and vitamins similar to those present in milk, cheese, and ice cream. Food technology startups such as Formo and Real Deal Milk, are using this technology to deliver equivalent taste, texture, and nutrition as conventional dairy products. Vitamins and minerals lacking in plant sources are being produced using precision fermentation and used for fortifying plant-based beverages to upgrade their nutritional quality.
In meat alternatives, engineered transglutaminases could contribute to improving the texture through the cross-linking of lysine between the γ-glutamyl residues and the ε-amino groups of lysine residues to extend protein chains [98]. Transglutaminase was added in textured protein formulations and showed a significant impact on textural properties such as elasticity and chewiness depending on the dose and the type of proteins [99]. Engineered transglutaminase can modulate the organoleptic properties of textured proteins (low moisture and high moisture). Engineered transglutaminase with specific crosslinking sites could drastically improve the bite and the mouthfeel of the meat alternatives. Engineered proteases could also play a role in improving the taste and contribute to creating an umami flavor such as through the application of the protease, Novozymes Protana®. Amano Enzyme developed a natural protease, Umamizyme™ Pulse, to hydrolyze plant proteins to produce a meaty (umami) flavor in proteins by increasing the glutamic acid and reducing the bitterness. Through protein engineering, the production of such enzymes could be optimized in terms of yield, cost, and sustainability. Established enzyme manufacturers can lead this sector by using their expertise to develop a custom portfolio of enzymes. Advanced computational biology and protein engineering would contribute to the design of new variants with novel functionalities [100]. Precision fermentation enabled the production of “heme” proteins such as soy leghemoglobin using Pichia pastoris and Hemami™ to imitate the flavor and color of meat.
4. Fermentation Scale-Up Challenges
Despite the advances in enzyme engineering, synthetic biology, gene editing, and recently cell-free systems, the limitation to scale-up production and manufacturing remains a significant challenge. Scale-up challenges in fermentation, be it submerged or solid-state, are not new. The technology has been in existence for a long time, but the knowledge base or expertise has been lacking. From a technical perspective, scale-up challenges include process development and optimization by judiciously selecting appropriate growth media, substrates, feedstocks, incubation temperature, and pH, and also considering the complexity of the fermentation process and downstream processing to obtain purified products [101,102]. Although precision fermentation allows for the specific molecule of interest to be produced efficiently, purification strategies can sometimes be still very complex. For many start-ups, feasibility and sustainability studies may not have been conducted a priori, and at the scale-up stage production costs become prohibitive [103]. Furthermore, scale-up facilities such as contract research organizations (CRO) or contract manufacturing organizations (CMO) are still not widely available or accessible.
To address scale-up challenges in fermentation, Ganeshan et al. [101] recently proposed adopting a Process Hazards Analysis (PHA) approach. PHA is essentially taking an approach to fermentation production by applying manufacturing practices widely employed in industrial engineering manufacturing by systematically, comprehensively, and analytically reviewing a process to enable the identification of process and operational hazards and their impacts [104]. In the food industry risk mitigating strategies such as Good Manufacturing Practices (GMP), Good Hygiene Practices (GHP), Quality Management Systems (QMS), and Hazard Analysis and Critical Control Points (HACCP) are already being followed and have been proven to be robust. However, in terms of fermentation, a more holistic approach needs to be followed to mitigate risks at every step of the process.
4.1. Submerged Liquid Fermentation (SmF)
Prior to embarking on scale-up production, it is imperative that a thorough investigation of available resources needs to be conducted. It is often suggested that submerged fermentation (SmF) scale-up be conducted in two stages: a pilot scale from 100–10,000 L and a demonstration scale from 10,000–100,000 L [105]. While this is true, there is always a tendency to expedite processes to achieve scale-up without adequate process development and optimization at smaller scales in the range of 10–50 L, and importantly inadequate technology transfer package development [101]. Consequently, sub-optimal yields are encountered, which contribute to uneconomical and unsustainable production [101]. To some extent, the yields can be addressed due to SmF being adaptable to continuous operation compared to a batch operation. This continuous fermentation process for enzyme production, besides allowing for increased yield, can also lead to cost savings. However, there is a risk of contamination as it involves replenishing the nutrient medium and harvesting at regular intervals.
While initial considerations of the aspects mentioned above are critical, it is important to note that SmF uses significantly large volumes of liquids and to this end, associated downstream processes to concentrate products need to be considered as well. For enzyme scale-up production, this implies an additional cost for purification and concentration, although SmF is preferred because of generally higher yields and lower risks of contamination [106]). With the advent of precision fermentation, yields of enzymes and risks of contamination are not of major concern, although concentration and purification of enzymes from the nutrient broth still need to be addressed.
4.2. Solid-State Fermentation (SSF)
The scale-up challenges thus far discussed are specifically referring to submerged liquid fermentation, be it conventional or precision. Solid-state fermentation (SSF), which has been in existence for centuries, is another option that is relevant in the context of operational cost, resources, and effectiveness. SSF-derived foods such as kimchi, tempeh, and koji are some of the familiar products. Indeed, SSF is amenable to customization and has low operational costs. Due to its significantly reduced moisture requirements (50–60% depending on applications), SSF has tremendous potential for enzyme production. While various small, pilot-scale, and large-scale SSF bioreactors are already available commercially, efforts have been made to develop pilot-scale SSF bioreactors specifically geared toward the production of enzymes. For example, a packed-bed SSF bioreactor was designed for the production of pectinases and lipases from fungi [107]. Requiring minimal water, SSF is well-suited for the use of the substrate itself as a matrix for immobilizing the microbial cells and producing enzymes that can alter the texture, flavor and aroma profiles without the need for purifying enzymes to achieve the desired results; essentially the traditional approach to SSF. Nonetheless, studies have shown that enzymes can be recovered from SSF, although scale-up production still needs to be demonstrated. Using grape pomace and wheat bran as substrate, a mutant strain of Aspergillus niger 3T5B8 was shown to produce hydrolytic enzymes, which were subsequently used as a cocktail for the release of bioactive compounds from grape pomace [108]. Similarly, α-amylase and protease were produced and extracted at a laboratory scale using Rhizopus oryzae and waste bread as substrate [109]. In another study, SSF was used at the laboratory scale and pilot scale (600 L) to produce α-amylase from Aspergillus oryzae and edible oil as substrate [110]. These studies not only demonstrate the potential of SSF for enzyme production, but also for the use of waste feedstocks or residues readily available. With the advent of precision fermentation and the need to move towards more sustainable practices, SSF serves as an ideal platform for further exploring its potential for bioengineered enzyme production and the use of inexpensive feedstocks commonly available. Recently, CRISPR/Cas9-edited genes in yeast were demonstrated to produce higher CO2 for bread leavening and reduced acrylamide in baked bread and potato chips [111]. Although this study was at a laboratory scale, it augurs well for the use of edited enzyme genes in microorganisms to catalyze specific changes in foods via SSF, besides further exploring a solid-state precision fermentation (SSPF) approach for the large-scale production of bioengineered enzymes.
SSF as an economically more sustainable approach for the production of enzymes is not without its limitations. One of its major limitations is the availability of large-scale systems. Also, at larger scales heat transfer and aeration can prevent the optimal growth of microorganisms. Due to the minimal use of moisture, extraction of secreted enzymes can be challenging. Therefore, methods need to be developed to process enzymes out of the solid substrates [112]. To reduce the duration of the SSF, high-density inoculum also needs to be used. Thus, the production of the latter introduces another additional step in the process, which may preclude any cost or time-saving. Therefore, fungal species for the production of enzymes by SSF appear to be more amenable than bacterial species. Furthermore, fungal species are more versatile and effective for the use of agricultural feedstocks as substrates. For example, using agricultural residues, cellulase production at high titres was demonstrated in SSF from Aspergilllus tubingensis NKBP-55 [113]. In fact, many agricultural feedstocks have been used as substrates with a variety of fungal species for the production of enzymes using SSF [112], and as mentioned above with investment of resources SSF can become a more effective approach to enzyme production in the precision fermentation landscape.
5. Concluding Remarks
Microbial food fermentation has been practiced for thousands of years. However, unbeknownst in those days was the fact that many of the textural, aromatic, and flavor profile changes were due to enzyme-catalyzed modifications. Subsequently, the realization that these enzymes could be produced, isolated, and purified at an industrial scale through microbial fermentation, led to the emergence of a completely new food processing approach. However, the efficiency of wild-type enzyme catalytic activity tended to be low. Thus, engineered enzymes for improved catalytic activity became more prevalent. Beyond the traditional rational design, directed evolution, and semi-rational design approaches, the advent of targeted gene editing, AI-based in silico design, and testing approaches further broadened the scope of manipulation of enzymes for improved activity, functionality, stability, and efficiency. However, the bottleneck currently lies in the scale-up production of these enzymes due to limited capacity, know-how, and accessibility. To derive maximum value from the novel enzymes emanating from synthetic biology and precision fermentation, it is imperative that scale-up challenges be addressed. Additionally, the cost of production could be reduced by exploring SSF approaches and inexpensive feedstocks.
Author Contributions
Conceptualization, F.B., S.G., Y.W., M.Ç.T. and M.T.N.; writing—original draft preparation, F.B., S.G., Y.W., M.Ç.T. and M.T.N.; writing—review and editing, F.B., S.G., Y.W., M.Ç.T. and M.T.N.; project administration, S.G.; funding acquisition, M.Ç.T. All authors have read and agreed to the published version of the manuscript.
Funding
Work at the Saskatchewan Food Industry Development Centre is supported by funds from Protein Industries Canada and the Saskatchewan Pulse Growers.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The author (Fatma Boukid) is employed by the company ClonBio Group Ltd. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Hassoun, A.; Bekhit, A.E.; Jambrak, A.R.; Regenstein, J.M.; Chemat, F.; Morton, J.D.; Gudjónsdóttir, M.; Carpena, M.; Prieto, M.A.; Varela, P.; et al. The fourth industrial revolution in the food industry-part II: Emerging food trends. Crit. Rev. Food Sci. Nutr. 2022, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Teng, T.S.; Chin, Y.L.; Chai, K.F.; Chen, W.N. Fermentation for future food systems. EMBO Rep. 2021, 22, e52680. [Google Scholar] [CrossRef] [PubMed]
- Fasim, A.; More, V.S.; More, S.S. Large-scale production of enzymes for biotechnology uses. Curr. Opin. Biotechnol. 2021, 69, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Mesbah, N.M. Industrial Biotechnology Based on Enzymes from Extreme Environments. Front. Bioeng. Biotechnol. 2022, 10, 870083. [Google Scholar] [CrossRef]
- Al-Ghanayem, A.A.; Joseph, B. Current prospective in using cold-active enzymes as eco-friendly detergent additive. Appl. Microbiol. Biotechnol. 2020, 104, 2871–2882. [Google Scholar] [CrossRef]
- Rocha, R.A.; Speight, R.E.; Scott, C. Engineering Enzyme Properties for Improved Biocatalytic Processes in Batch and Continuous Flow. Org. Process. Res. Dev. 2022, 26, 1914–1924. [Google Scholar] [CrossRef]
- Ali, M.; Ishqi, H.M.; Husain, Q. Enzyme engineering: Reshaping the biocatalytic functions. Biotechnol. Bioeng. 2020, 117, 1877–1894. [Google Scholar] [CrossRef]
- Jones, A.; Lamsa, M.; Frandsen, T.P.; Spendler, T.; Harris, P.; Sloma, A.; Xu, F.; Nielsen, J.B.; Cherry, J.R. Directed evolution of a maltogenic α-amylase from Bacillus sp. TS-25. J. Biotechnol. 2008, 134, 325–333. [Google Scholar] [CrossRef]
- Li, S.; Yang, Q.; Tang, B. Improving the thermostability and acid resistance of Rhizopus oryzae α-amylase by using multiple sequence alignment based site-directed mutagenesis. Biotechnol. Appl. Biochem. 2020, 67, 677–684. [Google Scholar] [CrossRef]
- da Silva Amatto, I.V.; da Rosa-Garzon, N.G.; de Oliveira Simões, F.A.; Santiago, F.; da Silva Leite, N.P.; Raspante Martins, J.; Cabral, H. Enzyme engineering and its industrial applications. Biotechnol. Appl. Biochem. 2022, 69, 389–409. [Google Scholar] [CrossRef]
- Seyedhosseini Ghaheh, H.; Sajjadi, S.; Shafiee, F.; Barzegari, E.; Moazen, F.; Sadeghi, H. Rational design of a new variant of Reteplase with optimized physicochemical profile and large-scale production in Escherichia coli. World J. Microbiol. Biotechnol. 2022, 38, 29. [Google Scholar] [CrossRef]
- Ruan, Y.; Zhang, R.; Xu, Y. Directed evolution of maltogenic amylase from Bacillus licheniformis R-53: Enhancing activity and thermostability improves bread quality and extends shelf life. Food Chem. 2022, 381, 132222. [Google Scholar] [CrossRef]
- Pouyan, S.; Lagzian, M.; Sangtarash, M.H. Enhancing thermostabilization of a newly discovered α-amylase from Bacillus cereus GL96 by combining computer-aided directed evolution and site-directed mutagenesis. Int. J. Biol. Macromol. 2022, 197, 12–22. [Google Scholar] [CrossRef]
- Xu, X.; Liu, M.-Q.; Huo, W.-K.; Dai, X.-J. Obtaining a mutant of Bacillus amyloliquefaciens xylanase A with improved catalytic activity by directed evolution. Enzym. Microb. Technol. 2016, 86, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-Q.; Li, J.-Y.; Rehman, A.U.; Xu, X.; Gu, Z.-J.; Wu, R.-C. Laboratory Evolution of GH11 Endoxylanase through DNA Shuffling: Effects of Distal Residue Substitution on Catalytic Activity and Active Site Architecture. Front. Bioeng. Biotechnol. 2019, 7, 350. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Huang, L.; Gui, S.; Zheng, D.; Jia, L.; Fu, Y.; Lu, F. Improvement in thermostability of an alkaline lipase I from Penicillium cyclopium by directed evolution. RSC Adv. 2017, 7, 38538–38548. [Google Scholar] [CrossRef]
- Guan, L.; Gao, Y.; Li, J.; Wang, K.; Zhang, Z.; Yan, S.; Ji, N.; Zhou, Y.; Lu, S. Directed Evolution of Pseudomonas fluorescens Lipase Variants with Improved Thermostability Using Error-Prone PCR. Front. Bioeng. Biotechnol. 2020, 8, 1034. [Google Scholar] [CrossRef] [PubMed]
- Rentschler, E.; Schwarz, T.; Stressler, T.; Fischer, L. Development and validation of a screening system for a β-galactosidase with increased specific activity produced by directed evolution. Eur. Food Res. Technol. 2016, 242, 2129–2138. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, T.; Zhang, Z.; Sun, T.; Wang, J.; Lu, F. Improvement of cold adaptation of Bacillus alcalophilus alkaline protease by directed evolution. J. Mol. Catal. B Enzym. 2014, 106, 117–123. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Shan, M.; Sang, J.; Li, Y.; Jia, L.; Wang, N.; Wang, S.; Shao, S.; Liu, F.; et al. Enhancing the activity and thermostability of Streptomyces mobaraensis transglutaminase by directed evolution and molecular dynamics simulation. Biochem. Eng. J. 2019, 151, 107333. [Google Scholar] [CrossRef]
- Ashraf, N.; Krishnagopal, A.; Hussain, A.; Kastner, D.; Sayed, A.; Mok, Y.K.; Swaminathan, K.; Zeeshan, N. Engineering of serine protease for improved thermostability and catalytic activity using rational design. Int. J. Biol. Macromol. 2018, 126, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Xiong, K.; Xiong, S.; Gao, S.; Li, Q.; Sun, B.; Li, X. Improving Hydrolysis Characteristics of Xylanases by Site-Directed Mutagenesis in Binding-Site Subsites from Streptomyces L10608. Int. J. Mol. Sci. 2018, 19, 834. [Google Scholar] [CrossRef]
- Niu, C.; Zhu, L.; Zhu, P.; Li, Q. Lysine-Based Site-Directed Mutagenesis Increased Rigid β-Sheet Structure and Thermostability of Mesophilic 1,3–1,4-β-Glucanase. J. Agric. Food Chem. 2015, 63, 5249–5256. [Google Scholar] [CrossRef]
- Lee, J.M.; Moon, S.Y.; Kim, Y.-R.; Kim, K.W.; Lee, B.-J.; Kong, I.-S. Improvement of thermostability and halostability of β-1,3-1,4-glucanase by substituting hydrophobic residue for Lys48. Int. J. Biol. Macromol. 2017, 94, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-W.; Huang, M.; Hsieh, Y.-H.; Luo, Y.-T.; Wu, T.-T.; Tsai, C.-W.; Chen, C.-S.; Shaw, J.-F. Simultaneous production of fatty acid methyl esters and diglycerides by four recombinant Candida rugosa lipase’s isozymes. Food Chem. 2014, 155, 140–145. [Google Scholar] [CrossRef]
- Costa, M.G.S.; Silva, Y.F.; Batista, P.R. Computational engineering of cellulase Cel9A-68 functional motions through mutations in its linker region. Phys. Chem. Chem. Phys. 2018, 20, 7643–7652. [Google Scholar] [CrossRef]
- Elatico, A.J.J.; Nellas, R.B. Computational reverse engineering of the lipase from Pseudomonas aeruginosa PAO1: α-helices. J. Mol. Graph. Model. 2020, 100, 107657. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Miao, H.; Ding, J.; Li, J.; Mu, Y.; Zhou, J.; Huang, Z. Improving the thermostability of a fungal GH11 xylanase via site-directed mutagenesis guided by sequence and structural analysis. Biotechnol. Biofuels 2017, 10, 133. [Google Scholar] [CrossRef]
- Takita, T.; Nakatani, K.; Katano, Y.; Suzuki, M.; Kojima, K.; Saka, N.; Mikami, B.; Yatsunami, R.; Nakamura, S.; Yasukawa, K. Increase in the thermostability of GH11 xylanase XynJ from Bacillus sp. strain 41M-1 using site saturation mutagenesis. Enzym. Microb. Technol. 2019, 130, 109363. [Google Scholar] [CrossRef]
- Wu, X.; Tian, Z.; Jiang, X.; Zhang, Q.; Wang, L. Enhancement in catalytic activity of Aspergillus niger XynB by selective site-directed mutagenesis of active site amino acids. Appl. Microbiol. Biotechnol. 2018, 102, 249–260. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiao, L.; Shen, J.; Chi, H.; Lu, Z.; Liu, H.; Lu, F.; Zhu, P. Enhancing the Catalytic Activity of Type II L-Asparaginase from Bacillus licheniformis through Semi-Rational Design. Int. J. Mol. Sci. 2022, 23, 9663. [Google Scholar] [CrossRef] [PubMed]
- Verhaeghe, T.; De Winter, K.; Berland, M.; De Vreese, R.; D’Hooghe, M.; Offmann, B.; Desmet, T. Converting bulk sugars into prebiotics: Semi-rational design of a transglucosylase with controlled selectivity. Chem. Commun. 2016, 52, 3687–3689. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xiao, Y.; Shakhnovich, E.I.; Zhang, W.; Mu, W. Semi-rational design and molecular dynamics simulations study of the thermostability enhancement of cellobiose 2-epimerases. Int. J. Biol. Macromol. 2020, 154, 1356–1365. [Google Scholar] [CrossRef]
- Iqbal, Z.; Sadaf, S. A patent-based consideration of latest platforms in the art of directed evolution: A decade long untold story. Biotechnol. Genet. Eng. Rev. 2022, 38, 133–246. [Google Scholar] [CrossRef] [PubMed]
- Ghahremani Nezhad, N.; Rahman, R.; Normi, Y.; Oslan, S.; Mohd Shariff, F.; Leow, T. Thermostability engineering of industrial enzymes through structure modification. Appl. Microbiol. Biotechnol. 2022, 106, 4845–4866. [Google Scholar] [CrossRef]
- Jang, W.D.; Kim, G.B.; Kim, Y.; Lee, S.Y. Applications of artificial intelligence to enzyme and pathway design for metabolic engineering. Curr. Opin. Biotechnol. 2022, 73, 101–107. [Google Scholar] [CrossRef]
- Freschlin, C.R.; Fahlberg, S.A.; Romero, P.A. Machine learning to navigate fitness landscapes for protein engineering. Curr. Opin. Biotechnol. 2022, 75, 102713. [Google Scholar] [CrossRef]
- Saito, Y.; Oikawa, M.; Sato, T.; Nakazawa, H.; Ito, T.; Kameda, T.; Tsuda, K.; Umetsu, M. Machine-Learning-Guided Library Design Cycle for Directed Evolution of Enzymes: The Effects of Training Data Composition on Sequence Space Exploration. ACS Catal. 2021, 11, 14615–14624. [Google Scholar] [CrossRef]
- Siedhoff, N.E.; Schwaneberg, U.; Davari, M.D. Machine learning-assisted enzyme engineering. In Methods in Enzymology; Tawfik, D.S., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 643, pp. 281–315. [Google Scholar]
- Siedhoff, N.E.; Illig, A.-M.; Schwaneberg, U.; Davari, M.D. PyPEF—An Integrated Framework for Data-Driven Protein Engineering. J. Chem. Inf. Model. 2021, 61, 3463–3476. [Google Scholar] [CrossRef]
- Yan, Q.; Zhang, X.; Chen, Y.; Guo, B.; Zhou, P.; Chen, B.; Huang, Q.; Wang, J.-B. From Semirational to Rational Design: Developing a Substrate-Coupled System of Glucose Dehydrogenase for Asymmetric Synthesis. ACS Catal. 2022, 12, 6746–6755. [Google Scholar] [CrossRef]
- Savino, S.; Desmet, T.; Franceus, J. Insertions and deletions in protein evolution and engineering. Biotechnol. Adv. 2022, 60, 108010. [Google Scholar] [CrossRef]
- Chi, H.; Wang, Y.; Xia, B.; Zhou, Y.; Lu, Z.; Lu, F.; Zhu, P. Enhanced Thermostability and Molecular Insights for L-Asparaginase from Bacillus licheniformis via Structure- and Computation-Based Rational Design. J. Agric. Food Chem. 2022, 70, 14499–14509. [Google Scholar] [CrossRef]
- Suzuki, M.; Date, M.; Kashiwagi, T.; Suzuki, E.; Yokoyama, K. Rational design of a disulfide bridge increases the thermostability of microbial transglutaminase. Appl. Microbiol. Biotechnol. 2022, 106, 4553–4562. [Google Scholar] [CrossRef]
- Leys, S.; Pauly, A.; Delcour, J.A.; Courtin, C.M. Modification of the Secondary Binding Site of Xylanases Illustrates the Impact of Substrate Selectivity on Bread Making. J. Agric. Food Chem. 2016, 64, 5400–5409. [Google Scholar] [CrossRef]
- Wang, X.; Du, J.; Zhao, B.; Wang, H.; Rao, S.; Du, G.; Zhou, J.; Chen, J.; Liu, S. Significantly Improving the Thermostability and Catalytic Efficiency of Streptomyces mobaraenesis Transglutaminase through Combined Rational Design. J. Agric. Food Chem. 2021, 69, 15268–15278. [Google Scholar] [CrossRef]
- Ferreira, P.; Fernandes, P.A.; Ramos, M.J. Modern computational methods for rational enzyme engineering. Chem. Catal. 2022, 2, 2481–2498. [Google Scholar] [CrossRef]
- Mehta, A.; Guleria, S.; Sharma, R.; Gupta, R. The lipases and their applications with emphasis on food industry. In Microbial Biotechnology in Food and Health; Ray, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 143–164. [Google Scholar]
- Anishchenko, I.; Pellock, S.J.; Chidyausiku, T.M.; Ramelot, T.A.; Ovchinnikov, S.; Hao, J.; Bafna, K.; Norn, C.; Kang, A.; Bera, A.K.; et al. De novo protein design by deep network hallucination. Nature 2021, 600, 547–552. [Google Scholar] [CrossRef]
- Schmitt, L.T.; Paszkowski-Rogacz, M.; Jug, F.; Buchholz, F. Prediction of designer-recombinases for DNA editing with generative deep learning. Nat. Commun. 2022, 13, 7966. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, J.Z.H. DenseCPD: Improving the Accuracy of Neural-Network-Based Computational Protein Sequence Design with DenseNet. J. Chem. Inf. Model. 2020, 60, 1245–1252. [Google Scholar] [CrossRef]
- Li, S.; Cao, L.; Yang, X.; Wu, X.; Xu, S.; Liu, Y. Simultaneously optimizing multiple properties of β-glucosidase Bgl6 using combined (semi-)rational design strategies and investigation of the underlying mechanisms. Bioresour. Technol. 2023, 374, 128792. [Google Scholar] [CrossRef]
- Pang, C.; Liu, S.; Zhang, G.; Zhou, J.; Du, G.; Li, J. Improving the catalytic efficiency of Pseudomonas aeruginosa lipoxygenase by semi-rational design. Enzym. Microb. Technol. 2023, 162, 110120. [Google Scholar] [CrossRef]
- Qu, G.; Sun, Z.; Reetz, M.T. Iterative Saturation Mutagenesis for Semi-rational Enzyme Design. In Protein Engineering; Wiley-VCH GmbH: Weinheim, Germany, 2021; pp. 105–132. [Google Scholar]
- Chen, L.; Jiang, K.; Zhou, Y.; Zhu, L.; Chen, X. Improving the Thermostability of α-Glucosidase from Xanthomonas campestris through Proline Substitutions Guided by Semi-rational Design. Biotechnol. Bioprocess. Eng. 2022, 27, 631–639. [Google Scholar] [CrossRef]
- Jia, R.; Wan, X.; Geng, X.; Xue, D.; Xie, Z.; Chen, C. Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives. Microorganisms 2021, 9, 1659. [Google Scholar] [CrossRef]
- Beerens, K.; De Winter, K.; Van de Walle, D.; Grootaert, C.; Kamiloglu, S.; Miclotte, L.; Van de Wiele, T.; Van Camp, J.; Dewettinck, K.; Desmet, T. Biocatalytic Synthesis of the Rare Sugar Kojibiose: Process Scale-Up and Application Testing. J. Agric. Food Chem. 2017, 65, 6030–6041. [Google Scholar] [CrossRef]
- Klug, A. The discovery of zinc fingers and their development for practical applications in gene regulation and genome manipulation. Q. Rev. Biophys. 2010, 43, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Joung, J.K.; Sander, J.D. TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Eleonora, I.I.; Matthew, T.N.Y.; Cian, S.-U.; Rohan, N.K.; Justin, L.; Lukas, V.; Wenyuan, Z.; Kaiyi, J.; Nathaniel, R.; Liyang, Z.; et al. Drag-and-drop genome insertion without DNA cleavage with CRISPR-directed integrases. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lopes, T.S.; Klootwijk, J.; Veenstra, A.E.; van der Aar, P.C.; van Heerikhuizen, H.; Raué, H.A.; Planta, R.J. High-copy-number integration into the ribosomal DNA of Saccharomyces cerevisiae: A new vector for high-level expression. Gene 1989, 79, 199–206. [Google Scholar] [CrossRef]
- Deckers, M.; Deforce, D.; Fraiture, M.A.; Roosens, N.H.C. Genetically Modified Micro-Organisms for Industrial Food Enzyme Production: An Overview. Foods 2020, 9, 326. [Google Scholar] [CrossRef]
- Zhang, K.; Su, L.; Wu, J. Enhanced extracellular pullulanase production in Bacillus subtilis using protease-deficient strains and optimal feeding. Appl. Microbiol. Biotechnol. 2018, 102, 5089–5103. [Google Scholar] [CrossRef]
- Salazar-Cerezo, S.; Kun, R.S.; de Vries, R.P.; Garrigues, S. CRISPR/Cas9 technology enables the development of the filamentous ascomycete fungus Penicillium subrubescens as a new industrial enzyme producer. Enzym. Microb. Technol. 2020, 133, 109463. [Google Scholar] [CrossRef]
- Park, J.; Yu, B.J.; Choi, J.-I.; Woo, H.M. Heterologous Production of Squalene from Glucose in Engineered Corynebacterium glutamicum Using Multiplex CRISPR Interference and High-Throughput Fermentation. J. Agric. Food Chem. 2019, 67, 308–319. [Google Scholar] [CrossRef]
- Dong, L.; Yu, D.; Lin, X.; Wang, B.; Pan, L. Improving expression of thermostable trehalase from Myceliophthora sepedonium in Aspergillus niger mediated by the CRISPR/Cas9 tool and its purification, characterization. Protein Expr. Purif. 2020, 165, 105482. [Google Scholar] [CrossRef]
- Waltz, E. GABA-enriched tomato is first CRISPR-edited food to enter market. Nat. Biotechnol. 2022, 40, 9–11. [Google Scholar] [CrossRef]
- González, M.N.; Massa, G.A.; Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.-S.; Storani, L.; Décima Oneto, C.A.; Hofvander, P.; Feingold, S.E. Reduced Enzymatic Browning in Potato Tubers by Specific Editing of a Polyphenol Oxidase Gene via Ribonucleoprotein Complexes Delivery of the CRISPR/Cas9 System. Front. Plant. Sci. 2020, 10, 1649. [Google Scholar] [CrossRef]
- EFSA Panel on Food Contact Materials, Enzymes, Processing Aids (CEP); Silano, V.; Barat Baviera, J.M.; Bolognesi, C.; Brüschweiler, B.J.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; et al. Safety evaluation of the food enzyme alpha-amylase from a genetically modified Bacillus subtilis (strain NBA). EFSA J. 2019, 17, e05681. [Google Scholar] [CrossRef]
- Budnik, L.T.; Scheer, E.; Burge, P.S.; Baur, X. Sensitising effects of genetically modified enzymes used in flavour, fragrance, detergence and pharmaceutical production: Cross-sectional study. Occup. Environ. Med. 2017, 74, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Kuske, M.; Berndt, K.; Spornraft-Ragaller, P.; Neumeister, V.; Raulf, M.; Sander, I.; Koschel, D.; Bickhardt, J.; Beissert, S.; Bauer, A. Occupational allergy to phytase: Case series of eight production workers exposed to animal feed additives. J. Dtsch. Dermatol. Ges. 2020, 18, 859–865. [Google Scholar] [CrossRef]
- Sewalt, V.; Shanahan, D.; Gregg, L.; La Marta, J.; Carrillo, R. The Generally Recognized as Safe (GRAS) Process for Industrial Microbial Enzymes. Ind. Biotechnol. 2016, 12, 295–302. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards; Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; de Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of qualified presumption of safety (QPS) recommended microorganisms intentionally added to food or feed as notified to EFSA. EFSA J. 2023, 21, e07747. [Google Scholar] [CrossRef]
- Seo, S.-O.; Jin, Y.-S. Next-Generation Genetic and Fermentation Technologies for Safe and Sustainable Production of Food Ingredients: Colors and Flavorings. Annu. Rev. Food Sci. Technol. 2022, 13, 463–488. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Paraskevopoulos, K.; Federici, S. Overview of EFSA and European national authorities’ scientific opinions on the risk assessment of plants developed through New Genomic Techniques. EFSA J. 2021, 19, e06314. [Google Scholar] [CrossRef] [PubMed]
- Rostoks, N. Implications of the EFSA Scientific Opinion on Site Directed Nucleases 1 and 2 for Risk Assessment of Genome-Edited Plants in the EU. Agronomy 2021, 11, 572. [Google Scholar] [CrossRef]
- Brookwell, A.; Oza, J.P.; Caschera, F. Biotechnology Applications of Cell-Free Expression Systems. Life 2021, 11, 1367. [Google Scholar] [CrossRef] [PubMed]
- Rasor, B.J.; Vögeli, B.; Landwehr, G.M.; Bogart, J.W.; Karim, A.S.; Jewett, M.C. Toward sustainable, cell-free biomanufacturing. Curr. Opin. Biotechnol. 2021, 69, 136–144. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. The fourth wave of biocatalysis is approaching. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2017, 376, 20170063. [Google Scholar] [CrossRef]
- Poppe, L.; Vértessy, B.G. The Fourth Wave of Biocatalysis Emerges—The 13th International Symposium on Biocatalysis and Biotransformations. ChemBioChem 2018, 19, 284–287. [Google Scholar] [CrossRef]
- Nirenberg, M.W.; Matthaei, J.H. The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. Natl. Acad. Sci. USA 1961, 47, 1588–1602. [Google Scholar] [CrossRef] [PubMed]
- Silverman, A.D.; Karim, A.S.; Jewett, M.C. Cell-free gene expression: An expanded repertoire of applications. Nat. Rev. Genet. 2020, 21, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Hershewe, J.; Kightlinger, W.; Jewett, M.C. Cell-free systems for accelerating glycoprotein expression and biomanufacturing. J. Ind. Microbiol. Biotechnol. 2020, 47, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Zawada, J.F.; Burgenson, D.; Yin, G.; Hallam, T.J.; Swartz, J.R.; Kiss, R.D. Cell-free technologies for biopharmaceutical research and production. Curr. Opin. Biotechnol. 2022, 76, 102719. [Google Scholar] [CrossRef]
- You, C.; Shi, T.; Li, Y.; Han, P.; Zhou, X.; Zhang, Y.-H.P. An in vitro synthetic biology platform for the industrial biomanufacturing of myo-inositol from starch. Biotechnol. Bioeng. 2017, 114, 1855–1864. [Google Scholar] [CrossRef]
- Zhong, C.; You, C.; Wei, P.; Zhang, Y.-H.P. Thermal Cycling Cascade Biocatalysis of myo-Inositol Synthesis from Sucrose. ACS Catal. 2017, 7, 5992–5999. [Google Scholar] [CrossRef]
- Cheng, K.; Zheng, W.; Chen, H.; Zhang, Y.-H.P.J. Upgrade of wood sugar d-xylose to a value-added nutraceutical by in vitro metabolic engineering. Metab. Eng. 2019, 52, 1–8. [Google Scholar] [CrossRef]
- Buntru, M.; Hahnengress, N.; Croon, A.; Schillberg, S. Plant-Derived Cell-Free Biofactories for the Production of Secondary Metabolites. Front. Plant. Sci. 2022, 12, 794999. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Hickisch, A.; Wani, A.A.; Eisner, P.; Schweiggert-Weisz, U. Enzymatic hydrolysis of lupin protein isolates—Changes in the molecular weight distribution, technofunctional characteristics, and sensory attributes. Food Sci. Nutr. 2019, 7, 2747–2759. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Prongjit, D.; Lekakarn, H.; Bunterngsook, B.; Aiewviriyasakul, K.; Sritusnee, W.; Champreda, V. Functional Characterization of Recombinant Raw Starch Degrading α-Amylase from Roseateles terrae HL11 and Its Application on Cassava Pulp Saccharification. Catalysts 2022, 12, 647. [Google Scholar] [CrossRef]
- Peng, H.; Li, R.; Li, F.; Zhai, L.; Zhang, X.; Xiao, Y.; Gao, Y. Extensive hydrolysis of raw rice starch by a chimeric α-amylase engineered with α-amylase (AmyP) and a starch-binding domain from Cryptococcus sp. S-2. Appl. Microbiol. Biotechnol. 2018, 102, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, Y.; Guan, R.; Jia, G.; Ma, Y.; Zhang, Y. Advances in research on calf rennet substitutes and their effects on cheese quality. Food Res. Int. 2021, 149, 110704. [Google Scholar] [CrossRef] [PubMed]
- Nicosia, F.D.; Puglisi, I.; Pino, A.; Caggia, C.; Randazzo, C.L. Plant Milk-Clotting Enzymes for Cheesemaking. Foods 2022, 11, 871. [Google Scholar] [CrossRef]
- Wen, Y.; Kim, H.W.; Park, H.J. Effects of transglutaminase and cooking method on the physicochemical characteristics of 3D-printable meat analogs. Innov. Food Sci. Emerg. Technol. 2022, 81, 103114. [Google Scholar] [CrossRef]
- Schlangen, M.; Ribberink, M.A.; Taghian Dinani, S.; Sagis, L.M.C.; van der Goot, A.J. Mechanical and rheological effects of transglutaminase treatment on dense plant protein blends. Food Hydrocoll. 2023, 136, 108261. [Google Scholar] [CrossRef]
- Boukid, F.; Hassoun, A.; Zouari, A.; Tülbek, M.Ç.; Mefleh, M.; Aït-Kaddour, A.; Castellari, M. Fermentation for Designing Innovative Plant-Based Meat and Dairy Alternatives. Foods 2023, 12, 1005. [Google Scholar] [CrossRef]
- Ganeshan, S.; Kim, S.H.; Vujanovic, V. Scaling-up production of plant endophytes in bioreactors: Concepts, challenges and perspectives. Bioresour. Bioprocess. 2021, 8, 63. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, G.; Ahmad, T.; Mansoor, S.; Kaur, B. Enzyme Engineering: Current Trends and Future Perspectives. Food Rev. Int. 2021, 37, 121–154. [Google Scholar] [CrossRef]
- Augustin, M.A.; Hartley, C.J.; Maloney, G.; Tyndall, S. Innovation in precision fermentation for food ingredients. Crit. Rev. Food Sci. Nutr. 2023, 1. [Google Scholar] [CrossRef]
- Nolan, D.P. Application of HAZOP and What-If Safety Reviews to the Petroleum, Petrochemical and Chemical Industries; Noyes Publications: Park Ridge, NJ, USA, 1994. [Google Scholar]
- Crater, J.; Lievense, J. Scale-up of Industrial Microbial Processes. FEMS Microbiol. Lett. 2018, 365, 1–5. [Google Scholar] [CrossRef]
- Al-Maqtari, Q.; Al-Ansi, W.; Mahdi, A. Microbial enzymes produced by fermentation and their applications in the food—A review. Int. J. Agric. Innov. Res. 2019, 8, 62–82. [Google Scholar]
- Mitchell, D.A.; Pitol, L.O.; Biz, A.; Finkler, A.T.J.; de Lima Luz, L.F.; Krieger, N. Design and Operation of a Pilot-Scale Packed-Bed Bioreactor for the Production of Enzymes by Solid-State Fermentation. In Solid. State Fermentation: Research and Industrial Applications; Steudler, S., Werner, A., Cheng, J.J., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 27–50. [Google Scholar]
- Teles, A.S.C.; Chávez, D.W.H.; Oliveira, R.A.; Bon, E.P.S.; Terzi, S.C.; Souza, E.F.; Gottschalk, L.M.F.; Tonon, R.V. Use of grape pomace for the production of hydrolytic enzymes by solid-state fermentation and recovery of its bioactive compounds. Food Res. Int. 2019, 120, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Benabda, O.; M’hir, S.; Kasmi, M.; Mnif, W.; Hamdi, M. Optimization of Protease and Amylase Production by Rhizopus oryzae Cultivated on Bread Waste Using Solid-State Fermentation. J. Chem. 2019, 2019, 3738181. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Jeevarathinam, G.; Kumar, S.K.S.; Muniraj, I.; Uthandi, S. Optimization and scale-up of α-amylase production by Aspergillus oryzae using solid-state fermentation of edible oil cakes. BMC Biotechnol. 2021, 21, 33. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-G.; Kim, B.-Y.; Bae, J.-M.; Wang, Y.; Jin, Y.-S. Genome-edited Saccharomyces cerevisiae strains for improving quality, safety, and flavor of fermented foods. Food Microbiol. 2022, 104, 103971. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Gottumukkala, L.D.; Rajasree, K.; Soccol, C.R.; Pandey, A. Solid-state fermentation—Current trends and future prospects. In Fermentation Microbiology and Biotechnology; El-Mansi, E.M.T., El-Mansi, M., Nielsen, J., Mousdale, D.M., Allman, T., Carlson, R., Eds.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Prajapati, B.P.; Kumar Suryawanshi, R.; Agrawal, S.; Ghosh, M.; Kango, N. Characterization of cellulase from Aspergillus tubingensis NKBP-55 for generation of fermentable sugars from agricultural residues. Bioresour. Technol. 2018, 250, 733–740. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).