
Enhanced ZBTB16 Levels by Progestin-Only Contraceptives Induces Decidualization and Inflammation
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Increased Endometrial ZBTB16 Levels in Women Receiving DMPA Treatment
2.2. Elevated Endometrial ZBTB16 Levels in Guinea Pigs Treated with MPA
2.3. Increased ZBTB16 Levels during In Vitro Decidualization of Primary Cultured HESCs
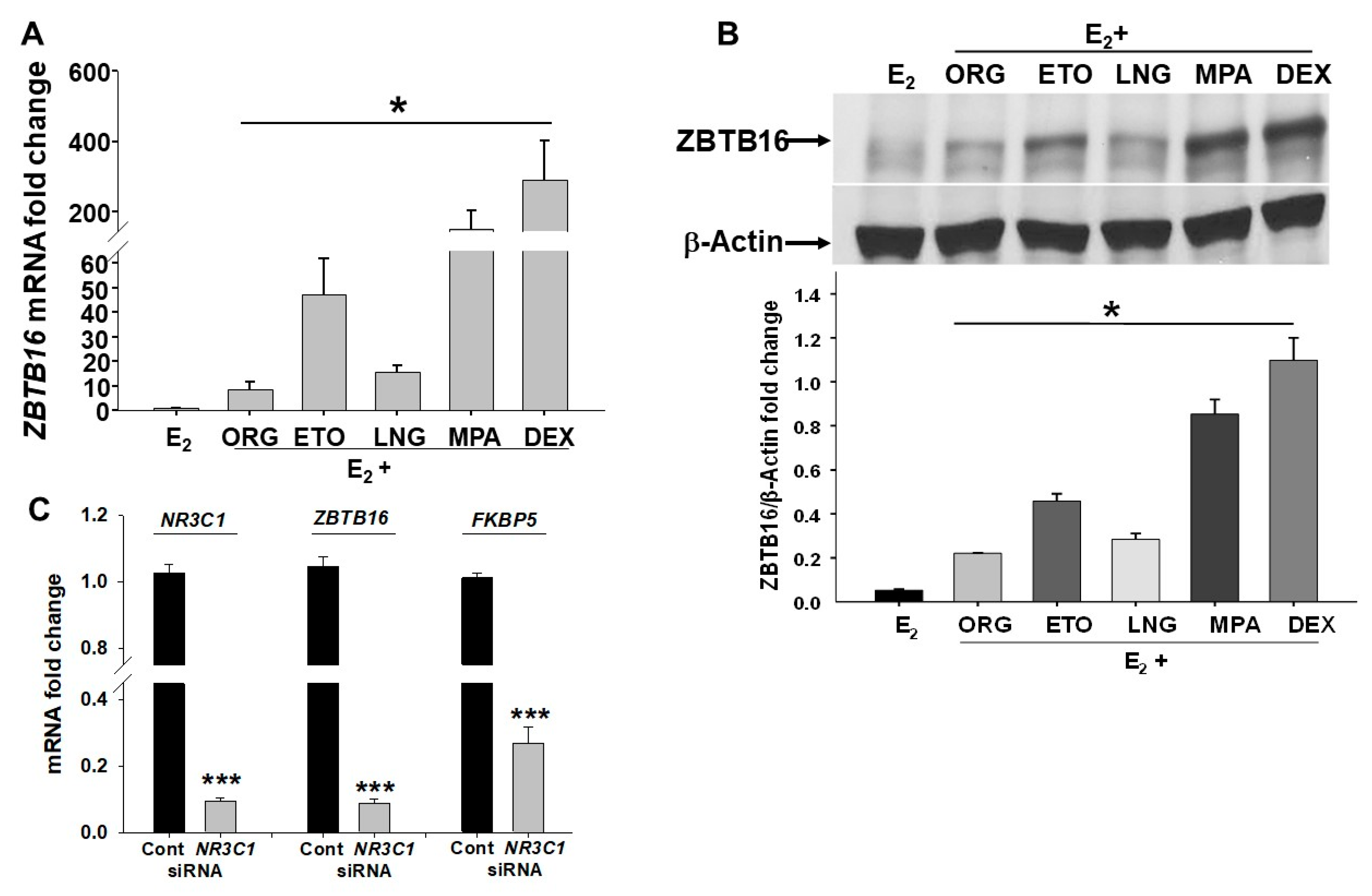
2.4. In Vitro Comparison of Induction of ZBTB16 Levels by Different pLARCs
2.5. ZBTB16 Overexpression Increases Expression of Decidualization Markers in HESCs
2.6. Elevated ZBTB16 in HESCs Induces Tissue Factor Expression
2.7. Elevated ZBTB16 in HESCs Promotes Inflammation by Increasing IL-8 and PTGS2 Expression
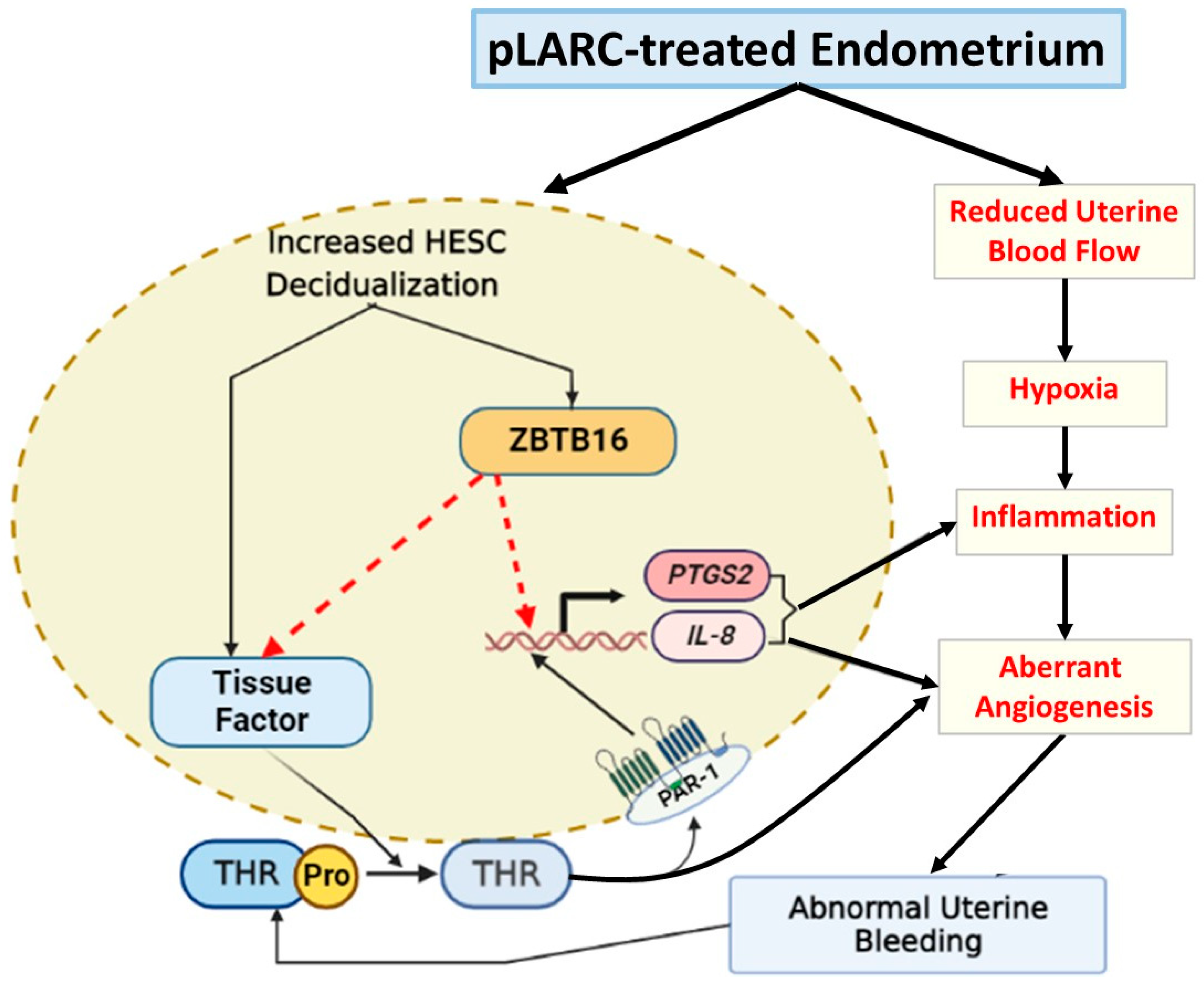
3. Discussion
4. Materials and Methods
4.1. Endometrial Tissue Collection
4.1.1. Human Endometrial Tissue Collection
4.1.2. Guinea Pig Endometrial Tissue Collection
4.1.3. Immunohistochemistry
4.2. Cell Culture
4.2.1. Quantitative Real-Time PCR
4.2.2. Immunoblotting Analyses
4.2.3. Transient Transfection of ZBTB16 Overexpression
4.2.4. siRNA-Mediated Silencing of Glucocorticoid Receptor
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grindlay, K.; Key, K.; Zuniga, C.; Wollum, A.; Grossman, D.; Blanchard, K. Experiences using a progestin-only pill in an over-the-counter environment in the United States: A cross-sectional survey. BMJ Sex. Reprod. Health 2023, 49, 27–34. [Google Scholar] [CrossRef]
- Haddad, L.B.; Herring, G.B.; Mehta, C.C.; Staple, T.; Young, M.R.; Govindaraj, S.; Velu, V.; Smith, A.K. Evaluating the impact of three progestin-based hormonal contraceptive methods on immunologic changes in the female genital tract and systemically (CHIME Study): A prospective cohort study protocol. BMC Womens Health 2022, 22, 456. [Google Scholar] [CrossRef]
- Pieh Holder, K.L. Contraception and Breastfeeding. Clin. Obs. Gynecol. 2015, 58, 928–935. [Google Scholar] [CrossRef]
- Burkett, A.M.; Hewitt, G.D. Progestin only contraceptives and their use in adolescents: Clinical options and medical indications. Adolesc. Med. Clin. 2005, 16, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.S.; Trussell, J.; Schwarz, E.B. Progestin-only contraceptive pill use among women in the United States. Contraception 2012, 86, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Affandi, B. An integrated analysis of vaginal bleeding patterns in clinical trials of Implanon. Contraception 1998, 58, 99S–107S. [Google Scholar]
- Regidor, P.A.; Colli, E.; Palacios, S. Overall and bleeding-related discontinuation rates of a new oral contraceptive containing 4 mg drospirenone only in a 24/4 regimen and comparison to 0.075 mg desogestrel. Gynecol. Endocrinol. 2021, 37, 1121–1127. [Google Scholar] [CrossRef]
- Hickey, M.; Krikun, G.; Kodaman, P.; Schatz, F.; Carati, C.; Lockwood, C.J. Long-term progestin-only contraceptives result in reduced endometrial blood flow and oxidative stress. J. Clin. Endocrinol. Metab. 2006, 91, 3633–3638. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Critchley, H.; Schatz, F.; Wan, L.; Caze, R.; Baergen, R.N.; Lockwood, C.J. Abnormal uterine bleeding during progestin-only contraception may result from free radical-induced alterations in angiopoietin expression. Am. J. Pathol. 2002, 161, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Booth, C.J.; Buchwalder, L.; Schatz, F.; Osol, G.; Mandala, M.; Lockwood, C.J. Effects of etonogestrel treatment in the reproductive organs and uterine arteries of nonoophorectomized guinea pigs. Reprod. Sci. 2012, 19, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Buhimschi, I.A.; Hickey, M.; Schatz, F.; Buchwalder, L.; Lockwood, C.J. Long-term progestin contraceptives (LTPOC) induce aberrant angiogenesis, oxidative stress and apoptosis in the guinea pig uterus: A model for abnormal uterine bleeding in humans. J. Angiogenes Res. 2010, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Schatz, F.; Finlay, T.; Kadner, S.; Mesia, A.; Gerrets, R.; Lockwood, C.J. Expression of angiopoietin-2 by human endometrial endothelial cells: Regulation by hypoxia and inflammation. Biochem. Biophys. Res. Commun. 2000, 275, 159–163. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Basar, M.; Shapiro, J.P.; Semerci, N.; Huang, J.S.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Long-acting progestin-only contraceptives enhance human endometrial stromal cell expressed neuronal pentraxin-1 and reactive oxygen species to promote endothelial cell apoptosis. J. Clin. Endocrinol. Metab. 2014, 99, E1957–E1966. [Google Scholar] [CrossRef]
- Lockwood, C.J.; Runic, R.; Wan, L.; Krikun, G.; Demopolous, R.; Schatz, F. The role of tissue factor in regulating endometrial haemostasis: Implications for progestin-only contraception. Hum. Reprod. 2000, 15 (Suppl. S3), 144–151. [Google Scholar]
- Runic, R.; Schatz, F.; Wan, L.; Demopoulos, R.; Krikun, G.; Lockwood, C.J. Effects of norplant on endometrial tissue factor expression and blood vessel structure. J. Clin. Endocrinol. Metab. 2000, 85, 3853–3859. [Google Scholar] [CrossRef]
- Knezevic, N.; Tauseef, M.; Thennes, T.; Mehta, D. The G protein betagamma subunit mediates reannealing of adherens junctions to reverse endothelial permeability increase by thrombin. J. Exp. Med. 2009, 206, 2761–2777. [Google Scholar] [CrossRef] [Green Version]
- Mhatre, M.V.; Potter, J.A.; Lockwood, C.J.; Krikun, G.; Abrahams, V.M. Thrombin Augments LPS-Induced Human Endometrial Endothelial Cell Inflammation via PAR1 Activation. Am. J. Reprod. Immunol. 2016, 76, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Krikun, G.; Huang, S.T.; Schatz, F.; Salafia, C.; Stocco, C.; Lockwood, C.J. Thrombin activation of endometrial endothelial cells: A possible role in intrauterine growth restriction. Thromb. Haemost. 2007, 97, 245–253. [Google Scholar]
- Lockwood, C.J.; Krikun, G.; Koo, A.B.; Kadner, S.; Schatz, F. Differential effects of thrombin and hypoxia on endometrial stromal and glandular epithelial cell vascular endothelial growth factor expression. J. Clin. Endocrinol. Metab. 2002, 87, 4280–4286. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, C.J.; Kumar, P.; Krikun, G.; Kadner, S.; Dubon, P.; Critchley, H.; Schatz, F. Effects of thrombin, hypoxia, and steroids on interleukin-8 expression in decidualized human endometrial stromal cells: Implications for long-term progestin-only contraceptive-induced bleeding. J. Clin. Endocrinol. Metab. 2004, 89, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, A.P.; Schatz, F.; Krikun, G.; Funai, E.F.; Kadner, S.; Lockwood, C.J. Mechanisms of abruption-induced premature rupture of the fetal membranes: Thrombin enhanced decidual matrix metalloproteinase-3 (stromelysin-1) expression. Am. J. Obs. Gynecol. 2004, 191, 1996–2001. [Google Scholar] [CrossRef]
- Rosen, T.; Schatz, F.; Kuczynski, E.; Lam, H.; Koo, A.B.; Lockwood, C.J. Thrombin-enhanced matrix metalloproteinase-1 expression: A mechanism linking placental abruption with premature rupture of the membranes. J. Matern. Fetal. Neonatal. Med. 2002, 11, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Murk, W.; Kayisli, U.A.; Buchwalder, L.F.; Huang, S.T.; Funai, E.F.; Krikun, G.; Schatz, F. Progestin and thrombin regulate tissue factor expression in human term decidual cells. J. Clin. Endocrinol. Metab. 2009, 94, 2164–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Brand, N.J.; Chen, A.; Chen, S.J.; Tong, J.H.; Wang, Z.Y.; Waxman, S.; Zelent, A. Fusion between a novel Kruppel-like zinc finger gene and the retinoic acid receptor-alpha locus due to a variant t(11;17) translocation associated with acute promyelocytic leukaemia. EMBO J. 1993, 12, 1161–1167. [Google Scholar] [PubMed]
- Cheng, Z.Y.; He, T.T.; Gao, X.M.; Zhao, Y.; Wang, J. ZBTB Transcription Factors: Key Regulators of the Development, Differentiation and Effector Function of T Cells. Front. Immunol. 2021, 12, 713294. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; English, M.A.; Ball, H.J.; Yeyati, P.L.; Waxman, S.; Licht, J.D. Sequence-specific DNA binding and transcriptional regulation by the promyelocytic leukemia zinc finger protein. J. Biol. Chem. 1997, 272, 22447–22455. [Google Scholar] [CrossRef] [Green Version]
- Bardwell, V.J.; Treisman, R. The POZ domain: A conserved protein-protein interaction motif. Genes Dev. 1994, 8, 1664–1677. [Google Scholar] [CrossRef] [Green Version]
- Suliman, B.A.; Xu, D.; Williams, B.R. The promyelocytic leukemia zinc finger protein: Two decades of molecular oncology. Front. Oncol. 2012, 2, 74. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.Q.; Unger, P.; Yang, Q.; Kinoshita, Y.; Singh, K.; McMahon, L.; Nastiuk, K.; Sha, K.; Krolewski, J.; Burstein, D. Loss of PLZF expression in prostate cancer by immunohistochemistry correlates with tumor aggressiveness and metastasis. PLoS ONE 2015, 10, e0121318. [Google Scholar] [CrossRef]
- Gaber, Z.B.; Butler, S.J.; Novitch, B.G. PLZF regulates fibroblast growth factor responsiveness and maintenance of neural progenitors. PLoS Biol. 2013, 11, e1001676. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Frank, G.D.; Ding, R.; Tan, Z.; Rachakonda, A.; Pandolfi, P.P.; Senbonmatsu, T.; Landon, E.J.; Inagami, T. Promyelocytic leukemia zinc finger protein activates GATA4 transcription and mediates cardiac hypertrophic signaling from angiotensin II receptor 2. PLoS ONE 2012, 7, e35632. [Google Scholar] [CrossRef] [Green Version]
- Chauchereau, A.; Mathieu, M.; de Saintignon, J.; Ferreira, R.; Pritchard, L.L.; Mishal, Z.; Dejean, A.; Harel-Bellan, A. HDAC4 mediates transcriptional repression by the acute promyelocytic leukaemia-associated protein PLZF. Oncogene 2004, 23, 8777–8784. [Google Scholar] [CrossRef] [Green Version]
- Barna, M.; Merghoub, T.; Costoya, J.A.; Ruggero, D.; Branford, M.; Bergia, A.; Samori, B.; Pandolfi, P.P. Plzf mediates transcriptional repression of HoxD gene expression through chromatin remodeling. Dev. Cell 2002, 3, 499–510. [Google Scholar]
- Hong, S.H.; David, G.; Wong, C.W.; Dejean, A.; Privalsky, M.L. SMRT corepressor interacts with PLZF and with the PML-retinoic acid receptor alpha (RARalpha) and PLZF-RARalpha oncoproteins associated with acute promyelocytic leukemia. Proc. Natl. Acad. Sci. USA 1997, 94, 9028–9033. [Google Scholar]
- Fahnenstich, J.; Nandy, A.; Milde-Langosch, K.; Schneider-Merck, T.; Walther, N.; Gellersen, B. Promyelocytic leukaemia zinc finger protein (PLZF) is a glucocorticoid- and progesterone-induced transcription factor in human endometrial stromal cells and myometrial smooth muscle cells. Mol. Hum. Reprod. 2003, 9, 611–623. [Google Scholar] [CrossRef]
- Kommagani, R.; Szwarc, M.M.; Vasquez, Y.M.; Peavey, M.C.; Mazur, E.C.; Gibbons, W.E.; Lanz, R.B.; DeMayo, F.J.; Lydon, J.P. The Promyelocytic Leukemia Zinc Finger Transcription Factor Is Critical for Human Endometrial Stromal Cell Decidualization. PLoS Genet. 2016, 12, e1005937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.H.; Cui, L.J.; Li, A.Y.; Zhang, J.P.; Liu, Y.; Zhao, J.S.; Xu, X.B.; He, B.; Wang, J.D.; Chu, L.; et al. Endometrial breakdown with sustained progesterone release involves NF-kappaB-mediated functional progesterone withdrawal in a mouse implant model. Mol. Reprod. Dev. 2016, 83, 780–791. [Google Scholar] [CrossRef]
- Chodankar, R.R.; Murray, A.; Nicol, M.; Whitaker, L.H.R.; Williams, A.R.W.; Critchley, H.O.D. The endometrial response to modulation of ligand-progesterone receptor pathways is reversible. Fertil. Steril. 2021, 116, 882–895. [Google Scholar] [CrossRef]
- Kayisli, U.A.; Basar, M.; Guzeloglu-Kayisli, O.; Semerci, N.; Atkinson, H.C.; Shapiro, J.; Summerfield, T.; Huang, S.J.; Prelle, K.; Schatz, F.; et al. Long-acting progestin-only contraceptives impair endometrial vasculature by inhibiting uterine vascular smooth muscle cell survival. Proc. Natl. Acad. Sci. USA 2015, 112, 5153–5158. [Google Scholar] [CrossRef] [Green Version]
- Guzeloglu Kayisli, O.; Kayisli, U.A.; Basar, M.; Semerci, N.; Schatz, F.; Lockwood, C.J. Progestins Upregulate FKBP51 Expression in Human Endometrial Stromal Cells to Induce Functional Progesterone and Glucocorticoid Withdrawal: Implications for Contraceptive-Associated Abnormal Uterine Bleeding. PLoS ONE 2015, 10, e0137855. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.C.; Jackson, J.A.; Ashmore, J.; Zhu, H.H.; Tseng, L. Regulation of insulin-like growth factor-binding protein-1 synthesis and secretion by progestin and relaxin in long term cultures of human endometrial stromal cells. J. Clin. Endocrinol. Metab. 1991, 72, 1014–1024. [Google Scholar] [CrossRef] [PubMed]
- Helmestam, M.; Lindgren, K.E.; Stavreus-Evers, A.; Olovsson, M. Mifepristone-exposured human endometrial endothelial cells in vitro. Reprod. Sci. 2014, 21, 408–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayisli, U.A.; Luk, J.; Guzeloglu-Kayisli, O.; Seval, Y.; Demir, R.; Arici, A. Regulation of angiogenic activity of human endometrial endothelial cells in culture by ovarian steroids. J. Clin. Endocrinol. Metab. 2004, 89, 5794–5802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krikun, G.; Schatz, F.; Taylor, R.; Critchley, H.O.; Rogers, P.A.; Huang, J.; Lockwood, C.J. Endometrial endothelial cell steroid receptor expression and steroid effects on gene expression. J. Clin. Endocrinol. Metab. 2005, 90, 1812–1818. [Google Scholar] [CrossRef] [Green Version]
- Daniels, K.; Abma, J.C. Current Contraceptive Status among Women Aged 15-49: United States, 2017–2019. In NCHS Data Brief; U.S. Department of Health & Human Services: Washington, DC, USA, 2020; pp. 1–8. [Google Scholar]
- Rosenberg, M.J.; Waugh, M.S.; Long, S. Unintended pregnancies and use, misuse and discontinuation of oral contraceptives. J. Reprod. Med. 1995, 40, 355–360. [Google Scholar]
- Shapiro, J.P.; Guzeloglu-Kayisli, O.; Kayisli, U.A.; Semerci, N.; Huang, S.J.; Arlier, S.; Larsen, K.; Fadda, P.; Schatz, F.; Lockwood, C.J. Thrombin impairs human endometrial endothelial angiogenesis; implications for progestin-only contraceptive-induced abnormal uterine bleeding. Contraception 2017, 95, 592–601. [Google Scholar] [CrossRef]
- Kawano, Y.; Furukawa, Y.; Kawano, Y.; Nasu, K.; Narahara, H. Thrombin-induced chemokine production in endometrial stromal cells. Hum. Reprod. 2011, 26, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Szwarc, M.M.; Hai, L.; Gibbons, W.E.; Peavey, M.C.; White, L.D.; Mo, Q.; Lonard, D.M.; Kommagani, R.; Lanz, R.B.; DeMayo, F.J.; et al. Human endometrial stromal cell decidualization requires transcriptional reprogramming by PLZF. Biol. Reprod. 2018, 98, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, C.J.; Schatz, F. A biological model for the regulation of peri-implantational hemostasis and menstruation. J. Soc. Gynecol. Investig. 1996, 3, 159–165. [Google Scholar] [CrossRef]
- Ujvari, D.; Jakson, I.; Babayeva, S.; Salamon, D.; Rethi, B.; Gidlof, S.; Hirschberg, A.L. Dysregulation of In Vitro Decidualization of Human Endometrial Stromal Cells by Insulin via Transcriptional Inhibition of Forkhead Box Protein O1. PLoS ONE 2017, 12, e0171004. [Google Scholar] [CrossRef] [Green Version]
- Logan, P.C.; Ponnampalam, A.P.; Steiner, M.; Mitchell, M.D. Effect of cyclic AMP and estrogen/progesterone on the transcription of DNA methyltransferases during the decidualization of human endometrial stromal cells. Mol. Hum. Reprod. 2013, 19, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Peppi, M.; Kujawa, S.G.; Sewell, W.F. A corticosteroid-responsive transcription factor, promyelocytic leukemia zinc finger protein, mediates protection of the cochlea from acoustic trauma. J. Neurosci. 2011, 31, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Wang, Z. Identification and characterization of PLZF as a prostatic androgen-responsive gene. Prostate 2004, 59, 426–435. [Google Scholar] [CrossRef]
- Naray-Fejes-Toth, A.; Boyd, C.; Fejes-Toth, G. Regulation of epithelial sodium transport by promyelocytic leukemia zinc finger protein. Am. J. Physiol. Ren. Physiol. 2008, 295, F18–F26. [Google Scholar] [CrossRef] [Green Version]
- Komane, M.; Avenant, C.; Louw-du Toit, R.; Africander, D.J.; Hapgood, J.P. Differential off-target glucocorticoid activity of progestins used in endocrine therapy. Steroids 2022, 182, 108998. [Google Scholar] [CrossRef]
- Sitruk-Ware, R. Pharmacological profile of progestins. Maturitas 2008, 61, 151–157. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Chen, Y.C.; Chen, T.H.; Sue, Y.M.; Cheng, T.H.; Chen, J.R.; Chen, C.H. Far-infrared therapy induces the nuclear translocation of PLZF which inhibits VEGF-induced proliferation in human umbilical vein endothelial cells. PLoS ONE 2012, 7, e30674. [Google Scholar] [CrossRef] [Green Version]
- Rho, S.B.; Choi, K.; Park, K.; Lee, J.H. Inhibition of angiogenesis by the BTB domain of promyelocytic leukemia zinc finger protein. Cancer Lett. 2010, 294, 49–56. [Google Scholar] [CrossRef]
- Joko, T.; Nanba, D.; Shiba, F.; Miyata, K.; Shiraishi, A.; Ohashi, Y.; Higashiyama, S. Effects of promyelocytic leukemia zinc finger protein on the proliferation of cultured human corneal endothelial cells. Mol. Vis. 2007, 13, 649–658. [Google Scholar]
- Hickey, M.; Crewe, J.; Mahoney, L.A.; Doherty, D.A.; Fraser, I.S.; Salamonsen, L.A. Mechanisms of irregular bleeding with hormone therapy: The role of matrix metalloproteinases and their tissue inhibitors. J. Clin. Endocrinol. Metab. 2006, 91, 3189–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, M.; Crewe, J.; Goodridge, J.P.; Witt, C.S.; Fraser, I.S.; Doherty, D.; Christiansen, F.T.; Salamonsen, L.A. Menopausal hormone therapy and irregular endometrial bleeding: A potential role for uterine natural killer cells? J. Clin. Endocrinol. Metab. 2005, 90, 5528–5535. [Google Scholar] [CrossRef] [Green Version]
- Babayev, S.N.; Kanchwala, M.; Xing, C.; Akgul, Y.; Carr, B.R.; Word, R.A. Thrombin Alters Human Endometrial Stromal Cell Differentiation during Decidualization. Reprod. Sci. 2019, 26, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.M.; Chen, C.L.; Tsai, Y.J.; Wu, P.H.; Wu, W.B. Thrombin induces cyclooxygenase-2 expression and prostaglandin E2 release via PAR1 activation and ERK1/2- and p38 MAPK-dependent pathway in murine macrophages. J. Cell Biochem. 2009, 108, 1143–1152. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsiao, L.D.; Shih, Y.F.; Hsu, C.K.; Hu, C.Y.; Yang, C.M. Thrombin Induces COX-2 and PGE2 Expression via PAR1/PKCalpha/MAPK-Dependent NF-kappaB Activation in Human Tracheal Smooth Muscle Cells. Mediat. Inflamm. 2022, 2022, 4600029. [Google Scholar] [CrossRef] [PubMed]
- Vincent-Fabert, C.; Platet, N.; Vandevelde, A.; Poplineau, M.; Koubi, M.; Finetti, P.; Tiberi, G.; Imbert, A.M.; Bertucci, F.; Duprez, E. PLZF mutation alters mouse hematopoietic stem cell function and cell cycle progression. Blood 2016, 127, 1881–1885. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.; Li, N.; Wu, J.; Zheng, L.; Zhai, Y.; Li, B.; Song, W.; Wang, J.; Zhu, H.; Li, G.; et al. PLZF-Induced Upregulation of CXCR4 Promotes Dairy Goat Male Germline Stem Cell Proliferation by Targeting Mir146a. J. Cell Biochem. 2016, 117, 844–852. [Google Scholar] [CrossRef]
- Bernardo, M.V.; Yelo, E.; Gimeno, L.; Campillo, J.A.; Parrado, A. Identification of apoptosis-related PLZF target genes. Biochem. Biophys. Res. Commun. 2007, 359, 317–322. [Google Scholar] [CrossRef]
- Gerart, S.; Siberil, S.; Martin, E.; Lenoir, C.; Aguilar, C.; Picard, C.; Lantz, O.; Fischer, A.; Latour, S. Human iNKT and MAIT cells exhibit a PLZF-dependent proapoptotic propensity that is counterbalanced by XIAP. Blood 2013, 121, 614–623. [Google Scholar] [CrossRef] [Green Version]
- Mao, A.P.; Constantinides, M.G.; Mathew, R.; Zuo, Z.; Chen, X.; Weirauch, M.T.; Bendelac, A. Multiple layers of transcriptional regulation by PLZF in NKT-cell development. Proc. Natl. Acad. Sci. USA 2016, 113, 7602–7607. [Google Scholar] [CrossRef] [Green Version]
- Wasim, M.; Carlet, M.; Mansha, M.; Greil, R.; Ploner, C.; Trockenbacher, A.; Rainer, J.; Kofler, R. PLZF/ZBTB16, a glucocorticoid response gene in acute lymphoblastic leukemia, interferes with glucocorticoid-induced apoptosis. J. Steroid Biochem. Mol. Biol. 2010, 120, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Wasim, M.; Mansha, M.; Kofler, A.; Awan, A.R.; Babar, M.E.; Kofler, R. Promyelocytic leukemia zinc finger protein (PLZF) enhances glucocorticoid-induced apoptosis in leukemic cell line NALM6. Pak. J. Pharm. Sci. 2012, 25, 617–621. [Google Scholar] [PubMed]
- Martin, P.J.; Delmotte, M.H.; Formstecher, P.; Lefebvre, P. PLZF is a negative regulator of retinoic acid receptor transcriptional activity. Nucl. Recept. 2003, 1, 6. [Google Scholar] [CrossRef]
- Schatz, F.; Soderland, C.; Hendricks-Munoz, K.D.; Gerrets, R.P.; Lockwood, C.J. Human endometrial endothelial cells: Isolation, characterization, and inflammatory-mediated expression of tissue factor and type 1 plasminogen activator inhibitor. Biol. Reprod. 2000, 62, 691–697. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arlier, S.; Kayisli, U.A.; Semerci, N.; Ozmen, A.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Guzeloglu-Kayisli, O. Enhanced ZBTB16 Levels by Progestin-Only Contraceptives Induces Decidualization and Inflammation. Int. J. Mol. Sci. 2023, 24, 10532. https://doi.org/10.3390/ijms241310532
Arlier S, Kayisli UA, Semerci N, Ozmen A, Larsen K, Schatz F, Lockwood CJ, Guzeloglu-Kayisli O. Enhanced ZBTB16 Levels by Progestin-Only Contraceptives Induces Decidualization and Inflammation. International Journal of Molecular Sciences. 2023; 24(13):10532. https://doi.org/10.3390/ijms241310532
Chicago/Turabian StyleArlier, Sefa, Umit A. Kayisli, Nihan Semerci, Asli Ozmen, Kellie Larsen, Frederick Schatz, Charles J. Lockwood, and Ozlem Guzeloglu-Kayisli. 2023. "Enhanced ZBTB16 Levels by Progestin-Only Contraceptives Induces Decidualization and Inflammation" International Journal of Molecular Sciences 24, no. 13: 10532. https://doi.org/10.3390/ijms241310532