Abstract
Clubroot is a soil-borne disease caused by Plasmodiophora brassicae, which can seriously affect the growth and production of cruciferous crops, especially Chinese cabbage crops, worldwide. At present, few studies have been conducted on the molecular mechanism of this disease’s resistance response. In this experiment, we analyzed the bioinformation of bra-miR167a, constructed a silencing vector (STTM167a) and an overexpression vector (OE-miR167a), and transformed them to Arabidopsis to confirm the role of miR167a in the clubroot resistance mechanism of Arabidopsis. Afterwards, phenotype analysis and expression level analysis of key genes were conducted on transgenic plants. From the result, we found that the length and number of lateral roots of silence transgenic Arabidopsis STTM167a was higher than that of WT and OE-miR167a. In addition, the STTM167a transgenic Arabidopsis induced up-regulation of disease resistance-related genes (PR1, PR5, MPK3, and MPK6) at 3 days after inoculation. On the other hand, the auxin pathway genes (TIR1, AFB2, and AFB3), which are involved in maintaining the balance of auxin/IAA and auxin response factor (ARF), were down-regulated. These results indicate that bra-miR167a is negative to the development of lateral roots and auxins, but positive to the expression of resistance-related genes. This also means that the STTM167a can improve the resistance of clubroot by promoting lateral root development and the level of auxin, and can induce resistance-related genes by regulating its target genes. We found a positive correlation between miR167a and clubroot disease, which is a new clue for the prevention and treatment of clubroot disease.
1. Introduction
Plasmodiophora brassicae is a soil-borne obligate biotrophic pathogenic protist that infects Brassica crops, including Chinese cabbage, cabbage, radish, kale, and broccoli. During infection, Plasmodiophora brassicae initially invades from the root hairs and infects the root cortex, causing swollen roots [1,2]. These swollen roots begin to hinder the absorption of water and nutrients, leading to abnormal growth [3,4,5]. In addition, with strong resilience, the pathogenic protist may maintain infectivity for 20 years in the soil, and its dormant spores can exist in the soil for a long time. Infection can start at any time during the cropping season and can affect crop growth and development [6,7,8]. This disease causes severe damage to crop quality and yield, resulting in a 10–15% yield loss of cruciferous plants globally [7,9].
Plant microRNAs (miRNAs) are a group of important endogenous small noncoding RNAs, up to 22 nucleotides in length, which regulate different developmental processes in plants. miRNAs are important regulatory molecules that mediate plant immune responses to biological stress [10]. Several miRNAs have been reported to be involved in plant defense against pathogens [11,12]. When plants are attacked by bacteria and fungi, pathogen-associated molecular pattern-triggered immunity and effector-triggered immunity can protect them against infection [13].
Auxin response factor (ARF), a transcription factor, binds to the early responsive elements of auxins, such as the TGTCTC auxin response element, at the promoter, and can mediate auxin signal transduction to regulate plant growth and development [14]. Most ARF proteins have three domains: an N-terminal DNA-binding domain; a C-terminal dimerization domain; and a non-conserved intermediate region, known as a transcriptional activation or repression domain. The expression of downstream auxin-responsive factors is regulated by the binding of ARFs to AuxREs [14,15,16]. ARFs play an important role in plant growth and development. To date, 23 ARFs have been encoded in the model plant A. thaliana. They are involved in the establishment and activity of roots and stem meristems, as well as the formation of embryonic roots, buds, and lateral branches. The function of ARF has also been reported in other plants; ARF6 is involved in the regulation of potato development, with extremely high expression in cells and vascular tissues [17]. CsARF1 plays an important role in the maintenance and release of deep dormancy for overwintering the buds of tea plants, and ARF5 plays an important role in the development of secondary xylem in poplar trees [17]. miR167 and its targets, ARF6 and ARF8, also regulate lateral root and adventitious root development. In Arabidopsis, miR167 negatively regulates the formation of adventitious roots in the plants, while its target gene ARF6/8 positively regulates the formation of adventitious roots in the plants [18]. Meanwhile, ath-miR167 and ARF8 are expressed in the pericycle of roots, where lateral roots emerge and mediate the development of lateral roots [18]. In soybean plants, miR167-GmARF8 is regarded as a key regulatory module of root nodules and lateral development [19].
The miR167 target genes in A. thaliana are ARF6 and ARF8 [19]. Previous studies have shown that miR167 can regulate the reproductive development processes of A. thaliana by regulating the target genes ARF6/ARF8, such as stamen and pistil maturation [20]. In addition, miR167 is also involved in the stress response. miR167 targets BnNRAMP1 in response to cadmium stress in Brassica napus. Deletion of the NRAMP1 gene can increase resistance to cadmium stress in Brassica napus [21]. Based on the target mimicry technique, a short tandem target mimic (STTM) was developed [22]. This technique specifically blocks the expression of target miRNAs without affecting other miRNAs [23]. Currently, STTM technology has been widely used in Arabidopsis, rice, tomato, soybean, and other miRNA function research [24,25,26,27]. For example, the overexpression of STTM160 and STTM165/166 in Arabidopsis [28], rice [29], and cotton [30] inhibit leaf development and increase drought resistance [31]. The introduction of STTM482b into tomatoes can increase disease resistance [32].
Currently, miRNA167 is concentrated in Arabidopsis, rice, tomato, and other model plants, and most studies have focused on the regulation of plant growth and development. However, the role of miRNA167 in the development of clubroot disease and regulation of the target genes ARF6 and ARF8 has not yet been reported. But previous studies have shown that miR167 is related to plant resistance to biotic stress and can regulate auxin levels in plants [33], which means that miR167 may influence plants’ resistance to clubroot by regulating its target gene ARF8 to participate in the process of responding to Plasmodiophora brassicae. In this study, based on the transcript sequencing result of Chinese cabbage, STTM silencing and overexpression (OE) vectors were constructed and transformed into A. thaliana. We analyzed the phenotype and cytology of transgenic A. thaliana to explore the function mechanism of how bra-miR167a influences plants’ resistance to clubroot by impacting the plants’ immune systems and auxin levels. In a word, this study could provide a basis on which to further elucidate the role of bra-miR167a in regulating the response to clubroot disease in Chinese cabbage.
2. Results
2.1. Bioinformatic Analysis of miR167s Reveales the Involvement of Cis-Acting Regulatory Elements
Bioinformatic analysis was performed to preliminarily forecast the biological function of miR167a. There are four precursors of miR167-bra-miR167a/b/c/d. The secondary structures of bra-miR167b and c are the same (Figure 1A). All published sequences of plant miR167 were downloaded from the miRbase database (http://www.miRbase.org/ (accessed on 6 May 2022)) to analyze the conserved bases of miR167. We found that the miR167 family had highly conserved regions at bases 2, 7, 8, and 18 (Figure 1B). A phylogenetic tree was constructed using MEGAX, which showed that pre-miR167a existed both in monocotyledonous and dicotyledonous plants, and bra- miR167a and ath-miR167a were close. This indicates that miR167 is evolutionarily conserved, and that there was high homology between bra- miR167a and ath-miR167a (Figure 1C). To explore the regulatory role of bra-miR167a, we predicted the cis-acting elements in the 2000 bp upstream region of the start codon of miR167a/b/c/d. We found that the bra-miR167 precursor, in addition to containing essential acting elements, also included a jasmonic acid (JA) response element, a plant hormone response element, a stress response element, etc. This means that the expression of bra-miR167a precursors may be induced or inhibited by factors such as light, biotic and abiotic stresses, and hormones (Figure 1D).
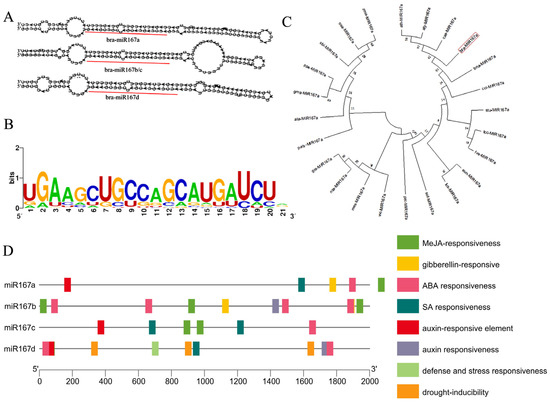
Figure 1.
Bioinformatic analysis of miR167s. (A) Predicted secondary structure of miR167 precursor. (B) Base conservation analysis of miR167 mature sequence. (C) Phylogenetic tree based on miR167a precursor sequence. (D) Cis-elements analysis of cis-regulatory elements in bra-miR167.
2.2. bra-miR167a Negatively Regulates the Expression of Targeted ARF and Auxin
To explore the regulation relationship among bra-miR167a, ARF8, and auxin levels in plants, we analyzed the targeting sites between bra-miR167a and ARF8. The miRNA, transcriptome sequencing, and phytohormone metabolism data were also used to analyze the regulation relationship between bra-miR167a and 31 ARFs genes in susceptible materials at four time periods of disease development. We found that target gene ARF8-b was directly targeted at 2171 bp and that bra-miR167a was bound to ARF8-b in the coding region of the target gene (Supplementary Figure S1A). We found that bra-miR167a was highly expressed at 3 and 9 days after inoculation (DAI), but was low-expressed at 0 and 20 DAI, whereas ARF8 was highly expressed at 0 and 20 DAI and low-expressed at 3 and 9 DAI (Figure 2A,B). bra-miR167a was shown to negatively regulate its target gene, ARF8. The IAA content was lower in the treated group, which was inoculated with clubroot at 3 and 9 DAI, but the IAA content was further increased at 20 DAI. In the control group, which was not inoculated with pathogen, the IAA content increased in the period of 3–9 DAI, but decreased at 9–20 DAI (Figure 2C). The expression level of bra-miR167a was negatively related to IAA, but positively related to ARF8 expression, at different time points after inoculation. In conclusion, bra-miR167a negatively regulate ARF8 gene expression and then auxin content.
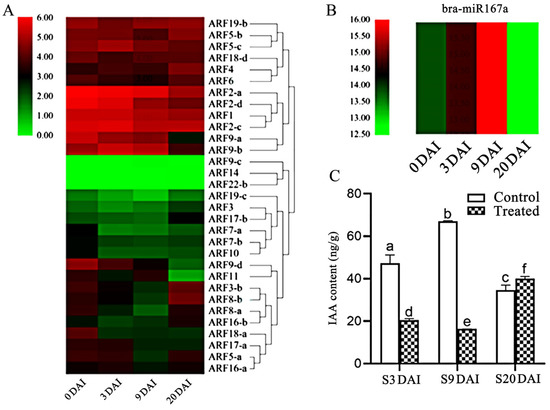
Figure 2.
Analysis of the expression patterns of bra-miR167a and ARFs in Chinese cabbage. (A) The heatmap analysis of ARFs expression; (B) the heatmap analysis of miR167a in susceptible materials at different periods after inoculation. (C) IAA content was measured in the treated group, which was inoculated with clubroot pathogen, and the control group was not inoculated for susceptible materials at different periods. The different lowercase letters indicate significant differences (p < 0.05) based on Duncan’s test.
2.3. STTM167 Negatively and OE-miR167a Positively Regulate the Expression of ARF
To construct the vectors of STTM167a and OE-miR167a, primers were designed for PCR amplification and separated by gel electrophoresis, and the PCR product with a single band and the desired product size was used as a template for synthesizing the sub-clone vector (Supplementary Figure S2A). The amplified STTM167a and OE-miR167a sequences were ligated into the PCAMBIA2301-KY vector at the restriction site KpnI + BamHI (Supplementary Figure S2C). The colony PCR showed that a single band was detected (Supplementary Figure S2B), and the sequencing result showed that the target gene was inserted into the vector (Supplementary Figure S2D).
The transgenic lines were confirmed by PCR amplification using the miRNA167a ST-RT primer [34]. Using quantitative real-time PCR (qRT-PCR), we found that the expression level of bra-miR167a was about 4-fold lower in the STTM167a transgenic plants and 47-fold higher in the OE-miR167a plants than that in the wild type (WT) A. thaliana. However, the expression level of ARF8 increased in the STTM167a transgenic plants, while ARF8 decreased in the OE-miR167a plants (Figure 3). This result was consistent with the result which we obtained previously, which also proved that bra-miR167a was negatively regulated with ARF8. GUS staining was also performed to verify the transgenic A. thaliana. The staining result of the major roots, lateral roots, and leaves in both STTM167a and OE-miR167a indicated successful transformation (Figure 4).
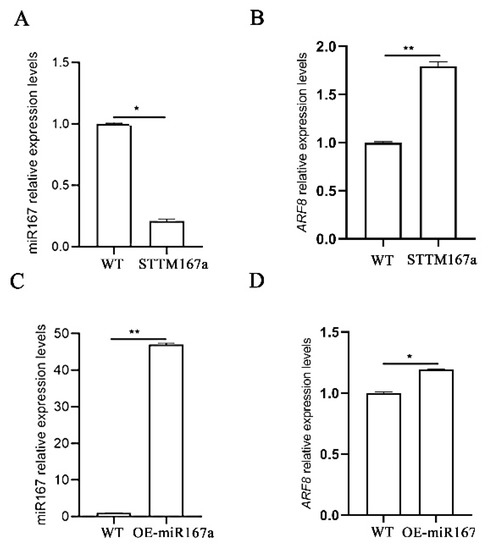
Figure 3.
Expression analysis of miR167a and ARF8 in STTM167a and OE-miR167a. (A) Analysis of miR167a and (B) ARF8 expression analysis in STTM167a plants; (C) analysis of miR167a and (D) ARF/8 expression analysis in OE-miR167a plants. Asterisks (* p < 0.05, ** p < 0.01) indicate significance based on t test (p < 0.05).
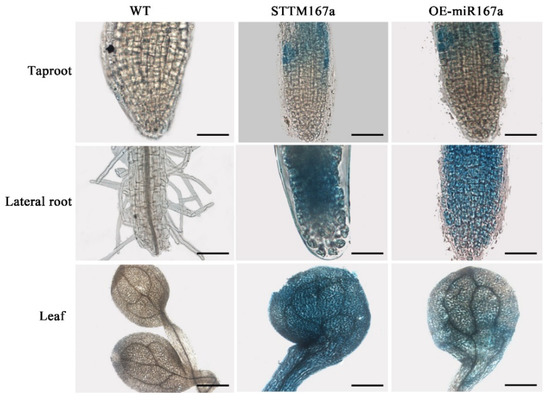
Figure 4.
GUS staining analysis of different tissues of WT, STTM167a, and OE-miR167a transgenic Arabidopsis. bar = 1 mm.
2.4. bra-miR167a Affects Lateral Root Development and Resistance to Clubroot Disease
To determine the role of bra-miR167a in lateral root development, we observed the phenotype in three different periods. From the root phenotype diagram, we can see that WT and STTM had more root development, with a higher number of longer lateral roots. However, OE had obvious main roots and shorter lateral roots, which were fewer in number (Figure 5A,B). To further clarify the specific differences of root development in different transgenic Arabidopsis thaliana varieties, we conducted a statistical analysis of the number and length of lateral roots in different transgenic plants. The results showed that the number of lateral roots of STTM167a was 56% higher and the length of lateral roots was 47% higher than that of WT at 15 d after sowing (DAS). OE-miR167a had half the number of lateral roots in transgenic lines that the WT did at 15 DAS, but the length of the lateral roots was not significantly different from the WT type (Figure 5A–F). This result indicates that low expression of bra-miR167a may contribute to the number and length of lateral roots, while its overexpression it may inhibit the number of lateral roots, but is not related to their length.
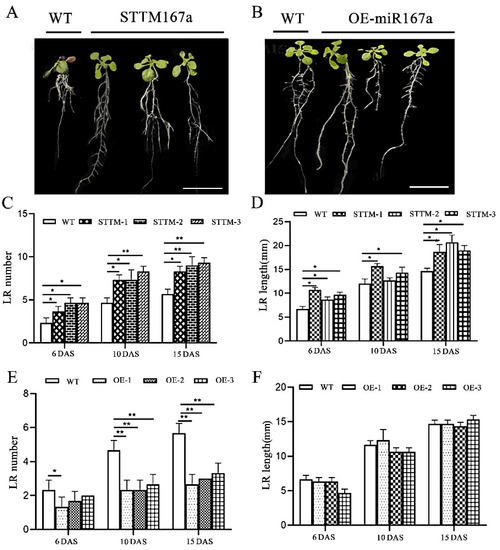
Figure 5.
Lateral root analysis of STTM167a and OE-miR167a transgenic plants. (A): STTM167a and (B): OE-miR167a root phenotype analysis at 20 DAS (bar = 50 mm). (C): STTM167a lateral root number analysis. (D) STTM167a lateral root length analysis. (E) OE-miR167a lateral root number analysis. (F) OE-miR167a lateral root length analysis. Asterisks (* p < 0.05, ** p < 0.01) indicate significance based on t test (p < 0.05).
To investigate the relationship between bra-miR167a and clubroot, we performed phenotypic observation and paraffin section, then calculated the disease index. From the phenotypic picture, there were no obvious differences at 0 DAI between WT, STTM167a, and OE-miR167a, but the roots swelling due to clubroot infection were irregular and varied in size at 30 DAI. The roots of STTM167a did not show significant swelling; OE-miR167a showed significant root swelling at both the lateral and main roots, especially at the junctions of roots and leaves; and WT showed low swelling at both the lateral and main roots (Figure 6A). We also performed paraffin section of root cells at 0 DAI and 30 DAI and found that both WT and OE-miR167a transgenic Arabidopsis root cells swelled irregularly at 30 DAI compared to the uninoculated control (Figure 6C). OE-miR167a was filled with a large number of resting spores, whereas STTM167a transgenic Arabidopsis showed a normal arrangement of root cells with a small quantity of spores (Figure 6C). Furthermore, the disease index of OE-miR167a Arabidopsis was the highest among the WT, STTM167a, and OE-miR167a varieties. The disease index of STTM167a was 58.7% lower than WT and 66.5% lower than OE-miR167a (Figure 6B). These results indicate that transgenic Arabidopsis of STTM167a s effectively reduces the occurrence of clubroot, while transgenic Arabidopsis of OE-miR167a has the opposite function. They also imply that the low expression of bra-miR167a could induce resistance of plants to clubroot.
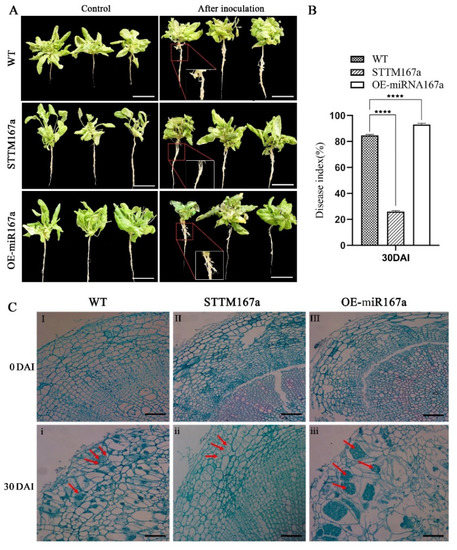
Figure 6.
The disease status of WT, STTM167a, and OE-miR167a Arabidopsis thaliana plants at 30 days after inoculation with P. brassicae (A) Root phenotype analysis of WT, STTM167a, and OE-miR167a Arabidopsis thaliana plants at 30 DAI with P. brassicae. (B) Disease index statistics (bar = 50 μm). (C) Observation of paraffin sections of WT, STTM167a, and OE-miR167a roots at 30 DAI. I and i are the cross-sections of the WT control, both uninoculated and at 30 DAI, respectively; II and ii are STTM167a, both uninoculated and at 30 DAI; III and iii are cross-sections of paraffin sections of the OE-miR167a control, both uninoculated and at 30 DAI, respectively (bar = 50 μm). The red arrow indicates the P. brassicae spores. Asterisks (**** p < 0.0001) indicate significance based on t test (p < 0.05).
2.5. bra-miR167a Regulate the Expression of Disease-Defense-Related Genes of Transgenic Arabidopsis under the Infection of Clubroot
Plants can not only rely on their own structure and some chemical substances to resist pathogen infection and external environmental stress, but can also use resistance genes (R genes) to monitor and identify pathogen effector factors, causing plants to produce hypersensitive responses (HR) to defend against pathogen invasions. R genes can trigger defense responses, including localized cell death (highly sensitive reactions) and acquired resistance, by encoding R proteins, which are related to the detection of pathogens and pests [35]. Three DAI is a critical period for disease resistance in clubroot [36]; thus, to further understand the mechanism of how bra-miR167a effects clubroots of Arabidopsis, four defense-related genes (PR1, PR5, MPK3, and MPK6) were selected in order for us to analyze their expression levels in different transgenic materials at 3 DAI and with the control (mock) treatment. We also detected the expression level of bra-miR167a and its target gene, ARF8, to explore the regulation relationship between bra-miR67a and the critical resistant genes mentioned above. As the result demonstrated, compared to the control (mock), the expression levels of bra-miR167a and ARF8 at 3 DAI were significantly lower. The expression level of bra-miR167a of OE-miR167a was significantly higher than STTM167a, while ARF8 was significantly lower than STTM67a. It indicates that plant could down-regulate miR167a to respond to clubroot stress. Meanwhile, the expression levels of these R genes, PR1, PR5, MPK3, and MPK6, in STTM167a transgenic Arabidopsis were higher than those in the WT and OE-miR167a transgenic Arabidopsis (Figure 7B). The expression levels of PR1, PR5, and MPK3 at 3 DAI were higher than those of the uninoculated control. This indicates that the low expression of bra-miR167a reduced the expression of R genes and then activated the plant defense pathway at 3 DAI. Root cross-sections were strained with toluidine blue to determine how the expression level of bra-miR167a impacted P. brassicae infection at the lateral stage. STTM167a showed few P. brassicae spores, while WT showed obvious spores and OE-miR167a contained numerous enlarged parenchyma cells filled with plasmodia (Figure 7C).
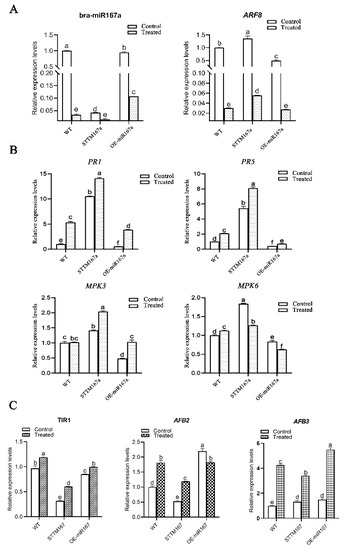
Figure 7.
(A) Expression changes of bra-miR167a and its target gene, ARF8, in different transgenic plants; (B) expression changes of disease-resistance-related genes in different transgenic plants; and (C) expression changes of TIR1, AFB2, and AFB3 in different transgenic plants at 3 DAI. The expression value at 3 DAI of WT of control was normalized as 1. The different lowercase letters indicate significant differences (p < 0.05) based on Duncan’s test.
2.6. Auxin Signaling Pathway Is Effected in OE-miR167a s under Infection with Clubroot
When plants are stressed by pathogens, auxin can promote the degradation of the indole-3-acetic acid (IAA) transcription repressor family and promote the up-regulation of auxin. The transport inhibitor response 1 (TIR1) is an auxin receptor that interacts with IAA protein and can encode the F-box protein that forms IAA [37]. When the concentration of auxin is high, Aux/IAA protein will combine with SCFTIR1/AFB protein complex and release the ARF gene by degrading 26S protease to regulate the transcription of auxin response gene. This then regulates the level of auxin in the plant [38]. Six genes of the TIR1/AFB protein family were totally expressed, but the expression of each TIR1/AFB protein was different in different organs. For example, TIR1, AFB1, and AFB2 were highly expressed in the root [39]. The target gene, ARF8, of bra-miR167a is a key gene in the auxin signaling pathway. To determine how the bra-miR167a regulates the auxin levels of Arabidopsis by regulating its target gene ARF8, we analyzed the expression levels of the crucial genes TIR1 (Bra007720), AFB2 (Bra032954), and AFB3 (Bra026953) of the auxin signaling pathway to assess the mechanism behind the change in the auxin level (Figure 7C). From the result, we found that the expression levels of TIR1, AFB2, and AFB3 in STTM167a transgenic Arabidopsis were significantly lower than those in OE-miR167a transgenic Arabidopsis, and had significantly decreased compared to the control after 3 DAI. This indicates that low expression of bra-miR167a may up-regulate the expression of auxin protein receptors by up-regulating the expression of ARF8 to induce resistance in clubroot.
3. Discussion
miR167 plays an important role in plant stress response, growth, and development. We found that miR167a plays an important role in lateral root development and root swelling in Arabidopsis with clubroot disease. From the bioinformatic analysis, we found that miR167 is conserved during phylogenetic evolution and has the highest homology with Arabidopsis miR167a. By data analysis, we found a negative relationship between miR167a and ARFs, where miR167a negatively regulated ARF8-b and auxin induction (Figure 2).
miR167 also plays a significant role in the response to biotic stresses; for example, miR167 is a positive regulator of nodule and lateral root development in Glycine max, which can directly and positively regulate the number of nodules by inhibiting the target genes GmARF8a and GmARF8b. In rice, Osa-miR167d can negatively regulate resistance to rice blast by regulating its target gene, ARF12. In Arabidopsis, miR167 is involved in defense against the bacterial pathogen Pseudomonas syringae. Overexpression of miR167 results in resistance, inhibition of the auxin response, and stimulation of salicylic acid (SA) signaling [40].
In this study, the silencing vector STTM167a and the OE-miR167a overexpression vector were constructed and transferred into A. thaliana using the Agrobacterium-mediated method. The expression levels of miR167 and its target genes, ARF6 and ARF8, in different tissues of the transgenic plants were analyzed using qRT-PCR. The expression level of bra-miR167a in STTM167a transgenic plants was significantly lower than that in WT plants, the expression of the target gene ARF8 was up-regulated, and the number and length of the lateral roots were significantly increased (Figure 5). However, in OE-miR167a plants, the expression of bra-miR167a was up-regulated, the expression of ARF8 was significantly inhibited, and the number of lateral roots was decreased. This indicated a significant role of bra-miR167a in lateral root development in A. thaliana. Similarly, the up-regulation of bra-miR167a in rice caused defects in adventitious roots [41], and the overexpression of bra-miR167a and mutation of ARF8 lines exhibited a loss of lateral root growth after nitrate stimulation in Arabidopsis [42].
bra-miR167a targets ARF6 and ARF8 as positive regulators of rooting, whereas miR160 targets ARF17 as a negative regulator of rooting [43]. In Arabidopsis, bra-miR167a can also adopt a root structure by targeting IAA-Ala Resistance 3 (IAR3) under osmotic stress, which can hydrolyzes an inactive form of auxin (indole-3-acetic acid [IAA] -alanine) to release IAA in order to promote root development. Under high osmotic stress, bra-miR167a down-regulation causes the up-regulation of IAR3, which promotes lateral root development [44]. This is consistent with our finding that bra-miR167a plays an important role in lateral root development, with an increasing number of lateral roots in STTM167a.
Plants can monitor and identify pathogen effectors by inducing resistance genes and producing a hypersensitive response in order to prevent pathogen invasion [45]. MAPK cascades are key to pathogen invasion in plants. They respond to the coercion process by converting the signals generated from plant cell surface pattern recognition receptors to stress responses [46,47,48]. Among the MAPK cascades, MPK3 and MPK6 respond quickly when biological and non-biological coercion are activated [49]. Pathogenesis-related (PR) genes are important in plants’ responses to stress [50].
Pathogenesis-related (PR) proteins have been recognized as proteins that are strongly induced upon biotic and abiotic stress. Recent progress has revealed that PR1 can activate the immune response by releasing a C-terminal CAPE1 peptide. Moreover, it can enhance the immune response of the host by forming compounds with PR5 [51]. It has been reported that PR1 responds to various stresses, such as drought and Fusarium infection [52]. Based on fluorescence quantitative analysis, we found that the expression levels of PR1, PR5, and MPK3 in STTM167a transgenic Arabidopsis were significantly higher than those in WT and OE-miR167a after 3 d of inoculation (Figure 7B). These results suggest that low expression levels of bra-miR167a could inhibit the development of clubroot disease by inducing resistance genes. Therefore, we suggest that STTM167a transgenic A. thaliana is resistant to clubroot.
In the early stages of pathogen infection, plants transmit and induce disease resistance signals through hormone signal transduction to trigger resistance genes and comprehensively upgrade primary metabolism, energy metabolism, and other metabolic pathways in order to reduce disease severity [53,54]. Transport inhibitor response 1(TIR1), an auxin receptor that interacts with AUX/IAA proteins [37], encodes F-box proteins and forms Aux/IAA complexes. F-box proteins form Aux/IAA-SCFTIR1 complexes with SKP1 and Cullin, which are degraded by the ubiquitin/26S proteasome pathway. Aux/IAA proteins can also bind ARF to inhibit the transcription of specific auxin response genes [55]. Therefore, we analyzed the expression of the Aux/IAA inhibitor protein receptors TIR1, AFB2, and AFB3 and found that the expression of TIR1, AFB2, and AFB3 in STTM167a transgenic Arabidopsis was significantly lower than that in OE-miR167a transgenic Arabidopsis (Figure 7C). The expression levels of TIR1, AFB2, and AFB3 were significantly decreased compared with the control at 3 DAI, indicating that STTM167a increased plant resistance to clubroot disease by down-regulating TIR1, AFB2, and AFB3.
Based on these results, a model of the auxin pathway that regulates the pathogenesis of clubroot disease is proposed in this study. When plants are infected by clubroot, the levels of plant hormones like JA, SA and auxin are up-regulated [36]. Then, the changes in plant hormones may up-regulate the auxin protein genes TIR1/AFB2/AFB3. In OE-miR167a transgenic A. thaliana, the high expression of bra-miR167a negatively regulates the ARF8, and then the low expression of ARF8 promotes the expression of TIR1, AFB2, and AFB3. This inhibits the combination of the auxin/IAA protein with the ARF8 protein and promotes the degradation of auxin, resulting in the up-regulation of auxin-responsive genes and increased plant susceptibility. In STTM167a transgenic Arabidopsis, the low expression of bra-miR167a positively regulates the ARF8, and then the high expression of ARF8 inhibits the expression of TIR1, AFB2, and AFB3, which promotes the combination of the auxin/IAA transcriptional repressor protein with ARF8 and inhibits the expression of auxin-responsive genes. Meanwhile, in STTM167a transgenic Arabidopsis, the resistance-related genes PR1, PR5, MPK3, and MPK6 were induced at 3 DAI, thus also increasing resistance to clubroot disease (Figure 8). From the schematic diagram, we can obviously determine how bra-miR167a regulates the resistance to clubroot disease. Neither bra-miR167a nor ARF8 directly regulates auxin levels in plants. bra-miR167a regulates the auxin level by influencing the expression level of ARF8, while ARF6/8 regulates it by regulating the response genes of auxin and the change in the expression levels of auxin response genes, which, in turn, effect the transcription levels of the crucial genes. In conclusion, this pattern illustrates the role of bra-miR167a in the response to clubroot disease in detail and provides new insights into the control of clubroot disease.
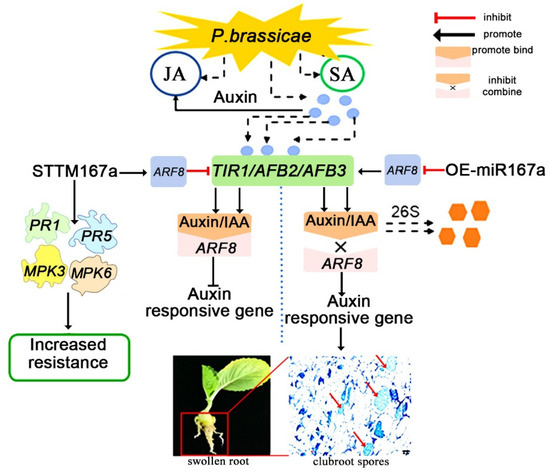
Figure 8.
A schematic diagram of the involvement of auxin in the process of clubroot disease. Auxin response factor 8 (ARF8), pathogenesis-related gene 1 (PR1), pathogenesis-related gene 5 (PR5), map kinase 3 (MPK3), and map kinase 6 (MPK6).
4. Materials and Methods
4.1. Sources of Plant Materials and Transformation Vectors
The material BrT24, which is resistant to P. brassicae, and the material Y510-9, which is susceptible to it [36], were multiplied in the laboratory of leafy vegetables at the Institute of Horticulture, Henan Academy of Agricultural Science, China. The P. brassicae strain used in this study was obtained from a clubroot-infected Chinese cabbage field (B. rapa) in Xinye County, Henan Province, China (113.97° E, 35.05° N), and was identified as race 4 by the Williams system [56]. The seeds of BrT24 and Y510-9 were sown into a 50-well tray and placed in an incubator at 25/20 °C with a 16/8 h light/dark cycle and 60% relative humidity. After 20 days of sowing, 20 mL of protist spore fluid of clubroot was inoculated into the roots in each well, and 20 mL sterile water was also injected into the roots of the control group. The root tissue was sampled at 0, 3, and 20 DAI and stored at −80 °C for subsequent experiments.
Arabidopsis thaliana accession Columbia (Columbia, 2n) was used for gene transformation. Arabidopsis thaliana was first seeded on ½ MS culture plates and, after 10 days, transplanted to a 50-well tray and placed in an incubator at 22/20 °C with a 16/8 h light/dark cycle and 60% relative humidity. After 20 days of sowing, 20 mL of protist spore fluid of clubroot was inoculated into the roots in each well, and 20 mL sterile water was also injected into the roots of the control group. The roots of Arabidopsis thaliana were sampled at 0, 3, and 30 DAI and stored at −80 °C for subsequent experiments.
Competent cells of Escherichia coli DH5α and Agrobacterium GV3101 were purchased from Shanghai Weidi Biological Company (Shanghai, China). The interference and overexpression vectors (pCAMBIA2301-KY (Supplementary Figure S1B)) were provided by Shanghai Kaiyi Biotechnology Co., Ltd. (Shanghai, China).
4.2. Prediction of bra-miR167 Target Gene and Bioinformatic Analyses
All plant precursor sequences were downloaded from the miRBase website (http://www.mirbase.org (accessed on 6 June 2022)) and used to construct a phylogenetic tree of miR167 with MEGAX software. The secondary structure was predicted using the RNAfold website, and the conserved regions of miR167 were analyzed using WebLogo (https://weblogo.berkeley.edu/logo.cgi (accessed on 6 June 2022)). The amino acid sequences of miR167 were obtained from Ensemble Plants (https://plants.ensembl.org/index.html (accessed on 6 June 2022)). The conserved domain of miR167 was analyzed using the CDD (Conserved Domain Database CDD) ((http://www.ncbi.nlm.nih.gov/Structuer/cdd/wrpsb.cgi (accessed on 8 June 2022)) of the National Center for Biotechnology Information. The evolutionary tree and protein conserved domains were visualized using TBtools software [57]. The target sites of bra-miR167a and ARF were analyzed using psRNATarget (http://plantgrn.noble.org/psRNATarget/ (accessed on 8 June 2022)), with all parameters set as the default. A heat map of bra-miR167a and its ARFs was constructed using TBtools, based on the expression obtained from miRNA and the transcriptome sequence results [58], at 0, 3, 9 and 20 DAI.
4.3. Real-Time Quantitative PCR Assay
RNA was extracted using the MiniBEST Plant RNA Extraction Kit (Takara BioTechnology (Dalian, China) Co., Ltd.) from plants, and MonScriptTM miRNA First Strand cDNA Synthesis Kit (Tailing Reaction kit Monad, Suzhou, China) was used for miRNA reverse transcription. MonScriptTM RTIII Super Mix, along with the daDNase kit (Monad, Suzhou, China), were used for mRNA reverse transcription. qRT-PCR analyses were performed in Roche LightCycler 480II (Roche, Basel, Switzerland). The program referred to instructions of the qRCR reagent, and the data were quantified using the comparative 2−∆∆Ct method after the PCR program. The expression analysis of each gene was performed with three biological replicates and three technical replicates. Then, the mean of the technical replicates for each biological replicate were calculated, and these values were used for the subsequent statistical analysis. All of the primers which were utilized are listed in Supplementary Table S1.
4.4. Transformation and Identification of Arabidopsis with STTM167a and OE-miR167a
The STTM vector was constructed by inserting TAG bases into the 10th and 11th bases to silence the mature miRNA sequence. The STTM miR167a intermediate formed a stem–loop structure on its own after being transcribed into RNA, with both ends present as single-stranded complementary portions to miR167a, which binds in a complementary manner to miR167a Synthetic sequences of OE-miR167a and STTM167a target genes were cloned into a pCAMBIA2301-KY (Supplementary Figure S1B) plant expression vector with control of the CaMV35S promoter to generate 35S: bra-miR167a constructs using USERTM cloning technology [59]. These were then transformed, above recombinant constructs, into Escherichia coli DH5α. The 35S: bra-miR167a constructs were transformed into Arabidopsis wild-type (Columbia, 2n) plants, and the oral-dip method was used to obtain OEmiR167a and STTM167a transgenic plants [60].
GUS and qRT-PCR were used to identify WT Arabidopsis and transgenic lines. The root tissues were sampled at 10 DAS for GUS staining. The tissue was treated with acetone (90%) for 20–30 min at room temperature (25 °C) to remove partial chlorophyll, and then the tissue was fixed with ether for 3 min. Afterward, they were washed with phosphate buffer and soaked in GUS staining solution for 8 h at 25–37 °C. Finally, they were transferred into 70% ethanol for decolorization, and this was repeated 2–3 times. The stained samples were observed under a fluorescence microscope and photographed. After 20 days of sowing, qRT-PCR experiments were performed to verify the transform result.
4.5. Preparation of Clubroot Spore Solution and Investigation of Disease Incidence
The swollen roots of Chinese cabbage were weighed according to the ratio of target pathogenic fluid volume:swollen root weight = 10:1. They were taken from a −20 °C refrigerator and poured it into a blender to be crushed. The crushed bacterial liquid was filtered, poured into a 50 mL tube, and centrifuged for 5 min (4500 rpm). The supernatant was discarded, and a small amount of sterile water was added into a centrifuge tube and stirred with a glass rod until the precipitate was well mixed. This operation was repeated three times. The supernatant was discarded the final time, and the volume of the supernatant was fixed to the corresponding volume according to the ratio of swelling weight/g:Aseptic water/mL = 1:10. Finally, the concentration of clubroot spore suspension was calculated using a 25 × 16 blood cell counting plate, and the average value was calculated by five-point counting method. The suspension was diluted to 1 × 107 spores/mL to inoculate Arabidopsis thaliana.
The roots of the plants were washed, and the incidence of disease was critically observed at 30 DAI. STTM167a, OE-miR167a, and WT were examined, each with three plates and one plate with 50 holes, for a total of 450 plants. Based on the size of the galls, they were graded as follows: grade 0 (no nodule in the lateral and main roots), grade 1 (small nodule in the lateral root), grade 2 (small nodule in the main root), grade 3 (nodule in both the lateral and main roots), and grade 4 (nodule in both the lateral and main roots) [61]. The disease index was calculated as follows: DI = (1 n1 + 2 n2 + 3 n3 + 4 n4) × 100/4 Nt (n represents the number of diseased plants/grade, and N represents the total number of diseased plants): DI = 0 (highly resistant), DI < 10 (resistant), 10 ≤ DI < 20 (moderately susceptible), 20 ≤ DI < 50 (susceptible), and DI ≥ 50 (highly susceptible). For each plate, a disease index was calculated, and each line of Arabidopsis thaliana received three disease indexes.
4.6. Paraffin Sectioning
Fresh tissue samples measuring about 1 cm was cut and placed it into a fixative solution. Then, they were taken from the solution, and the tissue was flatted and put into the embedding frame. Thereafter, the samples were placed in a dehydrating box into a dehydrator, wax dipping was performed according a series of treatments (75%—4 h; 85%—2 h; 90%—2 h; 95%—1 h; 100%—1 h; alcohol benzene—10 min; xylene—20 min; −65 °C—3 h). First, the melted wax was placed into the embedding frame before the tissue was removed and also placed into the embedding frame. After cooling on a −20 °C freezing platform, the solidified wax was removed from the embedding frame for flatting. The flattened wax block was sliced with a thickness of 4 μM. Slices were placed on a spreading machine to flatten the tissue, then placed in a 60 °C oven for baking. The baked wax was removed, and the sample was stored at room temperature for later use. Slices were placed into toluidine blue dye solution for about 2–5 min, then washed with water. Finally, microscopic inspection was conducted.
4.7. Data Analysis
The miRNA, transcriptome sequencing, and phytohormone metabolism data analysis methods referred to Xiaochun Wei [36]. All three repeated biological experiments were analyzed using IBM SPSS Statistics 2.6, and one-way or two-way variance analysis (ANOVA) was performed to determine the significance of the statistics. The given value is the average ± standard deviation (SD) for three biological replicates, and the SD values are shown by the error bars in the figures. The p value is corrected using Bonferroni and indicated by * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Moreover, different letters indicate significant differences at p < 0.05.
5. Conclusions
In this study, we explored the molecular mechanism by which bra-miR167a influences resistance to clubroot by bioinformatically analyzing miR167 and investigating the OE-miR167a and STTM167a. From the study’s results, we found that the number of lateral roots of STTM167a was significantly higher than that of OE-miR167a and WT, and the disease index of STTM167a was also significantly lower than that of OE-miR167a and WT, indicating that bra-miR167a was negatively correlated with both the number of lateral roots and resistance to clubroot disease. To further explore this, we detected the expression level of the resistance genes and auxin receptor genes. The results showed that the expression levels of the resistance genes of STTM167a were higher than those of OE-miR167a and WT, while the auxin receptor genes was even lower. This means that the low expression of bra-miR167a is able to up-regulate the resistance genes PR1, PR5, MPK3, and MPK6 and down-regulate the auxin/IAA auxin-inhibitory protein receptor genes TIR1, AFB2, and AFB3 to improve resistance to clubroot disease. In conclusion, bra-miR167a regulates the auxin pathway and responds to the immune system by regulating the target gene ARF8, the homeostasis of plant hormones, and resistance to clubroot disease. This indicates that we may be able to improve plant varieties by silencing or overexpressing the crucial miRNAs of some biological and abiotic stresses. However, our study simply clarified the mechanism of bra-miR167a-ARF8 in P. brassicae infection, but the deep interaction mechanism among miR167a, the immune system, plant hormones, and more miRNAs related to other biological and abiotic stresses should be studied further.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241411850/s1.
Author Contributions
Y.Y. and X.Z. conceived the study. F.W. and X.W. designed the experiments. L.L., S.Y. and H.S. performed the experiments and analyzed the data. Y.Z., Z.X. and Z.W. participated in the qRT-PCR and phenotype identification. R.L. and X.W. drafted the manuscript. U.K.N. and B.T. revised the manuscript. U.K.N. finalized the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation of China (Grant Nos.31801874), China Agricultural Research System (CARS-23-G15), Innovation Team of Henan Academy of Ag-ricultural Sciences (2023TD06), Joint Research on Agricultural Variety Improvement of Henan Province (2022010504), and Programs for Science and Technology Development of Henan Province (232102111105, 232102111114).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are publicly available. The data can be found in the National Center for Biotechnology Information (NCBI) BioProject database under accession numbers PRJNA868821 and PRJNA743585.
Acknowledgments
We thank Yingying Zhang, Life Origin Biotech, Wuhan City, for pre-experiment and data collection for improvement of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jia, H.; Wei, X.; Yang, Y.; Yuan, Y.; Wei, F.; Zhao, Y.; Yang, S.; Yao, Q.; Wang, Z.; Tian, B.; et al. Root RNA-seq analysis reveals a distinct transcriptome landscape between clubroot-susceptible and clubroot-resistant Chinese cabbage lines after Plasmodiophora brassicae infection. Plant Soil 2017, 421, 93–105. [Google Scholar] [CrossRef]
- Ludwig-Müller, J.; Prinsen, E.; Rolfe, S.A.; Scholes, J.D. Metabolism and Plant Hormone Action During Clubroot Disease. J. Plant Growth Regul. 2009, 28, 229–244. [Google Scholar] [CrossRef]
- Djavaheri, M.; Ma, L.; Klessig, D.F.; Mithofer, A.; Gropp, G.; Borhan, H. Mimicking the Host Regulation of Salicylic Acid: A Virulence Strategy by the Clubroot Pathogen Plasmodiophora brassicae. Mol. Plant Microbe Interact. 2019, 32, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Wen, X.U.; Yuan, Y.; Yao, Q.; Zhao, Y.; Wang, Z.; Jiang, W.; Zhang, X. Genome-wide Investigation of micro RNAs and Their Targets in Brassica rapa ssp. pekinensis Root with Plasmodiophora brassicae Infection. Hortic. Plant J. 2016, 2, 209–216. [Google Scholar] [CrossRef]
- Javed, M.A.; Schwelm, A.; Zamani-Noor, N.; Salih, R.; Silvestre Vañó, M.; Wu, J.; González García, M.; Heick, T.M.; Luo, C.; Prakash, P.; et al. The clubroot pathogen Plasmodiophora brassicae: A profile update. Mol. Plant. Pathol. 2023, 24, 89–106. [Google Scholar] [CrossRef]
- Kageyama, K.; Asano, T. Life Cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 2009, 28, 203–211. [Google Scholar] [CrossRef]
- Dixon, G.R. The Occurrence and Economic Impact of Plasmodiophora brassicae and Clubroot Disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Yang, L.; Fang, Z.; Zhang, Y.; Zhuang, M.; Lv, H.; Wang, Y.; Ji, J.; Liu, Y.; Li, Z.; Han, F. Recent advances of disease and stress resistance breeding of cabbage in china. Acta Hortic. Sin. 2020, 47, 1678–1688. [Google Scholar]
- Yuan, Y.; Qin, L.; Su, H.; Yang, S.; Zhang, X. Transcriptome and Coexpression Network Analyses Reveal Hub Genes in Chinese Cabbage (Brassica rapa L. ssp. pekinensis) During Different Stages of Plasmodiophora brassicae Infection. Front. Plant Sci. 2021, 12, 650252. [Google Scholar] [CrossRef]
- Feng, L.; Xia, R.; Liu, Y. Comprehensive Characterization of miRNA and PHAS Loci in the Diploid Strawberry (Fragaria vesca) Genome. Hortic. Plant J. 2019, 5, 255–267. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim Biophys. Acta 2012, 1819, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Gu, X.; Liu, J.; He, Z. Roles of small RNAs in crop disease resistance. Stress Biol. 2021, 1, 6. [Google Scholar] [CrossRef]
- Li, T.; Gonzalez, N.; Inze, D.; Dubois, M. Emerging Connections between Small RNAs and Phytohormones. Trends Plant Sci. 2020, 25, 912–929. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.B.; Hagen, G.; Guilfoyle, T. The roles of auxin response factor domains in auxin-responsive transcription. Plant Cell 2003, 15, 533–543. [Google Scholar] [CrossRef]
- Guilfoyle, T.; Hagen, G.; Ulmasov, T.; Murfett, J. How Does Auxin Turn On Genes? Plant Physiol. 1998, 118, 341–347. [Google Scholar] [CrossRef]
- Jiang, C.; Shen, Y.; Wang, Y.; Zhang, M.; Sun, C. Research Progress of Auxin Response Factor in Plants. Mol. Plant Breed. 2022, 1–8. [Google Scholar]
- Faivre-Rampant, O.; Cardle, L.; Marshall, D.; Viola, R.; Taylor, M.A. Changes in gene expression during meristem activation processes in Solanum tuberosum with a focus on the regulation of an auxin response factor gene. J. Exp. Bot. 2004, 55, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; Macpherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.A.; Chua, N.H. IAA-Ala Resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef]
- Wu, M.F.; Tian, Q.; Reed, J.W. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 2006, 133, 4211–4218. [Google Scholar] [CrossRef]
- Ru, P.; Xu, L.; Ma, H.; Huang, H. Plant fertility defects induced by the enhanced expression of microRNA167. Cell Res. 2006, 16, 457–465. [Google Scholar] [CrossRef]
- Kun, T.S. Functional Identificaiotn of Brassica Napus miR167 and Its Target under Cadmium Stress. Master Thesis, Nanjing Agricultural University, Nanjing, China, 2016. [Google Scholar]
- Wang, Y.; Li, K.; Chen, L.; Zou, Y.; Liu, H.; Tian, Y.; Li, D.; Wang, R.; Zhao, F.; Ferguson, B.J.; et al. MicroRNA167-Directed Regulation of the Auxin Response Factors GmARF8a and GmARF8b Is Required for Soybean Nodulation and Lateral Root Development. Plant Physiol. 2015, 168, 984–999. [Google Scholar] [CrossRef]
- Yan, J.; Gu, Y.; Jia, X.; Kang, W.; Pan, S.; Tang, X.; Chen, X.; Tang, G. Effective small RNA destruction by the expression of a short tandem target mimic in Arabidopsis. Plant Cell 2012, 24, 415–427. [Google Scholar] [CrossRef]
- Jia, X.; Ding, N.; Fan, W.; Yan, J.; Gu, Y.; Tang, X.; Li, R.; Tang, G. Functional plasticity of miR165/166 in plant development revealed by small tandem target mimic. Plant Sci. 2015, 233, 11–21. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Yan, J.; Gou, F.; Mao, Y.; Tang, G.; Botella, J.R.; Zhu, J.K. Short tandem target mimic rice lines uncover functions of miRNAs in regulating important agronomic traits. Proc. Natl. Acad. Sci. USA 2017, 114, 5277–5282. [Google Scholar] [CrossRef]
- Damodharan, S.; Zhao, D.; Arazi, T. A common miRNA160-based mechanism regulates ovary patterning, floral organ abscission and lamina outgrowth in tomato. Plant J. 2016, 86, 458–471. [Google Scholar] [CrossRef]
- Nizampatnam, N.R.; Schreier, S.J.; Damodaran, S.; Adhikari, S.; Subramanian, S. microRNA160 dictates stage-specific auxin and cytokinin sensitivities and directs soybean nodule development. Plant J. 2015, 84, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Gu, Y.; Zhang, Z.; Tang, G. The interaction between miR160 and miR165/166 in the control of leaf development and drought tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef]
- Chen, J.F.; Zhao, Z.X.; Li, Y.; Li, T.T.; Zhu, Y.; Yang, X.M.; Zhou, S.X.; Wang, H.; Zhao, J.Q.; Pu, M.; et al. Fine-tuning roles of Osa-miR159a in rice immunity against Magnaporthe oryzae and development. Rice 2021, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Huang, C.; Li, F.; Zhou, X. A versatile system for functional analysis of genes and microRNAs in cotton. Plant Biotechnol. J. 2014, 12, 638–649. [Google Scholar] [CrossRef]
- Liu, X.; Liu, S.; Wang, R.; Chen, X.; Zhou, T. Analyses of MiRNA Functions in Maize Using a Newly Developed ZMBJ-CMV-2bN81-STTM Vector. Front. Plant Sci. 2019, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Meng, J.; Cui, J.; Sun, G.; Luan, Y. Function identification of miR482b, a negative regulator during tomato resistance to Phytophthora infestans. Hortic. Res. 2018, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, S.; Xie, H. Advances in the regulation of plant development and stress response by miR167. FBL 2021, 26, 655–665. [Google Scholar]
- Caifu, C.; Ridzon, D.A.; Broomer, A.J.; Zhaohui, Z.; Lee, D.H.; Nguyen, J.T.; Maura, B.; Lan, X.N.; Mahuvakar, V.R.; Andersen, M.R. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 30, e179. [Google Scholar]
- Wu, D.; Cai, C.; Wei, G.; Xiang, Y. Genome-Wide Analysis of NBS-Encoding Disease Resistance Genes in Populus trichocarpa. China Sci. 2009, 45, 152–157. [Google Scholar]
- Wei, X.; Liao, R.; Zhang, X.; Zhao, Y.; Xie, Z.; Yang, S.; Su, H.; Wang, Z.; Zhang, L.; Tian, B.; et al. Integrative Transcriptome, miRNAs, Degradome, and Phytohormone Analysis of Brassica rapa L. in Response to Plasmodiophora brassicae. Int. J. Mol. Sci. 2023, 24, 2414. [Google Scholar] [CrossRef]
- Kepinski, S.; Leyser, O. The Arabidopsis F-box protein TIR1 is an Auxin receptor. Nature 2005, 435, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Li, W.; Zhu, Y.; Liu, Z.; Huang, W.; Zhan, J. Genome-wide identification, characterization and expression analysis of the auxin response factor gene family in Vitis vinifera. Plant Cell Rep. 2014, 33, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Matthieu, P.; Nikita, K.; Yi, Z.; Kathleen, G.; Whitnie, S.; Kumar, P.B.; Arvind, B.R.; Bennett, M.J.; Wolfgang, B.; et al. Genetic analysis of the Arabidopsis TIR1/AFB auxin receptors reveals both overlapping and specialized functions. eLife 2020, 9, e54740. [Google Scholar] [CrossRef]
- Caruana, J.C.; Dhar, N.; Raina, R. Overexpression of Arabidopsis microRNA167 induces salicylic acid-dependent defense against Pseudomonas syringae through the regulation of its targets ARF6 and ARF8. Plant Direct 2020, 4, e00270. [Google Scholar] [CrossRef]
- Meng, Y.; Huang, F.; Shi, Q.; Cao, J.; Chen, D.; Zhang, J.; Ni, J.; Wu, P.; Chen, M. Genome-wide survey of rice microRNAs and microRNA–target pairs in the root of a novel auxin-resistant mutant. Planta 2009, 230, 883–898. [Google Scholar] [CrossRef]
- Gifford, M.L.; Dean, A.; Gutierrez, R.A.; Coruzzi, G.M.; Birnbaum, K.D. Cell-specific nitrogen responses mediate developmental plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Bussell, J.D.; Pacurar, D.I.; Schwambach, J.; Pacurar, M.; Bellini, C. Phenotypic plasticity of adventitious rooting in Arabidopsis is controlled by complex regulation of AUXIN RESPONSE FACTOR transcripts and microRNA abundance. Plant Cell 2009, 21, 3119–3132. [Google Scholar] [CrossRef] [PubMed]
- Couzigou, J.-M.; Combier, J.-P. Plant microRNAs: Key regulators of root architecture and biotic interactions. New Phytol. 2016, 212, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Das, A.; Thakur, S.; Jalali, B. Recent Understanding on Structure, Function and Evolution of Plant Disease Resistance Genes. Proc. Indian Natl. Sci. Acad. 2015, 80, 83–93. [Google Scholar] [CrossRef]
- Bi, G.; Zhou, J.M. MAP Kinase Signaling Pathways: A Hub of Plant-Microbe Interactions. Cell Host Microbe 2017, 21, 270. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, Y. MAP kinase cascades in plant development and immune signaling. EMBO Rep. 2022, 23, e53817. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef]
- Zhang, S.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001, 6, 520–527. [Google Scholar] [CrossRef]
- Okushima, Y.; Koizumi, N.; Kusano, T.; Sano, H. Secreted proteins of tobacco cultured BY2 cells: Identification of a new member of pathogenesis-related proteins. Plant Mol. Biol. 2000, 42, 479–488. [Google Scholar] [CrossRef]
- Han, Z.; Xiong, D.; Schneiter, R.; Tian, C. The function of plant PR1 and other members of the CAP protein superfamily in plant-pathogen interactions. Mol. Plant Pathol. 2023, 24, 651–668. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P. Pathogenesis-Related Proteins and Peptides as Promising Tools for Engineering Plants with Multiple Stress Tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X. Salicylic Acid Inhibits Pathogen Growth in Plants through Repression of the Auxin Signaling Pathway. Curr. Biol. 2007, 17, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.X.; Tasset, C.; Hanemian, M.; Barlet, X.; Hu, J.; Tremousaygue, D.; Deslandes, L.; Marco, Y. Biological control of bacterial wilt in Arabidopsis thaliana involves abscissic acid signalling. New Phytol. 2012, 194, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O.; Leyser, O. Dynamic integration of auxin transport and signalling. Curr. Biol. 2006, 16, R424–R433. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Y.; Wei, X.; Yao, Q.; Jiang, W.; Wang, Z.; Yang, L.I.; Qian, X.U.; Yang, S.; Zhang, X. Pathotype Identification of Plasmodiophora brassicae Woron. Collected from Chinese Cabbage in Henan Province. J. Henan Agric. Sci. 2017, 46, 71–76. [Google Scholar]
- Cheng, X. Molecular Mechanism of Sly-miR319b Regulating TCPs in Response to Low Potassium Stress in Tomato. Master Thesis, Shenyang Agricultural University, Shenyang, China, 2020. [Google Scholar]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49. [Google Scholar] [CrossRef]
- Nour-Eldin, H.H.; Hansen, B.G.; Nørholm, M.H.; Jensen, J.K.; Halkier, B.A. Advancing uracil-excision based cloning towards an ideal technique for cloning PCR fragments. Nucleic Acids Res. 2006, 34, e122. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Jubault, M.; Lariagon, C.; Taconnat, L.; Renou, J.P.; Gravot, A.; Delourme, R.; Manzanares-Dauleux, M.J. Partial resistance to clubroot in Arabidopsis is based on changes in the host primary metabolism and targeted cell division and expansion capacity. Funct. Integr. Genom. 2013, 13, 191–205. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).