
Interactions between circRNAs and miR-141 in Cancer: From Pathogenesis to Diagnosis and Therapy

Abstract
:1. Introduction
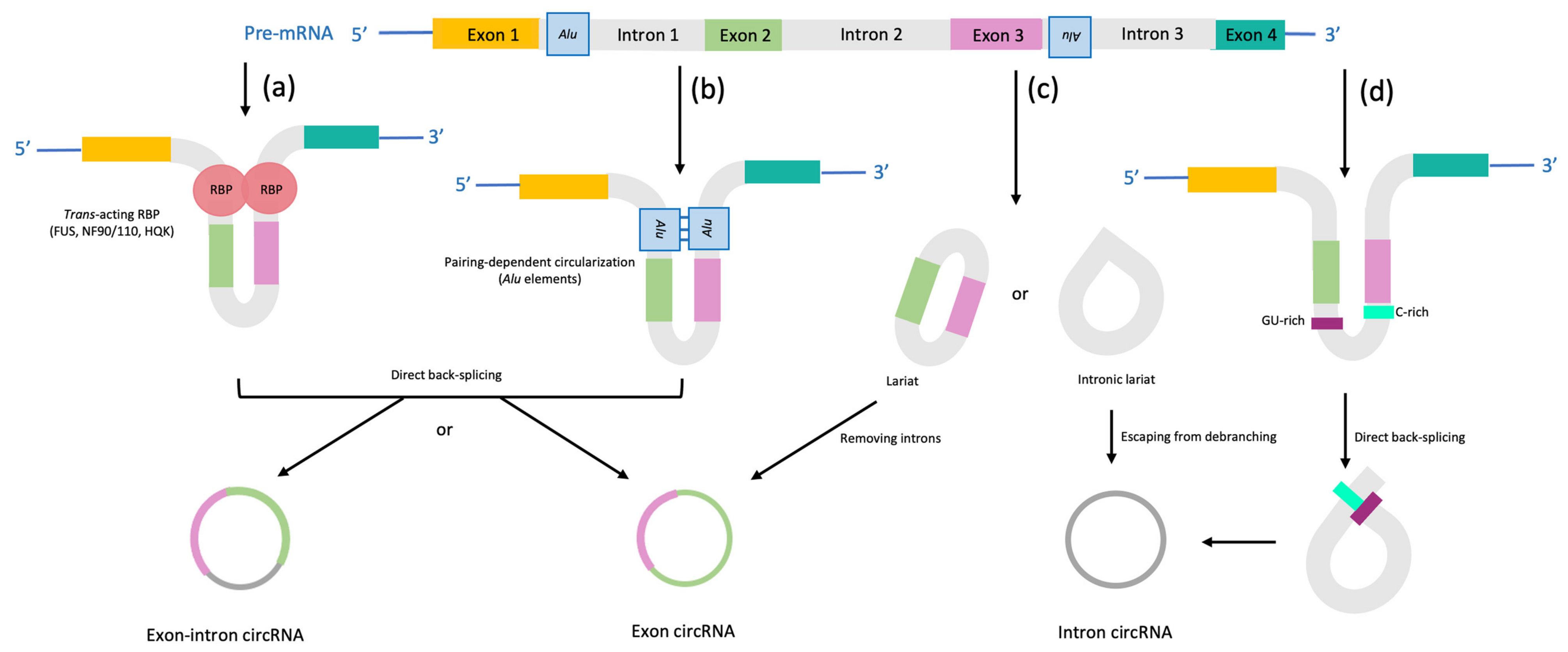
2. Biogenesis of miR-141, circRNAs and Importance of Their Interactions
3. miR-141—An Important Molecular Regulator of EMT in Cancer and EMT-Related Fibrosis in Non-Cancerous Diseases

{kind=link}
{kind=link}
| Cancer Type | ncRNA | Upregulated miR-141-3p Targets | Biological Effects of miR-141-3p and Its Targets | References |
|---|---|---|---|---|
| Gastric cancer (GC) | lncRNA H19 | ZEB1 | Inhibition of miR-141increases cell proliferation and invasion, and reduces apoptosis | [53] |
| lncRNA MAGI2-AS3 | ZEB1 | Sponging of miR-141/200a by lncRNA MAGI2-AS3 maintains ZEB1 overexpression and promotes GC progression through increased cell migration and invasion | [54] | |
| ZEB2 | Overexpression of miR-141 reverses EMT in human scirrhous GC and reduces invasion and peritoneal dissemination in nude mice orthotopic tumor model | [55] | ||
| Knock-out of ebv-miRNA (Bart9) increases expression of miR-141, CDH1, and inhibits proliferation and invasion of EBV-associated GC cells | [56] | |||
| Pancreatic cancer (PaC) | TM4SF1 | Overexpression of TM4SF1 promotes PaC cells, EMT, and angiogenesis in vitro and in vivo via Akt signaling pathway | [57] | |
| NRP1 | NRP1 is highly expressed in PaC tissues and promotes proliferation and migration of PaC cells by activation of EMT via TGF-β signaling pathway | [58] | ||
| ZEB1 | Downregulation of eIF4E increases expression of ZEB1 and vimentin, and downregulates CDH1 through repression of miR-200c and miR-141 | [59] | ||
| ZEB-1, TWIST-1 | Overexpression of miR-141 inhibits PDAC cell proliferation, migration, invasiveness, and chemoresistance in vitro. Overexpression of miR-141 and miR-720 downregulates ZEB1 and TWIST-1 leading to upregulation of CDH1 and decreased expression of MAP4K4 resulting in lower phosphorylation of JNK | [60] | ||
| Colorectal cancer (CRC) | EGFR | miR-141-3p increases cetuximab sensitivity and cetuximab-induced apoptosis, decreases EMT marker expression, suppresses proliferation, migration, and invasion in CRC cells by targeting EGFR pathway | [61] | |
| ZEB1 | Lower expression of miR-141 in oxaliplatin-resistant SW620 cells is associated with increased expression of ZEB1 and vimentin, and higher migration and invasion | [62] | ||
| ZEB1, ZEB2 | Knockdown of Ascl2 TF increases miR-141 expression, downregulatesZEB1/ZEB2 and can reverse EMT in vitro | [63] | ||
| ZEB2 | MiR-141 and CDH1 are downregulated in poorly differentiated clusters and tumor budding (TB), while ZEB2 is upregulated in TB in CRC | [64] | ||
| Non-small cell lung cancer (NSCLC) | Overexpression of miR-141 reduces KLF6 expression and increases the secretion of VEGFA in vitro and is related to higher microvessel density in cancer samples | [65] | ||
| PHLPP1, PHLPP2 | Overexpression of miR-141 promotes proliferation of NSCLC cells in vitro and cancer growth in vivo through downregulation of PI3K/AKT signaling antagonists, protein phosphatases PHLPP1 and PHLPP2 | [66] | ||
| lncRNA XIST | ZEB2 | Upregulated lncRNA XIST in metastatic NSCLC tissues decreases miR-141 expression leading to enhanced expression of ZEB2, induction of TGF-β-induced EMT, increased invasion and metastasis. | [67] | |
| ZEB1 | Overexpression of miR-141 downregulates ZEB1, increases expression of CDH1, enhances sensitivity of NSCLC cells to nintedanib and reverses the resistance of nintedanib-resistant NSCLC cells to nintedanib | [68] | ||
| lncRNA FAM83A-AS1 | Increased FAM83A-AS1 stimulates EMT, growth, migration, and invasion of LUAD cells through sponging miR-141-3p | [69] | ||
| CRC NSCLC | lncRNA LINC01296 | ZEB1, ZEB2 | High expression of LINC01296 in CRC and NSCLC cells upregulates ZEB1/ZEB2 by acting as miR-141-3p sponge to promote tumor EMT, invasion, and migration. | [70] |
| Breast cancer (BC) | Sec23A | miR-141 mimics delivered by chitosan nanoplexes to breast cancer cells diminish VEGF, metastasis, invasion, EMT and increase apoptosis | [71] | |
| ZEB1, ZEB2 | PELP1 recruits HDAC2 to repress miR-141 promoter which induces EMT, promotes migration and invasion of BC cells, and stimulates BC growth and metastasis in vivo | [72] | ||
| lnc-ATB | ZEB1, ZEB2 | High expression of lnc-ATB in BC cells and tissues decreases miR-141-3p expression to upregulate ZEB1 and ZEB2, stimulate EMT, cell migration and invasion. | [73] | |
| MiR-141 expression increases after treatment with PARP-1 inhibitor together with gemcitabine which suppresses EMT in TNBC with BRCA1 mutation in vitro | [74] | |||
| Prostate cancer (PC) | TRAF5, TRAF6 | Upregulation of miR-141-3p suppresses EMT, migration and invasion of PC cells and reduces bone metastasis in vivo via inhibition of NF-κB signaling; MiR-141-3p decreases in bone metastatic PC tissues (compared to non-bone metastatic PC); low miR-141-3p correlates with higher PSA level, Gleason grade (differentiation) and bone metastasis status | [75] | |
| BC and PC | Aberrant methylation of miR-141 CpG islands is associated with miR-141 silencing suggesting an important role for epigenetic mechanisms in the regulation of EMT | [52] | ||
| Renal cell carcinoma (RCC) | lncRNA CDKN2B-AS1 | CDKN2B-AS1, CCND1, CCND2, | Overexpression of CDKN2B-AS1 in RCC downregulates miR-141, which increases CCND1/CCND2 expression, stimulates proliferation, EMT, clonogenicity, invasion, migration, inhibits apoptosis in vitro and enhances tumor growth in vivo | [76] |
| ZEB2 | Honokiol, a biphenolic compound isolated from Magnolia spp. bark, upregulates miR-141 which suppresses expression of ZEB2, reverses EMT, inhibits formation of tumorspheres by cancer stem cells, and decreases proliferation, migration and invasion of RCC and tumor growth in vivo | [77] | ||
| Diffuse large B-cell lymphoma (DLBCL) | lncRNA LINC01857 | MAP4K4 | Upregulation of LINC01857 in DLBCL tissues and cells promotes proliferation, cell cycle, but suppresses apoptosis in DLBCL cells by sponging miR-141-3p, which decreases MAP4K4 and activates EMT and PI3K/mTOR pathway. | [78] |
| Head and neck squamous cell carcinoma (HNSCC) | ZEB1, ZEB2 | Enforced expression of miR-141 downregulates ZEB1/ZEB2, increases CDH1 expression, and reduces migration of HNSCC cells | [79] | |
| Ovarian cancer (OC) | ZEB | miR-141 mimic downregulates ZEB, upregulates CDH1, inhibits EMT and cell proliferation, decreases migration and invasion in vitro | [80] | |
| ZEB1, ZEB2, SNAl2 | Stable inhibition of miR-141 upregulates expression of ZEB1, ZEB2, SNAl2, vimentin and fibronectin, but downregulates CDH1 and decreases sensitivity of OC cells to paclitaxel and carboplatin | [81] | ||
| Laryngeal cancer (LC) | HOXC6 | Overexpression of miR-141 downregulates HOXC6, inhibits TGF-β signaling, represses EMT, migration, viability, and invasion of LC cells and decreases tumor growth and metastasis to lymph nodes in vivo | [82] | |
| Nasopharyngeal carcinoma (NPC) | DLC1 | Overexpression of miR-141-3p promotes cell proliferation, migration, invasion, and EMT in NPC cells by targeting DLC1 and activation of mTOR signaling pathway | [83] | |
| Hepatocellular carcinoma (HCC) | GP73 | Overexpression of miR-141-3p inhibits EMT, proliferation, invasion and migration of HCC cells in vitro, and tumor growth and lung metastasis in vivo | [84] | |
| ZEB1 | Downregulation of miR-141 in HCC with bile duct thrombus is associated with overexpression of ZEB1, TWIST, TGFβRII, vimentin, IL-6, Bmi1 and reduced level of IGFBP-4 | [85] | ||
| ZEB1 | Increased expression of ELF3 TF downregulates miR-141-3p to stimulate ZEB1 expression, EMT, cell proliferation, migration and invasion in vitro and metastasis in vivo | [86] | ||
| Endometrial cancer (EC) | ZEB1 | Lower expression of miR-141 in epoxomicin-resistant EC cells is associated with the upregulation of ZEB1, downregulation of CDH1 and induction of EMT. | [87] | |
| Osteosarcoma | AUF1 | miR-141 inhibits proliferation, invasion, and migration of osteosarcoma cells, and suppresses EMT through repression of RNA-binding protein AUF1 | [88] |
4. circRNAs as Sponges for miR-141 in Different Types of Human Cancers
4.1. Breast Cancer
4.2. Lung Cancer
4.3. Gastric Cancer
4.4. Prostate Cancer
4.5. Liver Cancer
4.6. Thyroid Cancer
4.7. Bladder Cancer
4.8. Oral Squamous Cell Carcinoma
4.9. Bone Cancer
5. Summary of the Importance of Interactions between ncRNAs and miR-141 in Cancer
5.1. miR-141 Acting as Tumor Suppressor
5.2. miR-141 Acting as an Oncogene
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.; Cheng, Z.; Pang, Y.; Cui, L.; Qian, T.; Quan, L.; Zhao, H.; Shi, J.; Ke, X.; Fu, L. Role of microRNAs, circRNAs and long noncoding RNAs in acute myeloid leukemia. J. Hematol. Oncol. 2019, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Han, T.-S.; Hur, K.; Cho, H.-S.; Ban, H.S. Epigenetic Associations between lncRNA/circRNA and miRNA in Hepatocellular Carcinoma. Cancers 2020, 12, 2622. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.L.; Hsieh, A.C. The Untranslated Regions of mRNAs in Cancer. Trends Cancer 2019, 5, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. miRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2020, 4, 1. [Google Scholar] [CrossRef]
- Xu, P.; Wu, Q.; Yu, J.; Rao, Y.; Kou, Z.; Fang, G.; Shi, X.; Liu, W.; Han, H. A Systematic Way to Infer the Regulation Relations of miRNAs on Target Genes and Critical miRNAs in Cancers. Front. Genet. 2020, 11, 278. [Google Scholar] [CrossRef]
- Zhou, K.; Liu, M.; Cao, Y. New Insight into microRNA Functions in Cancer: Oncogene–microRNA–Tumor Suppressor Gene Network. Front. Mol. Biosci. 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Claret, F.X.; Wu, W. MicroRNAs as therapeutic targets in nasopharyngeal carcinoma. Front. Oncol. 2019, 9, 756. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Chen, J. microRNAs as therapeutic targets in intestinal diseases. ExRNA 2019, 1, 23. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.; Krishna, M.; Singhal, J.; Horne, D.; Awasthi, S.; Salgia, R.; Singhal, S.S. Therapeutic targeting of miRNA-216b in cancer. Cancer Lett. 2020, 484, 16–28. [Google Scholar] [CrossRef]
- Miroshnichenko, S.; Patutina, O. Enhanced Inhibition of Tumorigenesis Using Combinations of miRNA-Targeted Therapeutics. Front. Pharmacol. 2019, 10, 488. [Google Scholar] [CrossRef]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef]
- Zhou, W.-Y.; Cai, Z.-R.; Liu, J.; Wang, D.-S.; Ju, H.-Q.; Xu, R.-H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Zhang, P.; Li, S.; Chen, M. Characterization and Function of Circular RNAs in Plants. Front. Mol. Biosci. 2020, 7, 91. [Google Scholar] [CrossRef]
- Liu, K.-S.; Pan, F.; Mao, X.-D.; Liu, C.; Chen, Y.-J. Biological functions of circular RNAs and their roles in occurrence of reproduction and gynecological diseases. Am. J. Transl. Res. 2019, 11, 1. [Google Scholar]
- Liu, J.; Zhang, X.; Yan, M.; Li, H. Emerging Role of Circular RNAs in Cancer. Front. Oncol. 2020, 10, 663. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Khanabdali, R.; Kalionis, B.; Tai, X.; Xia, S. Circular RNAs: Isolation, characterization and their potential role in diseases. RNA Biol. 2017, 14, 1715–1721. [Google Scholar] [CrossRef] [Green Version]
- Okholm, T.L.H.; Sathe, S.; Park, S.S.; Kamstrup, A.B.; Rasmussen, A.M.; Shankar, A.; Chua, Z.M.; Fristrup, N.; Nielsen, M.M.; Vang, S.; et al. Transcriptome-wide profiles of circular RNA and RNA-binding protein interactions reveal effects on circular RNA biogenesis and cancer pathway expression. Genome Med. 2020, 12, 112. [Google Scholar] [CrossRef]
- Wan, B.; Hu, H.; Wang, R.; Liu, W.; Chen, D. Therapeutic Potential of Circular RNAs in Osteosarcoma. Front. Oncol. 2020, 10, 370. [Google Scholar] [CrossRef]
- Senfter, D.; Madlener, S.; Krupitza, G.; Mader, R.M. The microRNA-200 family: Still much to discover. Biomol. Concepts 2016, 7, 311–319. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, B.; Han, S.; Zhang, K.; Chen, J.; Li, C.; Wang, R.; Chen, L. The roles of MicroRNA-141 in human cancers: From diagnosis to treatment. Cell Physiol. Biochem. 2016, 38, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Chen, W.; Wu, H.; Liu, C.; Zhang, J.; Chen, S. Mechanisms and Functions of MiR-200 Family in Hepatocellular Carcinoma. Onco Targets Ther. 2021, 13, 13479–13490. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, J.; Chen, Q.; Cheng, J.; Chen, X.; Mao, Y.; Chen, W.; Liu, C.; Wu, H.; Lv, Y.; et al. MicroRNA-200a and microRNA-141 have a synergetic effect on the suppression of epithelial-mesenchymal transition in liver cancer by targeting STAT4. Oncol. Lett. 2020, 21, 137. [Google Scholar] [CrossRef] [PubMed]
- Pitolli, C.; Marini, A.; Sette, C.; Pagliarini, V. Non-Canonical Splicing and Its Implications in Brain Physiology and Cancer. Int. J. Mol. Sci. 2022, 23, 2811. [Google Scholar] [CrossRef] [PubMed]
- Blakes, A.J.M.; Wai, H.A.; Davies, I.; Moledina, H.E.; Ruiz, A.; Thomas, T.; Bunyan, D.; Thomas, N.S.; Burren, C.P.; Greenhalgh, L.; et al. A systematic analysis of splicing variants identifies new diagnoses in the 100,000 Genomes Project. Genome Med. 2022, 14, 79. [Google Scholar] [CrossRef]
- Xiao, M.S.; Ai, Y.; Wilusz, J.E. Biogenesis and Functions of Circular RNAs Come into Focus. Trends Cell Biol. 2020, 30, 226–240. [Google Scholar] [CrossRef]
- Greene, J.; Baird, A.M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Geng, X.; Jia, Y.; Zhang, Y.; Shi, L.; Li, Q.; Zang, A.; Wang, H. Circular RNA: Biogenesis, degradation, functions and potential roles in mediating resistance to anticarcinogens. Epigenomics 2020, 12, 267–283. [Google Scholar] [CrossRef]
- Das, A.; Sinha, T.; Shyamal, S.; Panda, A.C. Emerging role of circular rna–protein interactions. Non Coding RNA 2021, 7, 48. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, Q. Mechanisms regulating abnormal circular RNA biogenesis in cancer. Cancers 2021, 13, 4185. [Google Scholar] [CrossRef]
- Wu, J.; Qi, X.; Liu, L.; Hu, X.; Liu, J.; Yang, J.; Yang, J.; Lu, L.; Zhang, Z.; Ma, S.; et al. Emerging Epigenetic Regulation of Circular RNAs in Human Cancer. Mol. Ther. Nucleic Acids 2019, 16, 589–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the epithelial–mesenchymal transition (EMT) with cisplatin resistance. Int. J. Mol. Sci. 2020, 21, 4002. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Li, Q.; Wu, F.; Lin, J.; Chen, J.; Zheng, H.; Guo, L. Epithelial-Mesenchymal Transition and Metabolic Switching in Cancer: Lessons From Somatic Cell Reprogramming. Front. Cell Dev. Biol. 2020, 8, 760. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef]
- Jo, H.; Shim, K.; Jeoung, D. Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics. Int. J. Mol. Sci. 2022, 23, 5881. [Google Scholar] [CrossRef]
- Cavallari, I.; Ciccarese, F.; Sharova, E.; Urso, L.; Raimondi, V.; Silic-Benussi, M.; D’Agostino, D.M.; Ciminale, V. The miR-200 Family of microRNAs: Fine Tuners of Epithelial-Mesenchymal Transition and Circulating Cancer Biomarkers. Cancers 2021, 13, 5874. [Google Scholar] [CrossRef]
- Huang, Y.; Tong, J.; He, F.; Yu, X.; Fan, L.; Hu, J.; Tan, J.; Chen, Z. miR-141 regulates TGF-β1-induced epithelial-mesenchymal transition through repression of HIPK2 expression in renal tubular epithelial cells. Int. J. Mol. Med. 2015, 35, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, Q.; Davis, F.; Mao, J.; Zhao, H.; Ma, D. Epithelial–mesenchymal transition in organ fibrosis development: Current understanding and treatment strategies. Burn. Trauma 2022, 10, tkac011. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, C.; Hou, L.; Wu, Y. Silencing of the lncRNA TUG1 attenuates the epithelial-mesenchymal transition of renal tubular epithelial cells by sponging miR-141-3p via regulating β-catenin. Am. J. Physiol. Renal Physiol. 2020, 319, F1125–F1134. [Google Scholar] [CrossRef]
- Yarani, R.; Shojaeian, A.; Palasca, O.; Doncheva, N.T.; Jensen, L.J.; Gorodkin, J.; Pociot, F. Differentially Expressed miRNAs in Ulcerative Colitis and Crohn’s Disease. Front. Immunol. 2022, 13, 865777. [Google Scholar] [CrossRef]
- García-Cuesta, E.M.; Santiago, C.A.; Vallejo-Díaz, J.; Juarranz, Y.; Rodríguez-Frade, J.M.; Mellado, M. The Role of the CXCL12/CXCR4/ACKR3 Axis in Autoimmune Diseases. Front. Endocrinol. 2019, 10, 585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Yoneyama, H. New endoscopic approach of anti-fibrotic therapy for inflammatory bowel disease. Ann. Transl. Med. 2017, 5, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, W.; Cai, X.; Qian, Q.; Peng, W.; Yu, J.; Zhang, X.; Tian, L.; Wang, C. lncRNA ZEB1-AS1 promotes pulmonary fibrosis through ZEB1-mediated epithelial–mesenchymal transition by competitively binding miR-141-3p. Cell Death Dis. 2019, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, A.; Gaudet, M.; Plesa, M.; Allakhverdi, Z.; Mogas, A.K.; Audusseau, S.; Baglole, C.J.; Eidelman, D.H.; Olivenstein, R.; Ludwig, M.S.; et al. Neutrophils from severe asthmatic patients induce epithelial to mesenchymal transition in healthy bronchial epithelial cells. Respir. Res. 2019, 20, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, S.; Comstock, A.T.; Sajjan, U.S. Barrier function of airway tract epithelium. Tissue Barriers 2013, 1, e24997. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.; Johansson, K.; Joo, A.; Bonser, L.R.; Koh, K.D.; Le Tonqueze, O.; Bolourchi, S.; Bautista, R.A.; Zlock, L.; Roth, T.L.; et al. Epithelial miR-141 regulates IL-13–induced airway mucus production. JCI Insight 2021, 6, A7468. [Google Scholar] [CrossRef]
- Viganò, P.; Ottolina, J.; Bartiromo, L.; Bonavina, G.; Schimberni, M.; Villanacci, R.; Candiani, M. Cellular Components Contributing to Fibrosis in Endometriosis: A Literature Review. J. Minim. Invasive Gynecol. 2020, 27, 287–295. [Google Scholar] [CrossRef]
- Yang, Y.-M.; Yang, W.-X. Epithelial-to-mesenchymal transition in the development of endometriosis. Oncotarget 2017, 8, 41679–41689. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Luo, Y.; Wang, G.; Yang, Q. CircATRNL1 promotes epithelial–mesenchymal transition in endometriosis by upregulating Yes-associated protein 1 in vitro. Cell Death Dis. 2020, 11, 594. [Google Scholar] [CrossRef]
- Liang, Z.-H.; Pan, Y.-C.; Lin, S.-S.; Qiu, Z.-Y.; Zhang, Z. LncRNA MALAT1 promotes wound healing via regulating miR-141-3p/ZNF217 axis. Regen. Ther. 2020, 15, 202–209. [Google Scholar] [CrossRef]
- Rajić, J.; Dinić, S.; Uskoković, A.; Arambašić Jovanović, J.; Tolić, A.; Đorđević, M.; Đorđević, M.; Poznanović, G.; Mihailović, M.; Inic-Kanada, A.; et al. DNA methylation of miR-200 clusters promotes epithelial to mesenchymal transition in human conjunctival epithelial cells. Exp. Eye Res. 2020, 197, 108047. [Google Scholar] [CrossRef]
- Vrba, L.; Jensen, T.J.; Garbe, J.C.; Heimark, R.L.; Cress, A.E.; Dickinson, S.; Stampfer, M.R.; Futscher, B.W. Role for DNA Methylation in the Regulation of miR-200c and miR-141 Expression in Normal and Cancer Cells. PLoS ONE 2010, 5, e8697. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ye, F.; Yin, C.; Zhuang, Y.; Yue, G.; Zhang, G. The Interaction Between MiR-141 and lncRNA-H19 in Regulating Cell Proliferation and Migration in Gastric Cancer. Cell Physiol. Biochem. 2015, 36, 1440–1452. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, J.; Zhang, M.; Hu, X.; She, J.; Qiu, X.; Zhang, X.; Xu, L.; Liu, Y.; Qin, S. LncRNA MAGI2-AS3 Is Regulated by BRD4 and Promotes Gastric Cancer Progression via Maintaining ZEB1 Overexpression by Sponging miR-141/200a. Mol. Ther. Nucleic Acids 2020, 19, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Takei, Y.; Shen, G.; Morita-Kondo, A.; Hara, T.; Mihara, K.; Yanagihara, K. MicroRNAs Associated with Epithelial-Mesenchymal Transition Can Be Targeted to Inhibit Peritoneal Dissemination of Human Scirrhous Gastric Cancers. Pathobiology 2018, 85, 232–246. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Liu, Y.Y.; Liu, K.H.; Hsu, J.T.; Chen, T.C.; Chiu, C.T.; Yeh, T. Sen Comprehensive profiling of virus microRNAs of Epstein–Barr virus-associated gastric carcinoma: Highlighting the interactions of ebv-Bart9 and host tumor cells. J. Gastroenterol. Hepatol. 2017, 32, 82–91. [Google Scholar] [CrossRef]
- Xu, D.; Yang, F.; Wu, K.; Xu, X.; Zeng, K.; An, Y.; Xu, F.; Xun, J.; Lv, X.; Zhang, X.; et al. Lost miR-141 and upregulated TM4SF1 expressions associate with poor prognosis of pancreatic cancer: Regulation of EMT and angiogenesis by miR-141 and TM4SF1 via AKT. Cancer Biol. Ther. 2020, 21, 354–363. [Google Scholar] [CrossRef]
- Ma, L.; Zhai, B.; Zhu, H.; Li, W.; Jiang, W.; Lei, L.; Zhang, S.; Qiao, H.; Jiang, X.; Sun, X. The miR-141/neuropilin-1 axis is associated with the clinicopathology and contributes to the growth and metastasis of pancreatic cancer. Cancer Cell Int. 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.; Chow, C.R.; Ebine, K.; Arslan, A.D.; Kwok, B.; Bentrem, D.J.; Eckerdt, F.D.; Platanias, L.C.; Munshi, H.G. Differential regulation of ZEB1 and EMT by MAPK-interacting protein kinases (MNK) and eIF4E in pancreatic cancer. Mol. Cancer Res. 2016, 14, 216–227. [Google Scholar] [CrossRef] [Green Version]
- Lemberger, M.; Loewenstein, S.; Lubezky, N.; Nizri, E.; Pasmanik-Chor, M.; Barazovsky, E.; Klausner, J.M.; Lahat, G. MicroRNA profiling of pancreatic ductal adenocarcinoma (PDAC) reveals signature expression related to lymph node metastasis. Oncotarget 2019, 10, 2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Jing, H.; Zhang, Y.; Suo, J.; Qian, M. MicroRNA-141-3p affected proliferation, chemosensitivity, migration and invasion of colorectal cancer cells by targeting EGFR. Int. J. Biochem. Cell Biol. 2020, 118, 105643. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Hosokawa, M.; Yonezawa, T.; Hayashi, W.; Ueda, K.; Iwakawa, S. Induction of epithelial-mesenchymal transition and down-regulation of MIR-200c and MIR-141 in oxaliplatin-resistant colorectal cancer cells. Biol. Pharm. Bull. 2015, 38, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Pan, Q.; Shang, Y.; Zhu, R.; Ye, J.; Liu, Y.; Zhong, X.; Li, S.; He, Y.; Chen, L.; et al. MicroRNA-200 (miR-200) cluster regulation by achaete scute-like 2 (Ascl2) impact on the epithelial-mesenchymal transition in colon cancer cells. J. Biol. Chem. 2014, 289, 36101–36115. [Google Scholar] [CrossRef] [Green Version]
- Pavlič, A.; Boštjančič, E.; Kavalar, R.; Ilijevec, B.; Bonin, S.; Zanconati, F.; Zidar, N. Tumour budding and poorly differentiated clusters in colon cancer—Different manifestations of partial epithelial–mesenchymal transition. J. Pathol. 2022, 258, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Tejero, R.; Navarro, A.; Campayo, M.; Viñolas, N.; Marrades, R.M.; Cordeiro, A.; Ruíz-Martínez, M.; Santasusagna, S.; Molins, L.; Ramirez, J.; et al. MiR-141 and miR-200c as markers of overall survival in early stage non-small cell lung cancer adenocarcinoma. PLoS ONE 2014, 9, e101899. [Google Scholar] [CrossRef]
- Mei, Z.; He, Y.; Feng, J.; Shi, J.; Du, Y.; Qian, L.; Huang, Q.; Jie, Z. MicroRNA-141 promotes the proliferation of non-small cell lung cancer cells by regulating expression of PHLPP1 and PHLPP2. FEBS Lett. 2014, 588, 3055–3061. [Google Scholar] [CrossRef]
- Li, C.; Wan, L.; Liu, Z.; Xu, G.; Wang, S.; Su, Z.; Zhang, Y.; Zhang, C.; Liu, X.; Lei, Z.; et al. Long non-coding RNA XIST promotes TGF-β-induced epithelial-mesenchymal transition by regulating miR-367/141-ZEB2 axis in non-small-cell lung cancer. Cancer Lett. 2018, 418, 185–195. [Google Scholar] [CrossRef]
- Nishijima, N.; Seike, M.; Soeno, C.; Chiba, M.; Miyanaga, A.; Noro, R.; Sugano, T.; Matsumoto, M.; Kubota, K.; Gemma, A. miR-200/ZEB axis regulates sensitivity to nintedanib in non-small cell lung cancer cells. Int. J. Oncol. 2016, 48, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Yang, C.; Zhang, Q.; Zhuo, T.; Li, X.; Li, N.; Zhu, L.; Luo, C.; Gan, J.; Wu, Y. Long non-coding RNA FAM83A antisense RNA 1 (lncRNA FAM83A-AS1) targets microRNA-141-3p to regulate lung adenocarcinoma cell proliferation, migration, invasion, and epithelial-mesenchymal transition progression. Bioengineered 2022, 13, 4964–4977. [Google Scholar] [CrossRef]
- Sun, Z.; Shao, B.; Liu, Z.; Dang, Q.; Guo, Y.; Chen, C.; Guo, Y.; Chen, Z.; Liu, J.; Hu, S.; et al. LINC01296/miR-141-3p/ZEB1-ZEB2 axis promotes tumor metastasis via enhancing epithelial-mesenchymal transition process. J. Cancer 2021, 12, 2723–2734. [Google Scholar] [CrossRef]
- Kaban, K.; Salva, E.; Akbuga, J. Modulation of the dual-faced effects of miR-141 with chitosan/miR-141 nanoplexes in breast cancer cells. J. Gene Med. 2019, 21, e3116. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.S.; Gonugunta, V.K.; Bandyopadhyay, A.; Rao, M.K.; Goodall, G.J.; Sun, L.Z.; Tekmal, R.R.; Vadlamudi, R.K. Significance of PELP1/HDAC2/miR-200 regulatory network in EMT and metastasis of breast cancer. Oncogene 2014, 33, 3707–3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, J.; Jia, S.; Wang, Y.; Kang, Y.; Zhang, W. Down-regulation of lncRNA-ATB inhibits epithelial-mesenchymal transition of breast cancer cells by increasing miR-141-3p expression. Biochem. Cell Biol. 2019, 97, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, Y.; Sasaki, A.; Sakamoto, N.; Haruyama, Y.; Nashimoto, M.; Koshida, Y.; Fukuma, E. Effects of olaparib, a PARP-1 inhibitor, on triple negative breast cancer cells with a BRCA1 mutation. Jpn. J. Cancer Chemother. 2021, 48, 53–56. [Google Scholar]
- Huang, S.; Wa, Q.; Pan, J.; Peng, X.; Ren, D.; Huang, Y.; Chen, X.; Tang, Y. Downregulation of miR-141-3p promotes bone metastasis via activating NF-κB signaling in prostate cancer. J. Exp. Clin. Cancer Res. 2017, 36, 173. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, P.; Kulkarni, P.; Majid, S.; Hashimoto, Y.; Shiina, M.; Shahryari, V.; Bhat, N.S.; Tabatabai, L.; Yamamura, S.; Saini, S.; et al. LncRNA CDKN2B-AS1/miR-141/cyclin D network regulates tumor progression and metastasis of renal cell carcinoma. Cell Death Dis. 2020, 11, 660. [Google Scholar] [CrossRef]
- Li, W.; Wang, Q.; Su, Q.; Ma, D.; An, C.; Ma, L.; Liang, H. Honokiol suppresses renal cancer cells’ metastasis Via dual-blocking epithelial-mesenchymal transition and cancer stem cell properties through modulating miR-141/ZEB2 signaling. Mol. Cells 2014, 37, 383. [Google Scholar] [CrossRef]
- Li, Q.; Li, B.; Lu, C.L.; Wang, J.Y.; Gao, M.; Gao, W. LncRNA LINC01857 promotes cell growth and diminishes apoptosis via PI3K/mTOR pathway and EMT process by regulating miR-141-3p/MAP4K4 axis in diffuse large B-cell lymphoma. Cancer Gene Ther. 2021, 28, 1046–1057. [Google Scholar] [CrossRef]
- Tamagawa, S.; Beder, L.B.; Hotomi, M.; Gunduz, M.; Yata, K.; Grenman, R.; Yamanaka, N. Role of miR-200c/miR-141 in the regulation of epithelial-mesenchymal transition and migration in head and neck squamous cell carcinoma. Int. J. Mol. Med. 2014, 33, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Lei, L.; Shao, L.; Shi, J.; Jia, J.; Tong, X. MicroRNA-141 inhibits epithelial-mesenchymal transition, and ovarian cancer cell migration and invasion. Mol. Med. Rep. 2017, 16, 6743–6749. [Google Scholar] [CrossRef] [Green Version]
- Brozovic, A.; Duran, G.E.; Wang, Y.C.; Francisco, E.B.; Sikic, B.I. The miR-200 family differentially regulates sensitivity to paclitaxel and carboplatin in human ovarian carcinoma OVCAR-3 and MES-OV cells. Mol. Oncol. 2015, 9, 1678–1693. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sun, D.-Z.; Fu, Y.-G.; Yang, P.-Z.; Lv, H.-Q.; Gao, Y.; Zhang, X.-Y. Upregulation of microRNA-141 suppresses epithelial-mesenchymal transition and lymph node metastasis in laryngeal cancer through HOXC6-dependent TGF-β signaling pathway. Cell Signal. 2020, 66, 109444. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.W.; Zhou, X.Y.; Wang, Q.J.; Han, L.H.; Jiao, J.B. MicroRNA-141-3p promoted the progression of nasopharyngeal carcinoma through targeting DLC1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11105–11113. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Yang, L.; Jiang, X.; Liu, Z.; Li, X.; Xie, S.; Li, G.; Liu, J. Role of microRNA-141-3p in the progression and metastasis of hepatocellular carcinoma cell. Int. J. Biol. Macromol. 2019, 128, 331–339. [Google Scholar] [CrossRef]
- Yeh, T.S.; Wang, F.; Chen, T.C.; Yeh, C.N.; Yu, M.C.; Jan, Y.Y.; Chen, M.F. Expression profile of MicroRNA-200 family in hepatocellular carcinoma with bile duct tumor thrombus. Ann. Surg. 2014, 259, 346–354. [Google Scholar] [CrossRef]
- Zheng, L.; Xu, M.; Xu, J.; Wu, K.; Fang, Q.; Liang, Y.; Zhou, S.; Cen, D.; Ji, L.; Han, W.; et al. ELF3 promotes epithelial–mesenchymal transition by protecting ZEB1 from miR-141-3p-mediated silencing in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 387. [Google Scholar] [CrossRef]
- Asakura, T.; Yamaguchi, N.; Ohkawa, K.; Yoshida, K. Proteasome inhibitor-resistant cells cause EMT-induction via suppression of E-cadherin by miR-200 and ZEB1. Int. J. Oncol. 2015, 46, 2251–2260. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalaf, H.H.; Aboussekhra, A. MicroRNA-141 and microRNA-146b-5p inhibit the prometastatic mesenchymal characteristics through the RNA-binding protein AUF1 targeting the transcription factor ZEB1 and the protein kinase AKT. J. Biol. Chem. 2014, 289, 31433–31447. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Sakshi, S.; Jayasuriya, R.; Ganesan, K.; Xu, B.; Ramkumar, K.M. Role of circRNA-miRNA-mRNA interaction network in diabetes and its associated complications. Mol. Ther. Nucleic Acids 2021, 26, 1291–1302. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Kuo, H.-C. The emerging roles and functions of circular RNAs and their generation. J. Biomed. Sci. 2019, 26, 29. [Google Scholar] [CrossRef]
- Wu, Q.; Li, P.; Wu, M.; Liu, Q. Deregulation of Circular RNAs in Cancer From the Perspectives of Aberrant Biogenesis, Transport and Removal. Front. Genet. 2019, 10, 16. [Google Scholar] [CrossRef]
- Huang, J.; Yu, S.; Ding, L.; Ma, L.; Chen, H.; Zhou, H.; Zou, Y.; Yu, M.; Lin, J.; Cui, Q. The Dual Role of Circular RNAs as miRNA Sponges in Breast Cancer and Colon Cancer. Biomedicines 2021, 9, 1590. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zou, Y.; Li, X.; Yang, A.; Ye, F.; Zhang, J.; Wei, W.; Kong, Y. circGNB1 Facilitates Triple-Negative Breast Cancer Progression by Regulating miR-141-5p-IGF1R Axis. Front. Genet. 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhao, X.; Wang, H. circ_0075943 Dominates the miR-141-3p/AK2 Network to Support the Development of Breast Carcinoma. J. Oncol. 2021, 2021, 4098270. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, F.; Sun, D.; Liu, J.; Liu, B.K.; He, Y.L.; Pang, S.; Shi, B.W.; Zhou, F.C.; Yao, L.; et al. CircKEAP1 Suppresses the Progression of Lung Adenocarcinoma via the miR-141-3p/KEAP1/NRF2 Axis. Front. Oncol. 2021, 11, 672586. [Google Scholar] [CrossRef]
- Chen, D.-L.; Sheng, H.; Zhang, D.-S.; Jin, Y.; Zhao, B.-T.; Chen, N.; Song, K.; Xu, R.-H. The circular RNA circDLG1 promotes gastric cancer progression and anti-PD-1 resistance through the regulation of CXCL12 by sponging miR-141-3p. Mol. Cancer 2021, 20, 166. [Google Scholar] [CrossRef]
- Nan, C.; Wang, Y.; Yang, S.; Chen, Y. circCRKL suppresses the progression of prostate cancer cells by regulating the miR-141/KLF5 axis. Pathol. Res. Pract. 2020, 216, 153182. [Google Scholar] [CrossRef]
- Chao, F.; Song, Z.; Wang, S.; Ma, Z.; Zhuo, Z.; Meng, T.; Xu, G.; Chen, G. Novel circular RNA circSOBP governs amoeboid migration through the regulation of the miR-141-3p/MYPT1/p-MLC2 axis in prostate cancer. Clin. Transl. Med. 2021, 11, e360. [Google Scholar] [CrossRef]
- Chen, D.; Chou, F.J.; Chen, Y.; Tian, H.; Wang, Y.; You, B.; Niu, Y.; Huang, C.P.; Yeh, S.; Xing, N.; et al. Targeting the radiation-induced TR4 nuclear receptor-mediated QKI/circZEB1/miR-141-3p/ZEB1 signaling increases prostate cancer radiosensitivity. Cancer Lett. 2020, 495, 100–111. [Google Scholar] [CrossRef]
- Huang, X.-Y.; Huang, Z.-L.; Zhang, P.-B.; Huang, X.-Y.; Huang, J.; Wang, H.-C.; Xu, B.; Zhou, J.; Tang, Z.-Y. CircRNA-100338 Is Associated With mTOR Signaling Pathway and Poor Prognosis in Hepatocellular Carcinoma. Front. Oncol. 2019, 9, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.Y.; Huang, Z.L.; Xu, Y.H.; Zheng, Q.; Chen, Z.; Song, W.; Zhou, J.; Tang, Z.Y.; Huang, X.Y. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-100338/MIR-141-3p pathway in hepatitis B-related hepatocellular carcinoma. Sci. Rep. 2017, 7, 5428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Tian, P.; Zheng, W.; Yan, X. Piplartine attenuates the proliferation of hepatocellular carcinoma cells via regulating hsa_circ_100338 expression. Cancer Med. 2020, 9, 4265–4273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Liu, G.; Sun, Y.; Gao, Y.; Ouyang, X.; Chang, C.; Gong, L.; Yeh, S. Targeting the estrogen receptor alpha (ERα)-mediated circ-SMG1.72/miR-141-3p/Gelsolin signaling to better suppress the HCC cell invasion. Oncogene 2020, 39, 2493–2508. [Google Scholar] [CrossRef]
- Peng, N.; Shi, L.; Zhang, Q.; Hu, Y.; Wang, N.; Ye, H. Microarray profiling of circular RNAs in human papillary thyroid carcinoma. PLoS ONE 2017, 12, e0170287. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wu, S.; Mou, Z.; Zhou, Q.; Dai, X.; Ou, Y.; Chen, X.; Chen, Y.; Xu, C.; Hu, Y.; et al. Exosome-derived circTRPS1 promotes malignant phenotype and CD8+ T cell exhaustion in bladder cancer microenvironments. Mol. Ther. 2022, 30, 1054–1070. [Google Scholar] [CrossRef]
- Dou, Z.; Li, S.; Ren, W.; Wang, Q.; Liu, J.; Kong, X.; Gao, L.; Zhi, K. Decreased expression of hsa:circ_0072387 as a valuable predictor for oral squamous cell carcinoma. Oral Dis. 2019, 25, 1302–1308. [Google Scholar] [CrossRef]
- Yu, Y.; Dong, G.; Li, Z.; Zheng, Y.; Shi, Z.; Wang, G. circ-LRP6 contributes to osteosarcoma progression by regulating the miR-141-3p/HDAC4/HMGB1 axis. Int. J. Oncol. 2022, 60, 38. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Lüönd, F.; Tiede, S.; Christofori, G. Breast cancer as an example of tumour heterogeneity and tumour cell plasticity during malignant progression. Br. J. Cancer 2021, 125, 164–175. [Google Scholar] [CrossRef]
- Turashvili, G.; Brogi, E. Tumor Heterogeneity in Breast Cancer. Front. Med. 2017, 4, 227. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, F.; Franken, A.; Fehm, T.; Neubauer, H. Navigation through inter- and intratumoral heterogeneity of endocrine resistance mechanisms in breast cancer: A potential role for Liquid Biopsies? Tumor Biol. 2017, 39, 101042831773151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dass, S.A.; Tan, K.L.; Selva Rajan, R.; Mokhtar, N.F.; Mohd Adzmi, E.R.; Wan Abdul Rahman, W.F.; Tengku Din, T.A.D.A.-A.; Balakrishnan, V. Triple Negative Breast Cancer: A Review of Present and Future Diagnostic Modalities. Medicina 2021, 57, 62. [Google Scholar] [CrossRef]
- Hsu, J.-Y.; Chang, C.-J.; Cheng, J.-S. Survival, treatment regimens and medical costs of women newly diagnosed with metastatic triple-negative breast cancer. Sci. Rep. 2022, 12, 729. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Zu, X.; Chen, Z.; Wen, G.; Zhong, J. Noncoding RNAs in triple negative breast cancer: Mechanisms for chemoresistance. Cancer Lett. 2021, 523, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Kong, Q.; Yin, J.; Zhang, J.; Jiang, Y. Insulin-like growth factor receptor signaling in tumorigenesis and drug resistance: A challenge for cancer therapy. J. Hematol. Oncol. 2020, 13, 64. [Google Scholar] [CrossRef]
- Klepinin, A.; Zhang, S.; Klepinina, L.; Rebane-Klemm, E.; Terzic, A.; Kaambre, T.; Dzeja, P. Adenylate Kinase and Metabolic Signaling in Cancer Cells. Front. Oncol. 2020, 10, 660. [Google Scholar] [CrossRef]
- Nooreldeen, R.; Bach, H. Current and Future Development in Lung Cancer Diagnosis. Int. J. Mol. Sci. 2021, 22, 8661. [Google Scholar] [CrossRef]
- Santos, R.M.; Moreno, C.; Zhang, W.C. Non-Coding RNAs in Lung Tumor Initiation and Progression. Int. J. Mol. Sci. 2020, 21, 2774. [Google Scholar] [CrossRef] [Green Version]
- Pouremamali, F.; Pouremamali, A.; Dadashpour, M.; Soozangar, N.; Jeddi, F. An update of Nrf2 activators and inhibitors in cancer prevention/promotion. Cell Commun. Signal. 2022, 20, 100. [Google Scholar] [CrossRef]
- Deshmukh, P.; Unni, S.; Krishnappa, G.; Padmanabhan, B. The Keap1–Nrf2 pathway: Promising therapeutic target to counteract ROS-mediated damage in cancers and neurodegenerative diseases. Biophys. Rev. 2017, 9, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Niu, M.; Xu, L.; Luo, S.; Wu, K. Regulation of PD-L1 expression in the tumor microenvironment. J. Hematol. Oncol. 2021, 14, 10. [Google Scholar] [CrossRef]
- Silva, R.; Gullo, I.; Carneiro, F. The PD-1:PD-L1 immune inhibitory checkpoint in Helicobacter pylori infection and gastric cancer: A comprehensive review and future perspectives. Porto Biomed. J. 2016, 1, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Chen, W.; Xu, Z.P.; Gu, W. PD-L1 Distribution and Perspective for Cancer Immunotherapy—Blockade, Knockdown, or Inhibition. Front. Immunol. 2019, 10, 2022. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gong, Y.; Lv, Z.; Li, L.; Yuan, Y. Expression of PD1/PDL1 in gastric cancer at different microsatellite status and its correlation with infiltrating immune cells in the tumor microenvironment. J. Cancer 2021, 12, 1698–1707. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727. [Google Scholar]
- Li, X.; Pan, K.; Vieth, M.; Gerhard, M.; Li, W.; Mejiías-luque, R. JAK-STAT1 Signaling Pathway Is an Early Response to Helicobacter pylori Infection and Contributes to Immune Escape and Gastric Carcinogenesis. Int. J. Mol. Sci. 2022, 23, 4147. [Google Scholar] [CrossRef] [PubMed]
- Ubukata, Y.; Ogata, K.; Sohda, M.; Yokobori, T.; Shimoda, Y.; Handa, T.; Nakazawa, N.; Kimura, A.; Kogure, N.; Sano, A.; et al. Role of PD-L1 Expression during the Progression of Submucosal Gastric Cancer. Oncology 2021, 99, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Spirina, L.; Avgustinovich, A.; Afanas’ev, S.; Volkov, M.; Dobrodeev, A.; Cheremisina, O.; Kostromitsky, D. PD-L1 Status in Gastric Cancers, Association with the Transcriptional, Growth Factors, AKT/mTOR Components Change, and Autophagy Initiation. Int. J. Mol. Sci. 2021, 22, 11176. [Google Scholar] [CrossRef]
- Pan, S.; Li, K.; Huang, B.; Huang, J.; Xu, H.; Zhu, Z. Efficacy and safety of immune checkpoint inhibitors in gastric cancer: A network meta-analysis of well-designed randomized controlled trials. Ann. Transl. Med. 2021, 9, 290. [Google Scholar] [CrossRef]
- Khare, T.; Bissonnette, M.; Khare, S. CXCL12-CXCR4/CXCR7 Axis in Colorectal Cancer: Therapeutic Target in Preclinical and Clinical Studies. Int. J. Mol. Sci. 2021, 22, 7371. [Google Scholar] [CrossRef]
- Kim, J.; Bae, J.-S. Tumor-Associated Macrophages and Neutrophils in Tumor Microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kwon, W.A.; Joung, J.Y. Impact of Benign Prostatic Hyperplasia and/or Prostatitis on the Risk of Prostate Cancer in Korean Patients. World J. Mens. Health 2021, 39, 358. [Google Scholar] [CrossRef] [PubMed]
- Glaser, A.; Shi, Z.; Wei, J.; Lanman, N.A.; Ladson-Gary, S.; Vickman, R.E.; Franco, O.E.; Crawford, S.E.; Lilly Zheng, S.; Hayward, S.W.; et al. Shared Inherited Genetics of Benign Prostatic Hyperplasia and Prostate Cancer. Eur. Urol. Open Sci. 2022, 43, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Shah, A.A.; Nandakumar, K.; Lobo, R. Mechanistic targets for BPH and prostate cancer–a review. Rev. Environ. Health 2021, 36, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, B.; Liu, M.; Fu, X.; Ci, X.; Jun, A.; Fu, C.; Dong, G.; Wu, R.; Zhang, Z.; et al. KLF5 Is Crucial for Androgen-AR Signaling to Transactivate Genes and Promote Cell Proliferation in Prostate Cancer Cells. Cancers 2020, 12, 748. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, C. The roles and regulation of the KLF5 transcription factor in cancers. Cancer Sci. 2021, 112, 2097–2117. [Google Scholar] [CrossRef]
- Ma, J.B.; Bai, J.Y.; Zhang, H.B.; Jia, J.; Shi, Q.; Yang, C.; Wang, X.; He, D.; Guo, P. KLF5 inhibits STAT3 activity and tumor metastasis in prostate cancer by suppressing IGF1 transcription cooperatively with HDAC1. Cell Death Dis. 2020, 11, 466. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, H.-B.; Shi, Q.; Yang, C.; Ma, J.-B.; Jin, B.; Wang, X.; He, D.; Guo, P. KLF5 downregulation desensitizes castration-resistant prostate cancer cells to docetaxel by increasing BECN1 expression and inducing cell autophagy. Theranostics 2019, 9, 5464–5477. [Google Scholar] [CrossRef]
- Duan, X.; Liu, J.; Zhu, C.-C.; Wang, Q.-C.; Cui, X.-S.; Kim, N.-H.; Xiong, B.; Sun, S.-C. Cell Cycle RhoA-mediated MLC2 regulates actin dynamics for cytokinesis in meiosis RhoA-mediated MLC2 regulates actin dynamics for cytokinesis in meiosis. Cell Cycle 2016, 15, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Tamihardja, J.; Lawrenz, I.; Lutyj, P.; Weick, S.; Guckenberger, M.; Polat, B.; Flentje, M. Propensity score-matched analysis comparing dose-escalated intensity-modulated radiation therapy versus external beam radiation therapy plus high-dose-rate brachytherapy for localized prostate cancer. Strahlenther. Und Onkol. 2022, 198, 735–743. [Google Scholar] [CrossRef]
- Shiota, M.; Fujimoto, N.; Kashiwagi, E.; Eto, M. The Role of Nuclear Receptors in Prostate Cancer. Cells 2019, 8, 602. [Google Scholar] [CrossRef] [Green Version]
- Jiliang, S.; Hui, L.; Gonghui, L.; Renan, J.; Liang, S.; Mingming, C.; Chawnshang, C.; Xiujun, C. TR4 nuclear receptor enhances the cisplatin chemo-sensitivity via altering the ATF3 expression to better suppress HCC cell growth. Oncotarget 2016, 7, 32088–32099. [Google Scholar] [CrossRef] [Green Version]
- Chianese, A.; Santella, B.; Ambrosino, A.; Stelitano, D.; Rinaldi, L.; Galdiero, M.; Zannella, C.; Franci, G. Oncolytic viruses in combination therapeutic approaches with epigenetic modulators: Past, present, and future perspectives. Cancers 2021, 13, 2761. [Google Scholar] [CrossRef] [PubMed]
- Zunica, E.R.M.; Heintz, E.C.; Axelrod, C.L.; Kirwan, J.P. Obesity Management in the Primary Prevention of Hepatocellular Carcinoma. Cancers 2022, 14, 4051. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhou, J.K.; Peng, Y.; He, W.; Huang, C. The role of long noncoding RNAs in hepatocellular carcinoma. Mol. Cancer 2020, 19, 77. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. MTOR signaling pathway and mTOR inhibitors in cancer: Progress and challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Paquette, M.; El-Houjeiri, L.; Pause, A. mTOR Pathways in Cancer and Autophagy. Cancers 2018, 10, 18. [Google Scholar] [CrossRef]
- Hao, P.; Yu, J.; Ward, R.; Liu, Y.; Hao, Q.; An, S.; Xu, T. Eukaryotic translation initiation factors as promising targets in cancer therapy. Cell Commun. Signal. 2020, 18, 1–20. [Google Scholar] [CrossRef]
- Meegan, M.J.; Nathwani, S.; Twamley, B.; Zisterer, D.M.; O’Boyle, N.M. Piperlongumine (piplartine) and analogues: Antiproliferative microtubule-destabilising agents. Eur. J. Med. Chem. 2017, 125, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.-H.; Fu, Q.-H.; Liu, Y.; Jiang, K.; Guo, Q.-M.; Chen, Q.-Y.; Yan, B.; Wang, Q.-Q.; Shen, J.-G. Piperine suppresses tumor growth and metastasis in vitro and in vivo in a 4T1 murine breast cancer model. Acta Pharmacol. Sin. 2012, 33, 523–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Sheng, H.; Zhao, Y.; Zhu, G. Piperine Inhibits Cell Proliferation and Induces Apoptosis of Human Gastric Cancer Cells by Downregulating Phosphatidylinositol 3-Kinase (PI3K)/Akt Pathway. Med. Sci. Monit. 2020, 27, e928403-1. [Google Scholar] [CrossRef]
- Parama, D.; Rana, V.; Girisa, S.; Verma, E.; Daimary, U.D.; Thakur, K.K.; Kumar, A.; Kunnumakkara, A.B. The promising potential of piperlongumine as an emerging therapeutics for cancer. Explor. Target Anti Tumor Ther. 2021, 2, 323–354. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Shi, H. Sex Hormones and Their Receptors Regulate Liver Energy Homeostasis. Int. J. Endocrinol. 2015, 2015, 294278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Liu, X. Therapeutic Value of Estrogen Receptor α in Hepatocellular Carcinoma Based on Molecular Mechanisms. J. Clin. Transl. Hepatol. 2022, 10, 140–146. [Google Scholar] [CrossRef]
- Wu, Y.; Zheng, J.; Yan, Y.; Liu, J.; Zhou, Y. Gelsolin Can Be a Prognostic Biomarker and Correlated with Immune Infiltrates in Gastric Cancer. Int. J. Gen. Med. 2022, 15, 927–936. [Google Scholar] [CrossRef]
- Kitahara, C.M.; Schneider, A.B. Epidemiology of Thyroid Cancer. Cancer Epidemiol. Biomark. Prev. 2022, 31, 1284–1297. [Google Scholar] [CrossRef]
- Bogović Crnčić, T. Risk Factors for Thyroid Cancer: What Do We Know So Far? Acta Clin. Croat. 2020, 59, 66–72. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Lee, E.J.; Huang, M.G.; Park, Y.I.; Khullar, A.; Plodkowski, R.A. Diagnosis and treatment of patients with thyroid cancer. Am. Health Drug Benefits 2015, 8, 30–38. [Google Scholar]
- Tabatabaeian, H.; Yang, S.P.; Tay, Y. Non-coding rnas: Uncharted mediators of thyroid cancer pathogenesis. Cancers 2020, 12, 3264. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, M.; Zhang, L.; Lou, J.; Zhou, F.; Fang, M. Non-coding RNA in thyroid cancer—Functions and mechanisms. Cancer Lett. 2021, 496, 117–126. [Google Scholar] [CrossRef]
- De Martino, M.; Esposito, F.; Capone, M.; Pallante, P. Noncoding RNAs in Thyroid-Follicular-Cell-Derived Carcinomas. Cancers 2022, 14, 3079. [Google Scholar] [CrossRef] [PubMed]
- Dobruch, J.; Oszczudłowski, M. Bladder cancer: Current challenges and future directions. Medicina 2021, 57, 749. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Trinh, Q.D. Diagnosis and Staging of Bladder Cancer. Hematol. Oncol. Clin. N. Am. 2015, 29, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, G.; Guo, X.; Yao, H.; Wang, G.; Li, C. Non-coding RNA in bladder cancer. Cancer Lett. 2020, 485, 38–44. [Google Scholar] [CrossRef]
- Yu, W.; Yang, X.Y.; Zhang, Q.; Sun, L.; Yuan, S.T.; Xin, Y.J. Targeting GLS1 to cancer therapy through glutamine metabolism. Clin. Transl. Oncol. 2021, 23, 2253–2268. [Google Scholar] [CrossRef]
- Elaiwy, O.; El Ansari, W.; AlKhalil, M.; Ammar, A. Epidemiology and pathology of oral squamous cell carcinoma in a multi-ethnic population: Retrospective study of 154 cases over 7 years in Qatar. Ann. Med. Surg. 2020, 60, 195–200. [Google Scholar] [CrossRef]
- Yasin, M.M.; Abbas, Z.; Hafeez, A. Correlation of histopathological patterns of OSCC patients with tumor site and habits. BMC Oral Health 2022, 22, 305. [Google Scholar] [CrossRef]
- Li, S.; Mai, Z.; Gu, W.; Ogbuehi, A.C.; Acharya, A.; Pelekos, G.; Ning, W.; Liu, X.; Deng, Y.; Li, H.; et al. Molecular Subtypes of Oral Squamous Cell Carcinoma Based on Immunosuppression Genes Using a Deep Learning Approach. Front. Cell Dev. Biol. 2021, 9, 687245. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Lou, Q.Y.; Yang, W.Y.; Wang, Y.R.; Chen, R.; Wang, L.; Xu, T.; Zhang, L. The role of non-coding RNAs in drug resistance of oral squamous cell carcinoma and therapeutic potential. Cancer Commun. 2021, 41, 981–1006. [Google Scholar] [CrossRef] [PubMed]
- Jafari, F.; Javdansirat, S.; Sanaie, S.; Naseri, A.; Shamekh, A.; Rostamzadeh, D.; Dolati, S. Osteosarcoma: A comprehensive review of management and treatment strategies. Ann. Diagn. Pathol. 2020, 49, 151654. [Google Scholar] [CrossRef]
- Misaghi, A.; Goldin, A.; Awad, M.; Kulidjian, A.A. Osteosarcoma: A comprehensive review. SICOT J. 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickel, K.; Fang, F.; Tao, J. Molecular genetics of osteosarcoma. Bone 2017, 102, 69–79. [Google Scholar] [CrossRef]
- Sittiju, P.; Chaiyawat, P.; Pruksakorn, D.; Klangjorhor, J.; Wongrin, W.; Phinyo, P.; Kamolphiwong, R.; Phanphaisarn, A.; Teeyakasem, P.; Kongtawelert, P.; et al. Osteosarcoma-Specific Genes as a Diagnostic Tool and Clinical Predictor of Tumor Progression. Biology 2022, 11, 698. [Google Scholar] [CrossRef]
- Smolle, M.A.; Pichler, M. The role of long non-coding RNAs in osteosarcoma. Non Coding RNA 2018, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Kong, Y.; Dai, Z.; Liu, W.; Zhao, Z. The circular RNA circCRIM1 inhibits osteosarcoma progression through sponging miR-513. Mamm. Genome 2021, 32, 495–502. [Google Scholar] [CrossRef]
- Ji, X.; Shan, L.; Shen, P.; He, M. Circular RNA circ_001621 promotes osteosarcoma cells proliferation and migration by sponging miR-578 and regulating VEGF expression. Cell Death Dis. 2020, 11, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, K.; Wang, H.; Fang, Y.; Wang, Y.; Wei, L.; Chen, X.; Jiang, Z.; Wei, X.; Hu, Y. Histone Deacetylase 4 Promotes Osteosarcoma Cell Proliferation and Invasion by Regulating Expression of Proliferating Cell Nuclear Antigen. Front. Oncol. 2019, 9, 870. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhang, G.; Gao, Y.; Chen, S.; Cao, C. Circular RNA hsa_circ_0005909 modulates osteosarcoma progression via the miR-936/HMGB1 axis. Cancer Cell Int. 2020, 20, 305. [Google Scholar] [CrossRef]
- Huang, J.; Ni, J.; Liu, K.; Yu, Y.; Xie, M.; Kang, R.; Vernon, P.; Cao, L.; Tang, D. HMGB1 promotes drug resistance in osteosarcoma. Cancer Res. 2012, 72, 230–238. [Google Scholar] [CrossRef] [Green Version]
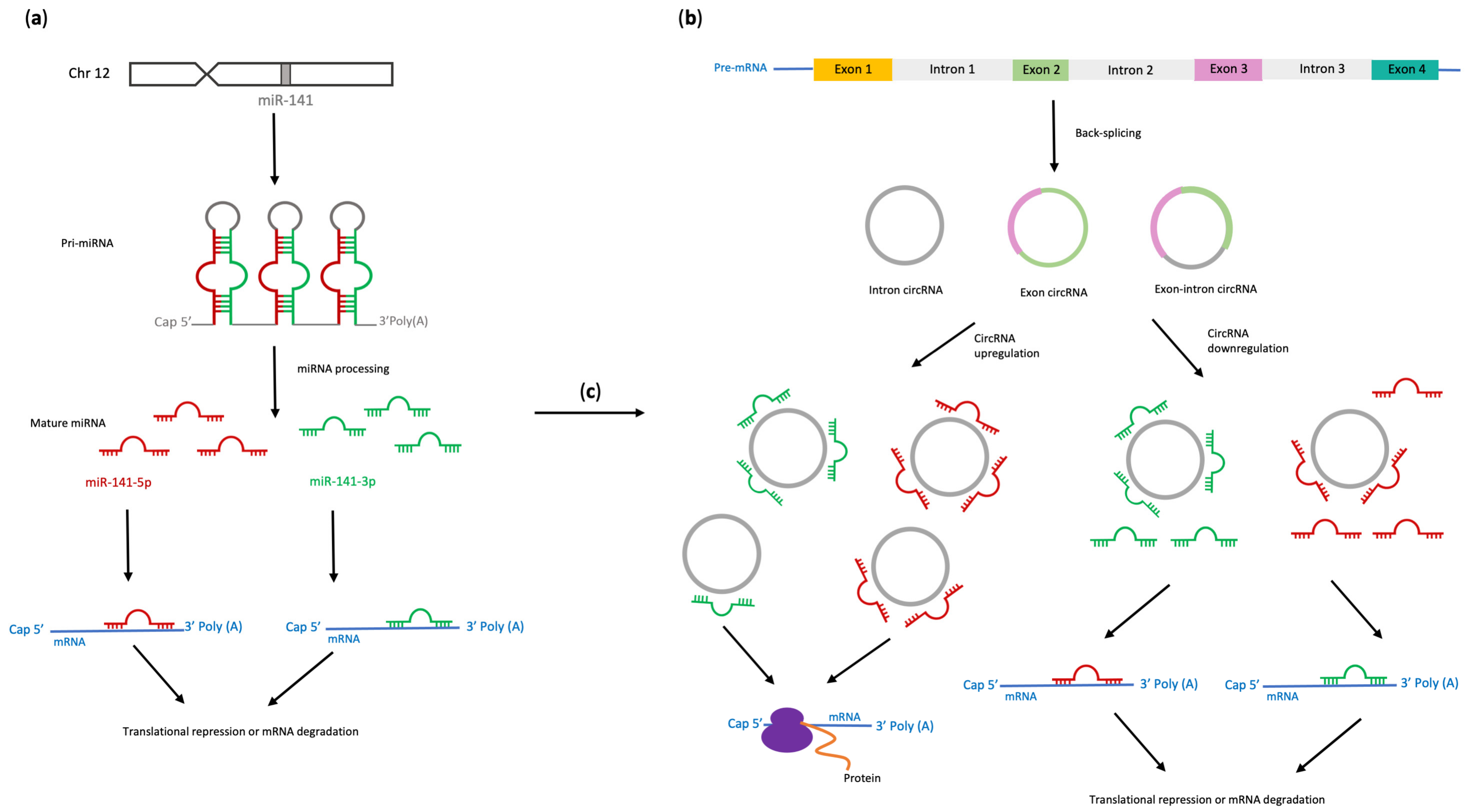
| Cancer Type | circRNA | Upregulated miR-141-3p Targets | Biological Effects | References |
|---|---|---|---|---|
| Breast cancer (BC) | hsa_circ_0009362 (circGNB1) | IGF1R | promotes proliferation and migration of TNBC cells; increases tumor growth and lung metastases in mouse xenograft model | [94] |
| hsa_circ_0075943 | AK2 | Knock-out of hsa_cicrc_0075943 suppresses the growth of cancer cells and induces apoptosis | [95] | |
| Lung adenocarcinoma (LUAD) | hsa_circ_102442 (circKEAP1) | KEAP1 | circKEAP1 sponges miR-141-3p which increases KEAP1 and decreases NRF2 expression and inhibits cancer growth | [96] |
| Gastric cancer (GC) | hsa_circ_0008583 (circDLG1) | CXCL12 | induces infiltration of myeloid-derived suppressor cells to impair the function of CD8+ T cells and promotes GC progression; enhances GC cells proliferation, migration, invasion, EMT, immune evasion, and metastasis in immunocompetent mice | [97] |
| Prostate cancer (PC) | hsa_circ_0001206 (circCRKL) | KLF5 | represses cell cycle, invasion, migration, stimulates apoptosis in vitro, and reduces tumor progression in vivo | [98] |
| hsa_circ_0001633 (circSOBP) | MYPT1 | inhibits migration and invasion of PC cells in vitro by the regulation of their ameboid migration and reduces metastasis in vivo | [99] | |
| hsa_circ_0004907 (circZEB1) | ZEB1 | TR4 induced circZEB1 expression decreases PC radiosensitivity via TR4/QKI/circZEB1/miR-141-3p/ZEB1 axis | [100] | |
| Hepatocellular carcinoma (HCC) | hsa_circ_100338 | RHEB | hsa_circ_100338 activates mTOR signaling pathway and is associated with poor prognosis | [101,102] |
| hsa_circ_100338 | ZEB1 | increases proliferation of HCC cells in vitro and in vivo | [103] | |
| hsa_circ_0008216 (circ-SMG1.72) | GSN | ERα suppresses circ-SMG1.72 and reduces HCC cell invasion via ERα/circ-SMG1.72/miR-141-3p/GSN axis | [104] | |
| Thyroid cancer (TC) | hsa_circ_0015278 (circRNA_100395) | circRNA_100395 is downregulated in papillary thyroid carcinoma tissues compared to normal thyroid tissues and benign thyroid lesions | [105] | |
| Bladder Cancer (BlC) | hsa_circ_0085361 (circTRPS1) | GLS1 | exosome-derived circTRPS1 decreases intracellular reactive oxygen species level and induces CD8+ T cell exhaustion in the tumor microenvironment which promotes BlC growth and metastasis via circTRPS1/miR-141-3p/GLS1 axis | [106] |
| Oral Squamous Cell Carcinoma (OSCC) | hsa_circ_0072387 | low hsa_circ_0072387 expression in OSCC tissues is associated with shorter overall survival of OSCC patients and related to higher TNM stage and tumor diameter | [107] | |
| Bone Cancer | circ-LRP6 | HDAC4, HMGB1 | promotes proliferation, migration, and invasion of osteosarcoma cells | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guz, M.; Jeleniewicz, W.; Cybulski, M. Interactions between circRNAs and miR-141 in Cancer: From Pathogenesis to Diagnosis and Therapy. Int. J. Mol. Sci. 2023, 24, 11861. https://doi.org/10.3390/ijms241411861
Guz M, Jeleniewicz W, Cybulski M. Interactions between circRNAs and miR-141 in Cancer: From Pathogenesis to Diagnosis and Therapy. International Journal of Molecular Sciences. 2023; 24(14):11861. https://doi.org/10.3390/ijms241411861
Chicago/Turabian StyleGuz, Małgorzata, Witold Jeleniewicz, and Marek Cybulski. 2023. "Interactions between circRNAs and miR-141 in Cancer: From Pathogenesis to Diagnosis and Therapy" International Journal of Molecular Sciences 24, no. 14: 11861. https://doi.org/10.3390/ijms241411861