Abstract
The gut microbiota has emerged as an important modulator of cardiovascular and renal homeostasis. The composition of gut microbiota in patients suffering from chronic kidney disease (CKD) is altered, where a lower number of bacteria producing short chain fatty acids (SCFAs) is observed. It is known that SCFAs, such as butyrate and acetate, have protective effects against cardiovascular diseases and CKD but their mechanisms of action remain largely unexplored. In the present study, we investigated the effect of butyrate and acetate on glomerular endothelial cells. Human glomerular microvascular endothelial cells (hgMVECs) were cultured and exposed to butyrate and acetate and their effects on cellular proliferation, mitochondrial mass and metabolism, as well as monolayer integrity were studied. While acetate did not show any effects on hgMVECs, our results revealed that butyrate reduces the proliferation of hgMVECs, strengthens the endothelial barrier through increased expression of VE-cadherin and Claudin-5 and promotes mitochondrial biogenesis. Moreover, butyrate reduces the increase in oxygen consumption induced by lipopolysaccharides (LPS), revealing a protective effect of butyrate against the detrimental effects of LPS. Taken together, our data show that butyrate is a key player in endothelial integrity and metabolic homeostasis.
1. Introduction
Chronic kidney disease (CKD) is a progressive disease affecting over 800 million people worldwide, with a high prevalence in patients with hypertension and type 2 diabetes mellitus (T2DM) [1]. CKD is characterized by glomerular injury [2], with the endothelial barrier being one of its main components. [3]. In the case of CKD, a high inflammatory state, increased oxidative stress levels and impaired nitric oxide (NO) production can damage the glomerular filtration system and cause endothelial dysfunction (ED) [4]. ED is characterized by a loss of cell-to-cell contacts which, together with an increased production of platelet-derived growth factor β (PDGF-β) and vascular endothelial growth factor A (VEGFA), promotes vascular instability and rarefaction [5,6,7,8]. This loss in endothelial integrity in CKD is also accompanied by altered mitochondrial function [9]. While mitochondrial homeostasis is regulated by cycles of fission–fusion and mitophagy–mitogenesis [10], this balance is skewed towards fission in T2DM and hypertension-driven hypoxia. This phenomenon causes excessive mitochondrial fragmentation, leading to mitochondrial dysfunction and a progressive reduction in the number of mitochondria [11,12].
The gut microbiota has recently emerged as a modulator of CKD pathology [13]. In CKD patients, the concentrations of microbiota-derived trimethylamine N-oxide (TMAO), indoxyl sulfate and p-cresyl sulfate in the blood are increased [14]. Because of poor renal function in CKD patients, these uremic toxins remain in the circulation, promote inflammation, thereby contributing to progressive kidney injury [15]. Moreover, these metabolites directly contribute to the microbiome imbalance by boosting uremic toxin-producing bacteria and lowering the ones producing short chain fatty acids [16,17]. Short chain fatty acids (SCFAs), including butyrate and acetate, are associated with beneficial effects on human health [18,19,20]. Previous studies showed that SCFAs also exert anti-inflammatory properties in cultured endothelial cells, as they inhibit leukocyte adhesion [21] and reduce expression of the pro-inflammatory proteins ICAM-1 and VCAM-1 [22]. Moreover, it has been reported that butyrate can reduce oxidative stress by improving mitochondrial function in STZ-treated pancreatic β-cells [23] and neuronal cells [24]. Butyrate is also able to counteract the pro-inflammatory activity of gut-derived lipopolysaccharides (LPS) [25], which, besides triggering inflammation and oxidative stress [26], promotes an excessive intestinal permeability [27]. Interestingly, CKD patients display an increased amount of LPS in both intestine and blood [28].
While previous studies showed that SCFAs can have protective effects on CKD progression [13,29], the direct effects of these metabolites on the glomerular endothelium remain still elusive. For this reason, we investigated the effect of butyrate and acetate on the function and metabolism of human glomerular microvascular endothelial cells (hgMVECs).
2. Results
2.1. Butyrate Decreases Proliferation of hgMVECs
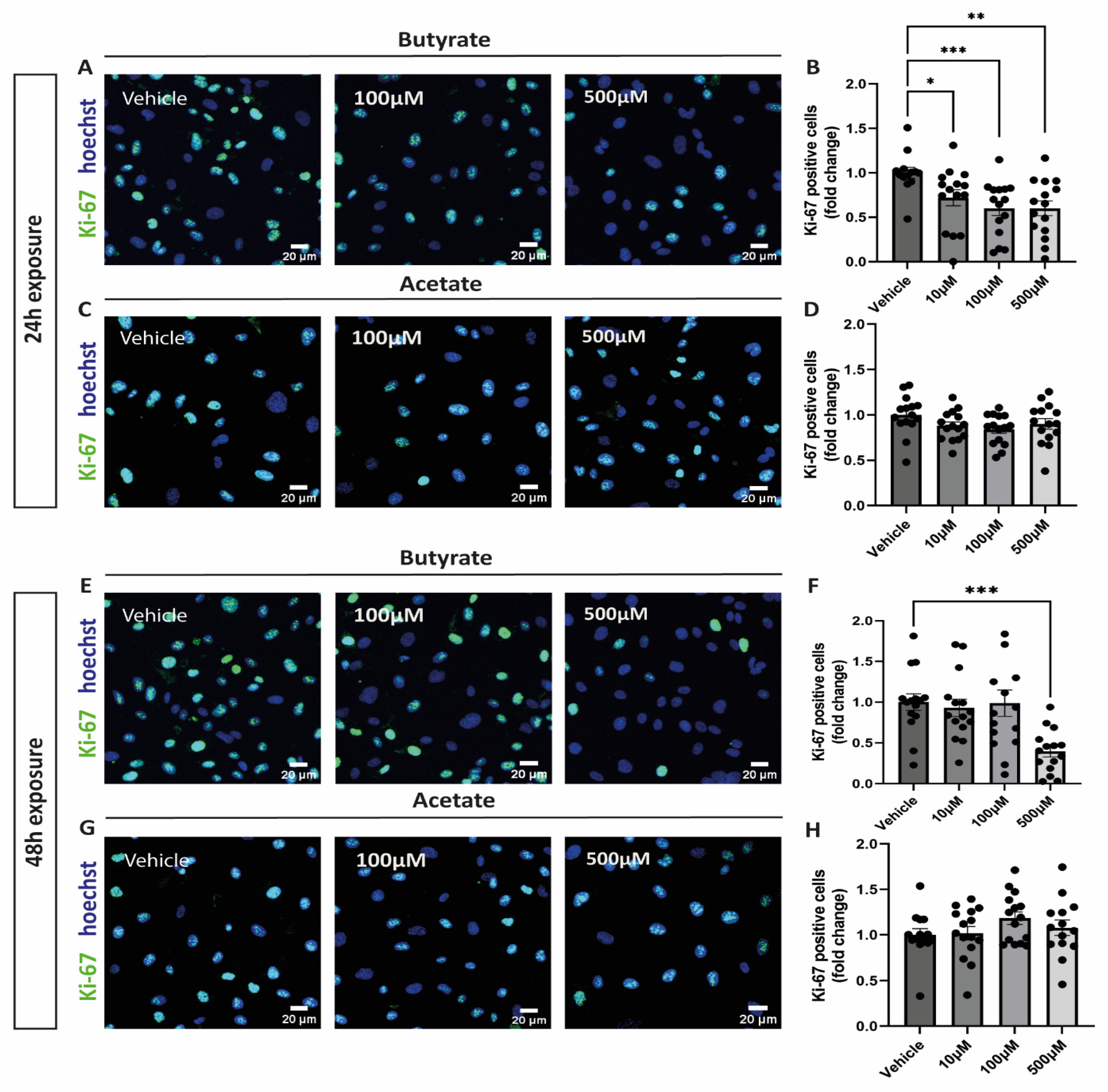
After exposing cells to increasing concentrations of butyrate or acetate for either 24 h or 48 h, cells were stained for ki-67 to identify proliferating cells. After 24 h incubation, butyrate significantly reduced proliferation of hgMVECs at every concentration tested (Figure 1A,B), unlike acetate (Figure 1C,D). After 48 h, only butyrate at 500 μM was effective in reducing proliferation of hgMVECs (Figure 1E,F).
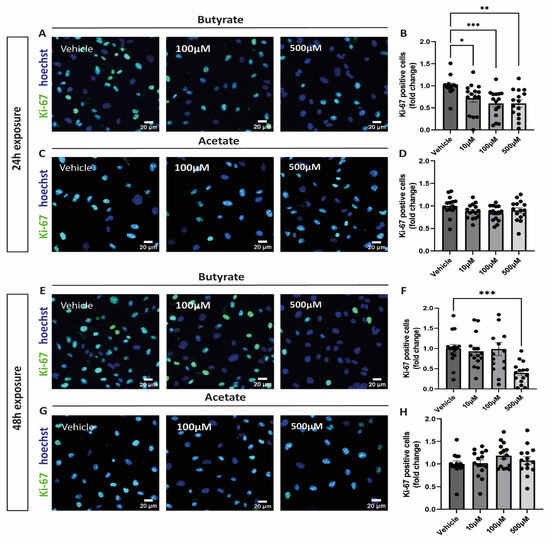
Figure 1.
Effect of butyrate and acetate on proliferation marker ki-67 in hgMVECs. hgMVECs have been exposed to either acetate or butyrate for 24 h (A–D) or 48 h (E–H). For the analysis, we counted the total number of nuclei (blue) in every picture and the percentage of ki-67 positive nuclei (green). The results are plotted as the fold change compared to vehicle. For every experiment (n = 3), we took 5 random pictures per condition, that were used for quantification. Data are presented as mean ± SEM. Statistical significance was analyzed by using One-Way ANOVA analysis, with * for p values < 0.05, ** for p values < 0.01 and *** for p values < 0.001.
2.2. Butyrate Improves Endothelial Monolayer Integrity by Increasing the Expression of Proteins Involved in Endothelial Junction Formation
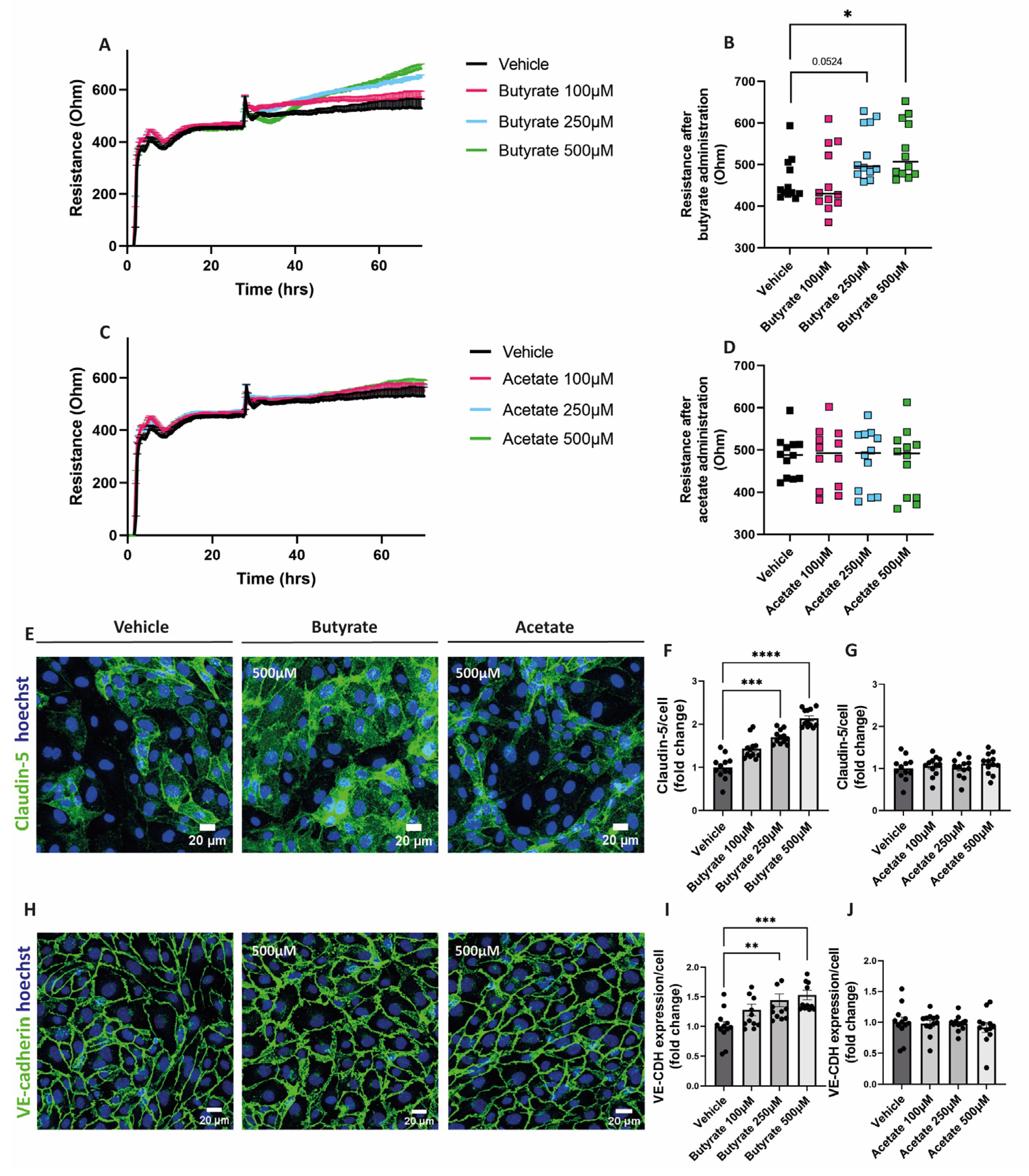
To assess the impact of the SCFAs on the endothelial monolayer resistance, cells were treated with either butyrate or acetate for at least 24 h after the formation of a stable monolayer. Butyrate at 250 μM and 500 μM increased the resistance of the endothelial monolayer (Figure 2A,B), while acetate did not show any effect on the resistance of the monolayer at any of the concentrations tested (Figure 2C,D).
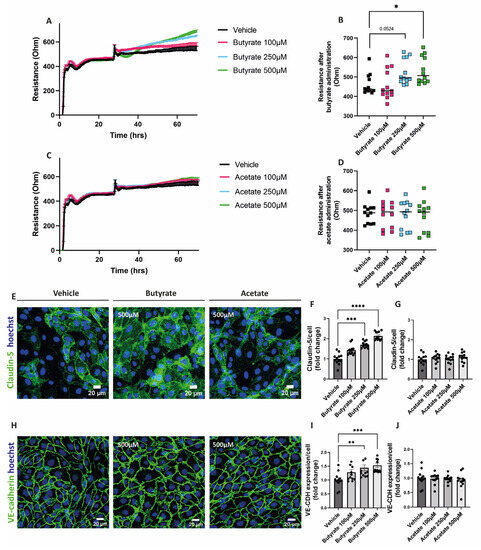
Figure 2.
Effect of butyrate and acetate on monolayer resistance and expression of claudin-5 and VE-cadherin in hgMVECs. For the ECIS experiments, hgMVECs were exposed to either butyrate (A) or acetate (C) for at least 24 h. For the analysis, we considered the time after we applied the treatment either with butyrate (B) or acetate (D). The data collected from every experiment (n = 3) are presented as mean ± SEM. For the stainings of claudin-5 (E–G) and VE-cadherin (H–J), hgMVECs were exposed to either butyrate or acetate for 48 h. For every experiment (n = 3), 4 random pictures per well were used for quantification purposes. The results of every experiment (n = 3) are plotted as the fold change compared to the vehicle. Data are presented as means ± SEM. Statistical significance was analyzed by using One-Way ANOVA analysis, with * for p values < 0.05, ** for p values < 0.01, *** for p values < 0.001 and **** for p values < 0.0001.
To visualize cell-to-cell contact, cells were fixed and stained for claudin-5 and VE-cadherin after 48 h exposure with butyrate or acetate. We observed that butyrate at 250 μM and at 500 μM increased the expression of both claudin-5 (Figure 2E,F) and VE-cadherin (Figure 2H,I). Acetate did not influence the expression of these proteins at any of the concentrations tested (Figure 2E,G,H,J).
2.3. Butyrate Increases Mitochondrial Mass and Influences the Expression of Genes Involved in Mitochondrial Homeostasis in hgMVECs
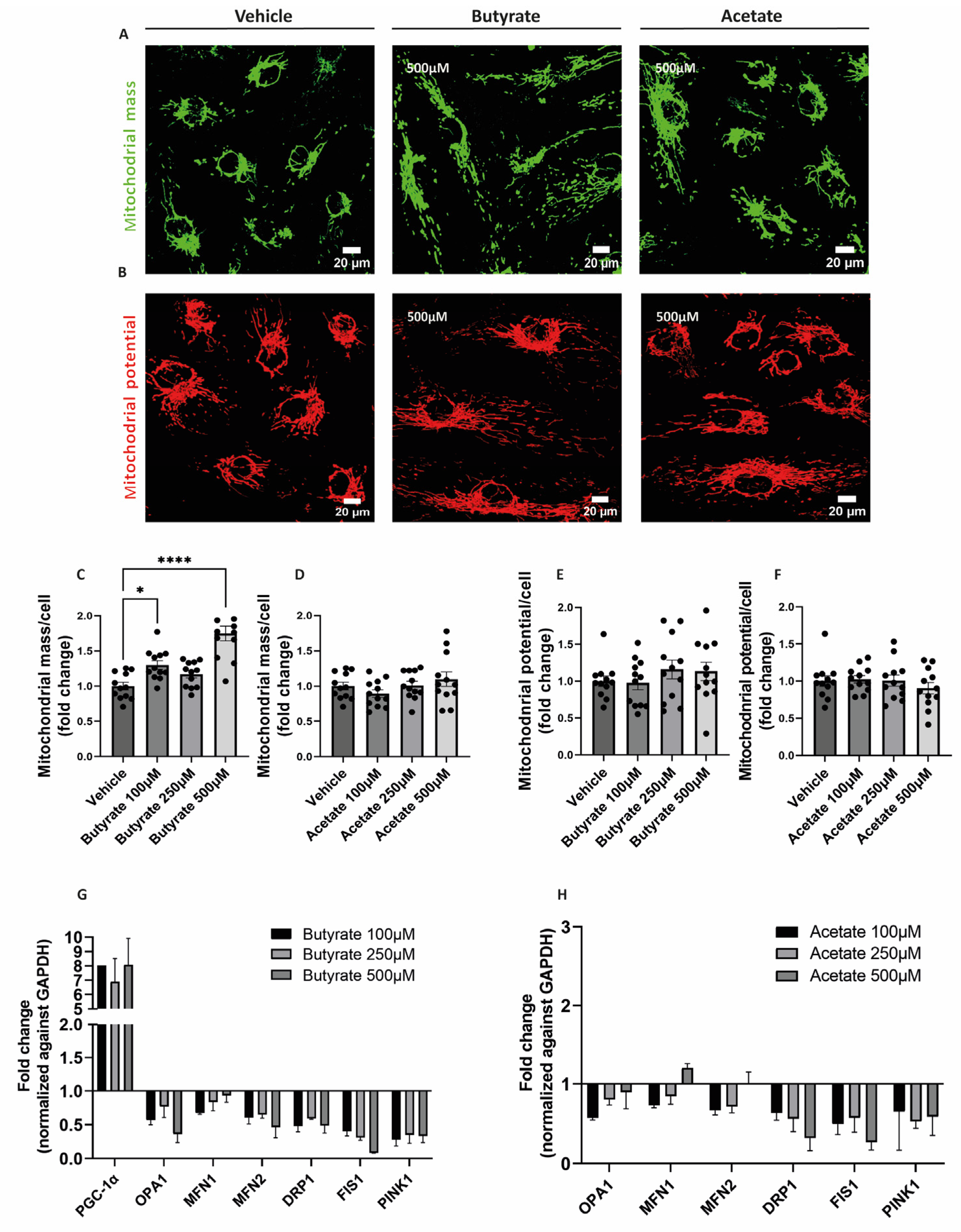
To visualize the effect of butyrate and acetate on mitochondrial mass and mitochondrial membrane potential, MitoTracker™ Green and MitoTracker™ Deep Red were used, respectively (Figure 2A,B). Upon 24 h-exposure to butyrate, the mitochondrial mass in hgMVECs was significantly increased, especially at the concentrations of 100 μM and 500 μM (Figure 2C). In contrast, acetate did not show the same effect on the mitochondrial mass at any of the concentrations tested (Figure 2D). While affecting the mitochondrial mass, butyrate did not influence mitochondrial membrane potential, nor did acetate (Figure 2E,F).
To assess the effect of butyrate and acetate on the expression of genes involved in mitochondrial metabolism, qPCR was performed on genes involved in mitochondrial fusion (OPA1, MFN1, MFN2) fission (DRP1, FIS1), mitophagy (PINK1), and mitochondrial biogenesis (PGC1-α). Both butyrate (Figure 3G) and acetate (Figure 3H) downregulated genes involved in mitochondrial fusion, fission and mitophagy. When exposing to butyrate, we were able to detect PGC1-α gene expression, which was expressed at a low level or not detected in control conditions. When exposing cells to acetate, expression of PGC1-α was not detected.
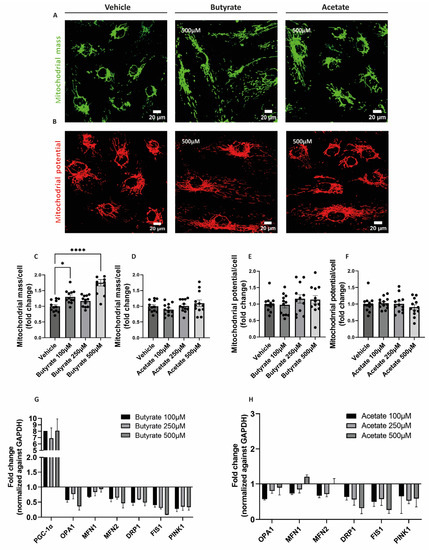
Figure 3.
Effect of butyrate and acetate on mitochondrial mass, mitochondrial membrane potential and gene expression of mitochondrial genes of hgMVECs. hgMVECs were exposed to butyrate or acetate for 24 h before staining with MitoTracker™ Green (A) or MitoTracker™ Deep Red (B). For the analysis of mitochondrial mass, we calculated the mitochondrial area, which was then normalized by the number of cells (C,D). For the mitochondrial potential, we calculated the intensity of the signal per cell (E,F). The results for both mitochondrial mass and mitochondrial potential were then plotted as fold change compared to the vehicle. For every experiment (n = 3) we took 4 random pictures per well, which were then used for quantification. For the qPCR experiments we collected mRNA from different experiments (n = 3) where cells were exposed to either butyrate or acetate for 24 h. The results of every experiment are plotted as the fold change compared to the control after normalization against GAPDH (G,H).The results of every experiment are plotted as the fold change compared to the vehicle. Data are presented as mean ± SEM. Statistical significance was calculated by using One-Way ANOVA analysis, with * for p values < 0.05 and **** for p values < 0.0001.
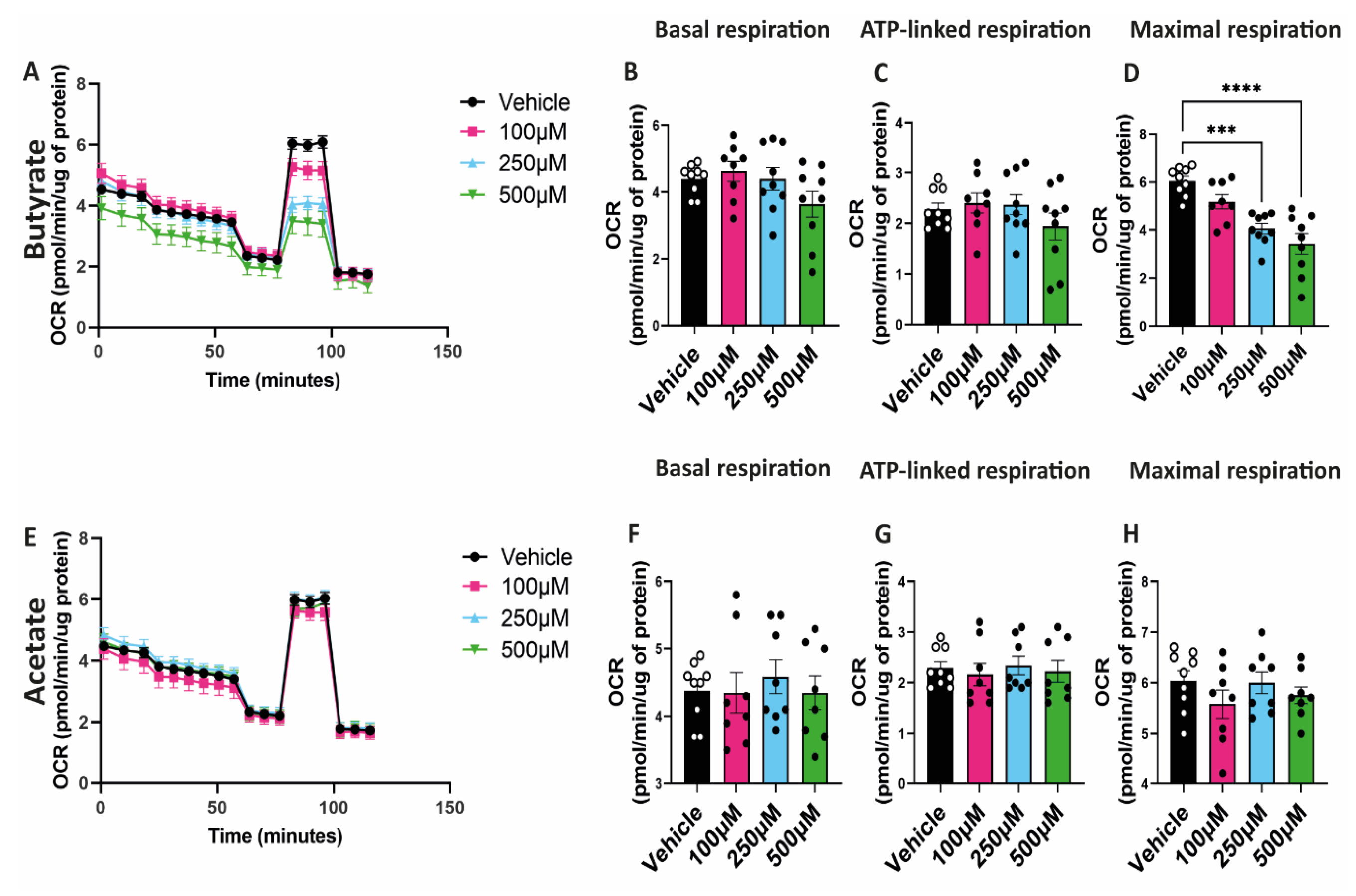
2.4. Butyrate Decreases Maximal Respiratory Capacity of hgMVECs
To study the effect of SCFAs on mitochondrial function, hgMVECs were exposed to butyrate or acetate for 24 h. Then, mitochondrial function was studied using a seahorse assay, specifically assessing the oxygen consumption rate. Butyrate decreased the maximal respiration of hgMVECs in a dose-dependent manner (Figure 4A,D). On the other hand, acetate did not show the same type of effect (Figure 4E,H). Unlike maximal respiration, basal respiration and ATP-linked respiration of hgMVECs were not significantly influenced by either butyrate (Figure 4B,C) or acetate (Figure 4F,G).
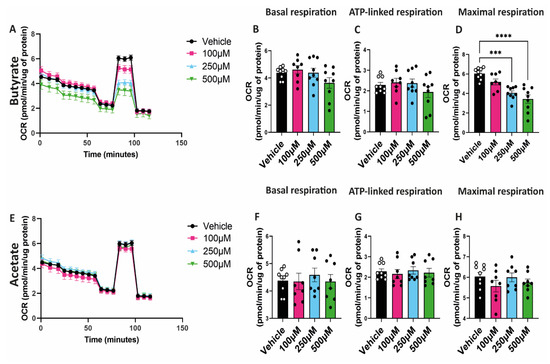
Figure 4.
Effect of butyrate and acetate on OCR of hgMVECs. hgMVECs were exposed to butyrate (A–D) or acetate (E–H) for 24 h before performing the seahorse assay. For every experiment (n = 3) we calculated basal respiration, ATP-linked respiration and maximal respiration. Data are presented as mean ± SEM. Statistical significance was calculated by using One-Way ANOVA analysis, with *** for p values < 0.001 and **** for p values < 0.0001.
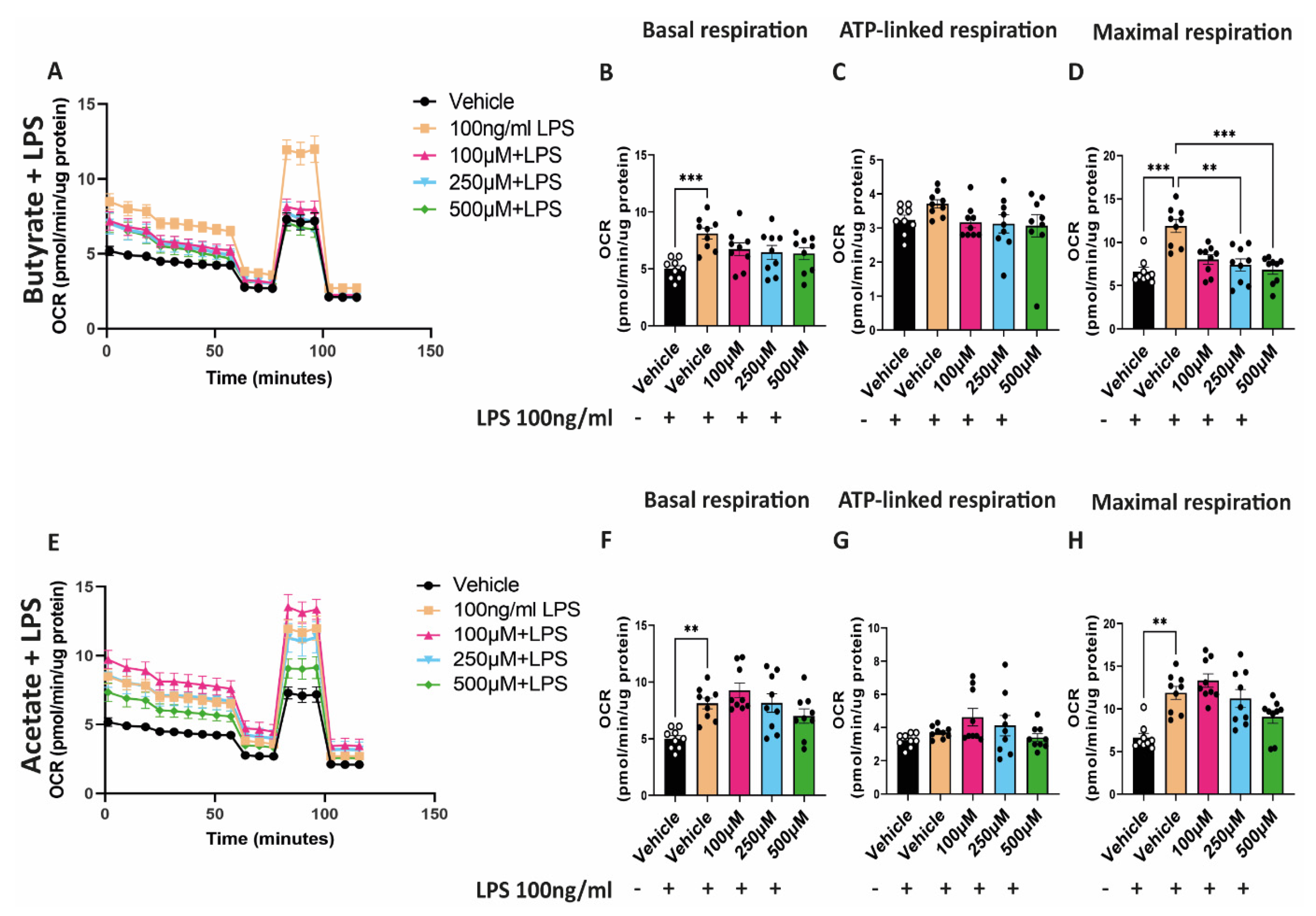
2.5. Butyrate Reduces the Increased Maximal Respiratory Capacity Induced by LPS
To study the effect of butyrate and acetate on mitochondrial function in combination with LPS, hgMVECs were exposed for 24 h to either butyrate or acetate and LPS. Exposure to LPS caused a general increase in OCR (Figure 5A,E), with a significant increase in basal respiration and maximal respiratory capacity (Figure 5B,D,F,H). Co-incubation of LPS with butyrate led to a significant decrease in the maximal respiratory capacity and in general to a lower OCR (Figure 5B–D). On the other hand, acetate did not show the same type of effect (Figure 5F–H). Although OCR seems to decrease when administering 500 μM of acetate, the difference when comparing to vehicle + 100 ng/mL LPS is not statistically significant.
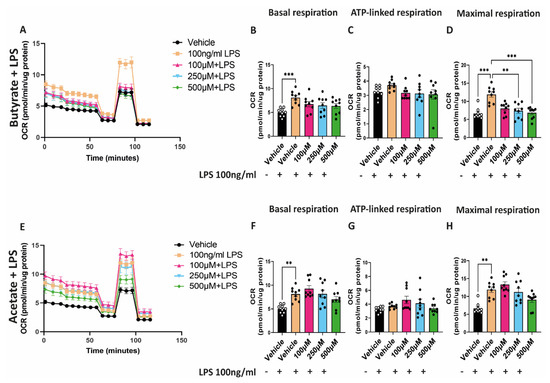
Figure 5.
Effect of butyrate (A–D) and acetate (E–H) in combination with LPS on OCR of hgMVECs. hgMVECs were co−incubated with butyrate or acetate and LPS for 24 h before performing seahorse assay. For every experiment (n = 3) we calculated basal respiration, ATP−linked respiration and maximal respiration. Data are presented as mean ± SEM. Statistical significance was calculated by using One-Way ANOVA analysis, with ** for p values < 0.01, *** for p values < 0.001.
3. Discussion
In the present study, we explored the effect of butyrate and acetate on human glomerular microvascular endothelial cells (hgMVECs). Specifically, we aimed to understand whether these compounds affect the homeostasis of glomerular endothelial cells and whether SCFAs might play a protective role against endothelial dysfunction (ED).
3.1. Butyrate Inhibits Endothelial Cell Proliferation
One of the main features of ED, especially in diabetic conditions, involves the uncontrolled proliferation of endothelial cells [30], which is mainly caused by an unbalanced production of VEGFA and NO. The latter helps to control proliferation of glomerular endothelial cells, but in diabetic conditions the producing enzyme, endothelial NO synthase (eNOS), is impaired. This leads to a lower production of NO which in combination with an increased production of VEGFA by podocytes causes uncontrolled proliferation [31,32]. In our study, we showed that butyrate can reduce proliferation of glomerular endothelial cells and potentially keep proliferation under control. This action of butyrate can be due to its ability to restore NO production through GPR41/43 activation [33] but also to its epigenetic activity. Butyrate is a well-known and potent histone deacetylase (HDAC) inhibitor and in this way, it can influence the expression of genes, also of the ones involved in cellular proliferation. For instance, a previous study showed that butyrate decreases the expression of the proliferative genes TES and PGF through HDC inhibition [34]. At the same time, SCFAs can have direct metabolic effects as they enter and boost the tricarboxylic (TCA) cycle, while reducing glycolysis. As a consequence, the proliferation of endothelial cells may also be reduced [35].
3.2. Butyrate and Increases the Resistance of the Endothelial Monolayer
The ability of butyrate to modulate the genetic profile of hgMVECs is evident also when looking at the expression of proteins involved in cell-to-cell adhesion. Treatment with butyrate resulted in the upregulated expression of VE-cadherin and claudin-5 and consequently in an increased resistance of the endothelial monolayer. The integrity of the endothelial monolayer is crucial for the functioning of any type of vessel, so that the proper degree of permeability is guaranteed. However, stressors such as hypoxia or oxidative stress can influence vascular permeability by hampering barrier function and integrity. For instance, exposure of endothelial cells to hydrogen peroxide promotes a loss of occludin and cadherin [36], while a previous study revealed that monolayer resistance of endothelial cells was impaired after exposure to a pro-inflammatory stimulus. Interestingly, barrier integrity was then restored upon incubation with vitamin D, which coincided with increased expression of VE-cadherin [37]. In a similar way, inflammation and oxidative stress can also hamper the integrity of the blood–brain barrier (BBB) via an increased activation of the Nf-kΒ/p65 pathway. This leads to a lower expression of claudin-5 and therefore to a weakened BBB. However, it has been shown that sodium butyrate can reduce the activity of the Nf-kΒ/p65 pathway and in this way restore the expression of claudin-5 [38].
Therefore, the increase in expression of VE-cadherin and also claudin-5 induced by butyrate suggest that this type of short chain fatty acid can play a pivotal role in protecting the integrity of the renal endothelial barrier against ED.
3.3. Butyrate Regulates Mitochondrial Biogenesis and Mitochondrial Function
As previously mentioned, excessive mitochondrial fragmentation and loss is one of the main features underlying mitochondrial dysfunction and, ultimately, kidney injury [39]. Interestingly, in our study, we observed a clear upregulation of the mitogenesis regulator PGC1-α upon butyrate exposure, which coincided with an increase in mitochondrial mass. Importantly, when staining with MitoTrackerTM Deep red, we could also observe that the increase in mitochondrial mass was not followed by a decrease in mitochondrial membrane potential, suggesting that this is not disturbed by butyrate. However, the higher number of mitochondria did not correspond to an augmented mitochondrial function, which declined instead in a dose-dependent manner. This might be explained by the action of butyrate as an HDAC inhibitor and by the presence of a potential negative feedback mechanism. When the ratio of acetyl-CoA/CoA increases excessively, acetyl-CoA has been shown to inhibit the activity of pyruvate dehydrogenases and consequently the amount of substrate available for mitochondrial respiration [40].
This could also partially explain how butyrate is able to restore mitochondrial function upon LPS treatment. LPS significantly increased basal respiration and maximal respiration of hgMVECs, which is necessary to provide energy to support protein and lipid production when endothelial cells are activated [41]. These cells respond to LPS stimulation by increasing mitochondrial respiration through the inhibition of the forkhead box protein O1/pyruvate dehydrogenase kinase 4 (FOXO1/PDK4) pathway, which regulates the amount of pyruvate that can enter the TCA cycle [42]. Interestingly, fatty acid supplementation seems to upregulate PKD4 [43] and this action could also contribute to the reduction in OCR when co-incubating butyrate and LPS. Taken all together, these observations suggest that butyrate can protect glomerular endothelial cells against mitochondrial dysfunction.
Unlike butyrate, acetate did not exert an effect on hgMVECs at any of the concentrations tested. One of the possible explanations underlying these observations is that butyrate is considered to be a more potent HDAC inhibitor compared to acetate [44] but also that butyrate and acetate have a different metabolic turnover. For instance, it is known that acetate shows a higher affinity for acyl-CoA short-chain synthetases (ACSS) than butyrate, and this induces a more rapid conversion of acetate in acetyl-CoA, when compared to butyrate [45].
3.4. Possible Clinical Implications
Our data suggest that butyrate plays a pivotal role in glomerular endothelial homeostasis, which has not only cellular but also potential clinical implications. With respect to CKD for instance, a recent study [16] showed that fecal and serum SCFAs are remarkably higher in healthy controls than in patients affected by CKD. Moreover, the progression of CKD negatively correlates with the amount of butyrate detected in the serum of the patients. This suggests that butyrate supplementation might have potential beneficial effects on kidney function, as it has been illustrated in several animal studies [46,47,48]. Besides helping to preserve kidney function, SCFAs seem to be beneficial for the whole cardiovascular system, by showing anti-hypertensive effects [49] and protecting from atherosclerosis [50] and heart failure [51]. Interestingly, the benefits of butyrate also go beyond the cardiovascular system, as they have been observed for instance acting on the central nervous system [52] and on intestinal barrier integrity [53,54]. The latter, in particular, is pivotal in preventing toxins produced in the gut from reaching other organs through the circulation. Importantly, butyrate administration might also have a long-term positive effect since it is able to reshape the gut microbiota composition and progressively boost and replenish SCFAs bacteria [55].
Although most of the studies have been conducted on cells and animals so far, all the gathered data suggest that butyrate supplementation might be an important ally in preventing the onset and the progression of cardiovascular and other severe diseases.
3.5. Study Limitations
The main limitation of this study is that cells have been treated with butyrate and acetate in a healthy setup, except for the experiments involving LPS. For this reason, in the future we are planning to study the effect of butyrate in a disease-mimicking environment, for instance by pre-incubating glomerular endothelial cells with diabetic serum and oxidative stress inducers. In this way we aim to further characterize the mechanisms of action of SCFAs on hgMVECs in endothelial dysfunction and CKD.
In conclusion, in our study we showed that in human glomerular endothelial cells butyrate decreases cellular proliferation and promotes mitochondrial biogenesis as well as monolayer integrity. Moreover, butyrate can counteract the respiration burst induced by LPS, suggesting a protective effect of butyrate against inflammation. Therefore, our data indicate that butyrate might play a pivotal role in preventing the onset and progression of endothelial dysfunction in CKD.
4. Materials and Methods
4.1. Primary Human Glomerular Microvascular Endothelial Cells
Primary human glomerular microvascular endothelial cells (hgMVECs) were purchased at Cell-System (Kirkland, WA, USA; ACBRI-128). Cells were cultured at 37 °C with 5% CO2 in full endothelial growth medium-2 (EGM-2) supplemented with 100 IU/mL penicillin and 100 μg/mL streptomycin. For the experiments, cells were left attached for 24 h and then treated for 24 h or 48 h with sodium butyrate (Sigma, St. Louis, MI, USA, 303410) or sodium acetate (Sigma, St. Louis, MI, USA; S2889-250G), or for 24 h with sodium butyrate/sodium acetate and 100 ng/mL of lipopolysaccharides from E. Coli O111:B4 (Sigma, St. Louis, MI, USA; L4391).
4.2. Ki-67 Staining for Proliferating Cells
Cells were stained with a monoclonal antibody against ki-67 (556003; BD-Pharmingen, San Diego, CA; USA) to visualize proliferating cells. First, cells were fixated with a solution at 4% of paraformaldehyde (PFA) for 10 min at room temperature. Afterwards, blocking and permeabilization steps were performed with a solution of 0.3% Triton-1% BSA for 10 min. Cells were then incubated with the primary antibody against ki-67 for 1.5 h at room temperature. After washing with PBS, cells were incubated with secondary antibody (Invitrogen, Waltham, MA, USA; A-11001) and stained with Hoechst (Thermo Fisher, Waltham, MA, USA; H3569) to visualize the nuclei for 45 min. After washing with PBS, coverslips were mounted with ProLong Gold (Thermo Fisher, Waltham, MA, USA; P36930) and cells were visualized using fluorescent microscopy.
4.3. Mitochondrial Staining with MitoTracker™
MitoTracker™ Green (Thermo Fisher, Waltham, MA, USA; M7514) and MitoTracker™ Deep Red (Thermo Fisher, Waltham, MA, USA; M22426), were used to stain for mitochondrial mass and mitochondrial membrane potential, respectively. Cells were incubated with 25 nM of MitoTracker™ Green or MitoTracker™ Deep Red for 30 min at 37 °C. Afterwards, cells were washed with warm medium and directly imaged with confocal microscopy.
4.4. Seahorse Assay
To study mitochondrial metabolism, seahorse assay experiments were performed. For this assay, cells were cultured in a Seahorse XF96 polystyrene tissue culture plate (Agilent technology, Santa Clara, CA, USA; 102416-100) coated with 1% gelatin. Cells were incubated with acetate or butyrate only, or with acetate/butyrate and LPS. Prior to the run, cells were incubated for 1 h at 37 °C without CO2 with Seahorse XF base medium (Agilent technology, Santa Clara, CA, USA; 103334-100) supplemented with L-glutamine. To measure oxygen consumption rate (OCR), we used the following compounds during the assay: 10 mM glucose (Thermo Fisher, Waltham, MA, USA; 15023021), 5 μM oligomycin A (Cayman Sanbio, Tokyo, Japan; 11342), 1 μM FCCP (Sigma, Waltham, MA, USA; C2920), 1 μM antimycin a (Sigma, Waltham, MA, USA; A8674) and 1 μM rotenone (Sigma, Waltham, MA, USA; R8875). Once the run terminated, cellular protein content was determined with a Pierce™ BCA-protein kit (Thermo Fisher, 23225). OCR values were then normalized to the protein content of the corresponding well.
4.5. Barrier Function Assay
Endothelial barrier resistance was analyzed by using an electric cell-substrate impedance sensing system (ECIS, Singapore, Applied BioPhysics). Cells were cultured on a ECIS Cultureware 96W20idf PET (Ibidi, Fitchburg, WI, USA; 72098) previously coated with 10 mM of L-cysteine, and then 1% gelatin. When cells were in the plate for 24 h and reached baseline resistance, butyrate or acetate were added to the plate.
4.6. Staining for VE-Cadherin and Claudin-5
Cells were stained with antibodies against VE-cadherin (Pharmingen, San Diego, CA, USA; 55561) and claudin-5 (Invitrogen, Waltham, MA, USA; 35-2500) to study the expression of these proteins as markers of endothelial junction formation. After incubating the cells with butyrate or acetate, we performed a fixation step with 4% PFA and a permeabilization-blocking step with 0.3% triton-1% BSA. Cells were then incubated with the primary antibody against either VE-cadherin or claudin-5 for 1.5 h at room temperature. After washing with PBS, cells were incubated with secondary antibody (Invitrogen, Waltham, MA, USA; A-11001) and stained with Hoechst (Thermo Fisher, Waltham, MA, USA; H3569) to visualize the nuclei for 45 min. After washing with PBS, cells were ready to be imaged with fluorescence microscopy.
4.7. Quantitative PCR
RNA was isolated by using an RNeasy Mini Kit (Qiagen, Singapore, 74106) according to manufacturer’s instructions. cDNA was then produced from the RNA using a M-MLV Reverse Transcriptase Kit (M1701; Promega, Singapore), while qPCR was performed with SYBR Select Master Mix (4472908, Waltham, MA, USA; Applied Biosystems). Forward and reverse primers of the following genes were used to perform qPCR analysis (Table 1).

Table 1.
Primer sequences for qPCR analysis.
4.8. Analysis and Statistics
Analysis of the images was performed with ImageJ-win64 (Fiji). The results are shown as mean ± SEM, while n represents the number of biological independent experiments. Differences between groups were assessed with One-Way ANOVA, where p values of <0.05 were considered statistically significant. Specifically, one star is used to indicate p values between 0.05 and 0.01, two stars for p values between 0.01 and 0.001 and three stars for p values between 0.001 and 0.0001.
Author Contributions
M.N.N., R.B., B.M.V.d.B. and J.I.R. contributed to designing the research; M.N.N. conducted the experiments and performed data collection and analysis; M.N.N. and J.I.R. wrote the manuscript; R.B., B.M.V.d.B. and A.J.V.Z. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Rembrandt Institute for Cardiovascular Science.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to acknowledge B. Evarts and A.J. van der Ham for helping with the seahorse assay and Z. Li for providing sodium butyrate in the first place. We would also like to thank X.M. Spijkers for proofreading and M.S. Zuurmond for helping with designing the graphical abstract.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Kovesdy, C.P. Epidemiology of chronic kidney disease: An update 2022. Kidney Int. Suppl. 2022, 12, 7–11. [Google Scholar] [CrossRef]
- Barton, M.; Sorokin, A. Endothelin and the Glomerulus in Chronic Kidney Disease. Semin. Nephrol. 2015, 35, 156–167. [Google Scholar] [CrossRef]
- Menon, M.C.; Chuang, P.Y.; He, C.J. The glomerular filtration barrier: Components and crosstalk. Int. J. Nephrol. 2012, 2012, 749010. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Mallamaci, F.; Zoccali, C. Endothelial Dysfunction in Chronic Kidney Dis-ease, from Biology to Clinical Outcomes: A 2020 Update. J. Clin. Med. 2020, 9, 2359. [Google Scholar] [CrossRef]
- Sol, M.; van den Born, J.K.J.; van den Heuvel, M.C.; van der Vlag, J.; Krenning, G.; Hillebrands, J.L. Glomerular Endothelial Cells as Instigators of Glomerular Sclerotic Diseases. Front. Pharmacol. 2020, 11, 573557. [Google Scholar] [CrossRef]
- Faller, D.V. Endothelial cell responses to hypoxic stress. Clin. Exp. Pharmacol. Physiol. 1999, 26, 74–84. [Google Scholar] [CrossRef]
- Eleftheriadis, T.; Antoniadi, G.; Pissas, G.; Liakopoulos, V.; Stefanidis, I. The renal endothelium in diabetic nephropathy. Ren. Fail. 2013, 35, 592–599. [Google Scholar] [CrossRef]
- Tufro, A.; Veron, D. VEGF and podocytes in diabetic nephropathy. Semin. Nephrol. 2012, 32, 385–393. [Google Scholar]
- Bhatia, D.; Capili, A.; Choi, M.E. Mitochondrial dysfunction in kidney injury, inflammation, and disease: Potential therapeutic approaches. Kidney Res. Clin. Pract. 2020, 39, 244–258. [Google Scholar] [CrossRef]
- Ma, K.; Chen, G.; Li, W.; Kepp, O.; Zhu, Y.; Chen, Q. Mitophagy, Mitochondrial Homeostasis, and Cell Fate. Front. Cell Dev. Biol. 2020, 8, 467. [Google Scholar]
- Ren, L.; Han, F.; Xuan, L.; Lv, Y.; Gong, L.; Yan, Y.; Wan, Z.; Guo, L.; Liu, H.; Xu, B.; et al. Clusterin ameliorates endothelial dysfunction in diabetes by suppressing mi-tochondrial fragmentation. Free Radic. Biol. Med. 2019, 145, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, D.C.; Brune, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [PubMed]
- Magliocca, G.; Mone, P.; Di Iorio, B.R.; Heidland, A.; Marzocco, S. Short-Chain Fatty Acids in Chronic Kidney Disease: Focus on Inflam-mation and Oxidative Stress Regulation. Int. J. Mol. Sci. 2022, 23, 5354. [Google Scholar] [CrossRef]
- Wehedy, E.; Shatat, I.F.; Al Khodor, S. The Human Microbiome in Chronic Kidney Dis-ease: A Double-Edged Sword. Front. Med. 2021, 8, 790783. [Google Scholar]
- Lau, W.L.; Kalantar-Zadeh, K.; Vaziri, N.D. The Gut as a Source of Inflammation in Chronic Kidney Disease. Nephron 2015, 130, 92–98. [Google Scholar] [PubMed]
- Wang, S.; Lv, D.; Jiang, S.; Jiang, J.; Liang, M.; Hou, F.; Chen, Y. Quantitative reduction in short-chain fatty acids, especially butyrate, contributes to the progression of chronic kidney disease. Clin. Sci. 2019, 133, 1857–1870. [Google Scholar]
- Gao, B.; Jose, A.; Alonzo-Palma, N.; Malik, T.; Shankaranarayanan, D.; Regunathan-Shenk, R.; Raj, D.S. Butyrate producing microbiota are reduced in chronic kidney diseases. Sci. Rep. 2021, 11, 23530. [Google Scholar] [CrossRef]
- Sanna, S.; van Zuydam, N.R.; Mahajan, A.; Kurilshikov, A.; Vila, A.V.; Vosa, U.; Mujagic, Z.; Masclee, A.A.M.; Jonkers, D.; Oosting, M.; et al. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar]
- Xiong, R.G.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Shang, A.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health Benefits and Side Effects of Short-Chain Fatty Acids. Foods 2022, 11, 2863. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar]
- Menzel, T.; Lührs, H.; Zirlik, S.; Schauber, J.; Kudlich, T.; Gerke, T.; Gostner, A.; Neumann, M.; Melcher, R.; Scheppach, W. Butyrate Inhibits Leukocyte Adhesion to Endothelial Cells viaModulation of VCAM-1. Inflamm. Bowel Dis. 2004, 10, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Zapolska-Downar, D.; Siennicka, A.; Kaczmarczyk, M.; Kolodziej, B.; Naruszewicz, M. Butyrate inhibits cytokine-induced VCAM-1 and ICAM-1 ex-pression in cultured endothelial cells: The role of NF-kappaB and PPARalpha. J. Nutr. Biochem. 2004, 15, 220–228. [Google Scholar] [PubMed]
- Hu, S.; Kuwabara, R.; de Haan, B.J.; Smink, A.M.; de Vos, P. Acetate and Butyrate Improve beta-cell Metabolism and Mitochondrial Respi-ration under Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 1542. [Google Scholar]
- Rose, S.; Bennuri, S.C.; Davis, J.E.; Wynne, R.; Slattery, J.C.; Tippett, M.; Delhey, L.; Melnyk, S.; Kahler, S.G.; MacFabe, D.F.; et al. Butyrate enhances mitochondrial function during oxidative stress in cell lines from boys with autism. Transl. Psychiatry 2018, 8, 42. [Google Scholar] [PubMed]
- Li, M.; van Esch, B.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory Effects of Short Chain Fatty Acids on Lipopolysaccha-ride- or Tumor Necrosis Factor alpha-Stimulated Endothelial Cells via Activation of GPR41/43 and Inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef]
- Qin, X.; Zou, H. The role of lipopolysaccharides in diabetic retinopathy. BMC Ophthalmol. 2022, 22, 86. [Google Scholar]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced in-flammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Kim, S.M.; Song, I.H. The clinical impact of gut microbiota in chronic kidney disease. Korean J. Intern. Med. 2020, 35, 1305–1316. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. The role and mechanism of gut microbiota-derived short-chain fatty in the prevention and treatment of diabetic kidney disease. Front. Immunol. 2022, 13, 1080456. [Google Scholar]
- Hohenstein, B.; Hausknecht, B.; Boehmer, K.; Riess, R.; Brekken, R.A.; Hugo, C.P. Local VEGF activity but not VEGF expression is tightly regulated during diabetic nephropathy in man. Kidney Int. 2006, 69, 1654–1661. [Google Scholar]
- Cha, D.R.; Kang, Y.S.; Han, S.Y.; Jee, Y.H.; Han, K.H.; Han, J.Y.; Kim, Y.S.; Kim, N.H. Vascular endothelial growth factor is increased during early stage of diabetic nephropathy in type II diabetic rats. J. Endocrinol. 2004, 183, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Harris, R.C. Renal Endothelial Dysfunction in Diabetic Nephropathy. Cardiovasc. Hematol. Disord. Targets 2014, 14, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; de la Visitacion, N.; Aguilera-Sanchez, N.; Redondo, J.M.; Duarte, J. Protective Effects of Short-Chain Fatty Acids on Endothelial Dysfunc-tion Induced by Angiotensin II. Front. Physiol. 2020, 11, 277. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg effect dictates the mechanism of butyrate-mediated his-tone acetylation and cell proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef]
- Eelen, G.; de Zeeuw, P.; Treps, L.; Harjes, U.; Wong, B.W.; Carmeliet, P. Endothelial Cell Metabolism. Physiol. Rev. 2018, 98, 3–58. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Endothelial Barrier and Its Abnor-malities in Cardiovascular Disease. Front. Physiol. 2015, 6, 365. [Google Scholar] [CrossRef]
- Schroder-Heurich, B.; von Hardenberg, S.; Brodowski, L.; Kipke, B.; Meyer, N.; Borns, K.; von Kaisenberg, C.S.; Brinkmann, H.; Claus, P.; von Versen-Hoynck, F. Vitamin D improves endothelial barrier integrity and counteracts inflammatory effects on endothelial progenitor cells. FASEB J. 2019, 33, 9142–9153. [Google Scholar] [CrossRef]
- Fock, E.; Parnova, R. Mechanisms of Blood-Brain Barrier Protection by Microbiota-Derived Short-Chain Fatty Acids. Cells 2023, 12, 657. [Google Scholar]
- Galvan, D.L.; Green, N.H.; Danesh, F.R. The hallmarks of mitochondrial dysfunction in chronic kidney disease. Kidney Int. 2017, 92, 1051–1057. [Google Scholar] [CrossRef]
- Shurubor, Y.I.; Cooper, A.J.L.; Krasnikov, A.B.; Isakova, E.P.; Deryabina, Y.I.; Beal, M.F.; Krasnikov, B.F. Changes of Coenzyme A and Acetyl-Coenzyme A Concentrations in Rats after a Single-Dose Intraperitoneal Injection of Hepatotoxic Thioacetamide Are Not Consistent with Rapid Recovery. Int. J. Mol. Sci. 2020, 21, 8918. [Google Scholar] [CrossRef]
- Raulien, N.; Friedrich, K.; Strobel, S.; Rubner, S.; Baumann, S.; von Bergen, M.; Korner, A.; Krueger, M.; Rossol, M.; Wagner, U. Fatty Acid Oxidation Compensates for Lipopolysaccharide-Induced Warburg Effect in Glucose-Deprived Monocytes. Front. Immunol. 2017, 8, 609. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; WOldham, M.; Priolo, C.; Pandey, A.K.; Loscalzo, J. Immunometabolic Endothelial Phenotypes: Integrating Inflammation and Glucose Metabolism. Circ. Res. 2021, 129, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, I.K.N.; Tusubira, D.; Ashrafi, H.; Dyrstad, S.E.; Hansen, L.; Liu, X.Z.; Nilsson, L.I.H.; Lovsletten, N.G.; Berge, K.; Wergedahl, H.; et al. Upregulated PDK4 expression is a sensitive marker of increased fatty acid oxidation. Mitochondrion 2019, 49, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Waldecker, M.; Kautenburger, T.; Daumann, H.; Busch, C.; Schrenk, D. Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. J. Nutr. Biochem. 2008, 19, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Moffett, J.R.; Puthillathu, N.; Vengilote, R.; Jaworski, D.M.; Namboodiri, A.M. Acetate Revisited: A Key Biomolecule at the Nexus of Metabolism, Epi-genetics and Oncogenesis-Part 1: Acetyl-CoA, Acetogenesis and Acyl-CoA Short-Chain Synthetases. Front Physiol. 2020, 11, 580167. [Google Scholar] [CrossRef]
- Cheng, X.; Zhou, T.; He, Y.; Xie, Y.; Xu, Y.; Huang, W. The role and mechanism of butyrate in the prevention and treatment of diabetic kidney disease. Front. Microbiol. 2022, 13, 961536. [Google Scholar]
- Gonzalez, A.; Krieg, R.; Massey, H.D.; Carl, D.; Ghosh, S.; Gehr, T.W.B.; Ghosh, S.S. Sodium butyrate ameliorates insulin resistance and renal failure in CKD rats by modulating intestinal permeability and mucin expression. Nephrol. Dial. Transplant. 2019, 34, 783–794. [Google Scholar] [CrossRef]
- Zybailov, B.L.; Glazko, G.V.; Rahmatallah, Y.; Andreyev, D.S.; McElroy, T.; Karaduta, O.; Byrum, S.D.; Orr, L.; Tackett, A.J.; Mackintosh, S.G.; et al. Metaproteomics reveals potential mechanisms by which dietary resistant starch supplementation attenuates chronic kidney disease progression in rats. PLoS ONE 2019, 14, e0199274. [Google Scholar] [CrossRef]
- Yukino-Iwashita, M.; Nagatomo, Y.; Kawai, A.; Taruoka, A.; Yumita, Y.; Kagami, K.; Yasuda, R.; Toya, T.; Ikegami, Y.; Masaki, N.; et al. Short-Chain Fatty Acids in Gut-Heart Axis: Their Role in the Pathology of Heart Failure. J. Pers. Med. 2022, 12, 1805. [Google Scholar]
- Aguilar, E.C.; da Silva, J.F.; Navia-Pelaez, J.M.; Leonel, A.J.; Lopes, L.G.; Menezes-Garcia, Z.; Ferreira, A.V.M.; Capettini, L.; Teixeira, L.G.; Lemos, V.S.; et al. Sodium butyrate modulates adipocyte expansion, adipogenesis, and insulin receptor signaling by upregulation of PPAR-gamma in obese Apo E knockout mice. Nutrition 2018, 47, 75–82. [Google Scholar] [CrossRef]
- Amiri, P.; Hosseini, S.A.; Ghaffari, S.; Tutunchi, H.; Ghaffari, S.; Mosharkesh, E.; Asghari, S.; Roshanravan, N. Role of Butyrate, a Gut Microbiota Derived Metabolite, in cardiovascular diseases: A comprehensive narrative review. Front. Pharmacol. 2021, 12, 837509. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Zhang, Z.; Sun, Y.; Zhu, R.Y.; Wang, F.X.; Ma, L.J.; Jiang, L.; Liu, H.D. Neuroprotective Effects of Sodium Butyrate by Restoring Gut Microbiota and Inhibiting TLR4 Signaling in Mice with MPTP-Induced Parkinson’s Disease. Nutrients 2023, 15, 930. [Google Scholar] [CrossRef] [PubMed]
- Anshory, M.; Effendi, R.M.R.A.; Kalim, H.; Dwiyana, R.F.; Suwarsa, O.; Nijsten, T.E.C.; Nouwen, J.L.; Thio, H.B. Butyrate Properties in Immune-Related Diseases: Friend or Foe? Fermentation 2023, 9, 205. [Google Scholar]
- Liu, S.; Zhang, S.; Wang, Y.; Lu, S.; Han, S.; Liu, Y.; Jiang, H.; Wang, C.; Liu, H. Dietary Sodium Butyrate Improves Intestinal Health of Triploid Oncorhynchus mykiss Fed a Low Fish Meal Diet. Biology 2023, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yi, C.X.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; van den Heuvel, J.K.; Meijer, O.C.; Berbee, J.F.P.; et al. Dietary butyrate ameliorates metabolic health associated with selective proliferation of gut Lachnospiraceae bacterium 28-4. JCI Insight 2023, 8, e166655. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).