Abstract
The SPL (SQUAMOSA-promoter binding protein-like) gene family is one of the largest plant transcription factors and is known to be involved in the regulation of plant growth, development, and stress responses. The genome-wide analysis of SPL gene members in a diverse range of crops has been elucidated. However, none of the genome-wide studies on the SPL gene family have been carried out for oil palm, an important oil-yielding plant. In this research, a total of 24 EgSPL genes were identified via a genome-wide approach. Phylogenetic analysis revealed that most of the EgSPLs are closely related to the Arabidopsis and rice SPL gene members. EgSPL genes were mapped onto the only nine chromosomes of the oil palm genome. Motif analysis revealed conservation of the SBP domain and the occurrence of 1–10 motifs in EgSPL gene members. Gene duplication analysis demonstrated the tandem duplication of SPL members in the oil palm genome. Heatmap analysis indicated the significant expression of SPL genes in shoot and flower organs of oil palm plants. Among the identified EgSPL genes, a total 14 EgSPLs were shown to be targets of miR156. Real-time PCR analysis of 14 SPL genes showed that most of the EgSPL genes were more highly expressed in female and male inflorescences of oil palm plants than in vegetative tissues. Altogether, the present study revealed the significant role of EgSPL genes in inflorescence development.
1. Introduction
The African oil palm (Elaeis guineensis) belongs to the palm family (Arecaceae) and is majorly cultivated as a source for palm oil. The palm oil is obtained from fruits of oil palm plants. The male and female inflorescences are formed separately in an alternating cycle on the same plant [1,2]. The inflorescences are continuously produced at the axil of each leaf along with the vegetative growth of oil palm plants [1,2]. The development of inflorescences occurs through different phases in a period of 2 to 3 years [3,4]. The successful development of inflorescence through differential phases is the key step for the formation of oil palm fruit for better oil yield [4]. However, differential gene expression also influence the development of oil palm inflorescences/flowers [4,5]. The differential expression of genes during the flowering are regulated at post-transcriptional level by various regulatory elements, including microRNAs [4]. To achieve the good yield of palm oil, the proper development of the oil palm fruit is needed, which is the source of palm oil. Prior to the fruit formation, growth and developmental stages of flowers are also most important. Recently, researchers identified the role of SPL genes in regulating the floral organ development by interacting with downstream genes that control the length and shape of inflorescence. So, it is noteworthy to identify the SPL genes in the oil palm genome and their specific expression in inflorescence for oil palm breeding.
Plant transcription factors (TFs) play a vital role in regulating the growth and development [6]. Among the plant transcription factors, the SPL (squamosa promoter binding protein, SBP-Box) gene family is an important plant specific transcription factor family known to regulate the growth and development of plants. The SPL gene was firstly identified in the cDNA library of Antirrhinum majus inflorescences as SPL proteins bind to the SQUAMOSA promoter of MADS-Box genes [7,8]. The SPL proteins contain a highly conserved SBP domain with 70 amino acid residues, including two tandem zinc fingers (Cys-Cys-His-Cys and Cys-Cys-Cys-His) and a C-terminus region with a nuclear localization signal (NLS) [9,10]. The NLS overlaps with the zinc finger structure to direct the SBP proteins into the nucleus for transcriptional regulation of downstream genes [11]. In recent years, various genome-wide studies have identified the occurrence of SPL gene members in various plants including 16 in sweet cherry [12], 23 in alfalfa [13], 18 in foxtail millet [14], 37 in trifolium [15], 15 in jatropha [16], 17 in Arabidopsis [17,18], 19 in rice [9], 56 in wheat [19], 24 in tartary buckwheat [20], 15 in pomegranate [21], 15 in tomato [22], 28 in poplar [23], 27 in apple [24], 18 in grape [25], and 17 in the orchardgrass genome [26]. However, no genome-wide studies have been carried out for identifying the SPL genes in the oil palm genome.
MicroRNAs (miRNAs) are endogenous non-coding RNAs known to suppress the expression of target genes at the post-transcriptional level [27,28]. Among all of the miRNAs, miR156 is highly conserved in plants and regulates the expression of SPL genes for transforming the plants from vegetative to reproductive phase [29,30,31]. The SPL genes regulated by miR156 are categorized into three groups including (i) SPL2, SPL9, SPL10, SPL11, SPL13, and SPL15 (promoting juvenile-to-adult vegetative transition and vegetative-to-reproductive transition); (ii) SPL3, SPL4, and SPL5 (promoting the floral meristem identify transition); and (iii) SPL6 (function not yet known) [18]. Various studies also demonstrated the involvement of SPL genes for regulating physiological aspects related to growth and development, including leaf development, flower and fruit formation, and abiotic and biotic stress response. The leaf development is also regulated by SPL genes [32]: for example, SPL3 inhibits leaf primordia development; SPL9 and SPL10 control the leaf blade shape [18,33]. The grain size and shape in rice are regulated by SPL13 and SPL16 [34,35]. Moreover, SPL genes also play a vital role in abiotic and biotic stress response in various plants. Maize SPL genes are upregulated by cold, salt, and drought stress [36]. Downregulation of SPL8 improved drought and salt stress tolerance of transgenic alfalfa [37]. Enhanced salt tolerance of rice was also reported by knocking out the SPL10 gene in rice. Downregulation of three target genes SPL14, SPL11, SPL4 of OsmiR535 reduced the tolerance of rice to cold stress [38]. Spatiotemporal expression of alfalfa SPL genes under drought, salt stress, and biotic stress (methyl jasmonate) was also reported [13]. Previous studies also have shown that various SPL genes have miR156-binding sites [30,39]. Overexpression of miR156 in Arabidopsis downregulated the expression of SPL genes which have miR156 target sites [40]. Various researchers reported that floral organ development is regulated by SPL2 by activating the ASYMMETRIC LEAVES 2 gene of Arabidopsis thaliana [41]. The expression of LEAFY (LFY), FRUITFULL (FUL), and APETALA1 (AP1) transcription factors in floral meristems is activated by the SPL3, SPL4 and SPL5 genes [42,43].
The aim of this study is to explore the oil palm inflorescence development mechanism via the SPL/miR156 module for genetic improvement and its utilization. Our genome- wide expression-profiling analysis of the SPL gene family in oil palm will provide a fundamental platform for candidate gene selection in oil palm biotechnology programs. In this study, we identified 24 SPL genes in the oil palm genome through a bioinformatics approach with the available oil palm genome sequencing data. This is the first ample report on gene structure, conserved motif analysis, chromosomal distribution, phylogenetic analysis, and duplication events of EgSPL genes in the oil palm genome. Heat map analysis from available transcriptome data of oil palm genome revealed the significant expression of EgSPL genes in shoot and floral tissues of oil palm plants. A total of 14 EgSPL genes possess the miR156 target sites. In addition, real time PCR analysis of EgSPL genes in vegetative and reproductive tissues revealed their significant expression in male and female inflorescences tissues. The expression levels of oil palm SPL genes in inflorescence provides some information to further study the biological functions in vegetative to floral transition and inflorescence development in this important oil-yielding crop. Altogether, our study provides the involvement of SPL genes during flower development in oil palm plants.
2. Results
2.1. SPL Genes Identification in E. guineensis Genome
A total of 24 SPL genes were identified in the oil palm genome through a genome-wide approach, and they were named as EgSPL1–EgSPL24. The gene IDs for all the oil palm SPL members are provided in Supplementary Table S2. The sequence information of CDS and protein for all the oil palm SPL members are provided in Supplementary Table S3. The length of the CDS for SPL genes ranges from 537 bp to 3282 bp, and the protein sequence length varies from 178 to 1093 amino acid residues (Supplementary Table S2).
2.2. EgSPL Gene Structural Features and Conserved Motif Analysis
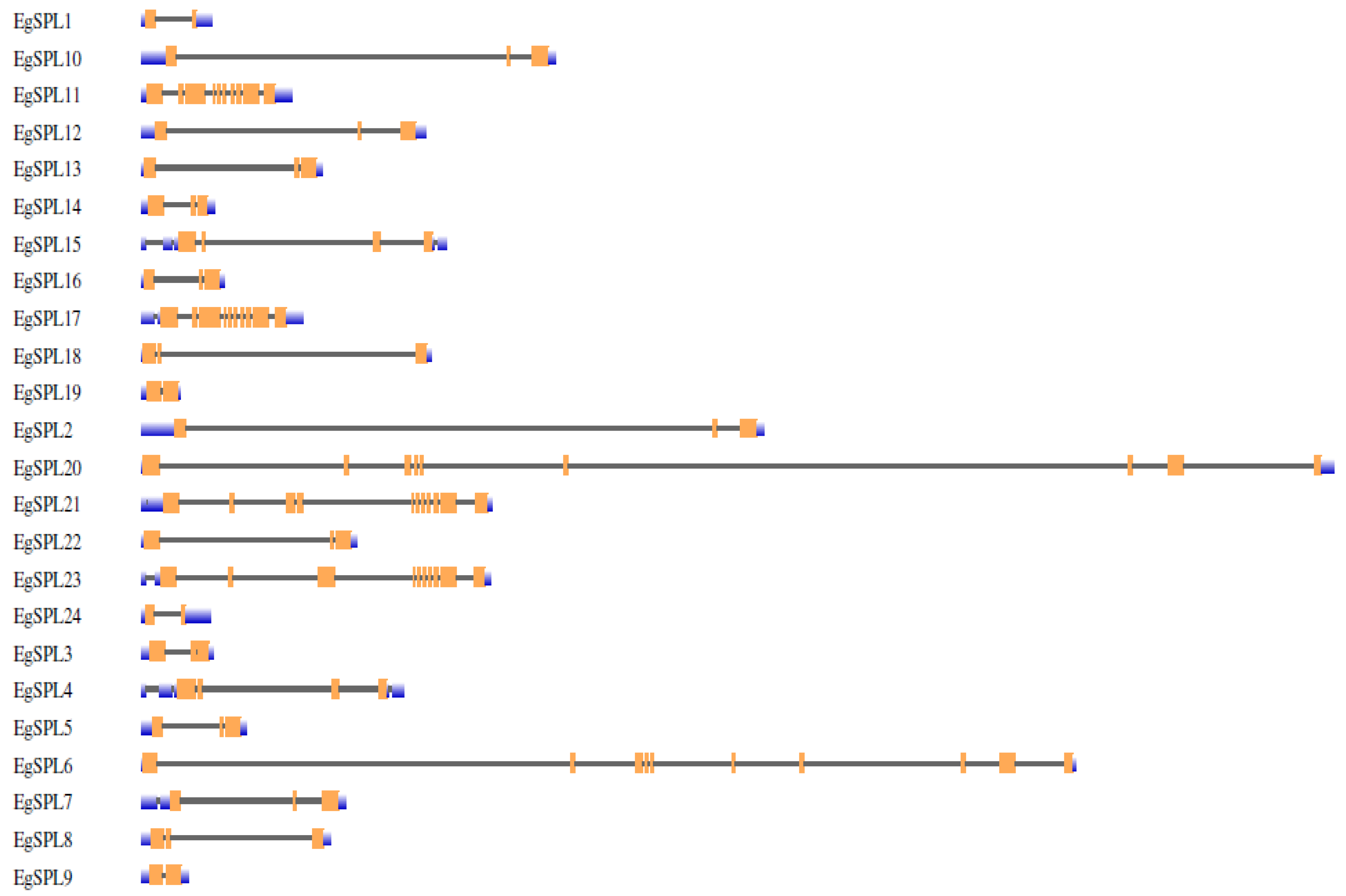
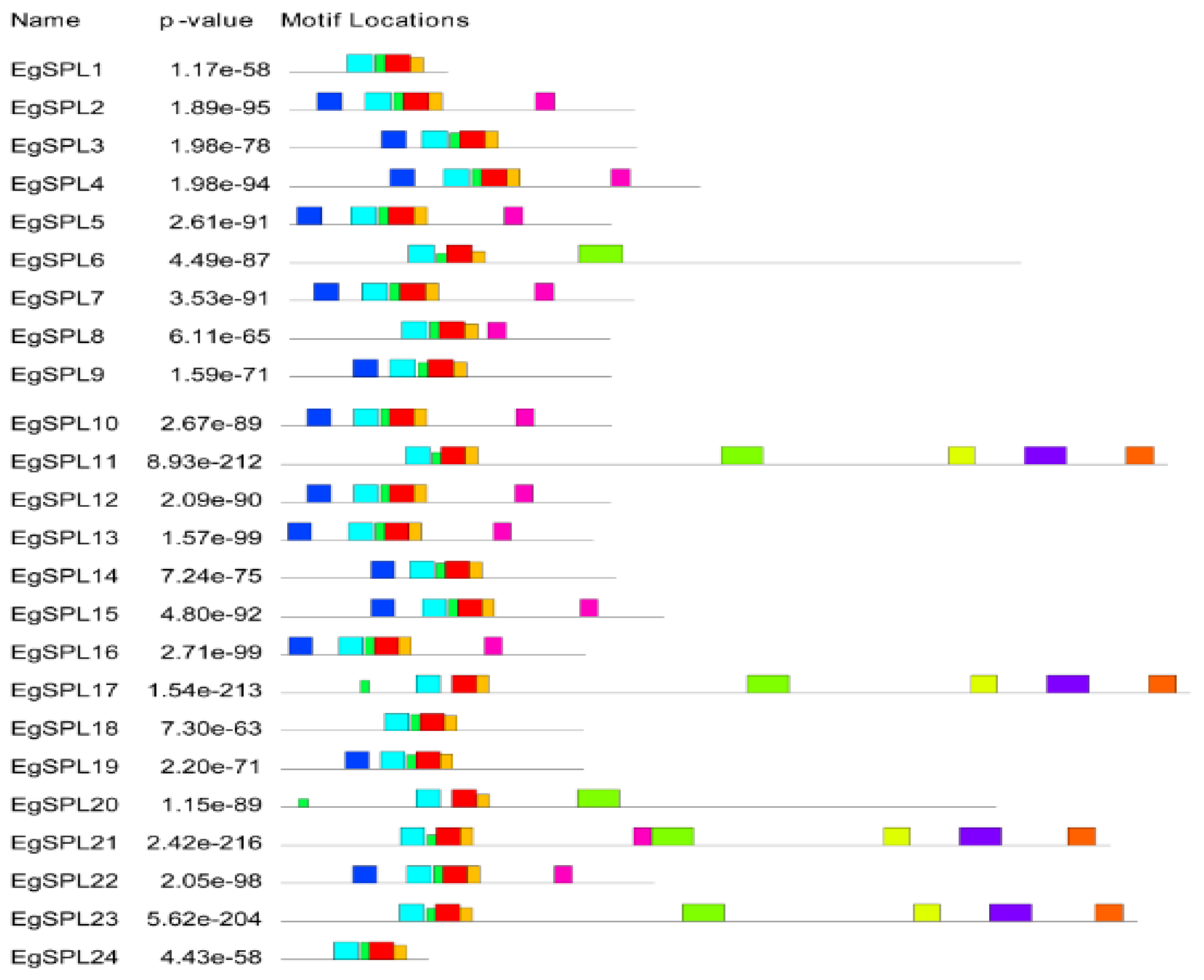
To gain further insight into the structural features of oil palm SPL genes, we used Gene Structural DiSPLay Server tool 2.0 (http://gsds.cbi.pku.edu.cn/, accessed on 1 June 2023) to analyze the exon/intron organization (Figure 1). Our analysis revealed the presence of a varied number of exons (2–12) among the identified 24 SPL genes. Highest numbers of exons (12) are possessed by EgSPL21, whereas EgSPL1, EgSPL9, EgSPL19, and EgSPL24 contained the lowest number of exons (Figure 1). Moreover, each SPL gene contained at least one intron, which varied among the SPL genes, indicating the functional role of introns in evolution. Further motif analysis revealed the existence of 10 motifs in SPL proteins which varied from 4 to 10 in each SPL protein (Figure 2). Moreover, motifs (1 and 2) related to SBP domains were found in all of the 24 oil palm SPL proteins (Figure 2). The presence of other motifs in addition to SBP domain motifs indicates the diverse functions of SPL genes.
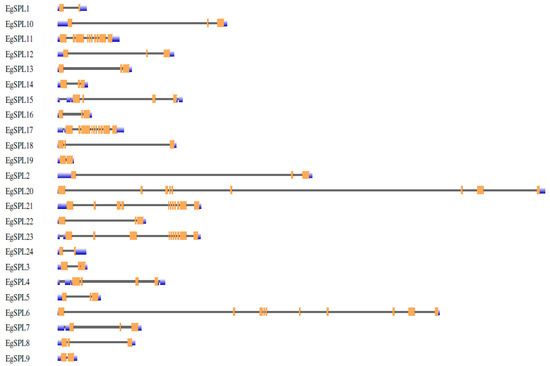
Figure 1.
Intron-exon organization of 24 EgSPL genes. Coding sequences (CDS), untranslated regions (3′ and 5′ UTRs). Intron regions are represented by orange, blue, and black color blocks, respectively, in the schematic presentation.
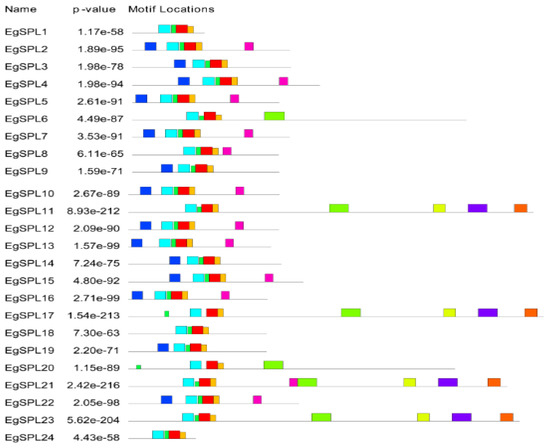

Figure 2.
Motif analysis of EgSPL proteins using the MEME tool. Presence of 1–10 motifs in EgSPL proteins. Each motif is represented with different color. The abundance of each amino acid in their motifs is represented by sequence logo of each motif.
2.3. miR156 Target Sites Prediction in Oil Palm SPL Genes
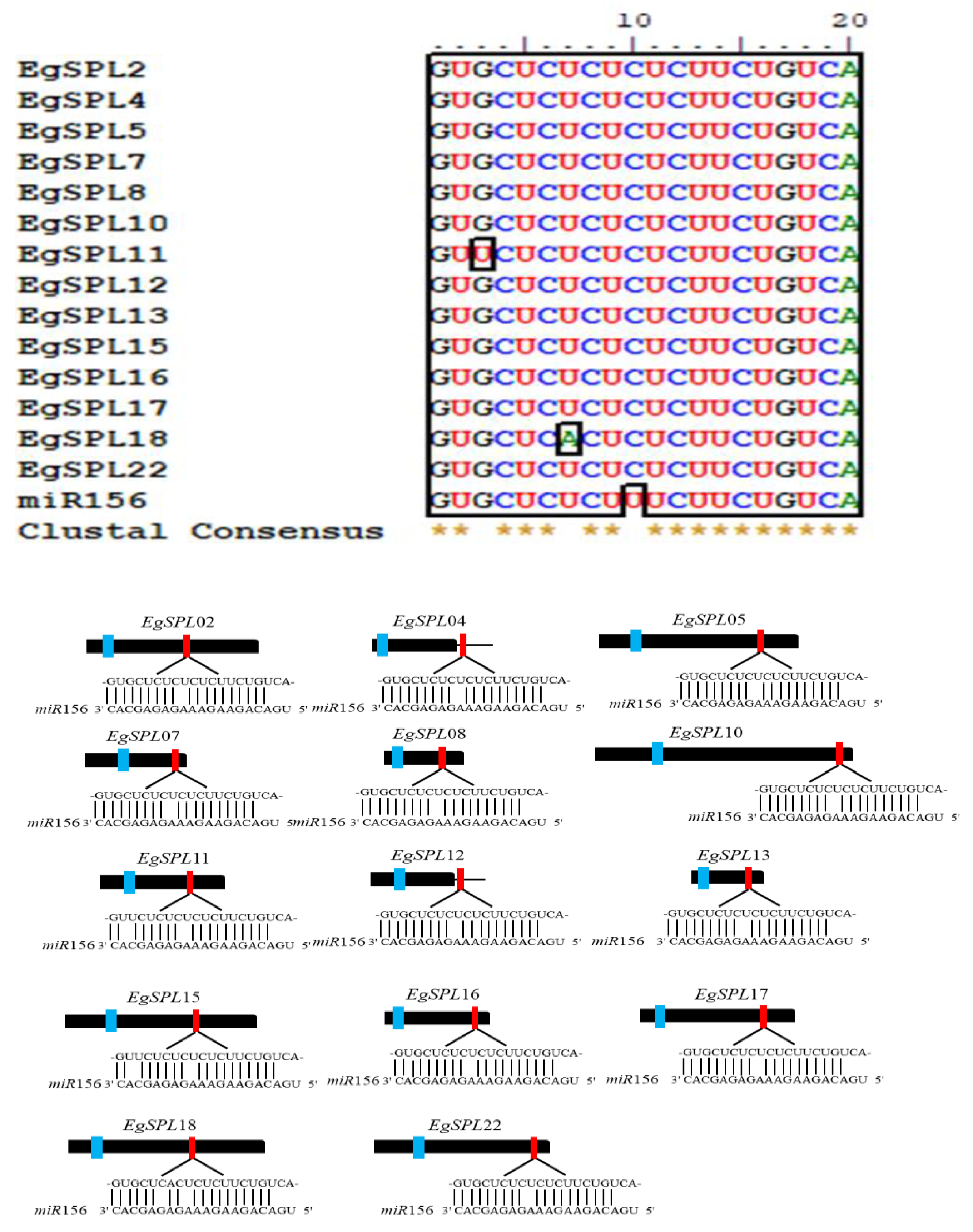
To identify the miR156 target sites in EgSPL genes, we searched the coding region and 3′-UTRs of all identified 24 SPL genes using the psRNATarget tool. We found that a total of 14 EgSPL genes (EgSPL2, EgSPL4, EgSPL5, EgSPL7, EgSPL8, EgSPL10, EgSPL11, EgSPL12, EgSPL13, EgSPL15, EgSPL 16, EgSPL17, EgSPL18, and EgSPL22) have miR156- binding sites either in coding or in 3′ UTR regions (Figure 3). The miR156-binding sites are present in the coding regions of 12 EgSPL (EgSPL2, EgSPL5, EgSPL7, EgSPL8, EgSPL10, EgSPL11, EgSPL13, EgSPL15, EgSPL16, EgSPL17, EgSPL18, and EgSPL22) members, whereas two SPLs (EgSPL4 and EgSPL12) contained miR156 sites in their 3′ UTR regions (Figure 3). Our results indicate that the regulation of EgSPL genes by miR156 is confined to a few genes among the identified 24 EgSPL genes.
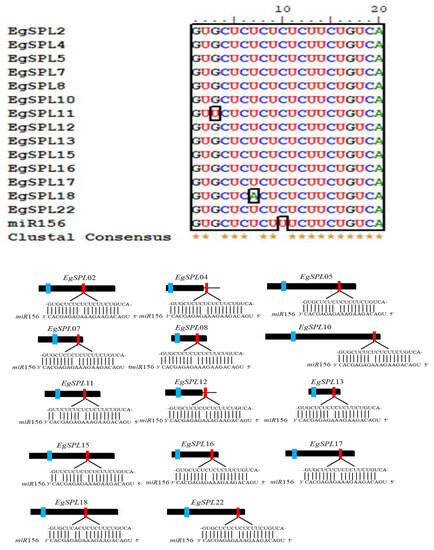
Figure 3.
Prediction of miR156 target sequences of 14 EgSPLs through psRNATarget tool. Multiple alignment of miR156 complementary sequences with their targets (upper panel). Lower panel diagram represents the gene structure of 14 EgSPLs. The black boxes represent CDS regions, the black lines represent 3′ and 5′ UTR regions, the blue boxes represent the SBP domains, and the red boxes represent miR156 target sites of EgSPL transcripts. In the expanded regions, the sequence direction of EgSPL is from 5′ to 3′ and the miR156 sequence direction is from 3′ to 5′.
2.4. Chromosomal Distribution and SPL Gene Duplication in Oil Palm Genome
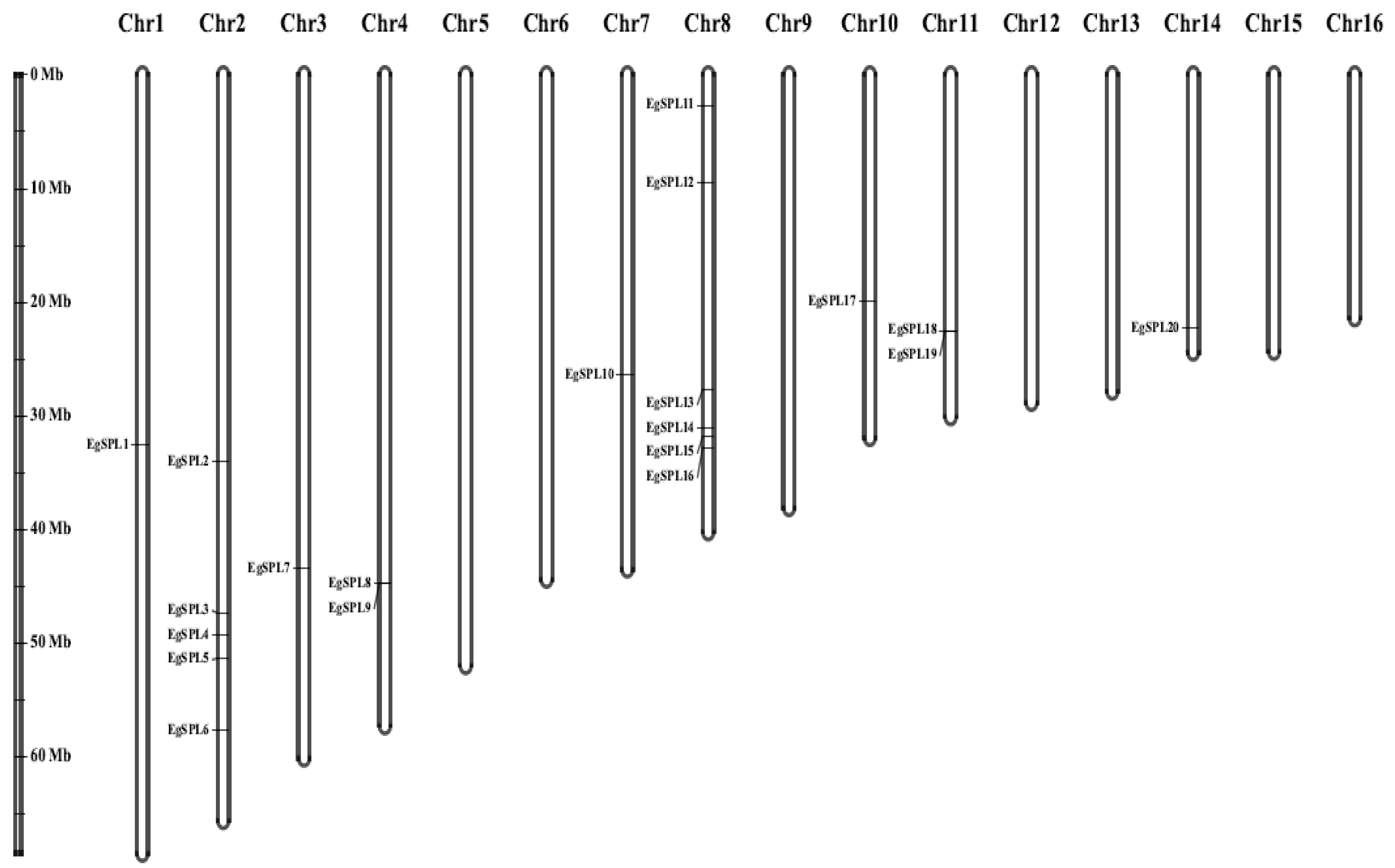
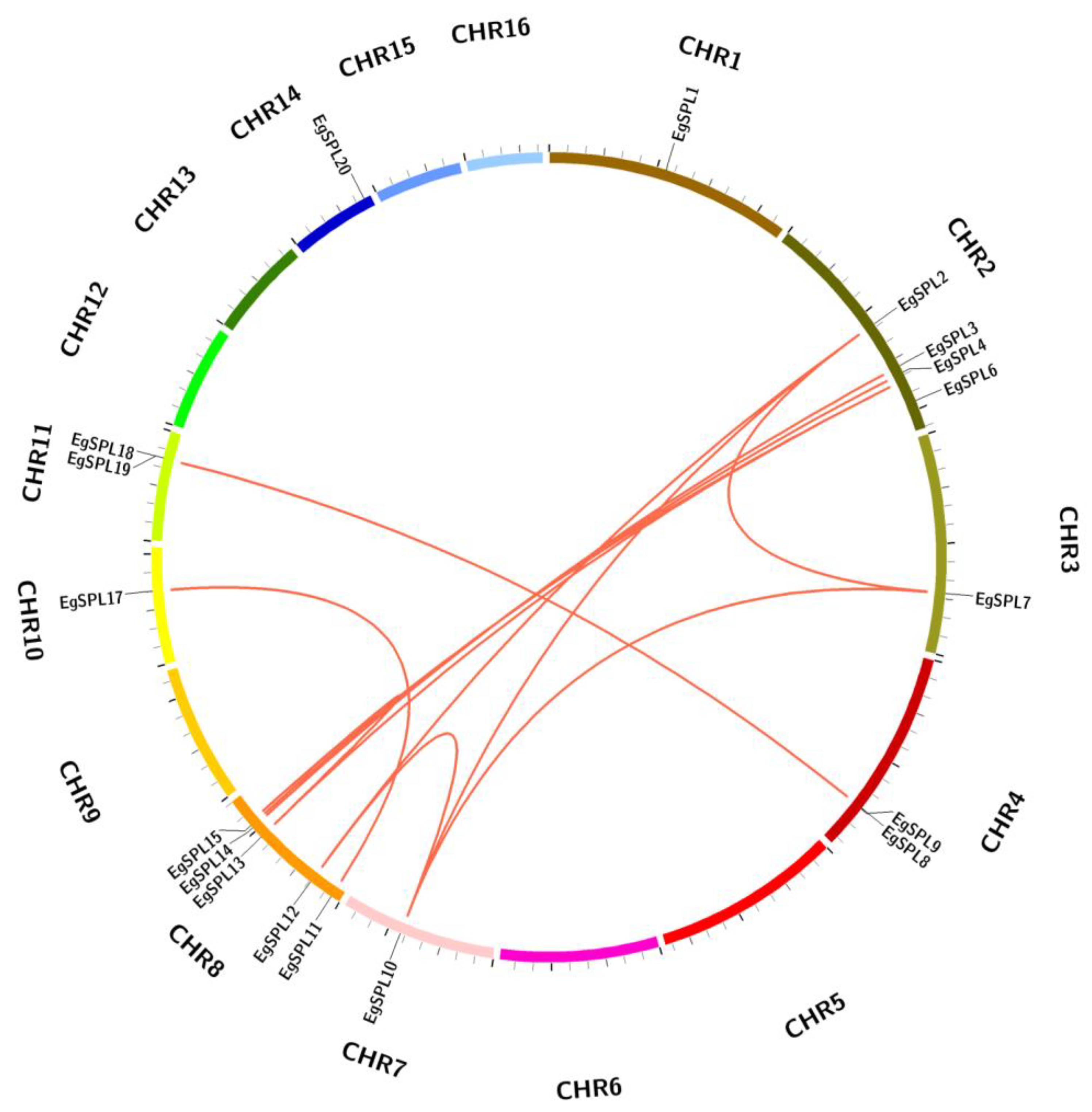
The mapping of all identified 24 EgSPL gene across 16 chromosomes of the oil palm genome was also studied. A total of 20 EgSPL members were mapped on the chromosomes (1, 2, 3, 4, 7, 8, 10, 11, and 14), and the remaining 4 SPL genes were not mapped to any of the chromosomes. We did not find any SPL genes distributed on chromosome 5, 6, 9, 12, 13, 15, and 16. Chromosome 8 and chromosome 2 contained the highest number of SPL genes (Figure 4). Chromosomes 1, 3, 7, 10, and 14 had only one SPL gene. These results suggest the uneven distribution of EgSPL genes on chromosomes of the oil palm genome (Figure 4). Further, we used the Circos algorithm to learn the information on expansion of SPL gene duplications in the oil palm genome. We found that a total of 16 pairs of SPL genes were duplicated among the 16 chromosomes of oil palm. The duplicated pairs were located on the different chromosomes of the genome. Chromosomes 2 and 8 have the largest number of duplicated pairs compared to other chromosomes (Figure 5). Most of the chromosomes did not contain any duplicated pairs of SPLs, and duplication existed only on seven chromosomes. Our findings indicate the expansion or duplication of SPLs only exists through the duplicated blocks of the genome (Figure 5).
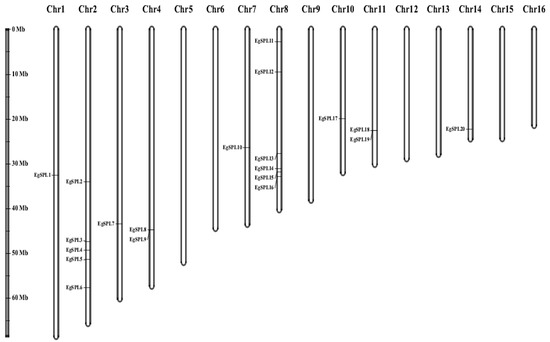
Figure 4.
Uneven distribution of EgSPL genes on 16 chromosomes of oil palm genome. Chromosome numbers from 1–16 are marked on the top portion of each chromosome. The length of the oil palm chromosomes is represented with the vertical greyscale on the left side.
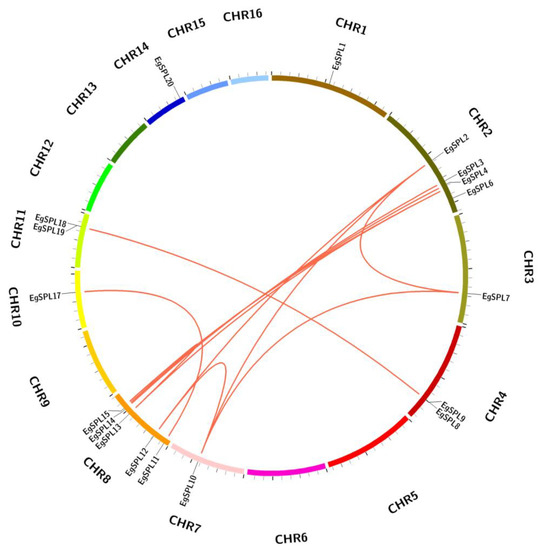
Figure 5.
EgSPL gene duplication events in oil palm genome. The duplicated gene pairs are linked by the red lines inside the circle view as revealed by MC ScanX tool. Each chromosomal block is represented by a different color.
2.5. Evolutionary Relationship of Oil Palm, Rice, and Arabidopsis SPL Genes
We constructed the Maximum Likelihood tree for analyzing the evolutionary relation between oil palm SPL genes with the SPL genes of rice and Arabidopsis. Interestingly, oil palm SPL genes are distantly related with both rice and Arabidopsis SPL members (Figure 6). However, some of them are closely present in the clades where rice SPL members are grouped. Our results demonstrate that EgSPLs were separately evolved during evolution (Figure 6).

Figure 6.
Phylogenetic analysis of EgSPL gene family with rice and Arabidopsis SPL family genes. Domain composition (middle) and gene structural organization (right) of oil palm, rice, and Arabidopsis SPL members are also represented in the illustration.
2.6. Expression Profiles of EgSPLs in Vegetative and Reproductive Tissues of Oil Palm
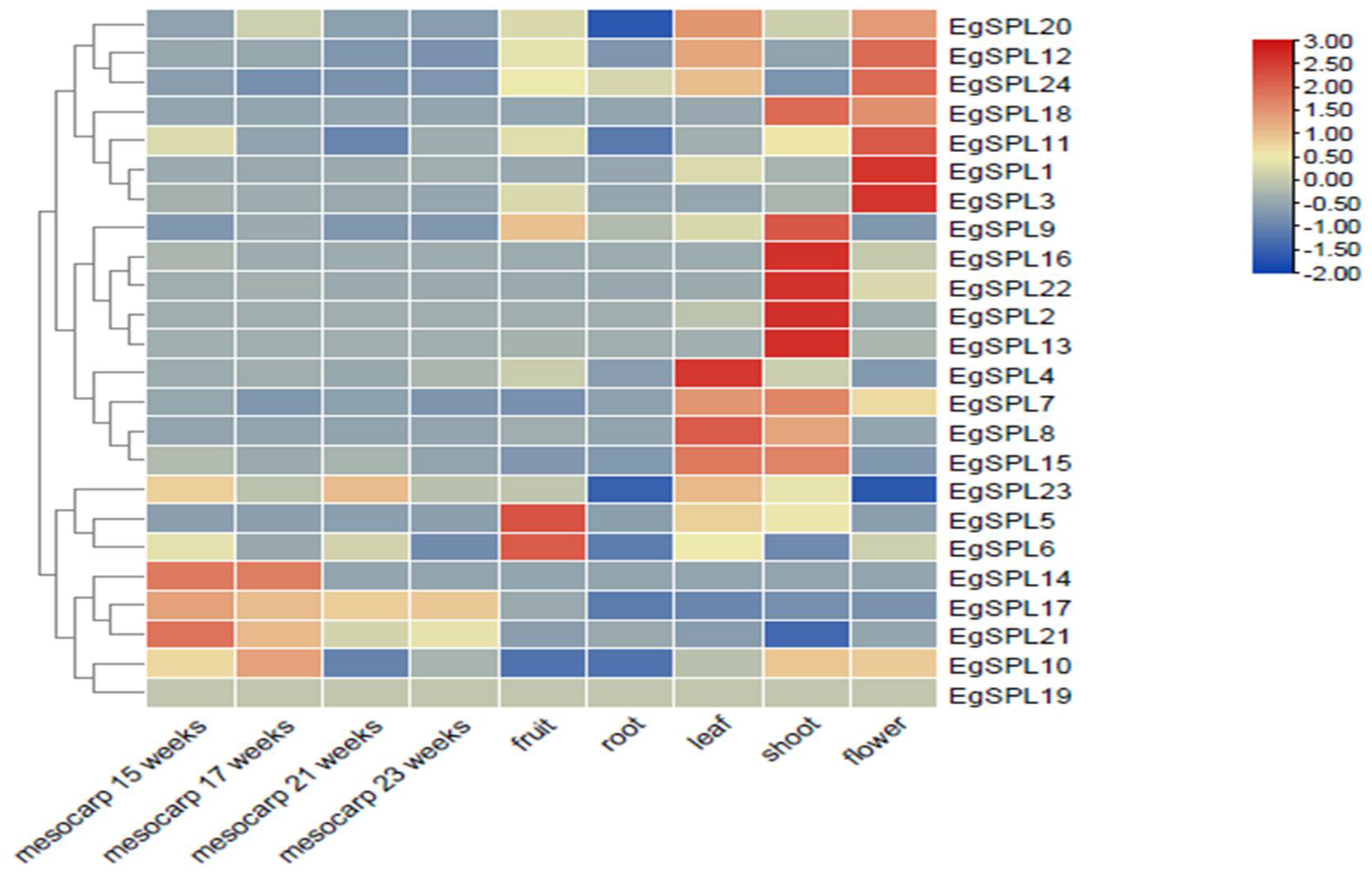
We examined the tissue-specific expression of 24 EgSPLs in vegetative (leaf, root, and shoot) and reproductive tissues (flower, fruit, and mesocarp (15, 17, 21, and 23 weeks old)) of oil palm plants using the available transcriptome data (SRR851096, SRR851071, SRR851067, SRR851108, SRR851103, SRR190698, SRR190699, SRR190701, and SRR190702). A total of two genes (EgSPL1, EgSPL3) in flower, a total of four genes (EgSPL2, EgSPL9, EgSPL13, EgSPL16, EgSPL22) in shoot, and a total of two genes (EgSPL5, EgSPL6) in fruits are highly expressed compared to other EgSPL genes (Figure 7). However, a majority of the EgSPLs were expressed in flower and shoot tissues. All the identified 24 SPL genes were downregulated in root tissues (Figure 7). Based on the expression data, we hypothesize that the oil palm SPL gene family might play important roles in oil palm plant growth development, i.e., EgSPL1 and EgSPL3 in floral meristem development; EgSPL2, EgSPL9, EgSPL13, EgSPL16, and EgSPL22 in shoot development; EgSPL5 and EgSPL6 in fruit development; EgSPL4 in leaf development.
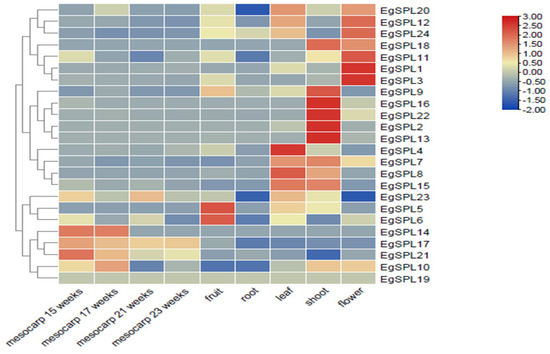
Figure 7.
Tissue-specific expression profile of EgSPL genes in vegetative (leaf, root, and shoot) and reproductive tissues (flower, mesocarp, and fruit) based on the available transcriptome data of oil palm. SRR190698 represents transcriptome of mesocarp (15 weeks); SRR190699 represents transcriptome of mesocarp (17 weeks); SRR190701 represents transcriptome of mesocarp (21 weeks); SRR190702 represents transcriptome of mesocarp (23 weeks); SRR851108 represents transcriptome of flower; SRR851067 represents transcriptome of fruit; SRR851096 represents transcriptome of leaf; SRR851071 represents transcriptome of root; SRR851103 represents transcriptome of shoot.
2.7. Real-Time Expression Analysis of EgSPLs Containing miR156-Binding Sites
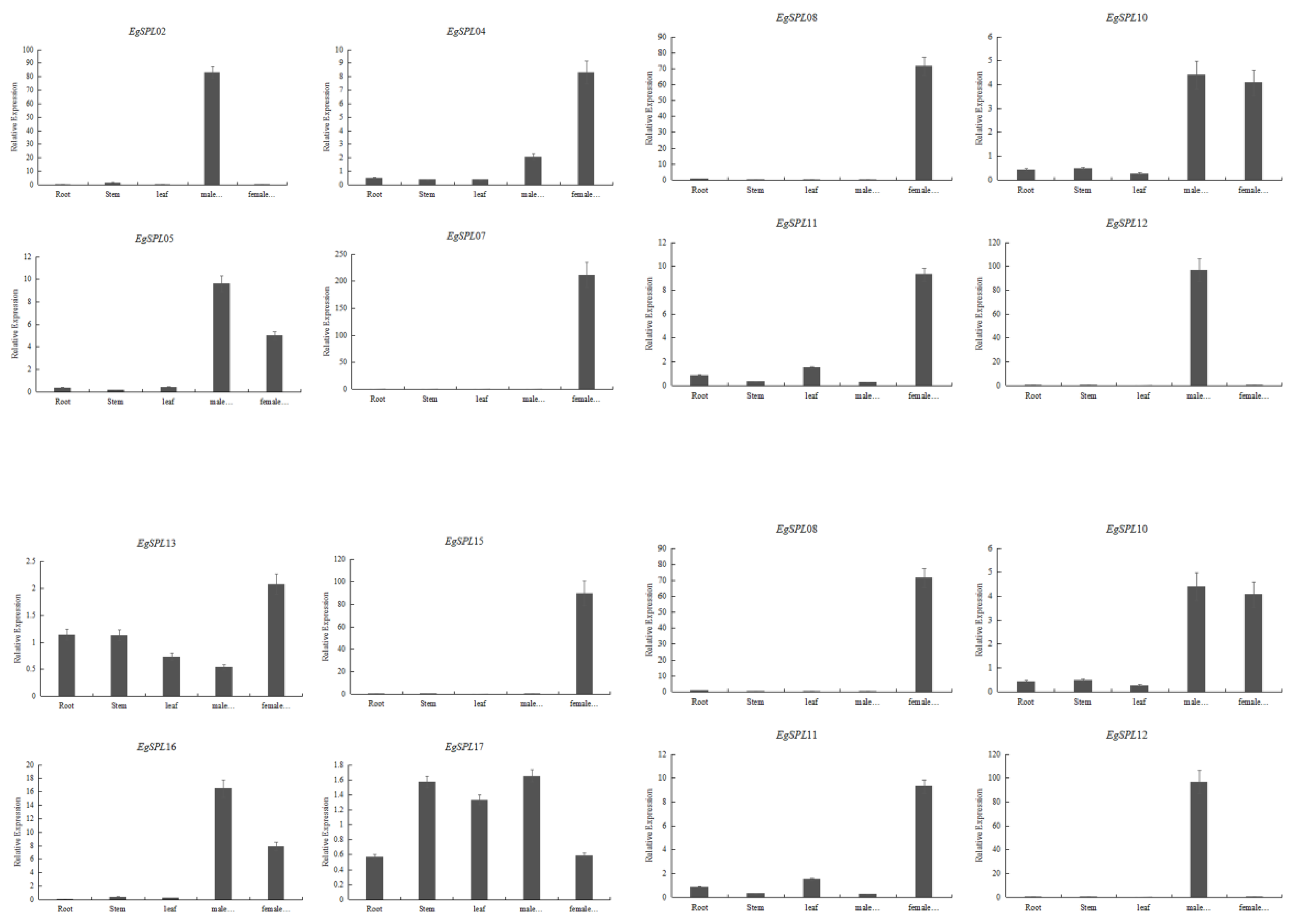
A quantitative real-time PCR experiment was performed to learn the relative expression of 14 oil palm SPL genes (containing miR156 sites) in various vegetative and reproductive tissues (male and female inflorescences) of oil palm plants. Our results demonstrate that expression of EgSPL02 and EgSPL12 is specifically confined to male inflorescences, whereas EgSPL07, EgSPL08, EgSPL18, and EgSPL22 are only confined to female inflorescences (Figure 8). A total of three genes (EgSPL05, EgSPL10, and EgSPL16) were only highly expressed in both the male and female inflorescences (Figure 8). A total of two genes, EgSPL13 and EgSPL17, were highly expressed in all the vegetative and reproductive tissues. Altogether, our results indicate the role of SPL genes in male and female inflorescence development in oil palm plants. We predict that EgSPL13 and EgSPL17 might play important roles in vegetative to reproductive phase transition, as both are expressed in vegetative and reproductive tissues of oil palm.
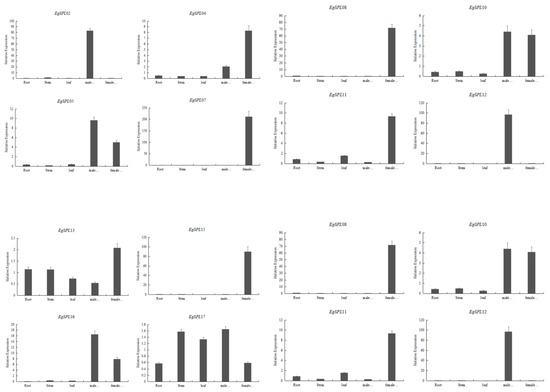
Figure 8.
Relative expression of 14 EgSPL transcription factors (containing miR156 sites) in vegetative and reproductive tissues. Data represent the mean ± SE of three replicates.
3. Discussion
SPLs are plant-specific transcription factors with a highly conserved SBP domain and are involved in regulating growth and development of plants. Various number of SPL genes were identified and characterized their expression in various crops including sweet cherry [12], alfalfa [13], quinoa [44], foxtail millet [14], wheat [45], pecan [46], soybean [47], Populas [48], and Jatropha [16]. However, some of the studies were carried out to characterize SPL gene expression during reproductive tissue development in various crops including pecan [46], Tartary buckwheat [20], rice [49], flowering cherry [50], Trifolium [15], and petunia [51]. However, to date none of the information is available on identification and characterization of SPL genes in oil palm during inflorescence development. In this study, we identified a total of 24 SPL genes in oil palm genome through a bioinformatics approach and dissected the expression of 14 SPLs in male and female inflorescences of oil palm plants through an experimental approach. The number of identified SPLs (24) in this study is fewer than the number of SPLs identified in sweet cherry (16 SPLs) [12], alfalfa (23 SPLs) [13], foxtail millet (18 SPLs) [14], and jatropha (15 SPLs) [16] and lesser than the number of SPLs identified in soybean (41 SPLs) [47], flowering cherry (32 SPLs) [50], wheat (56 SPLs) [45], Populus (33 SPLs) [48], and Trifolium (37 SPLs) [15] genomes. Present information on oil palm SPLs would elucidate the evolutionary process of SPLs across various plants.
The structural organization across the identified oil palm SPL genes showed that the variation in number of introns indicates the role of introns in SPL gene evolution. All the identified SPLs contained at least one intron and varied among them. Our results are consistent with the previous reports on identification of SPL genes in Jatropha [16], soybean [47], alfalfa [13], and sweet cherry [12]. The presence of the SBP domain is the key feature in the SPL gene family [9]. The motifs related to the SBP domain are found in all the identified EgSPL proteins. Conserved motif analysis revealed the occurrence of 10 motifs in oil palm SPL genes as reported in other plants including foxtail millet [14], alfalfa [13], and quinoa [44]. The post-transcriptional regulation of SPLs by miR156 determines fine-tuning functions of SPLs [52]. In our study, a total of 14 oil palm SPLs contained the miR156 sites, mostly in the coding regions and lesser in UTR regions. Our results are consistent with the previous reports on Jatropha, soybean, Medicago, and mustard SPL genes [16,47,53,54] indicating the conservation of miR156-mediated posttranscriptional regulation in plants. Phylogenetic analysis of 24 oil palm SPLs compared with Arabidopsis and rice SPLs suggested the diversification of oil palm SPL members among model-to-crop plants during the evolutionary process. The uneven distribution of 24 SPL genes on 16 chromosomes of oil palm also coincides with the previous reports on SPL gene family distribution in sweet cherry, alfalfa, and foxtail millet genomes [12,13,14].
SPL genes encode plant-specific transcription factors that play important roles in flower development, including vegetative to reproductive growth [55]. In our study, we analyzed the tissue specific expression of EgSPLs from the available transcriptome data and the majority of EgSPL genes expressed in flower and shoot. Our results are consistent with the previous reports on expression of SPL genes in blue berry [56]. Our qPCR data on expression of 14 SPL genes that contain miR156 sites revealed confined expression of some SPL genes in male and female inflorescences. The expression of miR156-targeted EgSPL02 and EgSPL12 was unique to male inflorescences, miR156-targeted EgSPL07, EgSPL08, EgSPL18, and EgSPL22 were unique to female inflorescences, and miR156-targeted EgSPL05, EgSPL10, and EgSPL16 were only highly expressed in both the male and female inflorescences, suggesting the involvement of EgSPL genes in inflorescence development. Our results are consistent with the previously reported involvement of miR156-targeted SPL genes in inflorescences [47,51].
4. Materials and Methods
4.1. Identification of SPL Gene Family in Oil Palm Genome
The known SPL protein sequences of Arabidopsis (16) and rice (19) were used as queries to retrieve the putative oil palm SPL family members against the oil palm (E. guineensis) genome database (http://palmxplore.mpob.gov.my/palmXplore/, accessed on 1 June 2023). Then, all the retrieved putative oil palm SPL family proteins were queried against the CDD (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 1 June 2023) and Pfam databases to confirm the occurrence of the conserved SBP domain. The length of each putative SPL gene coding sequence was also determined via Blastn search against the E. guineensis genome database.
4.2. Oil Palm SPL Gene Structure, Conserved Motif Analysis, and miR156 Target Site Prediction
The intron/exon structure analysis of 24 oil palm SPL gene structures was analyzed using Gene Structure DiSPLay Server (http://gsds.cbi.pku.edu.cn/, accessed on 1 June 2023). Further, the MEME tool was used for conserved motif analysis of all identified oil palm SPL protein members (http://meme-suite.org/tools/meme, accessed on 1 June 2023). The psRNATarget tool (http://plantgrn.noble.org/psRNATarget/?function=3, accessed on 1 June 2023) was used to predict the miR156 target site in oil palm full-length SPL gene sequences.
4.3. Phylogenetic Analysis, Duplication, and Chromosomal Distribution of Oil Palm SPL Genes
We generated the phylogenetic tree of oil palm SPL genes with the other known SPL genes of Arabidopsis and rice with the help of MEGA 7.0 tool by the Maximum Likelihood method, with a bootstrap value of 1000 replications [57]. We also investigated SPL gene members’ duplication events across the oil palm genome via the MCScanX tool with default parameters [58]. Further, we also mapped the distribution of 24 oil palm SPL genes on 16 chromosomes of oil palm from the oil palm genome database using TB tools software [59].
4.4. Plant Material and RNA Isolation
Healthy African oil palm (Elaeis guineensis) plants were grown in the field station of the Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences, Wenchang, China, and were all grown under institutional regulatory issues. The plant material used in this research work was collected by the corresponding author. To investigate the SPL gene expression analysis in different developmental stages, the samples including root, stem, leaf, male inflorescence, and female inflorescence were collected separately from the 6-year-old oil palm plants and quickly frozen in liquid nitrogen and stored at −80 °C for further experiments. Total RNA from root, stem, leaf, male inflorescence, and female inflorescence was isolated by following the method as described previously [60]. The yield, integrity, and purity of extracted RNA samples were quantified on Nanodrop and also electrophoresed on 1% agarose gel. The genomic DNA contamination was removed by treating the RNA samples with DNase I.
4.5. In Silico Expression Analysis of Oil Palm SPL Genes
To examine the SPL gene expression in various tissues of oil palm, the normalized RPKM values were retrieved from the available transcriptome data of six different tissues including leaf, root, flower, shoot, and mesocarp (15, 17, 21, and 23 weeks). The heatmap was generated to analyze the SPL gene expression in various tissues of oil palm using the pheatmap tool (https://cran.r-project.org/web/packages/pheatmap/index.html, accessed on 1 June 2023).
4.6. qRT-PCR Analysis of SPL Genes in Vegetative and Reproductive Tissues
To investigate the real-time expression of SPL genes in various vegetative and reproductive tissues, the real-time PCR expression analysis was performed using Mastercycler ep realplex4 Machine. The cDNA synthesis was carried out with the extracted RNA by using a MightyScript first-strand cDNA synthesis kit following the manufacturer’s instructions. The 2 × SYBR Green qPCR ProMix was used to carry out qRT-PCR reactions with Mastercycler. The oil palm SPL gene-specific primers (Supplementary Table S1) were designed by using the QuantPrime qPCR primer designing tool (https://quantprime.mpimp-golm.mpg.de/, accessed on 1 June 2023). Three biological and technical repeats were performed to determine the reliability of gene expression studies. The oil palm Actin1 gene was used as a housekeeping gene to check the relative expression of SPLs by employing the 2−ΔΔCt method. The statistical significance was determined at p < 0.05 and p < 0.01 using ANOVA.
5. Conclusions
To the best of our knowledge, this is the first report on genome-wide analysis of SPL genes in oil palm. In the current investigation, a total of 24 EgSPLs were identified, and the expression of 14 EgSPL (containing miR156 sites) genes in vegetative and reproductive tissues of oil palm was analyzed. Moreover, detailed information on SPL gene structure, their miR156 target sites, motif composition, chromosomal location, and phylogenetic analysis was also reported. Furthermore, the unique expression of EgSPLs (containing miR156 sites) in oil palm inflorescences was also revealed via qPCR analysis, predicting their putative role in male and female inflorescence development of oil palm.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241713658/s1.
Author Contributions
L.Z. conceived and designed the study. L.Z. and R.Y. performed the bioinformatics analysis. L.Z. and R.Y. analyzed the data. R.Y. drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research work is financially supported by Hainan Provincial Natural Science Foundation of China (323MS073).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data is available upon requesting the corresponding author.
Acknowledgments
We thank Rui Li for his help throughout this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adam, H.; Collin, M.; Richaud, F.; Beulé, T.; Cros, D.; Omoré, A.; Nodichao, L.; Nouy, B.; Tregear, J.W. Environmental regulation of sex determination in oil palm: Current knowledge and insights from other species. Ann. Bot. 2011, 108, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Adam, H.; Jouannic, S.; Escoute, J.; Duval, Y.; Verdeil, J.L.; Tregear, J.W. Reproductive developmental complexity in the African oil palm (Elaeis guineensis, Arecaceae). Am. J. Bot. 2005, 92, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Corley, R.H.V.; Gray, B.S. Growth and morphology. In Developments in Crop; Corley, R.H.V., Hardon, J.J., Wood, B.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1976. [Google Scholar]
- Ho, H.; Gudimella, R.; Ong-Abdullah, M. Expression of microRNAs during female inflorescence development in African oil palm (Elaeis uineensis Jacq.). Tree Genet. Genomes 2017, 13, 35. [Google Scholar] [CrossRef]
- Beulé, T.; Camps, C.; Debiesse, S.; Tranchant, C.; Dussert, S.; Sabau, X.; Jaligot, E.; Syed Alwee, S.S.R.; Tregear, J.W. Transcriptome analysis reveals differentially expressed genes associated with the mantled homeotic flowering abnormality in oil palm (Elaeis guineensis). Tree Genet. Genomes 2011, 7, 169–182. [Google Scholar] [CrossRef]
- Gonzalez, D.H. Introduction to transcription factor structure and function. In Plant Transcription Factors; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–11. [Google Scholar]
- Huijser, P.; Klein, J.; Lönnig, W.E.; Meijer, H.; Saedler, H.; Sommer, H. Bracteomania, an inflorescence anomaly, is caused by the loss of function of the MADS-box gene squamosa in Antirrhinum majus. EMBO J. 1992, 11, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.P. A new family of DNA binding proteins includes putative transcriptional regulators of the ANTIRRHINUM MAJUS floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. MGG 1996, 250, 7–16. [Google Scholar]
- Yang, Z.; Wang, X.; Gu, S.; Hu, Z.; Xu, H.; Xu, C. Comparative study of SBP-box gene family in Arabidopsis and rice. Gene 2008, 407, 1–11. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Jach, G.; Saedler, H.; Huijser, P. Functional dissection of the plant-specific SBP-domain: Overlap of the DNA-binding and nuclear localization domains. J. Mol. Biol. 2005, 352, 585–596. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Xiao, Y.; Zhang, X.; Du, B.; Turupu, M.; Wang, C.; Yao, Q.; Gai, S.; Huang, J.; et al. Genome-Wide Identification of the SQUAMOSA PROMOTER-BINDING PROTEIN-LIKE (SPL) Transcription Factor Family in Sweet Cherry Fruit. Int. J. Mol. Sci. 2023, 24, 2880. [Google Scholar] [CrossRef]
- Wang, Y.; Ruan, Q.; Zhu, X.; Wang, B.; Wei, B.; Wei, X. Identification of Alfalfa SPL gene family and expression analysis under biotic and abiotic stresses. Sci. Rep. 2023, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Fan, Y.; Xue, G.; He, A.; Yang, H.; He, C.; Li, Y.; Ruan, J.; Yan, J.; Cheng, J. Genome-wide identification and characterization of the SPL gene family and its expression in the various developmental stages and stress conditions in foxtail millet (Setaria italica). BMC Genom. 2022, 23, 389. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Nie, G.; Yang, Z.; Ma, S.; Fan, J.; Hu, R.; Wu, F.; Zhang, X. Genome-Wide Identification, Characterization, and Expression Profiling Analysis of SPL Gene Family during the Inflorescence Development in Trifolium repens. Genes 2022, 13, 900. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Yang, J.C.; Yin, G.T.; Li, R.S.; Zou, W.T. Genome-wide characterization of the SPL gene family involved in the age development of Jatropha curcas. BMC Genom. 2020, 21, 368. [Google Scholar] [CrossRef]
- Cardon, G.; Hohmann, S.; Klein, J.; Nettesheim, K.; Saedler, H.; Huijser, P. Molecular characterisation of the Arabidopsis SBP-box genes. Gene 1999, 237, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hu, T.; Zhao, J.; Park, M.Y.; Earley, K.W.; Wu, G.; Yang, L.; Poethig, R.S. Developmental functions of miR156-regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006263. [Google Scholar] [CrossRef]
- Zhu, T.; Liu, Y.; Ma, L.; Wang, X.Y.; Zhang, D.Z.; Han, Y.C.; Ding, Q.; Ma, L.J. Genome-wide identification, phylogeny and expression analysis of the SPL gene family in wheat. BMC Plant Biol. 2020, 20, 420. [Google Scholar] [CrossRef]
- Liu, M.; Sun, W.; Ma, Z.; Huang, L.; Wu, Q.; Tang, Z.Z.; Bu, T.L.; Li, C.L.; Chen, H. Genome-wide identification of the SPL gene family in Tartary Buckwheat (Fagopyrum tataricum) and expression analysis during fruit development stages. BMC Plant Biol. 2019, 19, 299. [Google Scholar] [CrossRef]
- Li, B.; Zhao, Y.; Wang, S.; Zhang, X.; Wang, Y.; Shen, Y.; Yuan, Z. Genome-wide identification, gene cloning, subcellular location and expression analysis of SPL gene family in P. granatum L. BMC Plant Biol. 2021, 21, 400. [Google Scholar] [CrossRef]
- Salinas, M.; Xing, S.; Höhmann, S.; Berndtgen, R.; Huijser, P. Genomic organization, phylogenetic comparison and differential expression of the SBP-box family of transcription factors in tomato. Planta 2012, 235, 1171–1184. [Google Scholar] [CrossRef]
- Li, C.; Lu, S. Molecular characterization of the SPL gene family in Populus trichocarpa. BMC Plant Biol. 2014, 14, 131. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, H.; Li, X.; Xiang, J.; Yin, X.; Gao, H.; Zheng, Y.; Bassett, C.L.; Wang, X.P. Genome-wide identification and analysis of the SBP-box family genes in apple (Malus × domestica Borkh.). Plant Physiol. Biochem. 2013, 70, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Li, J.; Gao, M.; Singer, S.; Wang, H.; Mao, L. Genomic organization, phylogenetic comparison and differential expression of the SBP-box family genes in grape. PLoS ONE 2013, 8, e59358. [Google Scholar] [CrossRef]
- Feng, G.; Han, J.; Yang, Z.; Liu, Q.; Shuai, Y.; Xu, X.; Nie, G.; Huang, L.; Liu, W.; Zhang, X. Genome-wide identification, phylogenetic analysis, and expression analysis of the SPL gene family in orchardgrass (Dactylis glomerata L.). Genomics 2021, 113, 2413–2425. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Breakfield, N.W.; Corcoran, D.L.; Petricka, J.J.; Shen, J.; Sae-Seaw, J.; Rubio-Somoza, I. High-resolution experimental and computational profiling of tissue specific known and novel miRNAs in Arabidopsis. Genome Res. 2012, 22, 163–176. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, W.; Wang, X.; Yang, R.; Wu, Z.; Wang, H. MiR156 regulates anthocyanin biosynthesis through SPL targets and other microRNAs in poplar. Horticult Res. 2020, 7, 6786–6787. [Google Scholar] [CrossRef]
- Ye, B.; Zhang, K.; Wang, J. The role of miR156 in rejuvenation in Arabidopsis thaliana. J. Integr. Plant Biol. 2020, 62, 550–555. [Google Scholar] [CrossRef]
- Teotia, S.; Tang, G. To bloom or not to bloom: Role of microRNAs in plant flowering. Mol. Plant 2015, 8, 359–377. [Google Scholar] [CrossRef]
- Usami, T.; Horiguchi, G.; Yano, S.; Tsukaya, H. The more and smaller cells mutants of Arabidopsis thaliana identify novel roles for squamosa promoter binding protein-like genes in the control of heteroblasty. Development 2009, 136, 955–964. [Google Scholar] [CrossRef]
- Martin, R.C.; Asahina, M.; Liu, P.P.; Kristof, J.R.; Coppersmith, J.L.; Pluskota, W.E.; Bassel, G.W.; Goloviznina, N.A.; Nguyen, T.T.; Martínez-Andújar, C.; et al. The microRNA156 and microRNA172 gene regulation cascades at post-germinative stages in Arabidopsis. Seed Sci. Res. 2010, 20, 79–87. [Google Scholar] [CrossRef]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950. [Google Scholar] [CrossRef]
- Mao, H.D.; Yu, L.J.; Li, Z.J.; Yan, Y.; Han, R.; Liu, H.; Ma, M. Genome-wide analysis of the SPL family transcription factors and their responses to abiotic stresses in maize. Plant Gene 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Gou, J.; Debnath, S.; Sun, L.; Flanagan, A.; Tang, Y.; Jiang, Q.; Wen, J.; Wang, Z. From model to crop: Functional characterization of SPL8 in M. truncatula led to genetic improvement of biomass yield and abiotic stress tolerance in alfalfa. Plant Biotech. J. 2018, 16, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Shen, Y.; Yang, J.; Cai, X.; Li, H.; Zhu, Y.; Jia, B.; Sun, X. miR535 negatively regulates cold tolerance in rice. Mol. Breed. 2020, 40, 14. [Google Scholar] [CrossRef]
- Munusamy, P.; Zolotarov, Y.; Meteignier, L.V.; Moffett, P.; Strömvik, M.V. Novo computational identification of stress-related sequencemotifs and microRNA target sites in untranslated regions of a plant translatome. Sci. Rep. 2017, 7, 43861. [Google Scholar] [CrossRef]
- Schwab, R.; Palatnik, J.F.; Riester, M.; Schommer, C.; Schmid, M.; Weigel, D. Specific effects of microRNAs on the plant transcriptome. Dev. Cell 2005, 8, 517–527. [Google Scholar] [CrossRef]
- Wang, Z.S.; Wang, Y.; Kohalmi, S.E.; Amyot, L.; Hannoufa, A. SQUAMOSA promoter binding PROTEIN-LIKE 2 controls floral organ development and plant fertility by activating ASYMMETRIC LEAVES 2 in Arabidopsis thaliana. Plant Mol. Biol. 2016, 9, 661–674. [Google Scholar] [CrossRef]
- Kim, J.J.; Lee, J.H.; Kim, W.; Jung, H.S.; Huijser, P.; Ahn, J.H. The miR156-SPL3 module regulates ambient temperature-responsive flowering via FT in Arabidopsis thaliana. Plant Physiol. 2012, 159, 461–478. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, P.J.; Kang, S.K.; Park, C.M. miR172 signals are incorporated into the miR156 signaling pathway at the SPL3/4/5 genes in Arabidopsis developmental transitions. Plant Mol. Biol. 2011, 76, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ma, R.; Fan, Y.; Zhao, B.; Cheng, P.; Fan, Y.; Wang, B. Genome-wide identification and expression analysis of the SPL transcription factor family and its response to abiotic stress in Quinoa (Chenopodium quinoa). BMC Genom. 2022, 23, 773. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Lu, Q.; Cang, J. Genome-wide identification and expression profiling of the SPL family genes in wheat. Botany 2021, 99, 185–198. [Google Scholar] [CrossRef]
- Wang, M.; Mo, Z.; Lin, R.; Zhu, C. Characterization and expression analysis of the SPL gene family during floral development and abiotic stress in pecan (Carya illinoinensis). PeerJ 2021, 9, e12490. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.K.; Goel, R.; Kumari, S.; Dahuja, A. Genomic organization, phylogenetic comparison, and expression profiles of the SPL family genes and their regulation in soybean. Dev. Genes. Evol. 2017, 227, 101–119. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Bao, L.H.; He, Z.G.; Li, C.L.; La, H.G.; Zhao, L.F. Identification and regulatory network analysis of SPL family transcription factors in Populus Euphratica Oliv. Heteromorphic Leaves. Sci. Rep. 2022, 12, 2856. [Google Scholar] [CrossRef] [PubMed]
- Giaume, F.; Fornara, F. SPL transcription factors prevent inflorescence reversion in rice. Mol. Plant 2021, 14, 1041–1043. [Google Scholar] [CrossRef]
- Gao, L.; Lyu, T.; Lyu, Y. Genome-Wide Analysis of the SPLGene Family and Expression Analysisduring Flowering Induction in Prunus yedoensis ‘Somei-yoshino’. Int. J. Mol. Sci. 2022, 23, 10052. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, S.; Chen, F.; Liu, B.; Wu, L.; Li, F.; Zhang, J.; Bao, M.; Liu, G. Genome-wide identification and characterization of the SBP-box gene family in Petunia. BMC Genom. 2018, 19, 193. [Google Scholar] [CrossRef]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Z.; Xu, Y.; Kong, L.C.; Shi, J.J.; Liu, Y.F.; Fu, C.X.; Wang, X.S.; Wang, Z.Y.; Zhou, C.E.; et al. Genome-wide characterization of SPL family in Medicago truncatula reveals the novel roles of miR156/SPL module in spiky pod development. BMC Genom. 2019, 20, 552. [Google Scholar] [CrossRef]
- Gao, J.; Peng, H.; Chen, F.; Liu, Y.; Chen, B.; Li, W. Genome-wide identification and characterization, phylogenetic comparison and expression profiles of SPL transcription factor family in B. juncea (Cruciferae). PLoS ONE 2019, 14, e0224704. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.; Liu, D.; Zhang, K.; Li, A.; Mao, L. SQUAMOSA promoter-binding protein-like transcription factors: Star players for plant growth and development. J. Integr. Plant Biol. 2010, 52, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhou, B.J.; Wu, X.L.; Wu, H.L.; Zhang, S.L.; Jiang, Y.; Wang, Y.P.; Zhang, Y.Q.; Cao, M.; Guo, B.S.; et al. Molecular characterization of SPL gene family during flower morphogenesis and regulation in blueberry. BMC Plant Biol. 2023, 23, 40. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucl. Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018. bioRxiv:10.1101.289660. [Google Scholar] [CrossRef]
- Badai, S.S.; Rasid, O.A.; Parveez, G.K.A.; Masani, M.Y.A. A rapid RNA extraction method from oil palm tissues suitable for reverse transcription quantitative real-time PCR (RT-qPCR). 3 Biotech 2020, 10, 530. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).