
Production of Recombinant Zika Virus Envelope Protein by Airlift Bioreactor as a New Subunit Vaccine Platform
 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Silico Analyses of Biochemical Properties of ZIKV Recombinant Envelope Protein (rEZIKV)
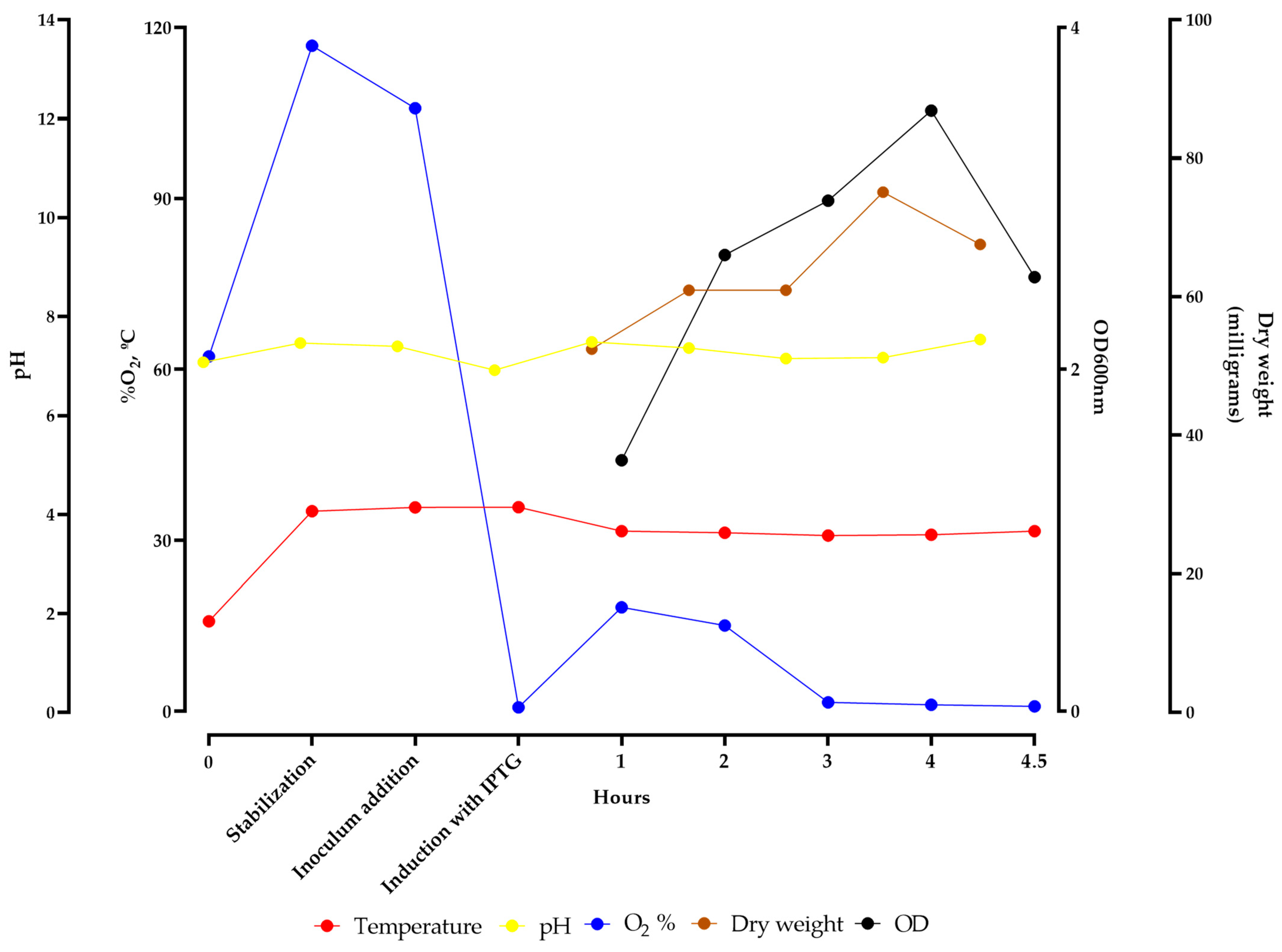
2.2. Expression and Purification of rEZIKV
2.3. Humoral Immune Response Evaluation after rEZIKV Immunization in Mice
2.4. Cross-Reactivity Evaluation between rEZIKV and OMV+ZIKV Sera and DENV Serotypes
2.5. rEZIKV Antigenic Potential Evaluation by ELISA
2.6. mRNA Cytokine Expression Profile Evaluation by RT-qPCR
3. Discussion
4. Materials and Methods
4.1. Bacterial Cells, Plasmids, ZIKV, and DENV Inactivated Particles
4.2. Ethics Statements
4.3. Sequence and Structure Analysis
4.4. Large-Scale Expression of ZIKV Recombinant Envelope Protein (rEZIKV) in an Airlift Bioreactor
4.5. Recombinant Protein Purification
4.6. SDS-PAGE and Western Blot for Purity Analysis of rEZIKV
4.7. BALB/c Mice Immunization
4.8. BALB/c Mice Splenocytes Stimulation in Cell Culture
4.9. mRNA Cytokine Expression from Stimulated Splenocytes
4.10. Enzyme Immunoassay to Evaluate the Immunogenic Potential of rEZIKV
4.11. Indirect ELISA for Evaluation of Anti-rEZIKV-Specific IgG Avidity Maturation
4.12. Detection of rEZIKV Protein by Immunoblotting
4.13. Cross-Reactivity Evaluation of rEZIKV Immunization by ELISA
4.14. Enzyme Immunoassay to Evaluate the Antigenic Potential of rEZIKV
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Isolation and Serological Specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Simpson, D.I.H. Zika Virus Infection in Man. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.-L.; Mallet, H.-P.; Sall, A.A.; Musso, D. Zika Virus, French Polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1085–1086. [Google Scholar] [CrossRef]
- Ferraris, P.; Yssel, H.; Missé, D. Zika Virus Infection: An Update. Microbes. Infect. 2019, 21, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.R.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Faria, N.R.; da Silva Azevedo, R.d.S.; Kraemer, M.U.G.; Souza, R.; Cunha, M.S.; Hill, S.C.; Thézé, J.; Bonsall, M.B.; Bowden, T.A.; Rissanen, I.; et al. Zika Virus in the Americas: Early Epidemiological and Genetic Findings. Science 2016, 352, 345–349. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastère, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré Syndrome Outbreak Associated with Zika Virus Infection in French Polynesia: A Case-Control Study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Miner, J.J.; Diamond, M.S. Zika Virus Pathogenesis and Tissue Tropism. Cell Host Microbe 2017, 21, 134–142. [Google Scholar] [CrossRef]
- de Araújo, T.V.B.; Rodrigues, L.C.; de Alencar Ximenes, R.A.; de Barros Miranda-Filho, D.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; de Albuquerque, M.d.F.P.M.; Souza, W.V.; Braga, C.; et al. Association between Zika Virus Infection and Microcephaly in Brazil, January to May, 2016: Preliminary Report of a Case-Control Study. Lancet Infect. Dis. 2016, 16, 1356–1363. [Google Scholar] [CrossRef]
- Shahrizaila, N.; Lehmann, H.C.; Kuwabara, S. Guillain-Barré Syndrome. Lancet 2021, 397, 1214–1228. [Google Scholar] [CrossRef]
- Brooks, R.B.; Carlos, M.P.; Myers, R.A.; White, M.G.; Bobo-Lenoci, T.; Aplan, D.; Blythe, D.; Feldman, K.A. Likely Sexual Transmission of Zika Virus from a Man with No Symptoms of Infection—Maryland, 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Roche, C.; Robin, E.; Nhan, T.; Teissier, A. Potential Sexual Transmission of Zika Virus. Emerg. Infect. Dis. 2015, 21, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Paz-Bailey, G.; Rosenberg, E.S.; Doyle, K.; Munoz-Jordan, J.; Santiago, G.A.; Klein, L.; Perez-Padilla, J.; Medina, F.A.; Waterman, S.H.; Gubern, C.G.; et al. Persistence of Zika Virus in Body Fluids—Final Report. N. Engl. J. Med. 2018, 379, 1234–1243. [Google Scholar] [CrossRef]
- Barjas-Castro, M.L.; Angerami, R.N.; Cunha, M.S.; Suzuki, A.; Nogueira, J.S.; Rocco, I.M.; Maeda, A.Y.; Vasami, F.G.S.; Katz, G.; Boin, I.F.S.F.; et al. Probable Transfusion-Transmitted Zika Virus in Brazil. Transfusion 2016, 56, 1684–1688. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8Å Resolution Cryo-EM Structure of Zika Virus. Science 2016, 352, 467–470. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Saiz, J.; Martín-acebes, M.A.; Bueno-marí, R.; Salomón, O.D.; Ortiga-carvalho, T.M.; Mendez-otero, R.; Rosado-de-castro, P.H. Zika Virus: What Have We Learnt Since the Start of the Recent Epidemic? Front. Microbiol. 2017, 8, 1554. [Google Scholar] [CrossRef]
- Dai, L.; Song, J.; Lu, X.; Deng, Y.Q.; Musyoki, A.M.; Cheng, H.; Zhang, Y.; Yuan, Y.; Song, H.; Haywood, J.; et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell Host Microbe 2016, 19, 696–704. [Google Scholar] [CrossRef]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F.; et al. Specificity, Cross-Reactivity, and Function of Antibodies Elicited by Zika Virus Infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.; Kim, K.; Lee, H.J.; Jung, Y.J.; Park, J.; Hahn, T.W. Vaccination with a Zika Virus Envelope Domain III Protein Induces Neutralizing Antibodies and Partial Protection against Asian Genotype in Immunocompetent Mice. Trop. Med. Health 2022, 50, 91. [Google Scholar] [CrossRef]
- Wang, Y.; Ling, L.; Zhang, Z.; Marin-Lopez, A. Current Advances in Zika Vaccine Development. Vaccines 2022, 10, 1816. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.; Varela, A.P.M.; Teixeira, T.F.; Cancela, M.P.; Sesterheim, P.; Souza, D.O.; Roehe, P.M.; Silveira, F. Zika Virus Envelope Domain III Recombinant Protein Delivered With Saponin-Based Nanoadjuvant From Quillaja Brasiliensis Enhances Anti-Zika Immune Responses, Including Neutralizing Antibodies and Splenocyte Proliferation. Front. Immunol. 2021, 12, 632714. [Google Scholar] [CrossRef]
- Kim, Y.C.; Dumoux, M.; Owens, R.J.; Reyes-Sandoval, A. Optimization of Small-Scale Production of Zika Virus Envelope Glycoprotein by Transient Expression in HEK293 Cells for ELISA. In Methods in Molecular Biology; Kobinger, G., Racine, T., Eds.; Humana: New York, NY, USA, 2020; pp. 103–112. ISBN 978-1-0716-0581-3. [Google Scholar]
- Kanno, A.I.; Leite, L.C.d.C.; Pereira, L.R.; de Jesus, M.J.R.; Andreata-Santos, R.; Alves, R.P.; Durigon, E.L.; Ferreira, L.C. Optimization and Scale-up Production of Zika Virus ΔNS1 in Escherichia Coli: Application of Response Surface Methodology. AMB Express 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Bioprocessing of Recombinant Proteins: Expression Hosts and Process Development. Front. Bioeng. Biotechnol. 2019, 7, 420. [Google Scholar] [CrossRef]
- Nikolay, A.; Castilho, L.R.; Reichl, U.; Genzel, Y. Propagation of Brazilian Zika Virus Strains in Static and Suspension Cultures Using Vero and BHK Cells. Vaccine 2018, 36, 3140–3145. [Google Scholar] [CrossRef]
- Nikolay, A.; Léon, A.; Schwamborn, K.; Genzel, Y.; Reichl, U. Process Intensification of EB66® Cell Cultivations Leads to High-Yield Yellow Fever and Zika Virus Production. Appl. Microbiol. Biotechnol. 2018, 102, 8725–8737. [Google Scholar] [CrossRef]
- Alvim, R.G.F.; Itabaiana, I.; Castilho, L.R. Zika Virus-like Particles (VLPs): Stable Cell Lines and Continuous Perfusion Processes as a New Potential Vaccine Manufacturing Platform. Vaccine 2019, 37, 6970–6977. [Google Scholar] [CrossRef] [PubMed]
- Campani, G.; Gonçalves da Silva, G.; Zangirolami, T.C.; Perencin de Arruda Ribeiro, M. Recombinant Escherichia Coli Cultivation in a Pressurized Airlift Bioreactor: Assessment of the Influence of Temperature on Oxygen Transfer and Uptake Rates. Bioprocess Biosyst. Eng. 2017, 40, 1621–1633. [Google Scholar] [CrossRef]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor Scale-up and Oxygen Transfer Rate in Microbial Processes: An Overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef]
- Warnock, J.N.; Al-Rubeai, M. Bioreactor Systems for the Production of Biopharmaceuticals from Animal Cells. Biotechnol. Appl. Biochem. 2006, 45, 1. [Google Scholar] [CrossRef] [PubMed]
- Merchuk, J.C. Why Use Air-Lift Bioreactors? Trends Biotechnol. 1990, 8, 66–71. [Google Scholar] [CrossRef]
- AL-Mashhadani, M.K.H.; Wilkinson, S.J.; Zimmerman, W.B. Airlift Bioreactor for Biological Applications with Microbubble Mediated Transport Processes. Chem. Eng. Sci. 2015, 137, 243–253. [Google Scholar] [CrossRef]
- Kim, Y.C.; Lopez-camacho, C.; Nettleship, J.E.; Rahman, N.; Hill, M.L.; Silva-reyes, L.; Ortiz-martinez, G.; Figueroa-aguilar, G.; Mar, M.A.; Vivanco-Cid, H.; et al. Optimization of Zika Virus Envelope Protein Production for ELISA and Correlation of Antibody Titers with Virus Neutralization in Mexican Patients from an Arbovirus Endemic Region. Virol. J. 2018, 15, 193. [Google Scholar] [CrossRef] [PubMed]
- Amaral, M.P.; Apostolico, J.d.S.; Tomita, N.; Coirada, F.C.; Lunardelli, V.A.S.; Fernandes, E.R.; Souza, H.F.S.; Astray, R.M.; Boscardin, S.B.; Rosa, D.S. Homologous Prime-Boost with Zika Virus Envelope Protein and Poly (I:C) Induces Robust Specific Humoral and Cellular Immune Responses. Vaccine 2020, 38, 3653–3664. [Google Scholar] [CrossRef]
- Salod, Z.; Mahomed, O. Mapping Potential Vaccine Candidates Predicted by VaxiJen for Different Viral Pathogens between 2017–2021—A Scoping Review. Vaccines 2022, 10, 1785. [Google Scholar] [CrossRef]
- Carbaugh, D.L.; Baric, R.S.; Lazear, H.M. Envelope Protein Glycosylation Mediates Zika Virus Pathogenesis. J. Virol. 2019, 93, e00113-19. [Google Scholar] [CrossRef] [PubMed]
- Carbaugh, D.L.; Lazear, H.M. Flavivirus Envelope Protein Glycosylation: Impacts on Viral Infection and Pathogenesis. J. Virol. 2020, 94, e00104-20. [Google Scholar] [CrossRef]
- Kostyuchenko, V.A.; Lim, E.X.Y.; Zhang, S.; Fibriansah, G.; Ng, T.; Ooi, J.S.G.; Shi, J.; Lok, S. Structure of the Thermally Stable Zika Virus. Nature 2016, 533, 425–428. [Google Scholar] [CrossRef]
- Liang, H.; Yang, R.; Liu, Z.; Li, M.; Liu, H.; Jin, X. Recombinant Zika Virus Envelope Protein Elicited Protective Immunity against Zika Virus in Immunocompetent Mice. PLoS ONE 2018, 13, e0194860. [Google Scholar] [CrossRef]
- Yang, C.; Zeng, F.; Gao, X.; Zhao, S.; Li, X.; Liu, S.; Li, N.; Deng, C.; Zhang, B.; Gong, R. Characterization of Two Engineered Dimeric Zika Virus Envelope Proteins as Immunogens for Neutralizing Antibody Selection and Vaccine Design. J. Biol. Chem. 2019, 294, 10638–10648. [Google Scholar] [CrossRef] [PubMed]
- Slon Campos, J.L.; Marchese, S.; Rana, J.; Mossenta, M.; Poggianella, M.; Bestagno, M.; Burrone, O.R. Temperature-Dependent Folding Allows Stable Dimerization of Secretory and Virus-Associated e Proteins of Dengue and Zika Viruses in Mammalian Cells. Sci. Rep. 2017, 7, 966. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, D.P. AIRLIFT BIOREACTOR TEC-BIO-P-30-AIR-LIFT. Available online: https://tecnal.com.br/en/produtos/lamina/5941_airlift_bioreactor (accessed on 18 February 2023).
- Han, J.; Qiu, Y.; Yu, J.; Wang, H.; Deng, Y.; Li, X.-F.; Zhao, H.; Sun, H.-X.; Qin, C.-F. Immunization with Truncated Envelope Protein of Zika Virus Induces Protective Immune Response in Mice. Sci. Rep. 2017, 7, 10047. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Dent, M.; Lai, H.; Sun, H.; Chen, Q. Immunization of Zika Virus Envelope Protein Domain III Induces Specific and Neutralizing Immune Responses against Zika Virus. Vaccine 2017, 35, 4287–4294. [Google Scholar] [CrossRef]
- Shin, M.; Kim, K.; Lee, H.-J.; Lee, R.; Jung, Y.-J.; Park, J.; Hahn, T.-W. Zika Virus Baculovirus-expressed Envelope Protein Elicited Humoral and Cellular Immunity in Immunocompetent Mice. Sci. Rep. 2022, 12, 660. [Google Scholar] [CrossRef]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika Virus Antigen in Microalgae: Towards Mucosal Vaccine Development. J. Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-Produced Zika Virus Envelope Protein Elicits Neutralizing Immune Responses That Correlate with Protective Immunity against Zika Virus in Mice. Plant Biotechnol. J. 2018, 16, 572–580. [Google Scholar] [CrossRef]
- Mossenta, M.; Marchese, S.; Poggianella, M.; Campos, J.L.S.; Burrone, O.R. Role of N-Glycosylation on Zika Virus E Protein Secretion, Viral Assembly and Infectivity. Biochem. Biophys. Res. Commun. 2017, 49, 579–586. [Google Scholar] [CrossRef]
- Martins, P.; Machado, D.; Theizen, T.H.; Guarnieri, J.P.O.; Bernardes, B.G.; Gomide, G.P.; Corat, M.A.F.; Abbehausen, C.; Módena, J.L.P.; Melo, C.F.O.R.; et al. Outer Membrane Vesicles from Neisseria Meningitidis (Proteossome) Used for Nanostructured Zika Virus Vaccine Production. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Bauer, G. The Potential Significance of High Avidity Immunoglobulin G (IgG) for Protective Immunity towards SARS-CoV-2. Int. J. Infect. Dis. 2021, 106, 61–64. [Google Scholar] [CrossRef]
- Bauer, G. High Avidity of Vaccine-Induced Immunoglobulin G against SARS-CoV-2: Potential Relevance for Protective Humoral Immunity. Explor. Immunol. 2022, 2, 133–156. [Google Scholar] [CrossRef]
- Chan, P.K.S.; Lim, P.L.; Liu, E.Y.M.; Cheung, J.L.K.; Leung, D.T.M.; Sung, J.J.Y. Antibody Avidity Maturation during Severe Acute Respiratory Syndrome-Associated Coronavirus Infection. J. Infect. Dis. 2005, 192, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Costa, H.H.M.; Orts, D.J.B.; Moura, A.D.; Duarte-neto, A.N.; Cirqueira, C.S.; Réssio, R.A.; Kanamura, C.T.; Miguita, K.; Ferreira, J.E.; Santos, R.T.M.; et al. RBD and Spike DNA-Based Immunization in Rabbits Elicited IgG Avidity Maturation and High Neutralizing Antibody Responses against SARS-CoV-2. Viruses 2023, 15, 555. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.D.; da Costa, H.H.M.; Correa, V.A.; Ana, A.K.; Lindoso, J.A.L.; De Gaspari, E.; Hong, M.A.; Cunha-Junior, J.P.; Prudencio, C.R. Assessment of Avidity Related to IgG Subclasses in SARS-CoV-2 Brazilian Infected Patients. Sci. Rep. 2021, 11, 17642. [Google Scholar] [CrossRef]
- Wang, R.; Gao, N.; Li, Y.; Fan, D.; Zhen, Z.; Feng, K.; Chen, H.; An, J. Cross-Protection against Four Serotypes of Dengue Virus in Mice Conferred by a Zika DNA Vaccine. Front. Cell. Infect. Microbiol. 2019, 9, 147. [Google Scholar] [CrossRef]
- De Alwis, R.; Smith, S.A.; Olivarez, N.P.; Messer, W.B.; Huynh, J.P.; Wahala, W.M.P.B.; White, L.J.; Diamond, M.S.; Baric, R.S.; Crowe, J.E.; et al. Identification of Human Neutralizing Antibodies That Bind to Complex Epitopes on Dengue Virions. Proc. Natl. Acad. Sci. USA 2012, 109, 7439–7444. [Google Scholar] [CrossRef]
- Williams, J. Vector Design for Improved DNA Vaccine Efficacy, Safety and Production. Vaccines 2013, 1, 225–249. [Google Scholar] [CrossRef]
- Oliveira, S.C.; Rosìnha, G.M.S.; De-Brito, C.F.A.; Fonseca, C.T.; Afonso, R.R.; Costa, M.C.M.S.; Goes, A.M.; Rech, E.L.; Azevedo, V. Immunological Properties of Gene Vaccines Delivered by Different Routes. Braz. J. Med. Biol. Res. 1999, 32, 207–214. [Google Scholar] [CrossRef]
- Fischer, C.; Jo, W.K.; Haage, V.; Moreira-Soto, A.; de Oliveira Filho, E.F.; Drexler, J.F. Challenges towards Serologic Diagnostics of Emerging Arboviruses. Clin. Microbiol. Infect. 2021, 27, 1221–1229. [Google Scholar] [CrossRef]
- Ndiaye, O.; Diagne, C.; Abd El Wahed, A.; Dia, F.; Dia, M.; Faye, A.; Leal, S.; dos Santos, M.; Lima Mendonça, M.; da Silva Leite, C.; et al. Use of Envelope Domain III Protein for the Detection of IgG Type Antibodies Specific to Zika Virus by Indirect ELISA. Diagnostics 2023, 13, 462. [Google Scholar] [CrossRef]
- Prudencio, C.R.; Costa, V.G.d.; Rocha, L.B.; Costa, H.H.M.; da Orts, D.J.B.; da Santos, F.R.; Rahal, P.; Lino, N.A.B.; da Conceição, P.J.P.; Bittar, C.; et al. Identification of Zika Virus NS1-Derived Peptides with Potential Applications in Serological Tests. Viruses 2023, 15, 654. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, I.C.L.; Souza, P.F.N.; Marques, L.E.C.; Girão, N.M.; Araújo, F.M.C.; Guedes, M.I.F. New Insights into the Recombinant Proteins and Monoclonal Antibodies Employed to Immunodiagnosis and Control of Zika Virus Infection: A Review. Int. J. Biol. Macromol. 2022, 200, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.K.B.; Mendonça, P.D.; Assis, L.L.K.S.; Prudêncio, C.R.; Guedes, M.I.F.; Marques, E.T.A.; Dutra, R.F. A Redox-Probe-Free Immunosensor Based on Electrocatalytic Prussian Blue Nanostructured Film One-Step-Prepared for Zika Virus Diagnosis. Biosensors 2022, 12, 623. [Google Scholar] [CrossRef] [PubMed]
- Pardy, R.D.; Richer, M.J. Protective to a T: The Role of T Cells during Zika Virus Infection. Cells 2019, 8, 820. [Google Scholar] [CrossRef] [PubMed]
- Ngono, A.E.; Vizcarra, E.A.; Tang, W.W.; Sheets, N.; Joo, Y.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Mapping and Role of the CD8+ T Cell Response During Primary Zika Virus Infection in Mice. Cell Host Microbe 2017, 21, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Ngono, A.E.; Young, M.P.; Bunz, M.; Xu, Z.; Hattakam, S.; Vizcarra, E.; Regla-Nava, J.A.; Tang, W.W.; Yamabhai, M.; Wen, J.; et al. CD4+ T Cells Promote Humoral Immunity and Viral Control during Zika Virus Infection. PLoS Pathog. 2019, 15, e1007474. [Google Scholar] [CrossRef]
- Pardy, R.D.; Rajah, M.M.; Condotta, S.A.; Taylor, N.G.; Sagan, S.M.; Richer, M.J. Analysis of the T Cell Response to Zika Virus and Identification of a Novel CD8+T Cell Epitope in Immunocompetent Mice. PLoS Pathog. 2017, 13, e1006184. [Google Scholar] [CrossRef]
- Tappe, D.; Pérez-Girón, J.V.; Zammarchi, L.; Rissland, J.; Ferreira, D.F.; Jaenisch, T.; Gómez-Medina, S.; Günther, S.; Bartoloni, A.; Muñoz-Fontela, C.; et al. Cytokine Kinetics of Zika Virus-Infected Patients from Acute to Reconvalescent Phase. Med. Microbiol. Immunol. 2016, 205, 269–273. [Google Scholar] [CrossRef]
- Zuñiga, J.; Choreño-Parra, J.A.; Jiménez-Alvarez, L.; Cruz-Lagunas, A.; Márquez-García, J.E.; Ramírez-Martínez, G.; Goodina, A.; Hernández-Montiel, E.; Fernández-López, L.A.; Cabrera-Cornejo, M.F.; et al. A Unique Immune Signature of Serum Cytokine and Chemokine Dynamics in Patients with Zika Virus Infection from a Tropical Region in Southern Mexico. Int. J. Infect. Dis. 2020, 94, 4–11. [Google Scholar] [CrossRef]
- Lum, F.M.; Lye, D.C.B.; Tan, J.J.L.; Lee, B.; Chia, P.Y.; Chua, T.K.; Amrun, S.N.; Kam, Y.W.; Yee, W.X.; Ling, W.P.; et al. Longitudinal Study of Cellular and Systemic Cytokine Signatures to Define the Dynamics of a Balanced Immune Environment during Disease Manifestation in Zika Virus–Infected Patients. J. Infect. Dis. 2018, 218, 814–824. [Google Scholar] [CrossRef]
- Silva, I.B.B.; França, R.K.A.d.O.; Silva, J.M.; Maranhão, A.Q.; Prudencio, C.R. Phage Display as a Strategy to Obtain Anti-Flavivirus Monoclonal Antibodies. In Dengue Fever in a One Health Perspective; Speranca, M.A., Ed.; IntechOpen: London, UK, 2020; pp. 1–20. ISBN 978-1-78985-649-1. [Google Scholar]
- Silva, I.B.B.; Silva, A.S.d.; Cunha, M.S.; Cabral, A.D.; de Oliveira, K.C.A.; De Gaspari, E.; Prudencio, C.R. Zika Virus Serological Diagnosis: Commercial Tests and Monoclonal Antibodies as Tools. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200019. [Google Scholar] [CrossRef] [PubMed]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The Arrive Guidelines 2.0: Updated Guidelines for Reporting Animal Research. PLoS Biol. 2020, 18, 1769–1777. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; Volume 112, pp. 531–552. [Google Scholar]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A Server for Prediction of Protective Antigens, Tumour Antigens and Subunit Vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Barbas, C.F.; Burton, D.R.; Scott, J.K.; Silverman, G.J. Phage Display: A Laboratory Manual, 1st ed.; Cold Springer Harbor Laboratory Press: New York, NY, USA, 2004; ISBN 978-087969740-2. [Google Scholar]
- Green, M.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Springer Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Zhu, D.; Qian, F.; Wu, Y.; Jones, D.S.; Rowe, C.; Narum, D.L.; Duffy, P.; Miller, L.H.; Saul, A. Determination of Protein Concentration for Protein-Protein Conjugates Using Ultraviolet Absorption. J. Immunol. Methods 2013, 387, 317–321. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar] [CrossRef]
- Barenco, P.V.C.; Lourenço, E.V.; Cunha, J.P.; Almeida, K.C.; Roque-Barreira, M.C.; Silva, D.A.O.; Araújo, E.C.B.; Coutinho, L.B.; Oliveira, M.C.; Mineo, T.W.P.; et al. Toxoplasma gondii 70 KDa Heat Shock Protein: Systemic Detection Is Associated with the Death of the Parasites by the Immune Response and Its Increased Expression in the Brain Is Associated with Parasite Replication. PLoS ONE 2014, 9, e96527. [Google Scholar] [CrossRef]
- Chen, C.; Xu, H.; Peng, Y.; Luo, H.; Huang, G.X.; Wu, X.J.; Dai, Y.C.; Luo, H.L.; Zhang, J.A.; Zheng, B.Y.; et al. Elevation in the Counts of IL-35-Producing B Cells Infiltrating into Lung Tissue in Mycobacterial Infection Is Associated with the Downregulation of Th1/Th17 and Upregulation of Foxp3+Treg. Sci. Rep. 2020, 10, 13212. [Google Scholar] [CrossRef]
- Overbergh, L.; Giulietti, A.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. The Use of Real-Time Reverse Transcriptase PCR for the Quantification of Cytokine Gene Expression. J. Biomol. Tech. 2003, 14, 33–43. [Google Scholar]
- Wu, Q.; Martin, R.J.; Rino, J.G.; Breed, R.; Torres, R.M.; Chu, H.W. IL-23-Dependent IL-17 Production Is Essential in Neutrophil Recruitment and Activity in Mouse Lung Defense against Respiratory Mycoplasma Pneumoniae Infection. Microbes Infect. 2007, 9, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Li, X.; Niu, W.; Wang, D.L.; Wu, B.; Yang, C.H. β-Sitosterol Regulated MicroRNAs in Endothelial Cells against an Oxidized Low-Density Lipoprotein. Food Funct. 2020, 11, 1881–1890. [Google Scholar] [CrossRef] [PubMed]
- Greiner, M. Two-Graph Receiver Operating Characteristic (TG-ROC): Update Version Supports Optimisation of Cut-off Values That Minimise Overall Misclassification Costs. J. Immunol. Methods 1996, 191, 93–94. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa, H.H.M.; Bielavsky, M.; Orts, D.J.B.; Araujo, S.; Adriani, P.P.; Nogueira, J.S.; Astray, R.M.; Pandey, R.P.; Lancellotti, M.; Cunha-Junior, J.P.; et al. Production of Recombinant Zika Virus Envelope Protein by Airlift Bioreactor as a New Subunit Vaccine Platform. Int. J. Mol. Sci. 2023, 24, 13955. https://doi.org/10.3390/ijms241813955
da Costa HHM, Bielavsky M, Orts DJB, Araujo S, Adriani PP, Nogueira JS, Astray RM, Pandey RP, Lancellotti M, Cunha-Junior JP, et al. Production of Recombinant Zika Virus Envelope Protein by Airlift Bioreactor as a New Subunit Vaccine Platform. International Journal of Molecular Sciences. 2023; 24(18):13955. https://doi.org/10.3390/ijms241813955
Chicago/Turabian Styleda Costa, Hernan H. M., Monica Bielavsky, Diego J. B. Orts, Sergio Araujo, Patrícia P. Adriani, Juliana S. Nogueira, Renato M. Astray, Ramendra P. Pandey, Marcelo Lancellotti, Jair P. Cunha-Junior, and et al. 2023. "Production of Recombinant Zika Virus Envelope Protein by Airlift Bioreactor as a New Subunit Vaccine Platform" International Journal of Molecular Sciences 24, no. 18: 13955. https://doi.org/10.3390/ijms241813955
APA Styleda Costa, H. H. M., Bielavsky, M., Orts, D. J. B., Araujo, S., Adriani, P. P., Nogueira, J. S., Astray, R. M., Pandey, R. P., Lancellotti, M., Cunha-Junior, J. P., & Prudencio, C. R. (2023). Production of Recombinant Zika Virus Envelope Protein by Airlift Bioreactor as a New Subunit Vaccine Platform. International Journal of Molecular Sciences, 24(18), 13955. https://doi.org/10.3390/ijms241813955