
Regulation of Transporters for Organic Cations by High Glucose


Abstract
:1. Introduction
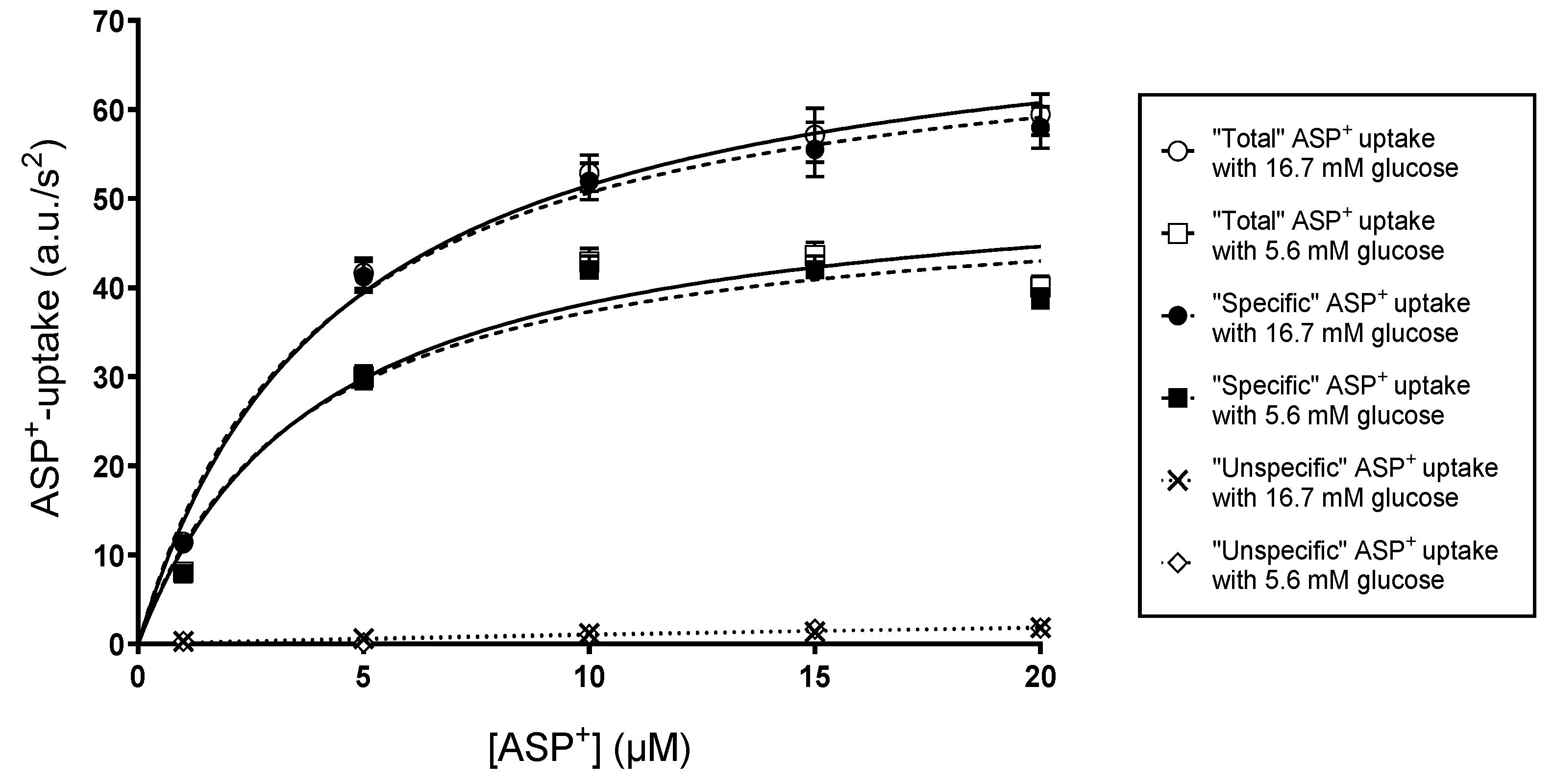
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Measurement of Transporter Function
4.3. Real-Time PCR Analysis
4.4. Biotinylation of Cell Surface Proteins
4.5. Human Samples
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nigam, S.K.; Granados, J.C. OAT, OATP, and MRP Drug Transporters and the Remote Sensing and Signaling Theory. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 637–660. [Google Scholar] [CrossRef]
- Schophuizen, C.M.S.; Wilmer, M.J.; Jansen, J.; Gustavsson, L.; Hilgendorf, C.; Hoenderop, J.G.J.; van den Heuvel, L.P.; Masereeuw, R. Cationic Uremic Toxins Affect Human Renal Proximal Tubule Cell Functioning through Interaction with the Organic Cation Transporter. Pflugers Arch. 2013, 465, 1701–1714. [Google Scholar] [CrossRef]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic Toxins in the Progression of Chronic Kidney Disease and Cardiovascular Disease: Mechanisms and Therapeutic Targets. Toxins 2021, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- De Deyn, P.P.; Vanholder, R.; Eloot, S.; Glorieux, G. Guanidino Compounds as Uremic (Neuro)Toxins. Semin. Dial. 2009, 22, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, M.; Kinoshita, Y.; Hirata, S.; Otagiri, M.; Ohtani, H.; Sawada, Y. Effects of Uremic Serum and Uremic Toxins on Hepatic Uptake of Digoxin. Ther. Drug Monit. 2008, 30, 576–582. [Google Scholar] [CrossRef]
- Otsuka, M.; Matsumoto, T.; Morimoto, R.; Arioka, S.; Omote, H.; Moriyama, Y. A Human Transporter Protein That Mediates the Final Excretion Step for Toxic Organic Cations. Proc. Natl. Acad. Sci. USA 2005, 102, 17923–17928. [Google Scholar] [CrossRef] [PubMed]
- Neuhoff, S.; Ungell, A.-L.; Zamora, I.; Artursson, P. PH-Dependent Bidirectional Transport of Weakly Basic Drugs across Caco-2 Monolayers: Implications for Drug-Drug Interactions. Pharm. Res. 2003, 20, 1141–1148. [Google Scholar] [CrossRef]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific Organic Cation Transporters: Structure, Function, Physiological Roles, and Biopharmaceutical Implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Ahmadimoghaddam, D.; Staud, F. Transfer of Metformin across the Rat Placenta Is Mediated by Organic Cation Transporter 3 (OCT3/SLC22A3) and Multidrug and Toxin Extrusion 1 (MATE1/SLC47A1) Protein. Reprod. Toxicol. 2013, 39, 17–22. [Google Scholar] [CrossRef]
- Kimura, N.; Okuda, M.; Inui, K. Metformin Transport by Renal Basolateral Organic Cation Transporter hOCT2. Pharm. Res. 2005, 22, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Gorecki, M.; Müller, S.F.; Leidolf, R.; Geyer, J. Trospium Chloride Transport by Mouse Drug Carriers of the Slc22 and Slc47 Families. Int. J. Mol. Sci. 2020, 22, 22. [Google Scholar] [CrossRef]
- Wenge, B.; Geyer, J.; Bönisch, H. Oxybutynin and Trospium Are Substrates of the Human Organic Cation Transporters. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 203–208. [Google Scholar] [CrossRef]
- Kantauskaitė, M.; Hucke, A.; Reike, M.; Ahmed Eltayeb, S.; Xiao, C.; Barz, V.; Ciarimboli, G. Rapid Regulation of Human Multidrug and Extrusion Transporters hMATE1 and hMATE2K. Int. J. Mol. Sci. 2020, 21, 5157. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G.; Schröter, R.; Neugebauer, U.; Vollenbröker, B.; Gabriëls, G.; Brzica, H.; Sabolić, I.; Pietig, G.; Pavenstädt, H.; Schlatter, E.; et al. Kidney Transplantation Down-Regulates Expression of Organic Cation Transporters, Which Translocate β-Blockers and Fluoroquinolones. Mol. Pharm. 2013, 10, 2370–2380. [Google Scholar] [CrossRef]
- Ciarimboli, G.; Schlatter, E. Regulation of Organic Cation Transport. Pflugers Arch. 2005, 449, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Rutter, G.A.; Tavaré, J.M.; Palmer, D.G. Regulation of Mammalian Gene Expression by Glucose. Physiology 2000, 15, 149–154. [Google Scholar] [CrossRef]
- Kim, D.; Sands, J.M.; Klein, J.D. Changes in Renal Medullary Transport Proteins during Uncontrolled Diabetes Mellitus in Rats. Am. J. Physiol. Renal Physiol. 2003, 285, F303–F309. [Google Scholar] [CrossRef] [PubMed]
- Marks, J.; Carvou, N.J.C.; Debnam, E.S.; Srai, S.K.; Unwin, R.J. Diabetes Increases Facilitative Glucose Uptake and GLUT2 Expression at the Rat Proximal Tubule Brush Border Membrane. J. Physiol. 2003, 553, 137–145. [Google Scholar] [CrossRef]
- Grover, B.; Auberger, C.; Sarangarajan, R.; Cacini, W. Functional Impairment of Renal Organic Cation Transport in Experimental Diabetes. Pharmacol. Toxicol. 2002, 90, 181–186. [Google Scholar] [CrossRef]
- Thomas, M.C.; Tikellis, C.; Burns, W.C.; Thallas, V.; Forbes, J.M.; Cao, Z.; Osicka, T.M.; Russo, L.M.; Jerums, G.; Ghabrial, H.; et al. Reduced Tubular Cation Transport in Diabetes: Prevented by ACE Inhibition. Kidney Int. 2003, 63, 2152–2161. [Google Scholar] [CrossRef]
- Thomas, M.C.; Tikellis, C.; Kantharidis, P.; Burns, W.C.; Cooper, M.E.; Forbes, J.M. The Role of Advanced Glycation in Reduced Organic Cation Transport Associated with Experimental Diabetes. J. Pharmacol. Exp. Ther. 2004, 311, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.; Huettner, J.P.; Hacker, K.; Bernhardt, G.; König, J.; Buschauer, A. Esters of Bendamustine are by Far More Potent Cytotoxic Agents than the Parent Compound against Human Sarcoma and Carcinoma Cells. PLoS ONE 2015, 10, e0133743. [Google Scholar] [CrossRef] [PubMed]
- Biermann, J.; Lang, D.; Gorboulev, V.; Koepsell, H.; Sindic, A.; Schröter, R.; Zvirbliene, A.; Pavenstädt, H.; Schlatter, E.; Ciarimboli, G. Characterization of Regulatory Mechanisms and States of Human Organic Cation Transporter 2. Am. J. Physiol. Cell Physiol. 2006, 290, C1521–C1531. [Google Scholar] [CrossRef] [PubMed]
- Leprivier, G.; Rotblat, B. How Does MTOR Sense Glucose Starvation? AMPK Is the Usual Suspect. Cell Death Discov. 2020, 6, 27. [Google Scholar] [CrossRef]
- Byles, V.; Cormerais, Y.; Kalafut, K.; Barrera, V.; Hughes Hallett, J.E.; Sui, S.H.; Asara, J.M.; Adams, C.M.; Hoxhaj, G.; Ben-Sahra, I.; et al. Hepatic MTORC1 Signaling Activates ATF4 as Part of Its Metabolic Response to Feeding and Insulin. Mol. Metab. 2021, 53, 101309. [Google Scholar] [CrossRef]
- Kogot-Levin, A.; Hinden, L.; Riahi, Y.; Israeli, T.; Tirosh, B.; Cerasi, E.; Mizrachi, E.B.; Tam, J.; Mosenzon, O.; Leibowitz, G. Proximal Tubule MTORC1 Is a Central Player in the Pathophysiology of Diabetic Nephropathy and Its Correction by SGLT2 Inhibitors. Cell Rep. 2020, 32, 107954. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, N.; Slizgi, J.R.; Brouwer, K.L.R. Effect of Liver Disease on Hepatic Transporter Expression and Function. J. Pharm. Sci. 2017, 106, 2282–2294. [Google Scholar] [CrossRef]
- Oswald, S.; Müller, J.; Neugebauer, U.; Schröter, R.; Herrmann, E.; Pavenstädt, H.; Ciarimboli, G. Protein Abundance of Clinically Relevant Drug Transporters in The Human Kidneys. Int. J. Mol. Sci. 2019, 20, 5303. [Google Scholar] [CrossRef]
- Yang, G.; Francis, D.; Krycer, J.R.; Larance, M.; Zhang, Z.; Novotny, C.J.; Diaz-Vegas, A.; Shokat, K.M.; James, D.E. Dissecting the Biology of MTORC1 beyond Rapamycin. Sci. Signal 2021, 14, eabe0161. [Google Scholar] [CrossRef] [PubMed]
- Koepp, T.N.; Tokaj, A.; Nedvetsky, P.I.; Conchon Costa, A.C.; Snieder, B.; Schröter, R.; Ciarimboli, G. Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model. Int. J. Mol. Sci. 2021, 22, 9658. [Google Scholar] [CrossRef]
- Motohashi, H.; Nakao, Y.; Masuda, S.; Katsura, T.; Kamba, T.; Ogawa, O.; Inui, K.-I. Precise Comparison of Protein Localization among OCT, OAT, and MATE in Human Kidney. J. Pharm. Sci. 2013, 102, 3302–3308. [Google Scholar] [CrossRef]
- Motohashi, H.; Sakurai, Y.; Saito, H.; Masuda, S.; Urakami, Y.; Goto, M.; Fukatsu, A.; Ogawa, O.; Inui, K.-I. Gene Expression Levels and Immunolocalization of Organic Ion Transporters in the Human Kidney. J. Am. Soc. Nephrol. 2002, 13, 866–874. [Google Scholar] [CrossRef]
- Nigam, S.K. The SLC22 Transporter Family: A Paradigm for the Impact of Drug Transporters on Metabolic Pathways, Signaling, and Disease. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 663–687. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Ciarimboli, G.; Struwe, K.; Arndt, P.; Gorboulev, V.; Koepsell, H.; Schlatter, E.; Hirsch, J.R. Regulation of the Human Organic Cation Transporter HOCT1. J. Cell Physiol. 2004, 201, 420–428. [Google Scholar] [CrossRef]
- Martel, F.; Keating, E.; Calhau, C.; Gründemann, D.; Schömig, E.; Azevedo, I. Regulation of Human Extraneuronal Monoamine Transporter (hEMT) Expressed in HEK293 Cells by Intracellular Second Messenger Systems. Naunyn Schmiedebergs Arch. Pharmacol. 2001, 364, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Goralski, K.B.; Stupack, D.G.; Hatch, G.M.; Sitar, D.S. Perturbation of Rat Renal Tubule Transport of the Organic Cation Amantadine in Recent Onset Streptozotocin-Induced Diabetes and in Uninephrectomy. Can. J. Physiol. Pharmacol. 2001, 79, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Harrach, S.; Barz, V.; Pap, T.; Pavenstädt, H.; Schlatter, E.; Edemir, B.; Distler, J.; Ciarimboli, G.; Bertrand, J. Notch Signaling Activity Determines Uptake and Biological Effect of Imatinib in Systemic Sclerosis Dermal Fibroblasts. J. Investig. Dermatol. 2019, 139, 439–447. [Google Scholar] [CrossRef]
- Nakrani, M.N.; Wineland, R.H.; Anjum, F. Physiology, Glucose Metabolism; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar] [PubMed]
- Park, S.H.; Choi, H.J.; Lee, J.H.; Woo, C.H.; Kim, J.H.; Han, H.J. High Glucose Inhibits Renal Proximal Tubule Cell Proliferation and Involves PKC, Oxidative Stress, and TGF-Beta 1. Kidney Int. 2001, 59, 1695–1705. [Google Scholar] [CrossRef]
- Efanova, I.B.; Zaitsev, S.V.; Zhivotovsky, B.; Köhler, M.; Efendić, S.; Orrenius, S.; Berggren, P.O. Glucose and Tolbutamide Induce Apoptosis in Pancreatic Beta-Cells. A Process Dependent on Intracellular Ca2+ Concentration. J. Biol. Chem. 1998, 273, 33501–33507. [Google Scholar] [CrossRef] [PubMed]
- Henry, D.N.; Frank, R.N.; Hootman, S.R.; Rood, S.E.; Heilig, C.W.; Busik, J. V Glucose-Specific Regulation of Aldose Reductase in Human Retinal Pigment Epithelial Cells in Vitro. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1554–1560. [Google Scholar]
- Shirazi-Beechey, S.P.; Moran, A.W.; Batchelor, D.J.; Daly, K.; Al-Rammahi, M. Glucose Sensing and Signalling; Regulation of Intestinal Glucose Transport. Proc. Nutr. Soc. 2011, 70, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Ferrannini, E.; Holman, R.R.; Sherwin, R.; Zinman, B. Medical Management of Hyperglycaemia in Type 2 Diabetes Mellitus: A Consensus Algorithm for the Initiation and Adjustment of Therapy. Diabetologia 2009, 52, 17–30. [Google Scholar] [CrossRef]
- ElSayed, N.A.; Aleppo, G.; Aroda, V.R.; Bannuru, R.R.; Brown, F.M.; Bruemmer, D.; Collins, B.S.; Gaglia, J.L.; Hilliard, M.E.; Isaacs, D.; et al. 2. Classification and Diagnosis of Diabetes: Standards of Care in Diabetes—2023. Diabetes Care 2023, 46, S19–S40. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. Regulation of MTORC1 and Its Impact on Gene Expression at a Glance. J. Cell Sci. 2013, 126, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Monteiro, R.; Pestana, D.; Martel, F.; de Freitas, V.; Mateus, N.; Calhau, C. Impact of Culture Media Glucose Levels on the Intestinal Uptake of Organic Cations. Cytotechnology 2010, 62, 23–29. [Google Scholar] [CrossRef]
- Ma, L.; Wu, X.; Ling-Ling, E.; Wang, D.-S.; Liu, H.-C. The Transmembrane Transport of Metformin by Osteoblasts from Rat Mandible. Arch. Oral. Biol. 2009, 54, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced Glycation End Products and Oxidative Stress in Type 2 Diabetes Mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of Advanced Glycation End Products in Cellular Signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Grover, B.; Buckley, D.; Buckley, A.R.; Cacini, W. Reduced Expression of Organic Cation Transporters ROCT1 and ROCT2 in Experimental Diabetes. J. Pharmacol. Exp. Ther. 2004, 308, 949–956. [Google Scholar] [CrossRef]
- Tonneijck, L.; Muskiet, M.H.A.; Smits, M.M.; van Bommel, E.J.; Heerspink, H.J.L.; van Raalte, D.H.; Joles, J.A. Glomerular Hyperfiltration in Diabetes: Mechanisms, Clinical Significance, and Treatment. J. Am. Soc. Nephrol. 2017, 28, 1023–1039. [Google Scholar] [CrossRef]
- Schorn, S.; Dicke, A.-K.; Neugebauer, U.; Schröter, R.; Friedrich, M.; Reuter, S.; Ciarimboli, G. Expression and Function of Organic Cation Transporter 2 in Pancreas. Front. Cell Dev. Biol. 2021, 9, 688885. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, B.; Wu, A.; Shin, J.-I.; Sang, Y.; Alexander, G.C.; Secora, A.; Inker, L.A.; Coresh, J.; Chang, A.R.; Grams, M.E. Association of Metformin Use with Risk of Lactic Acidosis Across the Range of Kidney Function. JAMA Intern. Med. 2018, 178, 903. [Google Scholar] [CrossRef] [PubMed]
- Hucke, A.; Park, G.Y.; Bauer, O.B.; Beyer, G.; Köppen, C.; Zeeh, D.; Wehe, C.A.; Sperling, M.; Schröter, R.; Kantauskaitè, M.; et al. Interaction of the New Monofunctional Anticancer Agent Phenanthriplatin with Transporters for Organic Cations. Front. Chem. 2018, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-K.; Reichold, M.; Edemir, B.; Ciarimboli, G.; Warth, R.; Koepsell, H.; Thévenod, F. Organic Cation Transporters OCT1, 2, and 3 Mediate High-Affinity Transport of the Mutagenic Vital Dye Ethidium in the Kidney Proximal Tubule. Am. J. Physiol. Renal Physiol. 2009, 296, F1504–F1513. [Google Scholar] [CrossRef]
- Minematsu, T.; Giacomini, K.M. Interactions of Tyrosine Kinase Inhibitors with Organic Cation Transporters and Multidrug and Toxic Compound Extrusion Proteins. Mol. Cancer Ther. 2011, 10, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hOCT1 | hOCT2 | hMATE1 | ||||
|---|---|---|---|---|---|---|
| Km (µM) | Vmax (a.u./s2) | Km (µM) | Vmax (a.u./s2) | Km (µM) | Vmax (a.u./s2) | |
| 5.6 mM glucose | 5 ± 2 | 7 ± 1 | 4 ± 2 | 51 ± 6 | 13 ± 5 | 4 ± 1 |
| 16.7 mM glucose | 10 ± 1 | 18 ± 1 * | 4 ± 1 | 71 ± 4 * | 14 ± 3 | 10 ± 1 * |
| 5.6 mM glucose + 100 nM Torin-1 | 3 ± 2 | 4 ± 1 | 6 ± 2 | 51 ± 6 | 12 ± 6 | 3 ± 1 |
| 16.7 mM glucose + 100 nM Torin-1 | 9 ± 4 | 5 ± 1 | 4 ± 2 | 43 ± 5 # | 16 ± 16 | 6 ± 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinbüchel, M.; Menne, J.; Schröter, R.; Neugebauer, U.; Schlatter, E.; Ciarimboli, G. Regulation of Transporters for Organic Cations by High Glucose. Int. J. Mol. Sci. 2023, 24, 14051. https://doi.org/10.3390/ijms241814051
Steinbüchel M, Menne J, Schröter R, Neugebauer U, Schlatter E, Ciarimboli G. Regulation of Transporters for Organic Cations by High Glucose. International Journal of Molecular Sciences. 2023; 24(18):14051. https://doi.org/10.3390/ijms241814051
Chicago/Turabian StyleSteinbüchel, Martin, Johannes Menne, Rita Schröter, Ute Neugebauer, Eberhard Schlatter, and Giuliano Ciarimboli. 2023. "Regulation of Transporters for Organic Cations by High Glucose" International Journal of Molecular Sciences 24, no. 18: 14051. https://doi.org/10.3390/ijms241814051