
Epitranscriptomics as a New Layer of Regulation of Gene Expression in Skeletal Muscle: Known Functions and Future Perspectives

 ,
,  ,
,  , , , and
, , , and 
Abstract
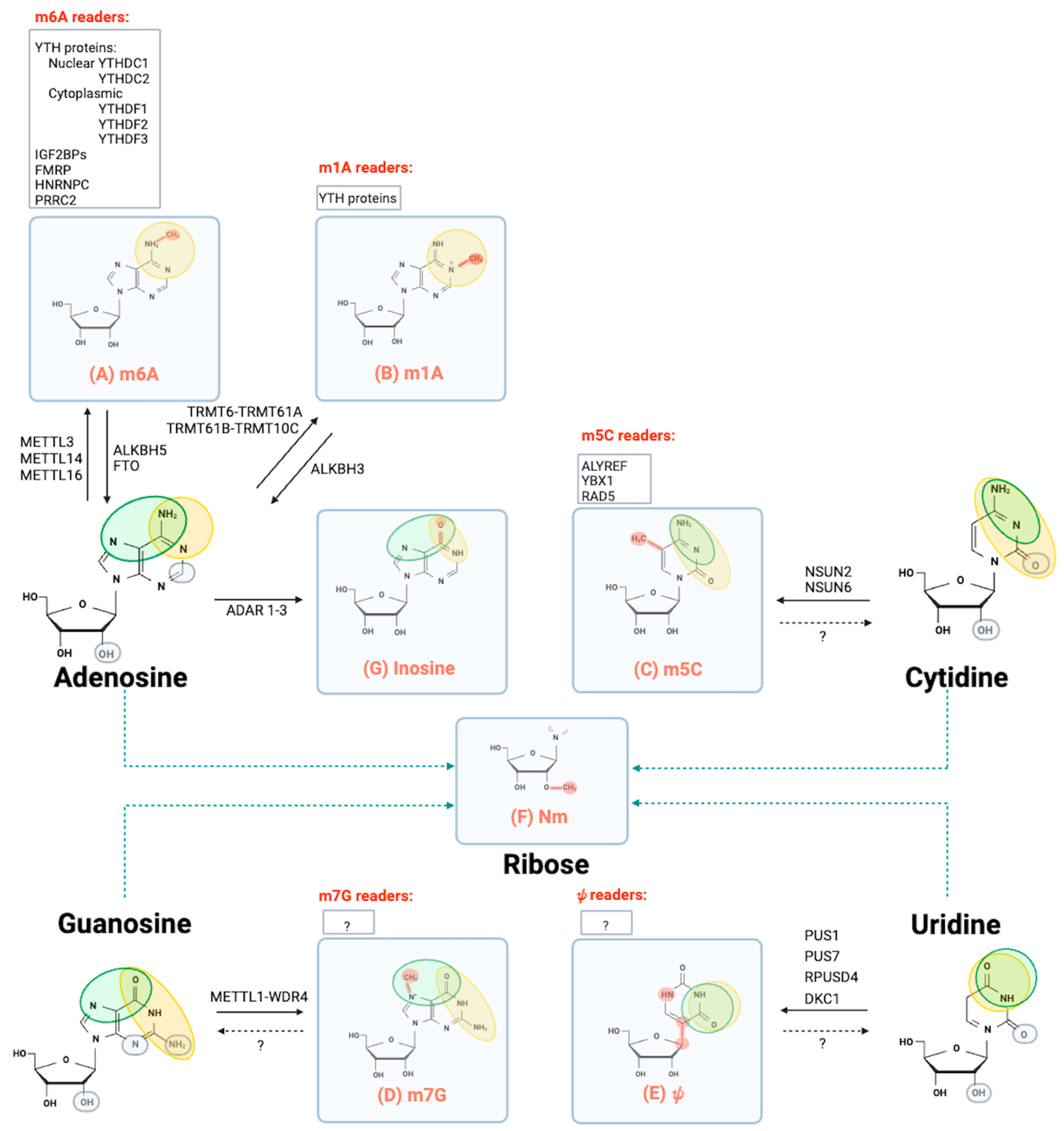
:1. Epitranscriptomics at a Glance
1.1. N⁶-Methyladenosine (m6A)
1.2. N1-Methyladenosine (m1A)
1.3. 5-Methylcytidine (m5C)
1.4. 7-Methylguanosine (m7G)
1.5. Pseudouridine (ψ)
1.6. 2′O-Methylation (Nm or 2′-O-Me)
1.7. Nucleoside Editing
2. Epitranscriptomics in Skeletal Muscle
2.1. N⁶-Methyladenosine Modification in Embryonic Myogenesis
2.2. N⁶-Methyladenosine Modification in Myoblast Differentiation
2.3. N⁶-Methyladenosine Modification in Skeletal Muscle Homeostasis
3. Epitranscriptomics as a Novel Pathogenetic Mechanism and a Potential Therapeutic Approach for Muscular Disorders
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- He, C. Grand Challenge Commentary: RNA Epigenetics? Nat. Chem. Biol. 2010, 6, 863–865. [Google Scholar] [CrossRef]
- Ribonucleic Acids from Yeast Which Contain a Fifth Nucleotide—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/13463012/ (accessed on 12 September 2023).
- Boccaletto, P.; Stefaniak, F.; Ray, A.; Cappannini, A.; Mukherjee, S.; Purta, E.; Kurkowska, M.; Shirvanizadeh, N.; Destefanis, E.; Groza, P.; et al. MODOMICS: A Database of RNA Modification Pathways. 2021 Update. Nucleic Acids Res. 2022, 50, D231–D235. [Google Scholar] [CrossRef]
- Raiber, E.A.; Hardisty, R.; van Delft, P.; Balasubramanian, S. Mapping and Elucidating the Function of Modified Bases in DNA. Nat. Rev. Chem. 2017, 1, 69. [Google Scholar] [CrossRef]
- Halder, S.; Bhattacharyya, D. RNA Structure and Dynamics: A Base Pairing Perspective. Prog. Biophys. Mol. Biol. 2013, 113, 264–283. [Google Scholar] [CrossRef]
- Iwanami, Y.; Brown, G.M. Methylated Bases of Ribosomal Ribonucleic Acid from HeLa Cells. Arch. Biochem. Biophys. 1968, 126, 8–15. [Google Scholar] [CrossRef]
- RajBhandary, U.L.; Stuart, A.; Faulkner, R.D.; Chang, S.H.; Khorana, H.G. Nucleotide Sequence Studies on Yeast Phenylalanine SRNA. Cold Spring Harb. Symp. Quant. Biol. 1966, 31, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Dunn, D.B. The Occurrence of 1-Methyladenine in Ribonucleic Acid. Biochim. Biophys. Acta 1961, 46, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-Methyladenosine in Nuclear RNA Is a Major Substrate of the Obesity-Associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Jimeno, S.; Balestra, F.R.; Huertas, P. The Emerging Role of RNA Modifications in DNA Double-Strand Break Repair. Front. Mol. Biosci. 2021, 8, 664872. [Google Scholar] [CrossRef]
- Nie, F.; Tang, Q.; Liu, Y.; Qin, H.; Liu, S.; Wu, M.; Feng, P.; Chen, W. RNAME: A Comprehensive Database of RNA Modification Enzymes. Comput. Struct. Biotechnol. J. 2022, 20, 6244–6249. [Google Scholar] [CrossRef]
- Jonkhout, N.; Tran, J.; Smith, M.A.; Schonrock, N.; Mattick, J.S.; Novoa, E.M. The RNA Modification Landscape in Human Disease. RNA 2017, 23, 1754–1769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, L.; Li, X. Detection Technologies for RNA Modifications. Exp. Mol. Med. 2022, 54, 1601–1616. [Google Scholar] [CrossRef]
- Li, X.; Xiong, X.; Yi, C. Epitranscriptome Sequencing Technologies: Decoding RNA Modifications. Nat. Methods 2016, 14, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Wiener, D.; Schwartz, S. The Epitranscriptome beyond M6A. Nat. Rev. Genet. 2021, 22, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yankova, E.; Blackaby, W.; Albertella, M.; Rak, J.; De Braekeleer, E.; Tsagkogeorga, G.; Pilka, E.S.; Aspris, D.; Leggate, D.; Hendrick, A.G.; et al. Small-Molecule Inhibition of METTL3 as a Strategy against Myeloid Leukaemia. Nature 2021, 593, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Xuan, J.J.; Sun, W.J.; Lin, P.H.; Zhou, K.R.; Liu, S.; Zheng, L.L.; Qu, L.H.; Yang, J.H. RMBase v2.0: Deciphering the Map of RNA Modifications from Epitranscriptome Sequencing Data. Nucleic Acids Res. 2018, 46, D327–D334. [Google Scholar] [CrossRef]
- Imbriano, C.; Molinari, S. Alternative Splicing of Transcription Factors Genes in Muscle Physiology and Pathology. Genes 2018, 9, 107. [Google Scholar] [CrossRef]
- Benarroch, L.; Bonne, G.; Rivier, F.; Hamroun, D. The 2023 Version of the Gene Table of Neuromuscular Disorders (Nuclear Genome). Neuromuscul. Disord. 2023, 33, 76–117. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.J.; Chen, Q.; et al. Reversible Methylation of M6Am in the 5′ Cap Controls MRNA Stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef]
- Akichika, S.; Hirano, S.; Shichino, Y.; Suzuki, T.; Nishimasu, H.; Ishitani, R.; Sugita, A.; Hirose, Y.; Iwasaki, S.; Nureki, O.; et al. Cap-Specific Terminal N6-Methylation of RNA by an RNA Polymerase II–Associated Methyltransferase. Science 2019, 363, 141. [Google Scholar] [CrossRef]
- Wei, C.-M.; Gershowitz, A.; Moss, B. Methylated Nucleotides Block 5′ Terminus of HeLa Cell Messenger RNA. Cell 1975, 4, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.; Lavi, U.; Darnell, J.E. The Absolute Frequency of Labeled N-6-Methyladenosine in HeLa Cell Messenger RNA Decreases with Label Time. J. Mol. Biol. 1978, 124, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene Expression Regulation Mediated through Reversible M6A RNA Methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Ashraf, S.; Huang, L.; Lilley, D.M.J. Effect of Methylation of Adenine N6 on Kink Turn Structure Depends on Location. RNA Biol. 2019, 16, 1377–1385. [Google Scholar] [CrossRef]
- Boulias, K.; Greer, E.L. Biological Roles of Adenine Methylation in RNA. Nat. Rev. Genet. 2023, 24, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.M.; Moss, B. Nucleotide Sequences at the N6-Methyladenosine Sites of HeLa Cell Messenger Ribonucleic Acid. Biochemistry 1977, 16, 1672–1676. [Google Scholar] [CrossRef]
- Schibler, U.; Kelley, D.E.; Perry, R.P. Comparison of Methylated Sequences in Messenger RNA and Heterogeneous Nuclear RNA from Mouse L Cells. J. Mol. Biol. 1977, 115, 695–714. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive Analysis of MRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; León-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-Wide Mapping Reveals Widespread Dynamic-Regulated Pseudouridylation of NcRNA and MRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5′ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef]
- Slobodin, B.; Han, R.; Calderone, V.; Vrielink, J.A.F.O.; Loayza-Puch, F.; Elkon, R.; Agami, R. Transcription Impacts the Efficiency of MRNA Translation via Co-Transcriptional N6-Adenosine Methylation. Cell 2017, 169, 326–337.e12. [Google Scholar] [CrossRef] [PubMed]
- Bokar, J.A.; Shambaugh, M.E.; Polayes, D.; Matera, A.G.; Rottman, F.M. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. RNA 1997, 3, 1233–1247. [Google Scholar] [PubMed]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Correspondence, N.K.C. The U6 SnRNA m6A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention in Brief an RNA Methyltransferase Controls Cellular SAM Levels through Post-Transcriptional Regulatory Feedback. Cell 2017, 169, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 Is a Mammalian RNA Demethylase That Impacts RNA Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Wu, R.; Li, A.; Sun, B.; Sun, J.G.; Zhang, J.; Zhang, T.; Chen, Y.; Xiao, Y.; Gao, Y.; Zhang, Q.; et al. A Novel M6A Reader Prrc2a Controls Oligodendroglial Specification and Myelination. Cell Res. 2019, 29, 23–41. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-Methyladenosine by IGF2BP Proteins Enhances MRNA Stability and Translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Edupuganti, R.R.; Geiger, S.; Lindeboom, R.G.H.; Shi, H.; Hsu, P.J.; Lu, Z.; Wang, S.Y.; Baltissen, M.P.A.; Jansen, P.W.T.C.; Rossa, M.; et al. N6-Methyladenosine (M6A) Recruits and Repels Proteins to Regulate MRNA Homeostasis. Nat. Struct. Mol. Biol. 2017, 24, 870–878. [Google Scholar] [CrossRef]
- Hsu, P.J.; Shi, H.; Zhu, A.C.; Lu, Z.; Miller, N.; Edens, B.M.; Ma, Y.C.; He, C. The RNA-Binding Protein FMRP Facilitates the Nuclear Export of N6-Methyladenosine-Containing MRNAs. J. Biol. Chem. 2019, 294, 19889–19895. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-Methyladenosine-Dependent RNA Structural Switches Regulate RNA-Protein Interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef]
- Haussmann, I.U.; Bodi, Z.; Sanchez-Moran, E.; Mongan, N.P.; Archer, N.; Fray, R.G.; Soller, M. M6A Potentiates Sxl Alternative Pre-MRNA Splicing for Robust Drosophila Sex Determination. Nature 2016, 540, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Luo, G.Z.; Zhang, Z.; Wang, X.; Zhou, T.; Cui, Y.; Sha, J.; Huang, X.; Guerrero, L.; Xie, P.; et al. YTHDC1 Mediates Nuclear Export of N6-Methyladenosine Methylated MRNAs. eLife 2017, 6, e31311. [Google Scholar] [CrossRef] [PubMed]
- Edens, B.M.; Vissers, C.; Su, J.; Arumugam, S.; Xu, Z.; Shi, H.; Miller, N.; Rojas Ringeling, F.; Ming, G.L.; He, C.; et al. FMRP Modulates Neural Differentiation through M6A-Dependent MRNA Nuclear Export. Cell Rep. 2019, 28, 845–854.e5. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the Human and Mouse M6A RNA Methylomes Revealed by M6A-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Garbo, S.; Zwergel, C.; Battistelli, C. M6A RNA Methylation and beyond—The Epigenetic Machinery and Potential Treatment Options. Drug Discov. Today 2021, 26, 2559–2574. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. The Expanding World of TRNA Modifications and Their Disease Relevance. Nat. Rev. Mol. Cell Biol. 2021, 22, 375–392. [Google Scholar] [CrossRef]
- Leptidis, S.; Papakonstantinou, E.; Diakou, K.I.O.; Pierouli, K.; Mitsis, T.; Dragoumani, K.; Bacopoulou, F.; Sanoudou, D.; Chrousos, G.P.; Vlachakis, D. Epitranscriptomics of Cardiovascular Diseases (Review). Int. J. Mol. Med. 2022, 49, 9. [Google Scholar] [CrossRef]
- Barbieri, I.; Kouzarides, T. Role of RNA Modifications in Cancer. Nat. Rev. Cancer 2020, 20, 303–322. [Google Scholar] [CrossRef]
- Yin, L.; Zhu, X.; Novák, P.; Zhou, L.; Gao, L.; Yang, M.; Zhao, G.J.; Yin, K. The Epitranscriptome of Long Noncoding RNAs in Metabolic Diseases. Clin. Chim. Acta 2021, 515, 80–89. [Google Scholar] [CrossRef]
- Dominissini, D.; Nachtergaele, S.; Moshitch-Moshkovitz, S.; Peer, E.; Kol, N.; Ben-Haim, M.S.; Dai, Q.; Di Segni, A.; Salmon-Divon, M.; Clark, W.C.; et al. The Dynamic N1-Methyladenosine Methylome in Eukaryotic Messenger RNA. Nature 2016, 530, 441–446. [Google Scholar] [CrossRef]
- Li, X.; Xiong, X.; Wang, K.; Wang, L.; Shu, X.; Ma, S.; Yi, C. Transcriptome-Wide Mapping Reveals Reversible and Dynamic N1-Methyladenosine Methylome. Nat. Chem. Biol. 2016, 12, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, K.; Liu, C.; Yi, C. Regulation and Functions of Non-M6A MRNA Modifications. Nat. Rev. Mol. Cell Biol. 2023, 24, 714–731. [Google Scholar] [CrossRef] [PubMed]
- Saikia, M.; Fu, Y.; Pavon-Eternod, M.; Chuan, H.; Pan, T. Genome-Wide Analysis of N1-Methyl-Adenosine Modification in Human TRNAs. RNA 2010, 16, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the Ribosome: The Influence of RRNA Modification on Eukaryotic Ribosome Biogenesis and Function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Safra, M.; Sas-Chen, A.; Nir, R.; Winkler, R.; Nachshon, A.; Bar-Yaacov, D.; Erlacher, M.; Rossmanith, W.; Stern-Ginossar, N.; Schwartz, S. The M1A Landscape on Cytosolic and Mitochondrial MRNA at Single-Base Resolution. Nature 2017, 551, 251–255. [Google Scholar] [CrossRef]
- Li, X.; Xiong, X.; Zhang, M.; Wang, K.; Chen, Y.; Zhou, J.; Mao, Y.; Lv, J.; Yi, D.; Chen, X.W.; et al. Base-Resolution Mapping Reveals Distinct M1A Methylome in Nuclear- and Mitochondrial-Encoded Transcripts. Mol. Cell 2017, 68, 993–1005.e9. [Google Scholar] [CrossRef]
- Zhou, H.; Rauch, S.; Dai, Q.; Cui, X.; Zhang, Z.; Nachtergaele, S.; Sepich, C.; He, C.; Dickinson, B.C. Evolution of a Reverse Transcriptase to Map N1-Methyladenosine in Human Messenger RNA. Nat. Methods 2019, 16, 1281–1288. [Google Scholar] [CrossRef]
- Zhou, H.; Kimsey, I.J.; Nikolova, E.N.; Sathyamoorthy, B.; Grazioli, G.; McSally, J.; Bai, T.; Wunderlich, C.H.; Kreutz, C.; Andricioaei, I.; et al. M(1)A and m(1)G Disrupt A-RNA Structure through the Intrinsic Instability of Hoogsteen Base Pairs. Nat. Struct. Mol. Biol. 2016, 23, 803–810. [Google Scholar] [CrossRef]
- Alriquet, M.; Calloni, G.; Martínez-Limón, A.; Delli Ponti, R.; Hanspach, G.; Hengesbach, M.; Tartaglia, G.G.; Vabulas, R.M. The Protective Role of M1A during Stress-Induced Granulation. J. Mol. Cell Biol. 2021, 12, 870–880. [Google Scholar] [CrossRef]
- Chujo, T.; Suzuki, T. Trmt61B Is a Methyltransferase Responsible for 1-Methyladenosine at Position 58 of Human Mitochondrial TRNAs. RNA 2012, 18, 2269–2276. [Google Scholar] [CrossRef]
- Dai, X.; Wang, T.; Gonzalez, G.; Wang, Y. Identification of YTH Domain-Containing Proteins as the Readers for N1-Methyladenosine in RNA. Anal. Chem. 2018, 90, 6380–6384. [Google Scholar] [CrossRef] [PubMed]
- Dubin, D.T.; Taylor, R.H. The Methylation State of Poly A-Containing Messenger RNA from Cultured Hamster Cells. Nucleic Acids Res. 1975, 2, 1653–1668. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Chen, W.; Liu, J.; Gu, N.; Zhang, R. Genome-Wide Identification of MRNA 5-Methylcytosine in Mammals. Nat. Struct. Mol. Biol. 2019, 26, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread Occurrence of 5-Methylcytosine in Human Coding and Non-Coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef] [PubMed]
- Legrand, C.; Tuorto, F.; Hartmann, M.; Liebers, R.; Jacob, D.; Helm, M.; Lyko, F. Statistically Robust Methylation Calling for Whole-Transcriptome Bisulfite Sequencing Reveals Distinct Methylation Patterns for Mouse RNAs. Genome Res. 2017, 27, 1589–1596. [Google Scholar] [CrossRef]
- Hajnic, M.; Gil, S.A.; Polyansky, A.A.; de Ruiter, A.; Zagrovic, B. Interaction Preferences between Protein Side Chains and Key Epigenetic Modifications 5-Methylcytosine, 5-Hydroxymethycytosine and N6-Methyladenine. Sci. Rep. 2022, 12, 19583. [Google Scholar] [CrossRef]
- Reid, R.; Greene, P.J.; Santi, D.V. Exposition of a Family of RNA m(5)C Methyltransferases from Searching Genomic and Proteomic Sequences. Nucleic Acids Res. 1999, 27, 3138–3145. [Google Scholar] [CrossRef]
- Goll, M.G.; Kirpekar, F.; Maggert, K.A.; Yoder, J.A.; Hsieh, C.L.; Zhang, X.; Golic, K.G.; Jacobsen, S.E.; Bestor, T.H. Methylation of TRNAAsp by the DNA Methyltransferase Homolog Dnmt2. Science 2006, 311, 395–398. [Google Scholar] [CrossRef]
- Selmi, T.; Hussain, S.; DIetmann, S.; Heiß, M.; Borland, K.; Flad, S.; Carter, J.M.; Dennison, R.; Huang, Y.L.; Kellner, S.; et al. Sequence- and Structure-Specific Cytosine-5 MRNA Methylation by NSUN6. Nucleic Acids Res. 2021, 49, 1006–1022. [Google Scholar] [CrossRef]
- Chen, Y.S.; Yang, W.L.; Zhao, Y.L.; Yang, Y.G. Dynamic Transcriptomic M5C and Its Regulatory Role in RNA Processing. Wiley Interdiscip. Rev. RNA 2021, 12, e1639. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, Q.; Shi, Y.; Shi, Q.; Jiang, Y.; Gu, Y.; Li, Z.; Li, X.; Zhao, K.; Wang, C.; et al. Tet2 Promotes Pathogen Infection-Induced Myelopoiesis through MRNA Oxidation. Nature 2018, 554, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, X.; Tang, H.; Jiang, B.; Dou, Y.; Gorospe, M.; Wang, W. NSUN2-Mediated M5C Methylation and METTL3/METTL14-Mediated M6A Methylation Cooperatively Enhance P21 Translation. J. Cell. Biochem. 2017, 118, 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, A.; Sun, B.F.; Yang, Y.; Han, Y.N.; Yuan, X.; Chen, R.X.; Wei, W.S.; Liu, Y.; Gao, C.C.; et al. 5-Methylcytosine Promotes Pathogenesis of Bladder Cancer through Stabilizing MRNAs. Nat. Cell Biol. 2019, 21, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-Methylcytosine Promotes MRNA Export—NSUN2 as the Methyltransferase and ALYREF as an M5C Reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef]
- Schumann, U.; Zhang, H.N.; Sibbritt, T.; Pan, A.; Horvath, A.; Gross, S.; Clark, S.J.; Yang, L.; Preiss, T. Multiple Links between 5-Methylcytosine Content of MRNA and Translation. BMC Biol. 2020, 18, 40. [Google Scholar] [CrossRef]
- Chen, H.; Yang, H.; Zhu, X.; Yadav, T.; Ouyang, J.; Truesdell, S.S.; Tan, J.; Wang, Y.; Duan, M.; Wei, L.; et al. M5C Modification of MRNA Serves a DNA Damage Code to Promote Homologous Recombination. Nat. Commun. 2020, 11, 2834. [Google Scholar] [CrossRef]
- Xue, C.; Zhao, Y.; Li, L. Advances in RNA Cytosine-5 Methylation: Detection, Regulatory Mechanisms, Biological Functions and Links to Cancer. Biomark. Res. 2020, 8, 43. [Google Scholar] [CrossRef]
- Huber, S.M.; Van Delft, P.; Mendil, L.; Bachman, M.; Smollett, K.; Werner, F.; Miska, E.A.; Balasubramanian, S. Formation and Abundance of 5-Hydroxymethylcytosine in RNA. ChemBioChem 2015, 16, 752–755. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Y. Mechanisms and Functions of Tet Protein-Mediated 5-Methylcytosine Oxidation. Genes Dev. 2011, 25, 2436–2452. [Google Scholar] [CrossRef]
- Yang, J.; Bashkenova, N.; Zang, R.; Huang, X.; Wang, J. The Roles of TET Family Proteins in Development and Stem Cells. Development 2020, 147, dev183129. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Both, G.W.; Furuichi, Y.; Shatkin, A.J. 5′-Terminal 7-Methylguanosine in Eukaryotic MRNA Is Required for Translation. Nature 1975, 255, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Murthy, K.G.K.; Park, P.; Manley, J.L. A Nuclear Micrococcal-Sensitive, ATP-Dependent Exoribonuclease Degrades Uncapped but Not Capped RNA Substrates. Nucleic Acids Res. 1991, 19, 2685–2692. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, Y. Discovery of m(7)G-Cap in Eukaryotic MRNAs. Proc. Jpn Acad. Ser. B Phys. Biol. Sci. 2015, 91, 394–409. [Google Scholar] [CrossRef] [PubMed]
- Cowling, V.H. Regulation of MRNA Cap Methylation. Biochem. J. 2010, 425, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Pillutla, R.C.; Yue, Z.; Maldonado, E.; Shatkin, A.J. Recombinant Human MRNA Cap Methyltransferase Binds Capping Enzyme/RNA Polymerase IIo Complexes. J. Biol. Chem. 1998, 273, 21443–21446. [Google Scholar] [CrossRef]
- Gonatopoulos-Pournatzis, T.; Dunn, S.; Bounds, R.; Cowling, V.H. RAM/Fam103a1 Is Required for MRNA Cap Methylation. Mol. Cell 2011, 44, 585–596. [Google Scholar] [CrossRef]
- Shaheen, R.; Abdel-Salam, G.M.H.; Guy, M.P.; Alomar, R.; Abdel-Hamid, M.S.; Afifi, H.H.; Ismail, S.I.; Emam, B.A.; Phizicky, E.M.; Alkuraya, F.S. Mutation in WDR4 Impairs TRNA m(7)G46 Methylation and Causes a Distinct Form of Microcephalic Primordial Dwarfism. Genome Biol. 2015, 16, 210. [Google Scholar] [CrossRef]
- Alexandrov, A.; Martzen, M.R.; Phizicky, E.M. Two Proteins That Form a Complex Are Required for 7-Methylguanosine Modification of Yeast TRNA. RNA 2002, 8, 1253–1266. [Google Scholar] [CrossRef]
- Haag, S.; Kretschmer, J.; Bohnsack, M.T. WBSCR22/Merm1 Is Required for Late Nuclear Pre-Ribosomal RNA Processing and Mediates N7-Methylation of G1639 in Human 18S RRNA. RNA 2015, 21, 180–187. [Google Scholar] [CrossRef]
- Leulliot, N.; Chaillet, M.; Durand, D.; Ulryck, N.; Blondeau, K.; van Tilbeurgh, H. Structure of the Yeast TRNA M7G Methylation Complex. Structure 2008, 16, 52–61. [Google Scholar] [CrossRef]
- Chu, J.M.; Ye, T.T.; Ma, C.J.; Lan, M.D.; Liu, T.; Yuan, B.F.; Feng, Y.Q. Existence of Internal N7-Methylguanosine Modification in MRNA Determined by Differential Enzyme Treatment Coupled with Mass Spectrometry Analysis. ACS Chem. Biol. 2018, 13, 3243–3250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.S.; Liu, C.; Ma, H.; Dai, Q.; Sun, H.L.; Luo, G.; Zhang, Z.; Zhang, L.; Hu, L.; Dong, X.; et al. Transcriptome-Wide Mapping of Internal N7-Methylguanosine Methylome in Mammalian MRNA. Mol. Cell 2019, 74, 1304–1316.e8. [Google Scholar] [CrossRef] [PubMed]
- Malbec, L.; Zhang, T.; Chen, Y.S.; Zhang, Y.; Sun, B.F.; Shi, B.Y.; Zhao, Y.L.; Yang, Y.; Yang, Y.G. Dynamic Methylome of Internal MRNA N7-Methylguanosine and Its Regulatory Role in Translation. Cell Res. 2019, 29, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Qing, Y.; Dong, L.; Han, L.; Wu, D.; Li, Y.; Li, W.; Xue, J.; Zhou, K.; Sun, M.; et al. QKI Shuttles Internal M7G-Modified Transcripts into Stress Granules and Modulates MRNA Metabolism. Cell 2023, 186, 3208–3226.e27. [Google Scholar] [CrossRef] [PubMed]
- Pandolfini, L.; Barbieri, I.; Bannister, A.J.; Hendrick, A.; Andrews, B.; Webster, N.; Murat, P.; Mach, P.; Brandi, R.; Robson, S.C.; et al. METTL1 Promotes Let-7 MicroRNA Processing via M7G Methylation. Mol. Cell 2019, 74, 1278–1290.e9. [Google Scholar] [CrossRef]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem. Sci. 2021, 46, 270–283. [Google Scholar] [CrossRef]
- Deb, I.; Popenda, Ł.; Sarzyńska, J.; Małgowska, M.; Lahiri, A.; Gdaniec, Z.; Kierzek, R. Computational and NMR Studies of RNA Duplexes with an Internal Pseudouridine-Adenosine Base Pair. Sci. Rep. 2019, 9, 16278. [Google Scholar] [CrossRef]
- Jack, K.; Bellodi, C.; Landry, D.M.; Niederer, R.O.; Meskauskas, A.; Musalgaonkar, S.; Kopmar, N.; Krasnykh, O.; Dean, A.M.; Thompson, S.R.; et al. RRNA Pseudouridylation Defects Affect Ribosomal Ligand Binding and Translational Fidelity from Yeast to Human Cells. Mol. Cell 2011, 44, 660–666. [Google Scholar] [CrossRef]
- Morais, P.; Adachi, H.; Yu, Y.T. Spliceosomal SnRNA Epitranscriptomics. Front. Genet. 2021, 12, 652129. [Google Scholar] [CrossRef]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine Profiling Reveals Regulated MRNA Pseudouridylation in Yeast and Human Cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef]
- Karijolich, J.; Yu, Y.T. Converting Nonsense Codons into Sense Codons by Targeted Pseudouridylation. Nature 2011, 474, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, X.; Kierzek, R.; Yu, Y.-T. A Flexible RNA Backbone within the Polypyrimidine Tract Is Required for U2AF65 Binding and Pre-MRNA Splicing In Vivo. Mol. Cell. Biol. 2010, 30, 4108–4119. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.M.; Su, A.; Burns, M.C.; Nussbacher, J.K.; Schaening, C.; Sathe, S.; Yeo, G.W.; Gilbert, W.V. Pseudouridine Synthases Modify Human Pre-MRNA Co-Transcriptionally and Affect Pre-MRNA Processing. Mol. Cell 2022, 82, 645–659.e9. [Google Scholar] [CrossRef] [PubMed]
- Bykhovskaya, Y.; Casas, K.; Mengesha, E.; Inbal, A.; Fischel-Ghodsian, N. Missense Mutation in Pseudouridine Synthase 1 (PUS1) Causes Mitochondrial Myopathy and Sideroblastic Anemia (MLASA). Am. J. Hum. Genet. 2004, 74, 1303–1308. [Google Scholar] [CrossRef]
- Dimitrova, D.G.; Teysset, L.; Carré, C. RNA 2′-O-Methylation (Nm) Modification in Human Diseases. Genes 2019, 10, 117. [Google Scholar] [CrossRef]
- Lo Monaco, P.; Marcel, V.; Diaz, J.J.; Catez, F. 2′-O-Methylation of Ribosomal RNA: Towards an Epitranscriptomic Control of Translation? Biomolecules 2018, 8, 106. [Google Scholar] [CrossRef]
- Decatur, W.A.; Fournier, M.J. RRNA Modifications and Ribosome Function. Trends Biochem. Sci. 2002, 27, 344–351. [Google Scholar] [CrossRef]
- Nachmani, D.; Bothmer, A.H.; Grisendi, S.; Mele, A.; Bothmer, D.; Lee, J.D.; Monteleone, E.; Cheng, K.; Zhang, Y.; Bester, A.C.; et al. Germline NPM1 Mutations Lead to Altered RRNA 2′-O-Methylation and Cause Dyskeratosis Congenita. Nat. Genet. 2019, 51, 1518–1529. [Google Scholar] [CrossRef]
- Sharma, S.; Marchand, V.; Motorin, Y.; Lafontaine, D.L.J. Identification of Sites of 2′-O-Methylation Vulnerability in Human Ribosomal RNAs by Systematic Mapping. Sci. Rep. 2017, 7, 11490. [Google Scholar] [CrossRef]
- Wilkinson, M.L.; Crary, S.M.; Jackman, J.E.; Grayhack, E.J.; Phizicky, E.M. The 2′-O-Methyltransferase Responsible for Modification of Yeast TRNA at Position 4. RNA 2007, 13, 404–413. [Google Scholar] [CrossRef]
- Endres, L.; Rose, R.E.; Doyle, F.; Rahn, T.; Lee, B.; Seaman, J.; McIntyre, W.D.; Fabris, D. 2′-O-Ribose Methylation of Transfer RNA Promotes Recovery from Oxidative Stress in Saccharomyces Cerevisiae. PLoS ONE 2020, 15, e0229103. [Google Scholar] [CrossRef]
- Picard-Jean, F.; Brand, C.; Tremblay-Létourneau, M.; Allaire, A.; Beaudoin, M.C.; Boudreault, S.; Duval, C.; Rainville-Sirois, J.; Robert, F.; Pelletier, J.; et al. 2′-O-Methylation of the MRNA Cap Protects RNAs from Decapping and Degradation by DXO. PLoS ONE 2018, 13, e0193804. [Google Scholar] [CrossRef]
- Dai, Q.; Moshitch-Moshkovitz, S.; Han, D.; Kol, N.; Amariglio, N.; Rechavi, G.; Dominissini, D.; He, C. Nm-Seq Maps 2′-O-Methylation Sites in Human MRNA with Base Precision. Nat. Methods 2017, 14, 695–698. [Google Scholar] [CrossRef]
- Choi, J.; Indrisiunaite, G.; Demirci, H.; Ieong, K.W.; Wang, J.; Petrov, A.; Prabhakar, A.; Rechavi, G.; Dominissini, D.; He, C.; et al. 2′-O-Methylation in MRNA Disrupts TRNA Decoding during Translation Elongation. Nat. Struct. Mol. Biol. 2018, 25, 208–216. [Google Scholar] [CrossRef]
- Elliott, B.A.; Ho, H.T.; Ranganathan, S.V.; Vangaveti, S.; Ilkayeva, O.; Abou Assi, H.; Choi, A.K.; Agris, P.F.; Holley, C.L. Modification of Messenger RNA by 2′-O-Methylation Regulates Gene Expression In Vivo. Nat. Commun. 2019, 10, 3401. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, F.; Stepinski, J.; Darzynkiewicz, E.; Pelletier, J. Characterization of HMTr1, a Human Cap1 2′-O-Ribose Methyltransferase. J. Biol. Chem. 2010, 285, 33037–33044. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.; Purta, E.; Kaminska, K.H.; Cymerman, I.A.; Campbell, D.A.; Mittra, B.; Zamudio, J.R.; Sturm, N.R.; Jaworski, J.; Bujnicki, J.M. 2′-O-Ribose Methylation of Cap2 in Human: Function and Evolution in a Horizontally Mobile Family. Nucleic Acids Res. 2011, 39, 4756–4768. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.M.; Wallis, S.C.; Pease, R.J.; Edwards, Y.H.; Knott, T.J.; Scott, J. A Novel Form of Tissue-Specific RNA Processing Produces Apolipoprotein-B48 in Intestine. Cell 1987, 50, 831–840. [Google Scholar] [CrossRef]
- Smith, H.C.; Bennett, R.P.; Kizilyer, A.; McDougall, W.M.; Prohaska, K.M. Functions and Regulation of the APOBEC Family of Proteins. Semin. Cell Dev. Biol. 2012, 23, 258–268. [Google Scholar] [CrossRef]
- Bass, B.L.; Weintraub, H. A Developmentally Regulated Activity That Unwinds RNA Duplexes. Cell 1987, 48, 607–613. [Google Scholar] [CrossRef]
- Keegan, L.P.; McGurk, L.; Palavicini, J.P.; Brindle, J.; Paro, S.; Li, X.; Rosenthal, J.J.C.; O’Connell, M.A. Functional Conservation in Human and Drosophila of Metazoan ADAR2 Involved in RNA Editing: Loss of ADAR1 in Insects. Nucleic Acids Res. 2011, 39, 7249–7262. [Google Scholar] [CrossRef] [PubMed]
- Slavov, D.; Crnogorac-Jurčević, T.; Clark, M.; Gardiner, K. Comparative Analysis of the DRADA A-to-I RNA Editing Gene from Mammals, Pufferfish and Zebrafish. Gene 2000, 250, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.D.Y.; Kim, T.T.Y.; Walsh, T.; Kobayashi, Y.; Matise, T.C.; Buyske, S.; Gabriel, A. Widespread RNA Editing of Embedded Alu Elements in the Human Transcriptome. Genome Res. 2004, 14, 1719–1725. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.; Li, Q.; Shanmugam, R.; Piskol, R.; Kohler, J.; Young, A.N.; Liu, K.I.; Zhang, R.; Ramaswami, G.; Ariyoshi, K.; et al. Dynamic Landscape and Regulation of RNA Editing in Mammals. Nature 2017, 550, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. A-to-I Editing of Coding and Non-Coding RNAs by ADARs. Nat. Rev. Mol. Cell Biol. 2016, 17, 83–96. [Google Scholar] [CrossRef]
- Mazloomian, A.; Meyer, I.M. Genome-Wide Identification and Characterization of Tissue-Specific RNA Editing Events in D. melanogaster and Their Potential Role in Regulating Alternative Splicing. RNA Biol. 2015, 12, 1391–1401. [Google Scholar] [CrossRef]
- Hwang, T.; Park, C.K.; Leung, A.K.L.; Gao, Y.; Hyde, T.M.; Kleinman, J.E.; Rajpurohit, A.; Tao, R.; Shin, J.H.; Weinberger, D.R. Dynamic Regulation of RNA Editing in Human Brain Development and Disease. Nat. Neurosci. 2016, 19, 1093–1099. [Google Scholar] [CrossRef]
- Huntley, M.A.; Lou, M.; Goldstein, L.D.; Lawrence, M.; Dijkgraaf, G.J.P.; Kaminker, J.S.; Gentleman, R. Complex Regulation of ADAR-Mediated RNA-Editing across Tissues. BMC Genom. 2016, 17, 61. [Google Scholar] [CrossRef]
- Wahlstedt, H.; Daniel, C.; Ensterö, M.; Öhman, M. Large-Scale MRNA Sequencing Determines Global Regulation of RNA Editing during Brain Development. Genome Res. 2009, 19, 978–986. [Google Scholar] [CrossRef]
- Oakes, E.; Anderson, A.; Cohen-Gadol, A.; Hundley, H.A. Adenosine Deaminase That Acts on RNA 3 (ADAR3) Binding to Glutamate Receptor Subunit B Pre-MRNA Inhibits RNA Editing in Glioblastoma. J. Biol. Chem. 2017, 292, 4326–4335. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, X. Genome Sequence-Independent Identification of RNA Editing Sites. Nat. Methods 2015, 12, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Cheng, Y.; Tan, B.C.M.; Kang, L.; Tian, Z.; Zhu, Y.; Zhang, W.; Liang, Y.; Hu, X.; Tan, X.; et al. Comprehensive Analysis of RNA-Seq Data Reveals Extensive RNA Editing in a Human Transcriptome. Nat. Biotechnol. 2012, 30, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Lagergren, J.; Öhman, M. RNA Editing of Non-Coding RNA and Its Role in Gene Regulation. Biochimie 2015, 117, 22–27. [Google Scholar] [CrossRef]
- Eggington, J.M.; Greene, T.; Bass, B.L. Predicting Sites of ADAR Editing in Double-Stranded RNA. Nat. Commun. 2011, 2, 319. [Google Scholar] [CrossRef]
- Xiang, J.F.; Yang, Q.; Liu, C.X.; Wu, M.; Chen, L.L.; Yang, L. N6-Methyladenosines Modulate A-to-I RNA Editing. Mol. Cell 2018, 69, 126–135.e6. [Google Scholar] [CrossRef] [PubMed]
- Rueter, S.M.; Dawson, T.R.; Emeson, R.B. Regulation of Alternative Splicing by RNA Editing. Nature 1999, 399, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Brümmer, A.; Yang, Y.; Chan, T.W.; Xiao, X. Structure-Mediated Modulation of MRNA Abundance by A-to-I Editing. Nat. Commun. 2017, 8, 1255. [Google Scholar] [CrossRef]
- Geula, S.; Moshitch-Moshkovitz, S.; Dominissini, D.; Mansour, A.A.F.; Kol, N.; Salmon-Divon, M.; Hershkovitz, V.; Peer, E.; Mor, N.; Manor, Y.S.; et al. M6A MRNA Methylation Facilitates Resolution of Naïve Pluripotency toward Differentiation. Science 2015, 347, 1002–1006. [Google Scholar] [CrossRef]
- Batista, P.J.; Molinie, B.; Wang, J.; Qu, K.; Zhang, J.; Li, L.; Bouley, D.M.; Lujan, E.; Haddad, B.; Daneshvar, K.; et al. M6A RNA Modification Controls Cell Fate Transition in Mammalian Embryonic Stem Cells. Cell Stem Cell 2014, 15, 707. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6-Methyladenosine Modification Destabilizes Developmental Regulators in Embryonic Stem Cells. Nat. Cell Biol. 2014, 16, 191–198. [Google Scholar] [CrossRef]
- Mendel, M.; Chen, K.M.; Homolka, D.; Gos, P.; Pandey, R.R.; McCarthy, A.A.; Pillai, R.S. Methylation of Structured RNA by the M6A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol. Cell 2018, 71, 986–1000.e11. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Yue, M.; Wang, J.; Kumar, S.; Wechsler-Reya, R.J.; Zhang, Z.; Ogawa, Y.; Kellis, M.; Duester, G.; et al. N6-Methyladenosine RNA Modification Regulates Embryonic Neural Stem Cell Self-Renewal through Histone Modifications. Nat. Neurosci. 2018, 21, 195–206. [Google Scholar] [CrossRef]
- Aguilo, F.; Zhang, F.; Sancho, A.; Fidalgo, M.; Di Cecilia, S.; Vashisht, A.; Lee, D.F.; Chen, C.H.; Rengasamy, M.; Andino, B.; et al. Coordination of M6A MRNA Methylation and Gene Transcription by ZFP217 Regulates Pluripotency and Reprogramming. Cell Stem Cell 2015, 17, 689. [Google Scholar] [CrossRef]
- Li, J.; Pei, Y.; Zhou, R.; Tang, Z.; Yang, Y. Regulation of RNA N6-Methyladenosine Modification and Its Emerging Roles in Skeletal Muscle Development. Int. J. Biol. Sci. 2021, 17, 1682. [Google Scholar] [CrossRef]
- Yu, B.; Liu, J.; Zhang, J.; Mu, T.; Feng, X.; Ma, R.; Gu, Y. Regulatory Role of RNA N6-Methyladenosine Modifications during Skeletal Muscle Development. Front. Cell Dev. Biol. 2022, 10, 929183. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, Y.; Han, J.; Yang, Y.; Chen, Y.; Tang, Z.; Gao, F. Longitudinal Epitranscriptome Profiling Reveals the Crucial Role of N6-Methyladenosine Methylation in Porcine Prenatal Skeletal Muscle Development. J. Genet. Genom. 2020, 47, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Xu, Z.; Lu, L.; Zeng, T.; Gu, L.; Huang, Y.; Zhang, S.; Yang, P.; Wen, Y.; Lin, D.; et al. Transcriptome-Wide Study Revealed M6A Regulation of Embryonic Muscle Development in Dingan Goose (Anser cygnoides orientalis). BMC Genom. 2021, 22, 270. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.; Fan, Y.; Liang, Y.; Cai, Y.; Zhang, G.; Deng, M.; Wang, Z.; Lu, J.; Shi, J.; Wang, F.; et al. FTO-Mediated Demethylation of GADD45B Promotes Myogenesis through the Activation of P38 MAPK Pathway. Mol. Ther. Nucleic Acids 2021, 26, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Liu, S.; Zhang, W.; Xiong, T.; Zhou, M.; Hu, X.; Mao, H.; Liu, S. Profiling Analysis of N6-Methyladenosine MRNA Methylation Reveals Differential M6A Patterns during the Embryonic Skeletal Muscle Development of Ducks. Animals 2022, 12, 2593. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, N.; Yang, M.; Wei, D.; Tai, H.; Han, X.; Gong, H.; Zhou, J.; Qin, J.; Wei, X.; et al. FTO Is Required for Myogenesis by Positively Regulating MTOR-PGC-1α Pathway-Mediated Mitochondria Biogenesis. Cell Death Dis. 2017, 8, e2702. [Google Scholar] [CrossRef]
- Ren, X.; Deng, R.; Zhang, K.; Sun, Y.; Li, Y.; Li, J.; Ren, X.; Deng, R.; Zhang, K.; Sun, Y.; et al. Single-Cell Imaging of M6A Modified RNA Using M6A-Specific In Situ Hybridization Mediated Proximity Ligation Assay (M6AISH-PLA). Angew. Chem. Int. Ed. 2021, 60, 22646–22651. [Google Scholar] [CrossRef]
- Chen, J.N.; Chen, Y.; Wei, Y.Y.; Raza, M.A.; Zou, Q.; Xi, X.Y.; Zhu, L.; Tang, G.Q.; Jiang, Y.Z.; Li, X.W. Regulation of M6A RNA Methylation and Its Effect on Myogenic Differentiation in Murine Myoblasts. Mol. Biol. 2019, 53, 384–392. [Google Scholar] [CrossRef]
- Gheller, B.J.; Blum, J.E.; Fong, E.H.H.; Malysheva, O.V.; Cosgrove, B.D.; Thalacker-Mercer, A.E. A Defined N6-Methyladenosine (M6A) Profile Conferred by METTL3 Regulates Muscle Stem Cell/Myoblast State Transitions. Cell Death Discov. 2020, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Mei, C.; Ma, X.; Du, J.; Wang, J.; Zan, L. M6 A Methylases Regulate Myoblast Proliferation, Apoptosis and Differentiation. Animals 2022, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wu, S.; Zhang, X.; Ren, S.; Tang, Z.; Gao, F. Coordinated Transcriptional and Post-Transcriptional Epigenetic Regulation during Skeletal Muscle Development and Growth in Pigs. J. Anim. Sci. Biotechnol. 2022, 13, 146. [Google Scholar] [CrossRef]
- Gao, X.; Shin, Y.H.; Li, M.; Wang, F.; Tong, Q.; Zhang, P. The Fat Mass and Obesity Associated Gene FTO Functions in the Brain to Regulate Postnatal Growth in Mice. PLoS ONE 2010, 5, e14005. [Google Scholar] [CrossRef]
- Church, C.; Moir, L.; McMurray, F.; Girard, C.; Banks, G.T.; Teboul, L.; Wells, S.; Brüning, J.C.; Nolan, P.M.; Ashcroft, F.M.; et al. Overexpression of Fto Leads to Increased Food Intake and Results in Obesity. Nat. Genet. 2010, 42, 1086–1092. [Google Scholar] [CrossRef]
- Xie, S.J.; Lei, H.; Yang, B.; Diao, L.T.; Liao, J.Y.; He, J.H.; Tao, S.; Hu, Y.X.; Hou, Y.R.; Sun, Y.J.; et al. Dynamic M6A MRNA Methylation Reveals the Role of METTL3/14-M6A-MNK2-ERK Signaling Axis in Skeletal Muscle Differentiation and Regeneration. Front. Cell Dev. Biol. 2021, 9, 744171. [Google Scholar] [CrossRef]
- Kudou, K.; Komatsu, T.; Nogami, J.; Maehara, K.; Harada, A.; Saeki, H.; Oki, E.; Maehara, Y.; Ohkawa, Y. The Requirement of Mettl3-Promoted MyoD MRNA Maintenance in Proliferative Myoblasts for Skeletal Muscle Differentiation. Open Biol. 2017, 7, 170119. [Google Scholar] [CrossRef]
- Liang, Y.; Han, H.; Xiong, Q.; Yang, C.; Wang, L.; Ma, J.; Lin, S.; Jiang, Y.Z. METTL3-Mediated M6A Methylation Regulates Muscle Stem Cells and Muscle Regeneration by Notch Signaling Pathway. Stem Cells Int. 2021, 2021, 9955691. [Google Scholar] [CrossRef]
- Brack, A.S.; Conboy, I.M.; Conboy, M.J.; Shen, J.; Rando, T.A. A Temporal Switch from Notch to Wnt Signaling in Muscle Stem Cells Is Necessary for Normal Adult Myogenesis. Cell Stem Cell 2008, 2, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ning, Y.; Abbas Raza, S.H.; Mei, C.; Zan, L. MEF2C Expression Is Regulated by the Post-Transcriptional Activation of the METTL3-M6A-YTHDF1 Axis in Myoblast Differentiation. Front. Vet. Sci. 2022, 9, 900924. [Google Scholar] [CrossRef] [PubMed]
- Diao, L.T.; Xie, S.J.; Lei, H.; Qiu, X.S.; Huang, M.C.; Tao, S.; Hou, Y.R.; Hu, Y.X.; Sun, Y.J.; Zhang, Q.; et al. METTL3 Regulates Skeletal Muscle Specific MiRNAs at Both Transcriptional and Post-Transcriptional Levels. Biochem. Biophys. Res. Commun. 2021, 552, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.J.; Tao, S.; Diao, L.T.; Li, P.L.; Chen, W.C.; Zhou, Z.G.; Hu, Y.X.; Hou, Y.R.; Lei, H.; Xu, W.Y.; et al. Characterization of Long Non-Coding RNAs Modified by M6A RNA Methylation in Skeletal Myogenesis. Front. Cell Dev. Biol. 2021, 9, 762669. [Google Scholar] [CrossRef]
- Petrosino, J.M.; Hinger, S.A.; Golubeva, V.A.; Barajas, J.M.; Dorn, L.E.; Iyer, C.C.; Sun, H.L.; Arnold, W.D.; He, C.; Accornero, F. The M6A Methyltransferase METTL3 Regulates Muscle Maintenance and Growth in Mice. Nat. Commun. 2022, 13, 168. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, M.; Liu, D.; Shi, Y.; Ren, J.; Wang, S.; Jing, Y.; Zhang, S.; Zhao, Q.; Li, H.; et al. M6A Epitranscriptomic Regulation of Tissue Homeostasis during Primate Aging. Nat. Aging 2023, 3, 705–721. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, T.; Wang, Q.; Fu, R.; Zhang, Z.; Chen, N.; Li, Z.; Gao, G.; Peng, S.; Yang, D. M6A Demethylase ALKBH5 Drives Denervation-Induced Muscle Atrophy by Targeting HDAC4 to Activate FoxO3 Signalling. J. Cachexia-Sarcopenia Muscle 2022, 13, 1210–1223. [Google Scholar] [CrossRef]
- Blanco, S.; Kurowski, A.; Nichols, J.; Watt, F.M.; Benitah, S.A.; Frye, M. The RNA-Methyltransferase Misu (NSun2) Poises Epidermal Stem Cells to Differentiate. PLoS Genet 2011, 7, e1002403. [Google Scholar] [CrossRef]
- Trixl, L.; Amort, T.; Wille, A.; Zinni, M.; Ebner, S.; Hechenberger, C.; Eichin, F.; Gabriel, H.; Schoberleitner, I.; Huang, A.; et al. RNA Cytosine Methyltransferase Nsun3 Regulates Embryonic Stem Cell Differentiation by Promoting Mitochondrial Activity. Cell. Mol. Life Sci. 2018, 75, 1483–1497. [Google Scholar] [CrossRef]
- Blanco, S.; Bandiera, R.; Popis, M.; Hussain, S.; Lombard, P.; Aleksic, J.; Sajini, A.; Tanna, H.; Cortés-Garrido, R.; Gkatza, N.; et al. Stem Cell Function and Stress Response Are Controlled by Protein Synthesis. Nature 2016, 534, 335–340. [Google Scholar] [CrossRef]
- Berdasco, M.; Esteller, M. Towards a Druggable Epitranscriptome: Compounds That Target RNA Modifications in Cancer. Br. J. Pharmacol. 2022, 179, 2868–2889. [Google Scholar] [CrossRef] [PubMed]
- Adachi, H.; Hengesbach, M.; Yu, Y.-T.; Morais, P. Biomedicines Review From Antisense RNA to RNA Modification: Therapeutic Potential of RNA-Based Technologies. Biomedicines 2021, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Rugowska, A.; Starosta, A.; Konieczny, P. Epigenetic Modifications in Muscle Regeneration and Progression of Duchenne Muscular Dystrophy. Clin. Epigenetics 2021, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Caretti, G.; Di Padova, M.; Micales, B.; Lyons, G.E.; Sartorelli, V. The Polycomb Ezh2 Methyltransferase Regulates Muscle Gene Expression and Skeletal Muscle Differentiation. Genes Dev. 2004, 18, 2627–2638. [Google Scholar] [CrossRef] [PubMed]
- Reardon, S. Step aside CRISPR, RNA Editing Is Taking Off. Nature 2020, 578, 24–27. [Google Scholar] [CrossRef]
- Takeshima, Y.; Yagi, M.; Okizuka, Y.; Awano, H.; Zhang, Z.; Yamauchi, Y.; Nishio, H.; Matsuo, M. Mutation Spectrum of the Dystrophin Gene in 442 Duchenne/Becker Muscular Dystrophy Cases from One Japanese Referral Center. J. Hum. Genet. 2010, 55, 379–388. [Google Scholar] [CrossRef]
- Fulco, M.; Schiltz, R.L.; Iezzi, S.; King, M.T.; Zhao, P.; Kashiwaya, Y.; Hoffman, E.; Veech, R.L.; Sartorelli, V. Sir2 Regulates Skeletal Muscle Differentiation as a Potential Sensor of the Redox State. Mol. Cell 2003, 12, 51–62. [Google Scholar] [CrossRef]
- Molinari, S.; Imbriano, C.; Moresi, V.; Renzini, A.; Belluti, S.; Lozanoska-Ochser, B.; Gigli, G.; Cedola, A. Histone Deacetylase Functions and Therapeutic Implications for Adult Skeletal Muscle Metabolism. Front. Mol. Biosci. 2023, 10, 1130183. [Google Scholar] [CrossRef]
- Hori, Y.S.; Kuno, A.; Hosoda, R.; Tanno, M.; Miura, T.; Shimamoto, K.; Horio, Y. Resveratrol Ameliorates Muscular Pathology in the Dystrophic Mdx Mouse, a Model for Duchenne Muscular Dystrophy. Experiment 2011, 338, 784–794. [Google Scholar] [CrossRef]
- Tonkin, J.; Villarroya, F.; Puri, P.L.; Vinciguerra, M. SIRT1 Signaling as Potential Modulator of Skeletal Muscle Diseases. Curr. Opin. Pharmacol. 2012, 12, 372–376. [Google Scholar] [CrossRef]
- Marroncelli, N.; Bianchi, M.; Bertin, M.; Consalvi, S.; Saccone, V.; De Bardi, M.; Puri, P.L.; Palacios, D.; Adamo, S.; Moresi, V. HDAC4 Regulates Satellite Cell Proliferation and Differentiation by Targeting P21 and Sharp1 Genes. Sci. Rep. 2018, 8, 3448. [Google Scholar] [CrossRef]
- Moresi, V.; Williams, A.H.; Meadows, E.; Flynn, J.M.; Potthoff, M.J.; Mcanally, J.; Shelton, J.M.; Backs, J.; Klein, W.H.; Richardson, J.A.; et al. Myogenin and Class II HDACs Control Neurogenic Muscle Atrophy by Inducing E3 Ubiquitin Ligases. Cell 2010, 143, 35–45. [Google Scholar] [CrossRef]
- Pigna, E.; Renzini, A.; Greco, E.; Simonazzi, E.; Fulle, S.; Mancinelli, R.; Moresi, V.; Adamo, S. HDAC4 Preserves Skeletal Muscle Structure Following Long-Term Denervation by Mediating Distinct Cellular Responses. Skelet. Muscle 2018, 8, 6. [Google Scholar] [CrossRef]
- Pigna, E.; Simonazzi, E.; Sanna, K.; Bernadzki, M.; Proszynski, T.; Heil, C.; Palacios, D.; Adamo, S.; Moresi, V. Histone Deacetylase 4 Protects from Denervation and Skeletal Muscle Atrophy in a Murine Model of Amyotrophic Lateral Sclerosis. EBioMedicine 2019, 40, 717–732. [Google Scholar] [CrossRef]
- Renzini, A.; Pigna, E.; Rocchi, M.; Cedola, A.; Gigli, G.; Moresi, V.; Coletti, D. Sex and HDAC4 Differently Affect the Pathophysiology of Amyotrophic Lateral Sclerosis in SOD1-G93A Mice. Int. J. Mol. Sci. 2023, 24, 98. [Google Scholar] [CrossRef]
- Renzini, A.; Marroncelli, N.; Cavioli, G.; Di Francescantonio, S.; Forcina, L.; Lambridis, A.; Di Giorgio, E.; Valente, S.; Mai, A.; Brancolini, C.; et al. Cytoplasmic HDAC4 Regulates the Membrane Repair Mechanism in Duchenne Muscular Dystrophy. J. Cachexia Sarcopenia Muscle 2022, 13, 1339–1359. [Google Scholar] [CrossRef]
- Shen, H.; Xie, K.; Li, M.; Yang, Q.; Wang, X. N6-Methyladenosine (M6A) Methyltransferase METTL3 Regulates Sepsis-Induced Myocardial Injury through IGF2BP1/HDAC4 Dependent Manner. Cell Death Discov. 2022, 8, 322. [Google Scholar] [CrossRef]
- Taylor, M.V.; Hughes, S.M. Mef2 and the Skeletal Muscle Differentiation Program. Semin. Cell Dev. Biol. 2017, 72, 33–44. [Google Scholar] [CrossRef]
- Nakka, K.; Ghigna, C.; Gabellini, D.; Dilworth, F.J. Diversification of the Muscle Proteome through Alternative Splicing. Skelet. Muscle 2018, 8, 8. [Google Scholar] [CrossRef]
- Zhu, B.; Ramachandran, B.; Gulick, T. Alternative Pre-MRNA Splicing Governs Expression of a Conserved Acidic Transactivation Domain in Myocyte Enhancer Factor 2 Factors of Striated Muscle and Brain. J. Biol. Chem. 2005, 280, 28749–28760. [Google Scholar] [CrossRef]
- Sebastian, S.; Faralli, H.; Yao, Z.; Rakopoulos, P.; Palii, C.; Cao, Y.; Singh, K.; Liu, Q.C.; Chu, A.; Aziz, A.; et al. Tissue-Specific Splicing of a Ubiquitously Expressed Transcription Factor Is Essential for Muscle Differentiation. Genes Dev. 2013, 27, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Baruffaldi, F.; Montarras, D.; Basile, V.; de Feo, L.; Badodi, S.; Ganassi, M.; Battini, R.; Nicoletti, C.; Imbriano, C.; Musarò, A.; et al. Dynamic Phosphorylation of the Myocyte Enhancer Factor 2Cα1 Splice Variant Promotes Skeletal Muscle Regeneration and Hypertrophy. Stem Cells 2017, 35, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Ganassi, M.; Badodi, S.; Polacchini, A.; Baruffaldi, F.; Battini, R.; Hughes, S.M.; Hinits, Y.; Molinari, S. Distinct Functions of Alternatively Spliced Isoforms Encoded by Zebrafish Mef2ca and Mef2cb. Biochim. Biophys. Acta Gene Regul. Mech. 2014, 1839, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Yogev, O.; Williams, V.C.; Hinits, Y.; Hughes, S.M. EIF4EBP3L Acts as a Gatekeeper of TORC1 in Activity-Dependent Muscle Growth by Specifically Regulating Mef2ca Translational Initiation. PLoS Biol. 2013, 11, e1001679. [Google Scholar] [CrossRef]
- Black, B.L.; Lu, J.; Olson, E.N. The MEF2A 3′ Untranslated Region Functions as a Cis-Acting Translational Repressor. Mol. Cell. Biol. 1997, 17, 2756–2763. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Luo, W.; Liu, S.; Jiang, H.; Liu, S.; Xu, J.; Chen, B. Cloning of the RNA M6A Methyltransferase 3 and Its Impact on the Proliferation and Differentiation of Quail Myoblasts. Vet. Sci. 2023, 10, 300. [Google Scholar] [CrossRef]
- Adachi, H.; De Zoysa, M.D.; Yu, Y.T. Post-Transcriptional Pseudouridylation in MRNA as Well as in Some Major Types of Noncoding RNAs. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 230–239. [Google Scholar] [CrossRef]
- Lin, T.Y.; Smigiel, R.; Kuzniewska, B.; Chmielewska, J.J.; Kosińska, J.; Biela, M.; Biela, A.; Kościelniak, A.; Dobosz, D.; Laczmanska, I.; et al. Destabilization of Mutated Human PUS3 Protein Causes Intellectual Disability. Hum. Mutat. 2022, 43, 2063–2078. [Google Scholar] [CrossRef]
- Shaheen, R.; Tasak, M.; Maddirevula, S.; Abdel-Salam, G.M.H.; Sayed, I.S.M.; Alazami, A.M.; Al-Sheddi, T.; Alobeid, E.; Phizicky, E.M.; Alkuraya, F.S. PUS7 Mutations Impair Pseudouridylation in Humans and Cause Intellectual Disability and Microcephaly. Hum. Genet. 2019, 138, 231–239. [Google Scholar] [CrossRef]
- Bachinski, L.L.; Sirito, M.; Böhme, M.; Baggerly, K.A.; Udd, B.; Krahe, R. ALTERED MEF2 ISOFORMS IN MYOTONIC DYSTROPHY AND OTHER NEUROMUSCULAR DISORDERS. Muscle Nerve 2010, 42, 856–863. [Google Scholar] [CrossRef]
- Timchenko, N.A.; Patel, R.; Iakova, P.; Cai, Z.J.; Quan, L.; Timchenko, L.T. Overexpression of CUG Triplet Repeat-Binding Protein, CUGBP1, in Mice Inhibits Myogenesis. J. Biol. Chem. 2004, 279, 13129–13139. [Google Scholar] [CrossRef]
- Lee, K.S.; Cao, Y.; Witwicka, H.E.; Tom, S.; Tapscott, S.J.; Wang, E.H. RNA-Binding Protein Muscleblind-like 3 (MBNL3) Disrupts Myocyte Enhancer Factor 2 (Mef2) β-Exon Splicing. J. Biol. Chem. 2010, 285, 33779–33787. [Google Scholar] [CrossRef]
- Basile, V.; Baruffaldi, F.; Dolfini, D.; Belluti, S.; Benatti, P.; Ricci, L.; Artusi, V.; Tagliafico, E.; Mantovani, R.; Molinari, S.; et al. NF-YA Splice Variants Have Different Roles on Muscle Differentiation. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2016, 1859, 627–638. [Google Scholar] [CrossRef]
- Rigillo, G.; Basile, V.; Belluti, S.; Ronzio, M.; Sauta, E.; Ciarrocchi, A.; Latella, L.; Saclier, M.; Molinari, S.; Vallarola, A.; et al. The Transcription Factor NF-Y Participates to Stem Cell Fate Decision and Regeneration in Adult Skeletal Muscle. Nat. Commun. 2021, 12, 6013. [Google Scholar] [CrossRef]
- Libetti, D.; Bernardini, A.; Sertic, S.; Messina, G.; Dolfini, D.; Mantovani, R. The Switch from NF-YAl to NF-YAs Isoform Impairs Myotubes Formation. Cells 2020, 9, 789. [Google Scholar] [CrossRef] [PubMed]
- Rigillo, G.; Belluti, S.; Campani, V.; Ragazzini, G.; Ronzio, M.; Miserocchi, G.; Bighi, B.; Cuoghi, L.; Mularoni, V.; Zappavigna, V.; et al. The NF-Y Splicing Signature Controls Hybrid EMT and ECM-Related Pathways to Promote Aggressiveness of Colon Cancer. Cancer Lett. 2023, 567, 216262. [Google Scholar] [CrossRef] [PubMed]
- Belluti, S.; Semeghini, V.; Rigillo, G.; Ronzio, M.; Benati, D.; Torricelli, F.; Reggiani Bonetti, L.; Carnevale, G.; Grisendi, G.; Ciarrocchi, A.; et al. Alternative Splicing of NF-YA Promotes Prostate Cancer Aggressiveness and Represents a New Molecular Marker for Clinical Stratification of Patients. J. Exp. Clin. Cancer Res. 2021, 40, 362. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, I.; Tzelepis, K.; Pandolfini, L.; Shi, J.; Millán-Zambrano, G.; Robson, S.C.; Aspris, D.; Migliori, V.; Bannister, A.J.; Han, N.; et al. Promoter-Bound METTL3 Maintains Myeloid Leukaemia by M6A-Dependent Translation Control. Nature 2017, 552, 126–131. [Google Scholar] [CrossRef]
- Paris, N.D.; Soroka, A.; Klose, A.; Liu, W.; Chakkalakal, J. V Smad4 Restricts Differentiation to Promote Expansion of Satellite Cell Derived Progenitors during Skeletal Muscle Regeneration. Abstract 2016, 5, e19484. [Google Scholar] [CrossRef]
- Forouhan, M.; Lim, W.F.; Zanetti-Domingues, L.C.; Tynan, C.J.; Roberts, T.C.; Malik, B.; Manzano, R.; Speciale, A.A.; Ellerington, R.; Garcia-Guerra, A.; et al. AR Cooperates with SMAD4 to Maintain Skeletal Muscle Homeostasis. Acta Neuropathol. 2022, 143, 713–731. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Modified Ribonucleoside | Stoichiometry | Target Sequence | mRNA Preferred Regions |
|---|---|---|---|
| m6A | 0.2–0.6% m6A/A | DRACH | 5′ UTR, near the stop codon, 3′ UTR |
| m1A | 0.01–0.05% m1A/A | GC-rich and GA-rich sequence, GUUCNANNC | All segments of the transcripts, enriched in: first splice site, GC-rich highly structured regions in the 5′ UTRs, translation start site |
| m5C | 0.03–0.1% m5C/C | CTCCA | 5′/3′ UTRs, next to argonaute-binding regions |
| m7G | 0.02–0.05% m7G/G | GA- or GG-enriched sequence motifs AG-rich sequence | 5′UTR near the start codon 3′UTR |
| ψ | 0.2–0.6% ψ/U | AU (PUS1-specific) GUUCNANYCY (PUS4) UGUAG PUS7) | Coding sequence and 3′UTR. |
| Nm | 0.01% Nm/N | NmAGAUC followed by a 5 nt-long AG-rich stretch | CDS, near splice sites, 5′/3′ UTR, introns, alternatively spliced regions |
| A to I | Not available | dsRNA, preference for U in the −1 position | Introns (Alu sequences) and UTRs; coding sequence |
| Gene | Gene ID | m6A | m1A | m5C | m7G | PseudoU | 2′-O-Me | RNA-Editing, A-I Sites | Total |
|---|---|---|---|---|---|---|---|---|---|
| ACTA1 | ENSG00000143632.14 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 3 |
| ANO5 | ENSG00000171714.11 | 23 | 0 | 0 | 0 | 0 | 0 | 3 | 26 |
| B3GALNT2 | ENSG00000162885.13 | 13 | 0 | 1 | 0 | 0 | 1 | 8 | 23 |
| B4GAT1 | ENSG00000174684.7 | 27 | 1 | 1 | 0 | 0 | 0 | 0 | 29 |
| BVES | ENSG00000112276.14 | 11 | 0 | 0 | 0 | 0 | 0 | 1 | 12 |
| CACNA1S | ENSG00000081248.11 | 6 | 0 | 2 | 0 | 0 | 0 | 0 | 8 |
| CAPN3 | ENSG00000092529.24 | 17 | 0 | 0 | 0 | 0 | 0 | 2 | 19 |
| CAV3 | ENSG00000182533.6 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 7 |
| CAVIN1 | ENSG00000177469.13 | 45 | 0 | 1 | 0 | 0 | 0 | 0 | 46 |
| CHKB | ENSG00000100288.19 | 25 | 0 | 0 | 0 | 0 | 0 | 1 | 26 |
| COL12A1 | ENSG00000111799.21 | 203 | 0 | 0 | 0 | 0 | 0 | 0 | 203 |
| COL6A1 | ENSG00000142156.14 | 93 | 0 | 0 | 0 | 0 | 0 | 0 | 93 |
| COL6A2 | ENSG00000142173.15 | 88 | 0 | 0 | 0 | 0 | 0 | 3 | 91 |
| COL6A3 | ENSG00000163359.15 | 144 | 0 | 1 | 0 | 0 | 0 | 5 | 150 |
| DAG1 | ENSG00000173402.11 | 138 | 0 | 2 | 0 | 0 | 1 | 6 | 147 |
| DES | ENSG00000175084.11 | 35 | 0 | 1 | 0 | 0 | 0 | 0 | 36 |
| DMD | ENSG00000198947.15 | 116 | 0 | 2 | 0 | 0 | 0 | 19 | 137 |
| DNAJB6 | ENSG00000105993.15 | 86 | 0 | 14 | 0 | 0 | 0 | 8 | 108 |
| DNM2 | ENSG00000079805.16 | 118 | 0 | 13 | 0 | 1 | 0 | 29 | 161 |
| DPM1 | ENSG00000000419.12 | 25 | 0 | 0 | 0 | 0 | 1 | 16 | 42 |
| DPM2 | ENSG00000136908.17 | 54 | 0 | 0 | 0 | 0 | 0 | 0 | 54 |
| DPM3 | ENSG00000179085.7 | 6 | 0 | 1 | 0 | 0 | 0 | 0 | 7 |
| DYSF | ENSG00000135636.14 | 33 | 0 | 4 | 0 | 0 | 0 | 1 | 38 |
| EMD | ENSG00000102119.10 | 22 | 0 | 10 | 0 | 0 | 0 | 0 | 32 |
| FHL1 | ENSG00000022267.17 | 76 | 0 | 0 | 0 | 4 | 0 | 2 | 82 |
| FKRP | ENSG00000181027.10 | 64 | 0 | 0 | 0 | 0 | 0 | 12 | 76 |
| FKTN | ENSG00000106692.14 | 49 | 0 | 0 | 0 | 1 | 0 | 4 | 54 |
| GAA | ENSG00000171298.13 | 69 | 0 | 6 | 0 | 2 | 0 | 0 | 77 |
| GGPS1 | ENSG00000152904.11 | 83 | 0 | 5 | 0 | 0 | 0 | 1 | 89 |
| GMPPB | ENSG00000173540.12 | 76 | 0 | 0 | 0 | 0 | 0 | 1 | 77 |
| GOLGA2 | ENSG00000167110.17 | 81 | 0 | 3 | 0 | 0 | 0 | 10 | 94 |
| GOSR2 | ENSG00000108433.16 | 139 | 0 | 0 | 0 | 0 | 0 | 3 | 142 |
| HNRNPDL | ENSG00000152795.17 | 94 | 0 | 3 | 0 | 0 | 1 | 0 | 98 |
| INPP5K | ENSG00000132376.20 | 62 | 0 | 1 | 0 | 0 | 0 | 1 | 64 |
| ITGA7 | ENSG00000135424.16 | 39 | 0 | 0 | 0 | 0 | 0 | 0 | 39 |
| JAG2 | ENSG00000184916.9 | 48 | 0 | 4 | 1 | 0 | 0 | 0 | 53 |
| LAMA2 | ENSG00000196569.12 | 21 | 0 | 0 | 0 | 0 | 1 | 3 | 25 |
| LARGE1 | ENSG00000133424.20 | 62 | 0 | 0 | 0 | 0 | 1 | 63 | 126 |
| LIMS2 | ENSG00000072163.19 | 27 | 0 | 0 | 0 | 0 | 0 | 2 | 29 |
| LMNA | ENSG00000160789.20 | 72 | 0 | 5 | 0 | 0 | 1 | 22 | 100 |
| LRIF1 | ENSG00000121931.16 | 72 | 0 | 0 | 0 | 0 | 0 | 0 | 72 |
| MPDU1 | ENSG00000129255.16 | 36 | 0 | 2 | 0 | 0 | 0 | 0 | 38 |
| MSTO1 | ENSG00000125459.15 | 33 | 0 | 1 | 0 | 1 | 0 | 0 | 35 |
| MYOT | ENSG00000120729.9 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 7 |
| PLEC | ENSG00000178209.15 | 167 | 0 | 26 | 1 | 2 | 0 | 21 | 217 |
| POGLUT1 | ENSG00000163389.12 | 37 | 0 | 1 | 0 | 1 | 0 | 1 | 40 |
| POMGNT1 | ENSG00000085998.14 | 59 | 0 | 1 | 0 | 0 | 0 | 0 | 60 |
| POMGNT2 | ENSG00000144647.6 | 49 | 0 | 1 | 0 | 0 | 0 | 1 | 51 |
| POMK | ENSG00000185900.9 | 24 | 0 | 0 | 0 | 0 | 0 | 0 | 24 |
| POMT1 | ENSG00000130714.16 | 53 | 0 | 4 | 0 | 0 | 0 | 16 | 73 |
| POMT2 | ENSG00000009830.11 | 66 | 0 | 2 | 0 | 0 | 0 | 5 | 73 |
| POPDC3 | ENSG00000132429.10 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 25 |
| PYROXD1 | ENSG00000121350.16 | 48 | 0 | 1 | 0 | 2 | 0 | 0 | 51 |
| RXYLT1 | ENSG00000118600.11 | 23 | 0 | 2 | 0 | 0 | 0 | 1 | 26 |
| RYR1 | ENSG00000196218.12 | 32 | 0 | 3 | 0 | 0 | 1 | 1 | 37 |
| SELENON | ENSG00000162430.17 | 59 | 0 | 4 | 0 | 0 | 0 | 15 | 78 |
| SGCA | ENSG00000108823.16 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| SGCB | ENSG00000163069.12 | 61 | 0 | 17 | 0 | 0 | 0 | 2 | 80 |
| SGCG | ENSG00000102683.7 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 6 |
| SMCHD1 | ENSG00000101596.15 | 153 | 0 | 2 | 0 | 0 | 0 | 16 | 171 |
| SYNE1 | ENSG00000131018.23 | 339 | 0 | 4 | 0 | 0 | 2 | 50 | 399 |
| SYNE2 | ENSG00000054654.16 | 482 | 0 | 2 | 0 | 0 | 0 | 22 | 506 |
| TCAP | ENSG00000173991.5 | 11 | 0 | 4 | 0 | 0 | 0 | 0 | 15 |
| TMEM43 | ENSG00000170876.7 | 89 | 0 | 2 | 0 | 0 | 0 | 0 | 91 |
| TNPO3 | ENSG00000064419.13 | 94 | 0 | 1 | 0 | 0 | 0 | 8 | 103 |
| TOR1AIP1 | ENSG00000143337.18 | 90 | 0 | 5 | 0 | 0 | 0 | 3 | 98 |
| TRAPPC11 | ENSG00000168538.16 | 67 | 0 | 0 | 0 | 0 | 0 | 3 | 70 |
| TRIM32 | ENSG00000119401.10 | 38 | 0 | 0 | 0 | 1 | 0 | 0 | 39 |
| TRIP4 | ENSG00000103671.9 | 23 | 0 | 0 | 0 | 0 | 0 | 11 | 34 |
| TTN | ENSG00000155657.26 | 249 | 0 | 5 | 0 | 0 | 1 | 0 | 255 |
| VCP | ENSG00000165280.16 | 84 | 0 | 10 | 0 | 0 | 6 | 0 | 100 |
| Gene | Gene ID | m6A | m1A | m5C | m7G | PseudoU | 2′-O-Me | RNA-Editing, A-I Sites | Total |
|---|---|---|---|---|---|---|---|---|---|
| EZH2 | ENSG00000106462.10 | 52 | 0 | 0 | 0 | 0 | 1 | 4 | 57 |
| HDAC4 | ENSG00000068024.16 | 80 | 0 | 17 | 0 | 0 | 0 | 12 | 109 |
| MEF2A | ENSG00000068305.17 | 128 | 0 | 1 | 0 | 2 | 0 | 49 | 180 |
| MEF2C | ENSG00000081189.15 | 41 | 0 | 0 | 0 | 0 | 0 | 3 | 44 |
| MEF2D | ENSG00000116604.18 | 38 | 0 | 2 | 0 | 0 | 0 | 8 | 48 |
| MYF5 | ENSG00000111049.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| MYH1 | ENSG00000109061.10 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| MYH2 | ENSG00000125414.19 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 3 |
| MYOD1 | ENSG00000129152.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| NFYA | ENSG00000001167.14 | 79 | 0 | 6 | 1 | 0 | 0 | 1 | 87 |
| PAX3 | ENSG00000135903.19 | 59 | 0 | 0 | 0 | 0 | 0 | 0 | 59 |
| PAX7 | ENSG00000009709.12 | 37 | 0 | 0 | 0 | 0 | 0 | 0 | 37 |
| SIRT1 | ENSG00000096717.12 | 76 | 0 | 0 | 0 | 0 | 0 | 27 | 103 |
| SMAD4 | ENSG00000141646.13 | 104 | 0 | 4 | 0 | 0 | 0 | 5 | 113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imbriano, C.; Moresi, V.; Belluti, S.; Renzini, A.; Cavioli, G.; Maretti, E.; Molinari, S. Epitranscriptomics as a New Layer of Regulation of Gene Expression in Skeletal Muscle: Known Functions and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 15161. https://doi.org/10.3390/ijms242015161
Imbriano C, Moresi V, Belluti S, Renzini A, Cavioli G, Maretti E, Molinari S. Epitranscriptomics as a New Layer of Regulation of Gene Expression in Skeletal Muscle: Known Functions and Future Perspectives. International Journal of Molecular Sciences. 2023; 24(20):15161. https://doi.org/10.3390/ijms242015161
Chicago/Turabian StyleImbriano, Carol, Viviana Moresi, Silvia Belluti, Alessandra Renzini, Giorgia Cavioli, Eleonora Maretti, and Susanna Molinari. 2023. "Epitranscriptomics as a New Layer of Regulation of Gene Expression in Skeletal Muscle: Known Functions and Future Perspectives" International Journal of Molecular Sciences 24, no. 20: 15161. https://doi.org/10.3390/ijms242015161
APA StyleImbriano, C., Moresi, V., Belluti, S., Renzini, A., Cavioli, G., Maretti, E., & Molinari, S. (2023). Epitranscriptomics as a New Layer of Regulation of Gene Expression in Skeletal Muscle: Known Functions and Future Perspectives. International Journal of Molecular Sciences, 24(20), 15161. https://doi.org/10.3390/ijms242015161