
Triterpenoids from Kochiae Fructus: Glucose Uptake in 3T3-L1 Adipocytes and α-Glucosidase Inhibition, In Silico Molecular Docking
Abstract
:
1. Introduction
2. Results and Discussion
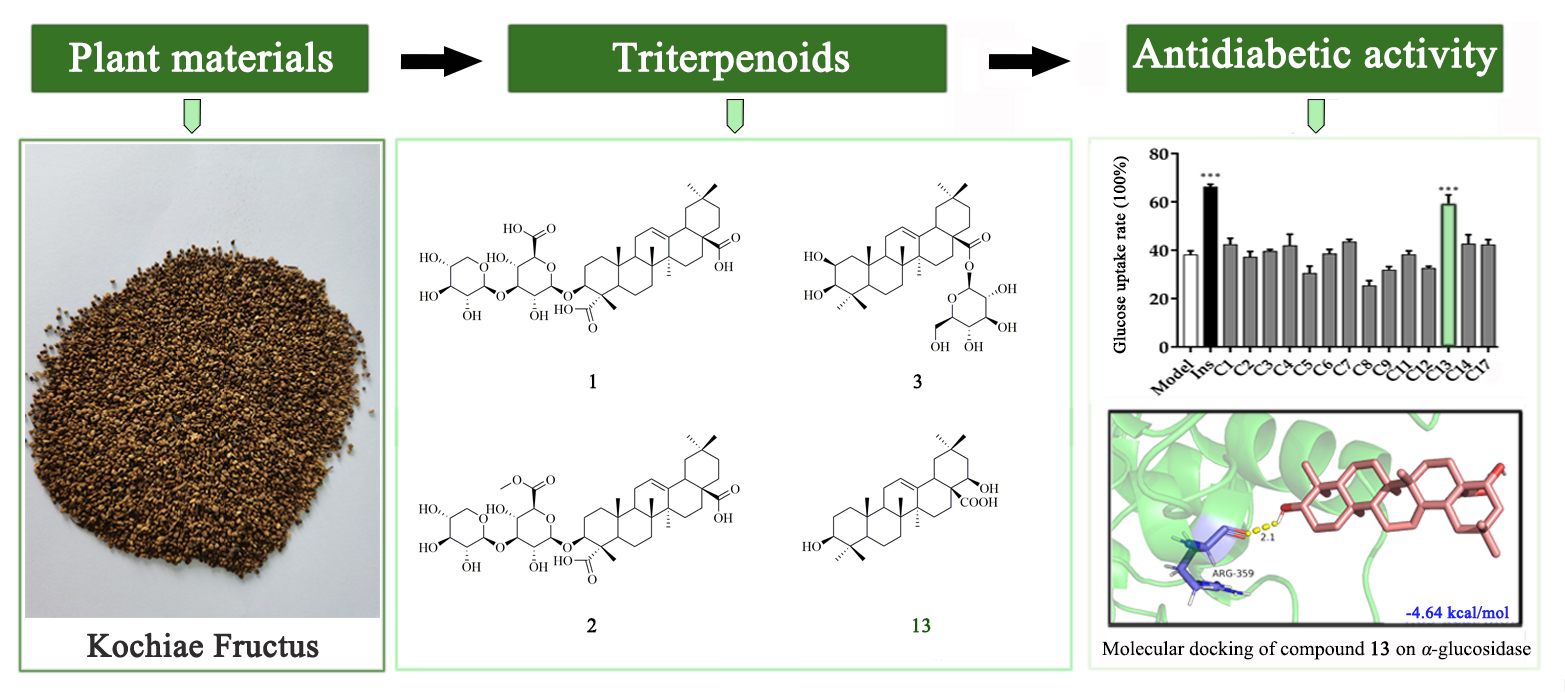
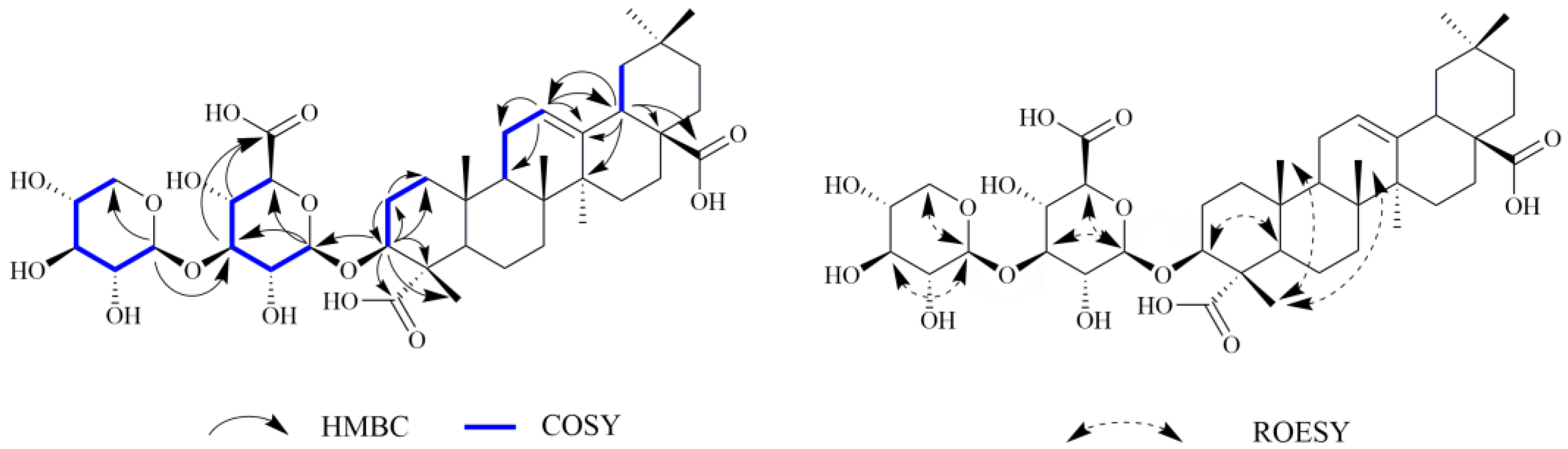
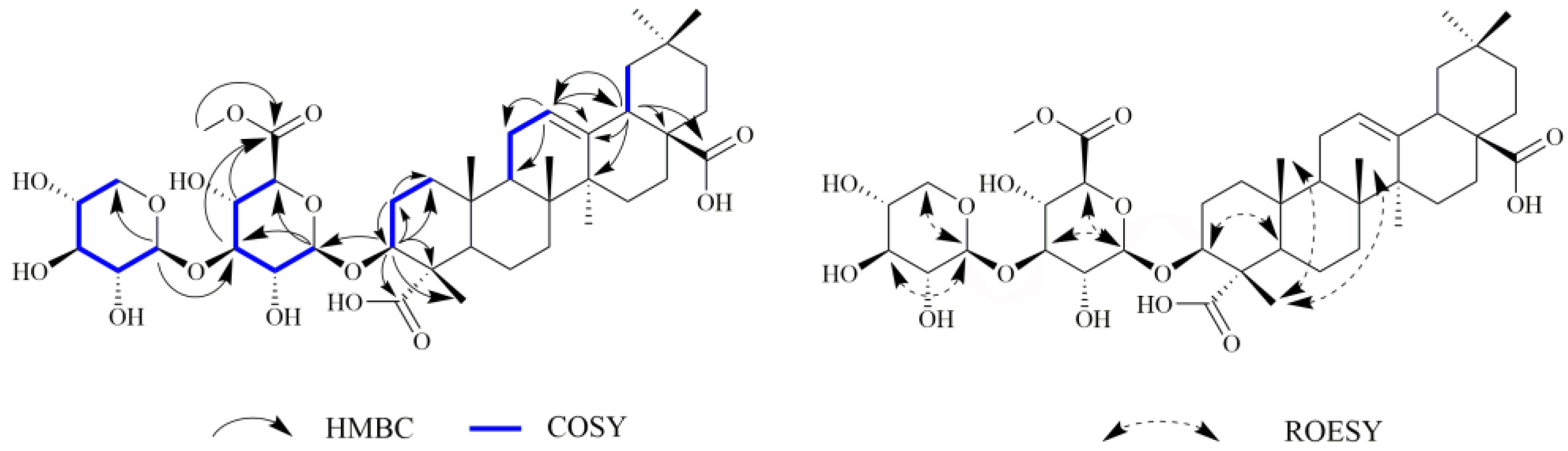
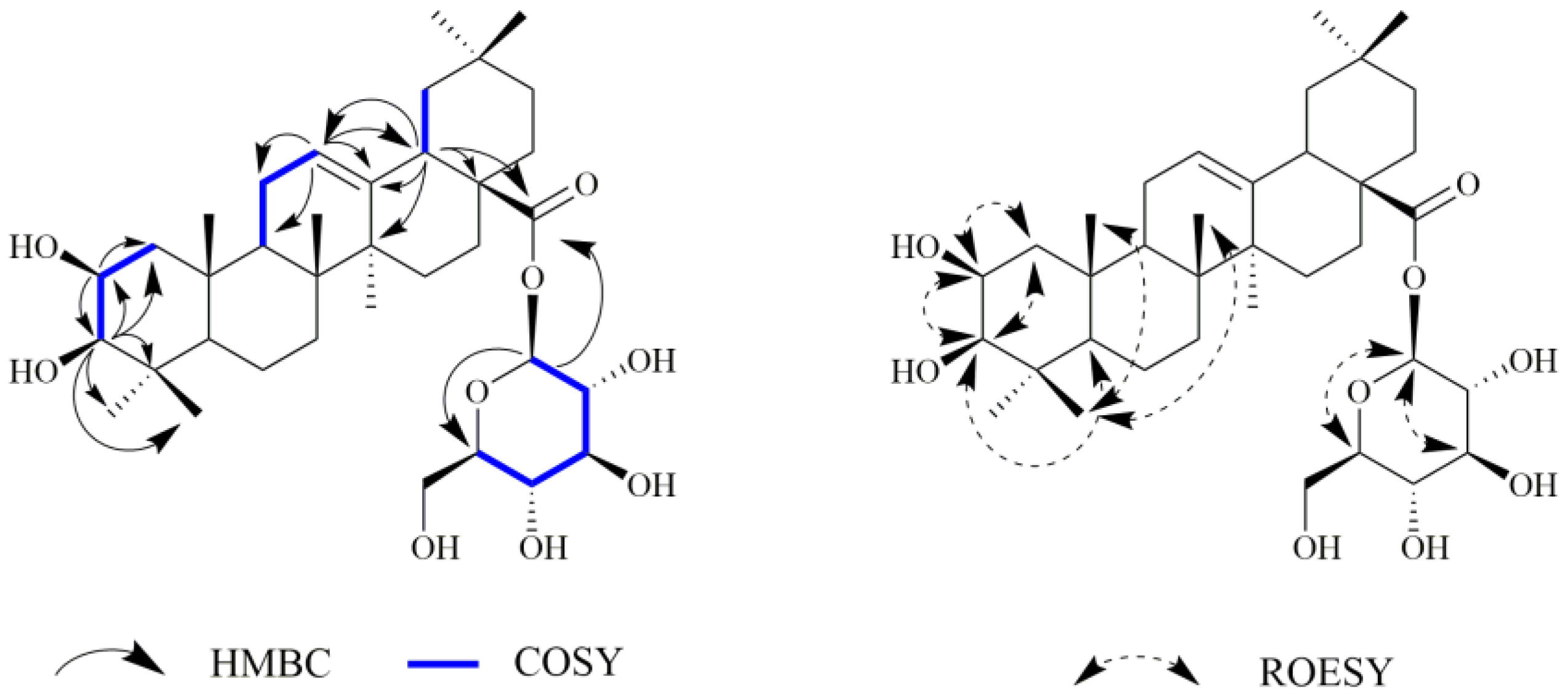
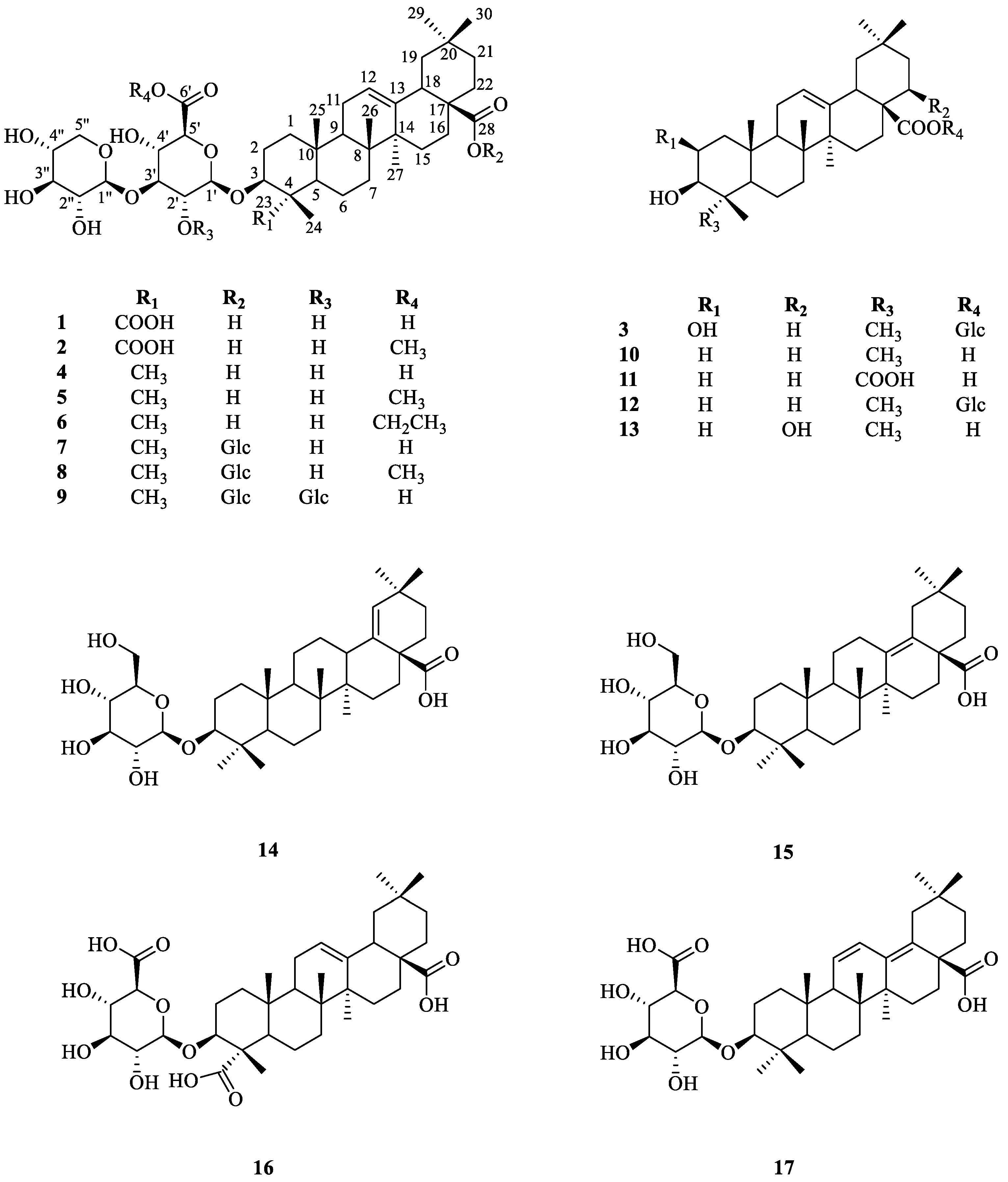
2.1. Chemistry
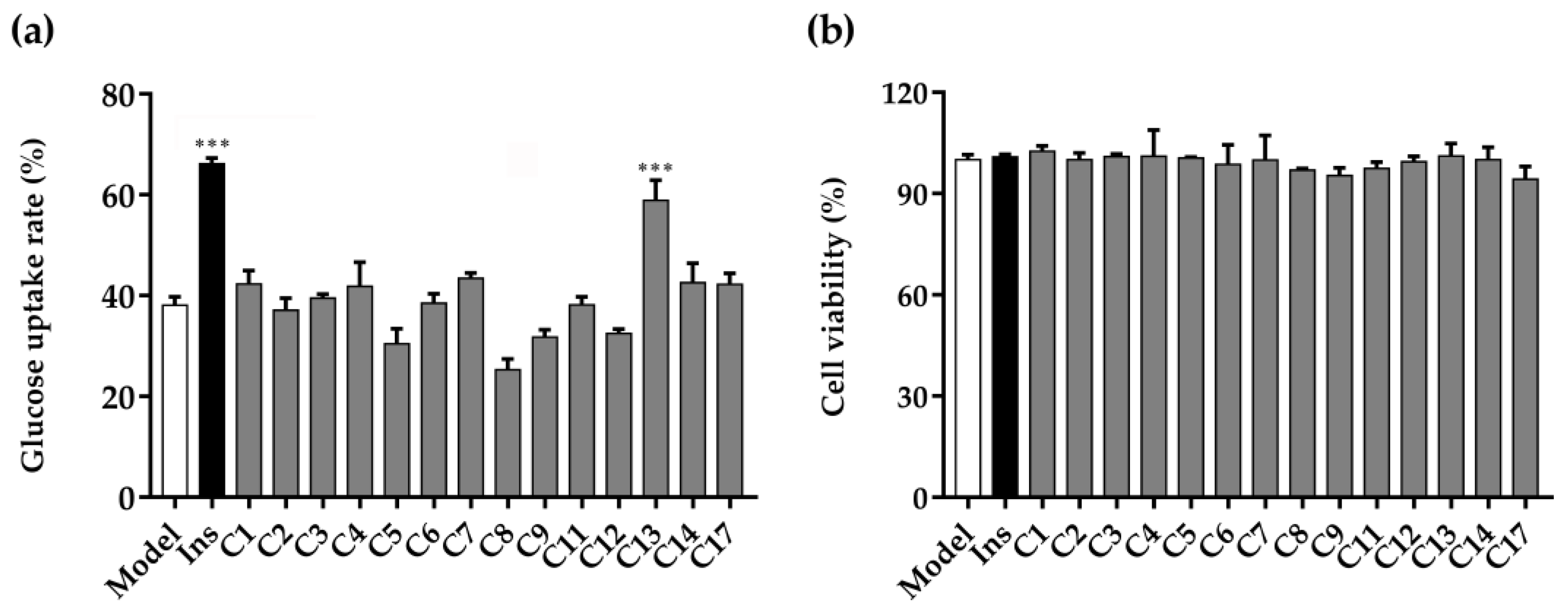
2.2. Glucose Uptake and Cell Viability
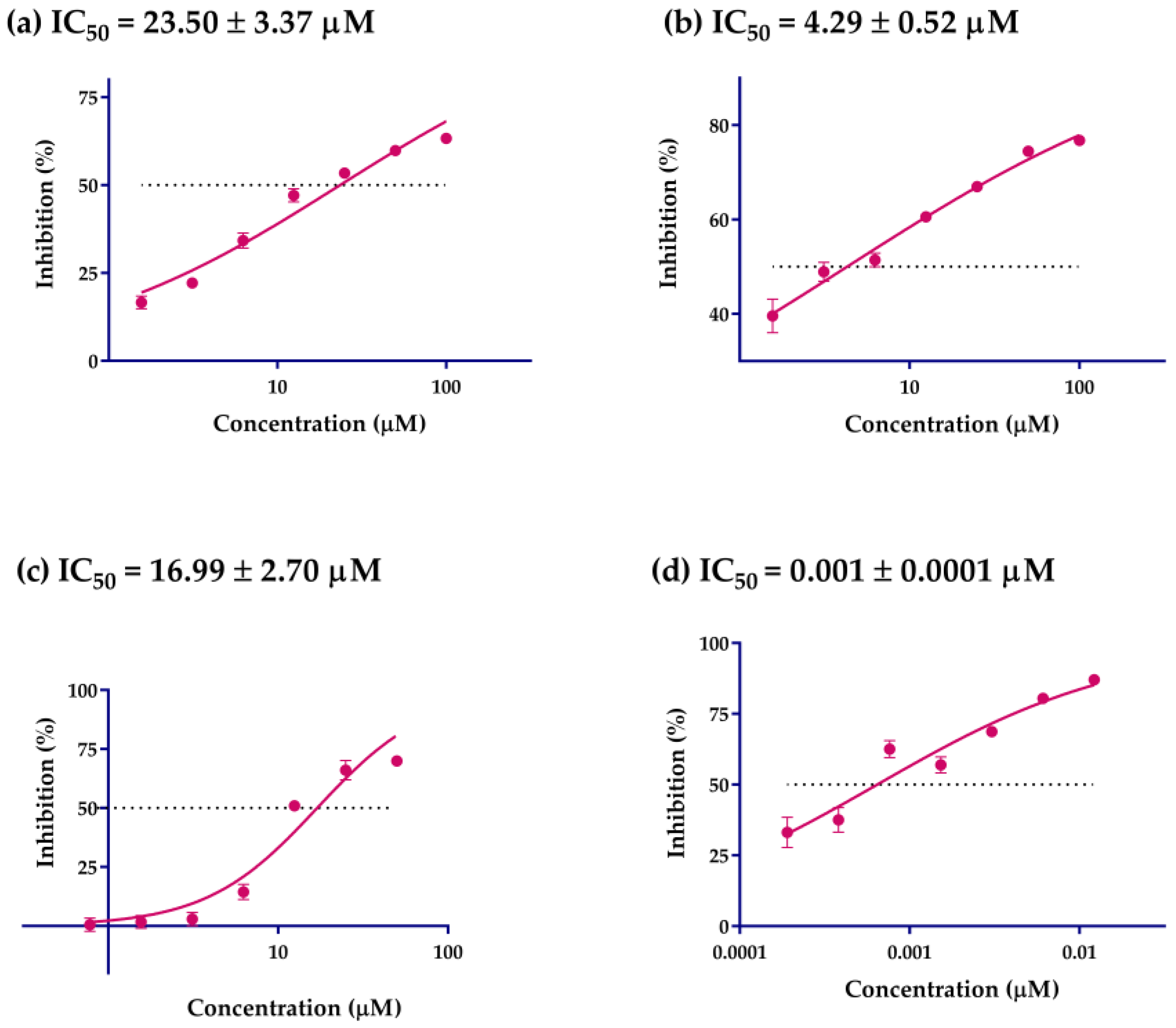
2.3. α-Glucosidase Inhibition Activity
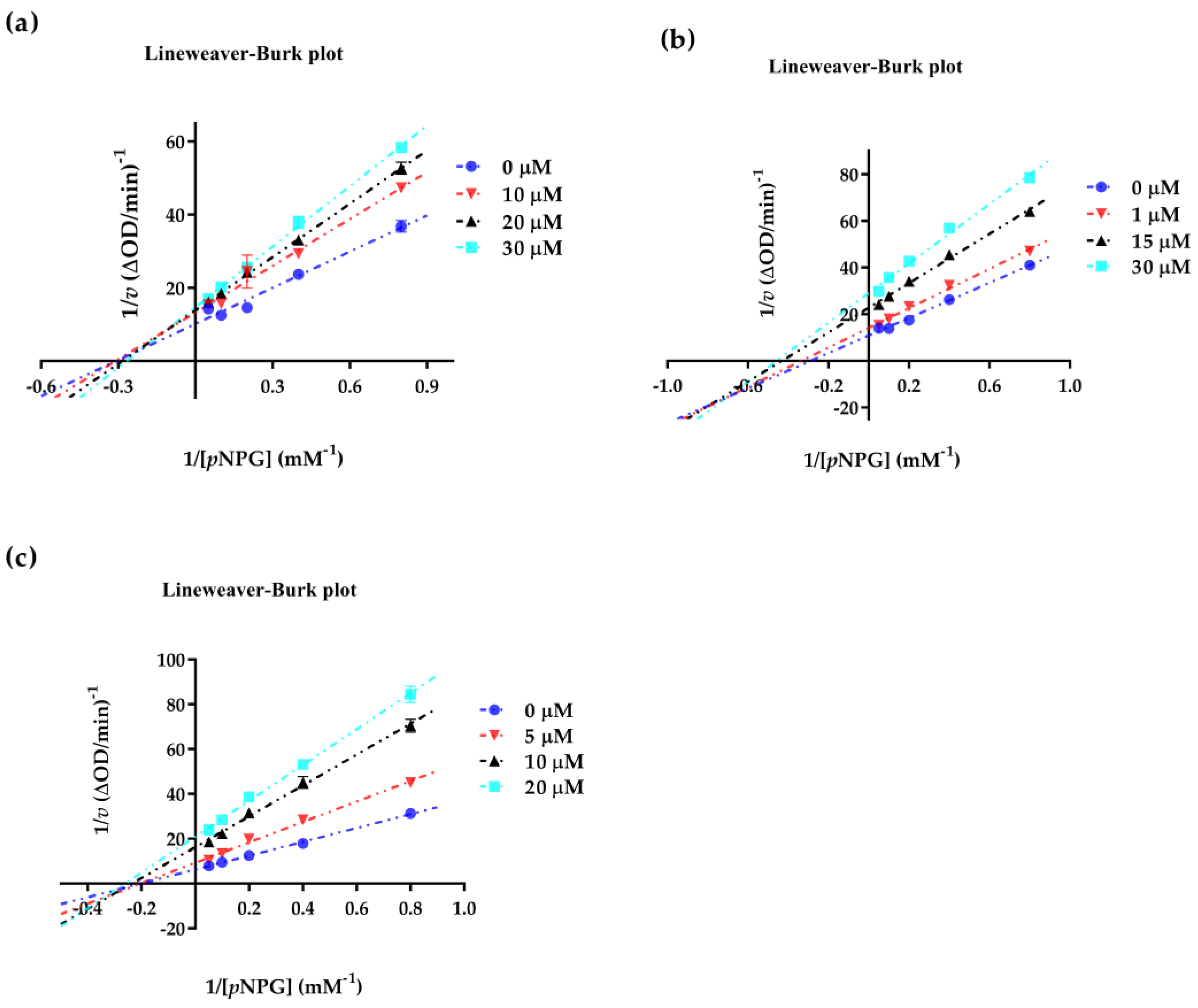
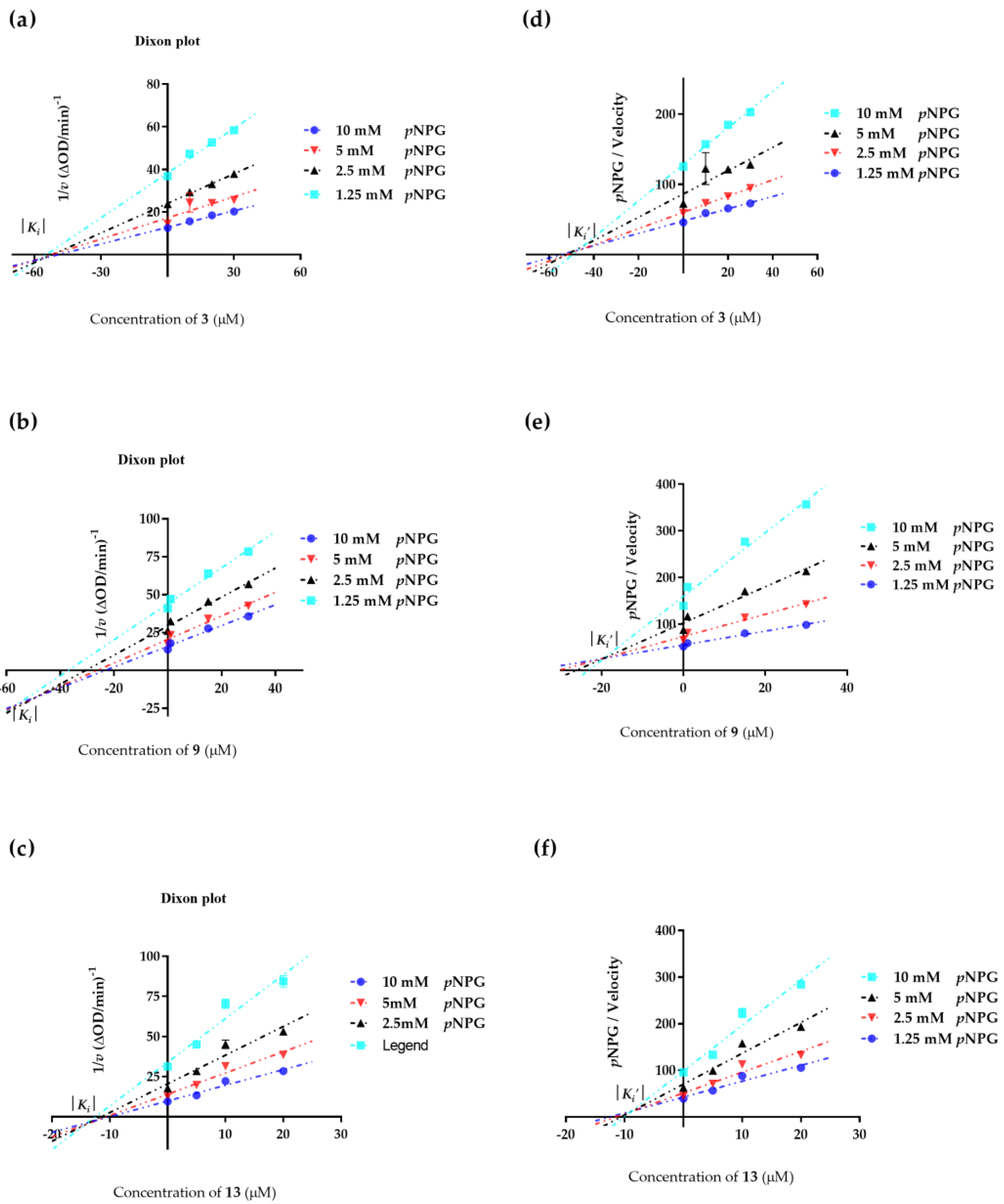
2.4. Enzyme Kinetic Equation
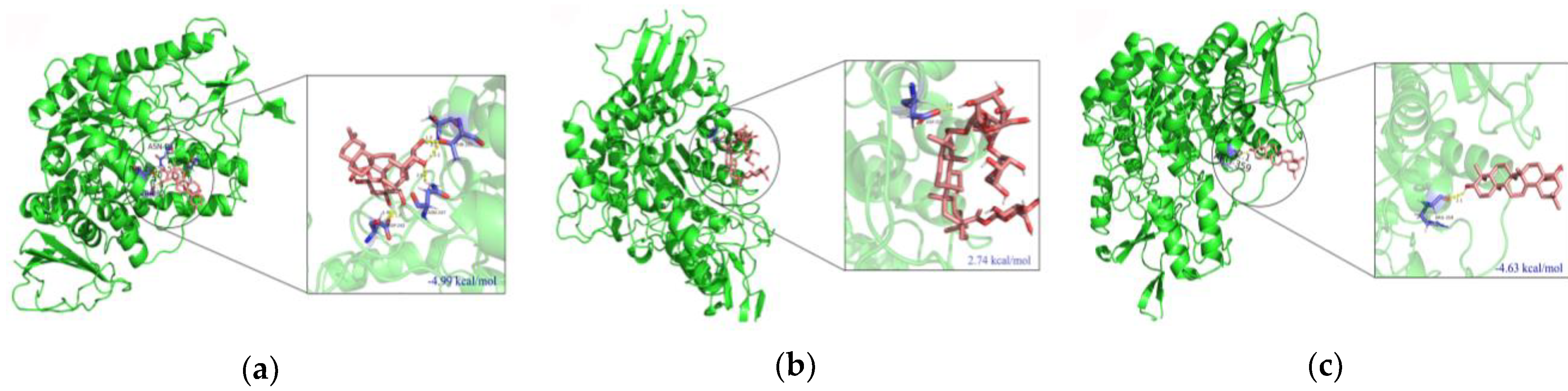
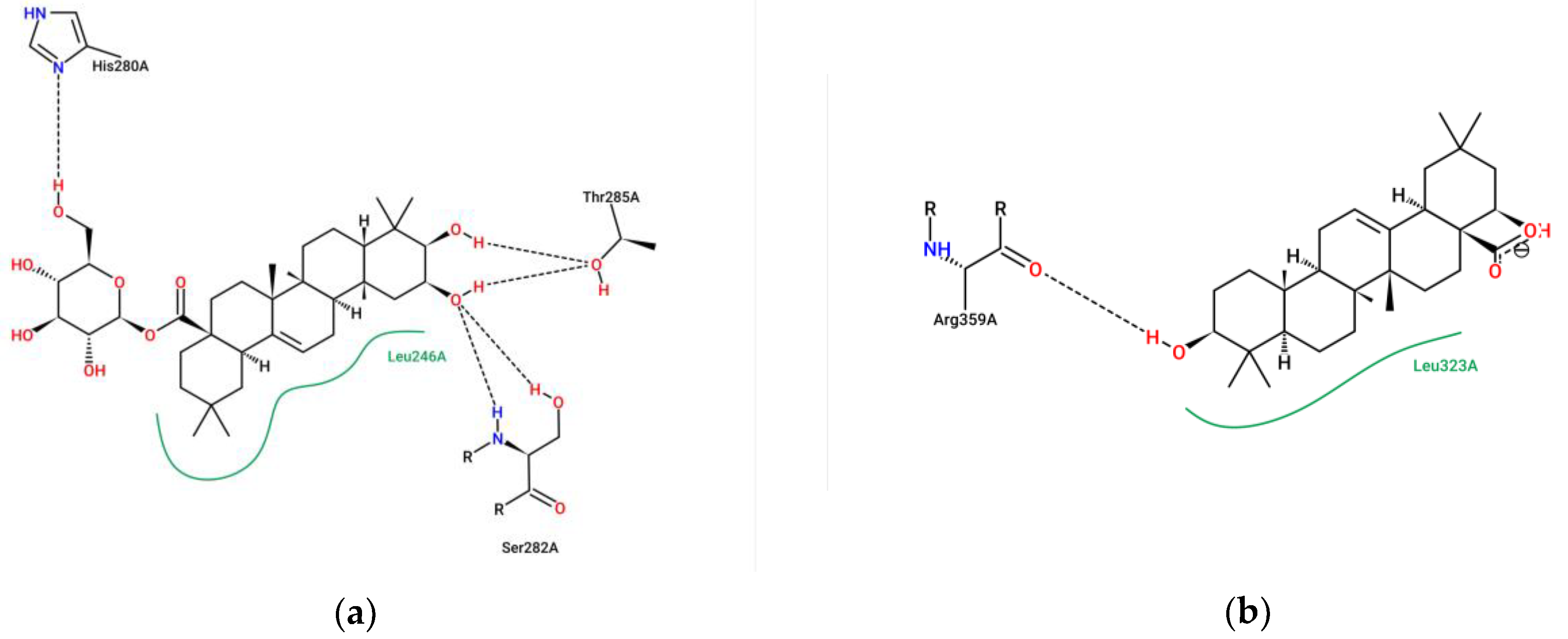
2.5. Molecular Docking
3. Materials and Methods
3.1. Chemicals, Reagents and Cell
3.2. Plant Materials
3.3. Extraction and Isolation
3.3.1. 3(β)-3-O-β-D-xylopyranosly(1→3)-β-D-glucopyranosiduronic acid-12-en-23,28-dioic acid (1)
3.3.2. 3(β)-3-O-β-D-xylopyranosly(1→3)-β-D-glucopyranosiduronic acid 6-methyl ester-12-en-23,28-dioic acid (2)
3.3.3. 2β,3β-dihydroxyolean-12-en-28-oic acid 28-O-β-D-glucopyranoside (3)
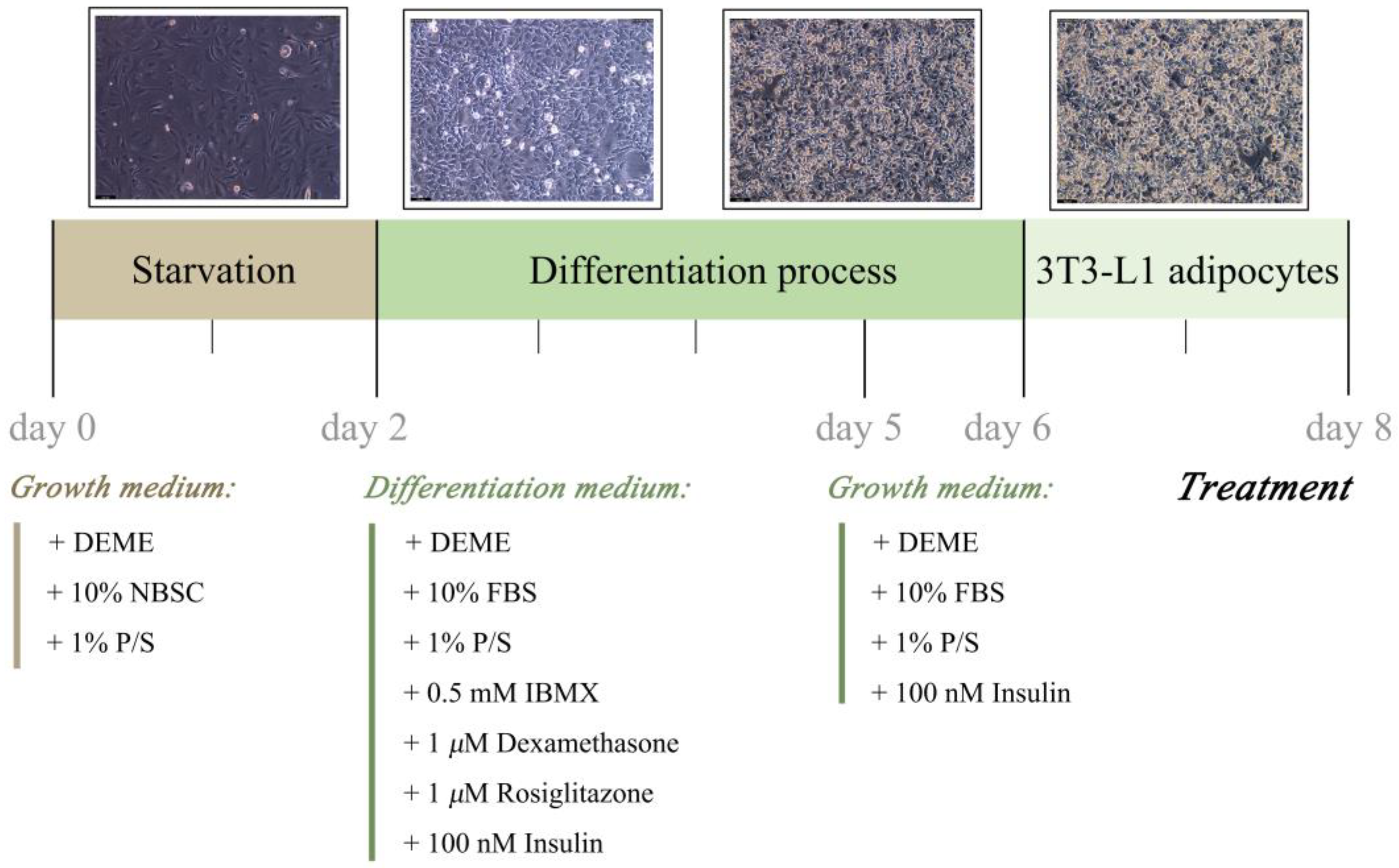
3.4. 3T3-L1 Preadipocytes Culture and Differentiation
3.5. Glucose Uptake and Cell Viability Assays
3.6. In Vitro α-Glucosidase Inhibition Assay
3.7. Kinetics Involved in the Inhibition of α-Glucosidase
3.8. Molecular Docking
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, D.; Tian, W.N.; An, M.Q.; Chen, Y.L.; Zeng, W.S.; Zhu, S.Y.; Li, P.; Du, B. Characterization of antidiabetic effects of Dendrobium officinale derivatives in a mouse model of type 2 diabetes mellitus. Food Chem. 2023, 399, 133974. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Tello, J.D.C.; Moretta, D.E.S.; HernÁndez, G.J.V.; Medina, L.S.H. Type 2 diabetes mellitus and management of isolated hyperglycemia: A review. Innovare J. Health Sci. 2022, 10, 1–4. [Google Scholar] [CrossRef]
- Solis-Herrera, C.; Triplitt, C.; Cersosimo, E.; DeFronzo, R.A. Site of Insulin Resistance in Type 2 Diabetes. Pathogenesis of Type 2 Diabetes Mellitus, 3rd ed.; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; Endotext [Internet]: South Dartmouth, MA, USA, 2021. [Google Scholar]
- Tomasik, P.; Horton, D. Enzymatic conversions of starch. Adv. Carbohydr. Chem. Biochem. 2012, 68, 59–436. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Yu, J.S.; Lee, H.S.; Choi, C.I.; Kim, K.H. Antidiabetic flavonoids from fruits of Morus alba promoting insulin-stimulated glucose uptake via Akt and AMP-activated protein kinase activation in 3T3-L1 adipocytes. Pharmaceutics 2021, 13, 526. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.K.; Hao, H.; Bian, Y.; Ge, Y.X.; Lu, S.; Xie, H.X.; Wang, K.M.; Tao, H.; Yuan, C.; Zhang, J.; et al. Discovery of new α-glucosidase inhibitors: Structure-based virtual screening and biological evaluation. Front. Chem. 2021, 9, 639279. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, D.; Srinivasan, H.; Patil, M.S.; Seetarama, S.; Agrawal, S.K.; Dixit, M.N.; Dhar, K. Novel method to differentiate 3T3-L1 cells in vitro to produce highly sensitive adipocytes for a GLUT4 mediated glucose uptake using fluorescent glucose analog. J. Cell. Commun. Signal. 2013, 7, 129–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Shin, E.J.; Kim, E.D.; Bayaraa, T.; Frost, S.C.; Hyun, C.K. Berberine activates GLUT1-mediated glucose uptake in 3T3-L1 adipocytes. Biol. Pharm. Bull. 2007, 30, 2120–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyni, G.L.; Kavitha, S.; Indu, S.; Arya, A.D.; Anusree, S.S.; Vineetha, V.P.; Vandana, S.; Sundaresan, A.; Raghu, K.G. Chebulagic acid from Terminalia chebula enhances insulin mediated glucose uptake in 3T3-L1 adipocytes via PPARγ signaling pathway. Biofactors 2014, 40, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Hwang, K.Y.; Oh, K.H.; Kim, Y.H.; Lee, J.Y.; Kim, K. Discovery of novel α-glucosidase inhibitors based on the virtual screening with the homology-modeled protein structure. Bioorg. Med. Chem. 2008, 16, 284–292. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.Y.; Jin, C.H. Mechanistic investigation of anthocyanidin derivatives as α-glucosidase inhibitors. Bioorg. Chem. 2019, 87, 803–809. [Google Scholar] [CrossRef]
- Li, D.Q.; Qian, Z.M.; Li, S.P. Inhibition of three selected beverage extracts on α-glucosidase and rapid identification of their active compounds using HPLC-DAD-MS/MS and biochemical detection. J. Agric. Food Chem. 2010, 58, 6608–6613. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Dai, Y.; Shimada, H.; Morikawa, T.; Matsumura, N.; Yoshizumi, S.; Matsuda, H.; Matsuda, H.; Kubo, M. Studies on Kochiae Fructus. II. On the saponin constituents from the fruit of chinese Kochia scoparia (Chenopodiaceae): Chemical structures of kochianosides I, II, III, and IV. Chem. Pharm. Bull. 1997, 45, 1052–1055. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Li, Y.; Murakami, T.; Matsumura, N.; Yamahara, J.; Yoshikawa, M. Antidiabetic principles of natural medicines. III. Structure-related inhibitory activity and action mode of oleanolic acid glycosides on hypoglycemic activity. Chem. Pharm. Bull. 1998, 46, 1399–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.H.; Huang, H.; Zhang, N.; Kong, D.Y.; Hua, M.L. Studies on the flavone glycosides from Fructus Kochiae. J. Asian Nat. Prod. Res. 2014, 16, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.H.; Xu, X.D.; Fu, H.W.; Chen, B.; Tian, J.K.; Zhang, L. Study on chemical constituents of Kochia scoparia. Chin. Pharm. J. 2012, 47, 338–342. [Google Scholar]
- Houlihan, A.J.; Conlin, P.; Chee-Sanford, J.C. Water-soluble exudates from seeds of Kochia scoparia exhibit antifungal activity against Colletotrichum graminicola. PLoS ONE 2019, 14, e0218104. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, Q.; Cai, J.; Yang, S.B. Supercritical CO2 extraction and GC-MS analysis of Kochia Fructus oil. Zhong Yao Cai 2003, 7, 494. [Google Scholar] [CrossRef]
- Zou, W.; Tang, Z.; Long, Y.; Xiao, Z.Q.; Ouyang, B.; Liu, M.H. Kochiae Fructus, the fruit of common Potherb Kochia scoparia (L.) Schrad: A review on phytochemistry, pharmacology, toxicology, quality control, and pharmacokinetics. Evid. Based Complement. Alternat. Med. 2021, 2021, 5382684. [Google Scholar] [CrossRef] [PubMed]
- National Pharmacopoeia Commission. The Pharmacopoeia of the People’s Republic of China, 1st ed.; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]
- Jo, S.; Ryu, J.; Han, H.Y.; Lee, G.; Ryu, M.H.; Kim, H. Anti-inflammatory activity of Kochia scoparia fruit on contact dermatitis in mice. Mol. Med. Rep. 2016, 13, 1695–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Xia, Y.F.; Lin, S.L. Studies on the hypoglycemic mechanism of n-butanol fraction from Fructus Kochiae. Pharmacol. Clin. Chin. Mater. Med. 2003, 19, 21–24. [Google Scholar]
- Yan, M.R.; Zhang, W.B.; Luo, G.P.; Wang, X.N.; Feng, S.M.; Zhao, N. Screening chemical constituents with antioxidative activity from the Kochiae Fructus. Chin. Med. J. Res. Pract. 2019, 33, 29–32. [Google Scholar]
- Wang, J.; Han, Y.; Wang, M.; Zhao, Q.Q.; Chen, X.F.; Liu, X.B. Natural triterpenoid saponin momordin Ic suppresses HepG2 cell invasion via COX-2 inhibition and PPARγ activation. Toxicol. In Vitro 2020, 65, 104784. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.N.; Liu, Q.Y.; Yao, C.H.; Chen, S.J.; Song, L.Y.; Guan, P.; Shao, L.J.; She, Y.Y.; Chi, H.Y. Normative research on bacteriostasis and relieving itching external therapeutic function of Kochiae Fructus. Zhong Yao Cai 2012, 35, 1974–1977. [Google Scholar] [CrossRef]
- Kubo, M.; Matsuda, H.; Dai, Y.; Ido, Y.; Yoshikawa, M. Studies on Kochiae Fructus. I. Antipruritogenic effect of 70% ethanol extract from kochiae fructus and its active component. Yakugaku Zasshi 1997, 117, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, H.; Dai, Y.; Ido, Y.; Yoshikawa, M.; Kubo, M. Studies on Kochiae Fructus. IV. Anti-allergic effects of 70% ethanol extract and its component, momordin Ic from dried fruits of Kochia scoparia L. Biol. Pharm. Bull. 1997, 20, 1165–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Xia, Y.F.; Chen, H.B.; Matsuda, H.; Kubo, M. Inhibition of immediate and delayed type hypersensitivity by the 70% ethanolic extract from Fructus kochiae. Chin. J. Mod. Appl. Pharm. 2001, 1, 8–9. [Google Scholar] [CrossRef]
- Kim, N.Y.; Lee, M.K.; Park, M.J.; Kim, S.J.; Park, H.J.; Choi, J.; Kim, S.H.; Cho, S.Y.; Lee, J. Momordin Ic and oleanolic acid from Kochiae Fructus reduce carbon tetrachloride-induced hepatotoxicity in rats. J. Med. Food 2005, 8, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Dai, Y.; Ido, Y.; Murakami, T.; Matsuda, H.; Yoshikawa, M.; Kubo, M. Studies on Kochiae Fructus. V. Antipruritic effects of oleanolic acid glycosides and the structure-requirement. Biol. Pharm. Bull. 1998, 21, 1231–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amimoto, K.; Yoshikawa, K.; Arihara, S. Triterpenoid saponins of Aquifoliaceous plants. VIII. Ilexosides XXIX-XXXII from the leaves of Ilex rotunda THUNB. Chem. Pharm. Bull. 1992, 40, 3138–3141. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Chen, Y.; Cui, Z.; Li, J.; Wang, Z. Triterpenoid glycosides from the fruits of Kochia scoparia. Planta Med. 1995, 61, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Han, L.K.; Nose, R.; Li, W.; Gong, X.J.; Zheng, Y.N.; Yoshikawa, M.; Koike, K.; Nikaido, T.; Okuda, H.; Kimura, Y. Reduction of fat storage in mice fed a high-fat diet long term by treatment with saponins prepared from Kochia scoparia fruit. Phytother. Res. 2006, 20, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, N.; Watanabe, H.; Oshio, H. Saponins from roots of Momordica cochinchinensis. Phytochemistry 1988, 27, 3585–3591. [Google Scholar] [CrossRef]
- Hossain, M.A.; Ismail, Z. Isolation and characterization of triterpenes from the leaves of Orthosiphon stamineus. Arab. J. Chem. 2013, 6, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.X.; Zhang, H.C.; Wang, H.Y.; Wang, Y.; Wang, F.L.; Sun, J.Y. Two new triterpenoids from Gypsophila oldhamiana. Nat. Prod. Res. 2016, 30, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Suthar, S.K.; Hooda, A.; Sharma, A.; Bansal, S.; Monga, J.; Chauhan, M.; Sharma, M. Isolation optimisation, synthesis, molecular docking and in silico ADMET studies of lantadene a and its derivatives. Nat. Prod. Res. 2021, 35, 3939–3944. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Xie, L.; Liu, B.R.; Yu, L.X. Triterpenoids from the fruits of Forsythia suspensa. Chin. J. Nat. Med. 2010, 8, 414–418. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.B.; Hu, K.W.; Xu, Q.L.; Wen, X.A.; Cheng, K.G.; Chen, C.P.; Yuan, H.L.; Dai, L.; Sun, H.B. Synthesis and anti-inflammatory activity of saponin derivatives of δ-oleanolic acid. Eur. J. Med. Chem. 2021, 209, 112932. [Google Scholar] [CrossRef] [PubMed]
- Borel, C.; Gupta, M.P.; Hostettmann, K. Molluscicidal saponins from Swartzia simplex. Phytochemistry 1987, 26, 2685–2689. [Google Scholar] [CrossRef]
- Kuroda, M.; Aoshima, T.; Haraguchi, M.; Young, M.C.M.; Sakagami, H.; Mimaki, Y. New oleanane glycosides from the roots of Gomphrena macrocephala. Nat. Prod. Commun. 2006, 1, 431–439. [Google Scholar] [CrossRef]
- Lahrita, L.; Kato, E.; Kawabata, J. Uncovering potential of indonesian medicinal plants on glucose uptake enhancement and lipid suppression in 3T3-L1 adipocytes. J. Ethnopharmacol. 2015, 168, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalra, S. Alpha glucosidase inhibitors. J. Pak. Med. Assoc. 2014, 64, 474–476. [Google Scholar] [PubMed]
- Huang, L.R.; Luo, H.; Li, Q.J.; Wang, D.P.; Zhang, J.X.; Hao, X.J.; Yang, X.S. Pentacyclic triterpene derivatives possessing polyhydroxyl ring A inhibit gram-positive bacteria growth by regulating metabolism and virulence genes expression. Eur. J. Med. Chem. 2015, 95, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, L.; Chen, W.H.; Park, H.; Ke, Z.F.; Wang, B. Binding mechanism and synergetic effects of xanthone derivatives as noncompetitive α-glucosidase inhibitors: A theoretical and experimental study. J. Phys. Chem. B 2013, 117, 13464–13471. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Zhang, G.W.; Liao, Y.J.; Gong, D.M. Inhibitory mechanism of morin on α-glucosidase and its anti-glycation properties. Food Funct. 2016, 7, 3953–3963. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Mukherjee, A.; Goswami, R.; Basu, S. Hypoglycemic activity of the antioxidant saponarin, characterized as α-glucosidase inhibitor present in Tinospora cordifolia. J. Enzyme Inhib. Med. Chem. 2009, 24, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Lim-Wilby, M. Molecular docking. Methods Mol. Biol. 2008, 443, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Addepalli, R.; Netzel, M.E.; Tinggi, U.; Fletcher, M.T.; Sultanbawa, Y.; Osborne, S.A. Antioxidant-rich extracts of Terminalia ferdinandiana interfere with estimation of cell viability. Antioxidants 2019, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Chen, X.L.; Zhao, X.; Ni, J.Y.; Wang, H.L.; Han, M.; Zhang, Y.M. Antidiabetic potential of Catechu via assays for α-glucosidase, α-amylase, and glucose uptake in adipocytes. J. Ethnopharmacol. 2022, 291, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Miyake, H.; Kusunoki, M.; Osaki, S. Crystal structures of isomaltase from Saccharomyces cerevisiae and in complex with its competitive inhibitor maltose. FEBS J. 2010, 277, 4205–4214. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| δH, (J in Hz) | δC | δH, (J in Hz) | δC | δH, (J in Hz) | δC | |
| 1 | 1.67, overlapped | 39.7 | 1.66, overlapped | 39.7 | 1.92, m | 48.3 |
| 1.08, overlapped | 1.08, overlapped | 0.88, m | ||||
| 2 | 1.87, m | 26.6 | 1.83, m | 26.6 | 3.62, m | 69.6 |
| 1.68, m | 1.67, m | |||||
| 3 | 4.05, dd (11.5, 4.0) | 86.6 | 4.05, dd (11.5, 4.5) | 86.6 | 2.90, d (9.5) | 84.6 |
| 4 | 54.0 | 54.1 | 40.7 | |||
| 5 | 1.49, m | 53.1 | 1.49, m | 53.1 | 0.84, m | 56.9 |
| 6 | 2.00, td (13.5, 3.5) | 24.2 | 2.01, td (13.5, 4.0) | 24.2 | 2.04, td (13.5, 3.5) | 24.1 |
| 1.59, m | 1.60, m | 1.70, m | ||||
| 7 | 1.53, overlapped | 33.7 | 1.53, overlapped | 33.7 | 1.56, overlapped | 34.0 |
| 1.53, overlapped | 1.53, overlapped | 1.56, overlapped | ||||
| 8 | 41.0 | 41.0 | 40.9 | |||
| 9 | 1.65, overlapped | 49.1 | 1.64, overlapped | 49.1 | 1.62, overlapped | 49.2 |
| 10 | 37.6 | 37.6 | 39.4 | |||
| 11 | 1.92, m | 24.6 | 1.93, m | 24.6 | 1.92, m | 24.8 |
| 1.90, m | 1.91, m | 1.90, m | ||||
| 12 | 5.23, brs | 123.6 | 5.24, brs | 123.6 | 5.27, brs | 123.8 |
| 13 | 145.3 | 145.6 | 145.2 | |||
| 14 | 43.0 | 43.1 | 43.1 | |||
| 15 | 1.76, overlapped | 29.0 | 1.76, overlapped | 29.0 | 1.79, overlapped | 29.0 |
| 1.05, overlapped | 1.05, overlapped | 1.05, overlapped | ||||
| 16 | 1.57, overlapped | 21.9 | 1.57, overlapped | 21.9 | 1.52, overlapped | 19.7 |
| 1.57, overlapped | 1.57, overlapped | 1.52, overlapped | ||||
| 17 | 47.8 | 47.8 | 48.2 | |||
| 18 | 2.84, dd (13.5, 4.0) | 42.9 | 2.85, dd (14.0, 4.0) | 42.9 | 2.86, dd (14.0, 4.0) | 42.7 |
| 19 | 1.71, m | 47.4 | 1.69, m | 47.4 | 1.70, m | 47.4 |
| 1.11, m | 1.11, m | 1.13, m | ||||
| 20 | 31.8 | 31.8 | 31.7 | |||
| 21 | 1.38, td (13.5, 3.5) | 35.0 | 1.39, td (13.5, 4.0) | 35.0 | 1.39, m | 35.0 |
| 1.21, m | 1.22, m | 1.21, m | ||||
| 22 | 1.73, overlapped | 33.9 | 1.72, overlapped | 34.0 | 1.72, overlapped | 33.3 |
| 1.27, overlapped | 1.27, overlapped | 1.32, overlapped | ||||
| 23 | 181.5 | 181.7 | 1.01, s | 29.4 | ||
| 24 | 1.14, s | 12.2 | 1.14, s | 12.4 | 0.80, s | 17.9 |
| 25 | 0.96, s | 16.4 | 0.96, s | 16.4 | 1.01, s | 17.3 |
| 26 | 0.80, s | 17.7 | 0.81, s | 17.7 | 0.80, s | 17.6 |
| 27 | 1.16, s | 26.4 | 1.17, s | 26.5 | 1.16, s | 26.5 |
| 28 | 181.9 | 182.0 | 178.2 | |||
| 29 | 0.90, s | 33.7 | 0.91, s | 33.7 | 0.91, s | 33.6 |
| 30 | 0.93, s | 24.1 | 0.94, s | 24.1 | 0.94, s | 24.1 |
| No. | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| δH, (J in Hz) | δC | δH, (J in Hz) | δC | δH, (J in Hz) | δC | |
| GlcA-at-C-3 | Glc-at-C-28 | |||||
| 1′ | 4.37, d (7.5) | 105.5 | 4.39, d (7.5) | 105.5 | 5.38, d (8.0) | 95.9 |
| 2′ | 3.35, d (7.5) | 74.6 | 3.34, d (7.5) | 74.6 | 3.31, m | 74.1 |
| 3′ | 3.50, m | 86.3 | 3.52, m | 86.1 | 3.40, m | 78.5 |
| 4′ | 3.56, m | 71.8 | 3.57, m | 71.7 | 3.34, m | 71.3 |
| 5′ | 3.76, m | 76.4 | 3.83, d (9.5) | 76.5 | 3.34, m | 78.9 |
| 6′ | 171.4 | 171.2 | 3.81, dd (12.0, 1.5) | 62.6 | ||
| 3.68, dt (12.0, 2.0) | ||||||
| 6′-OMe | 3.77, s | 53.0 | ||||
| Xyl→glc-C-3 | ||||||
| 1″ | 4.50, d (7.5) | 105.9 | 4.52, d (7.5) | 105.9 | ||
| 2″ | 3.24, m | 75.4 | 3.24, m | 75.4 | ||
| 3″ | 3.32, m | 77.7 | 3.32, m | 77.7 | ||
| 4″ | 3.47, m | 71.2 | 3.49, m | 71.2 | ||
| 5″ | 3.89, dd (11.5, 5.5) | 67.2 | 3.89, dd (11.5, 5.5) | 67.2 | ||
| 3.20, dd (11.5, 5.5) | 3.20, dd (11.5, 5.5) | |||||
| Compounds | Inhibitory Rate (%) | IC50 (μM) |
|---|---|---|
| Acarbose A | 87.04 ± 0.29 a | 0.001 ± 0.00 |
| 1 B | 13.71 ± 0.54 j | >50 |
| 2 B | 14.05 ± 4.90 j | >50 |
| 3 B | 59.79 ± 1.04 d | 23.50 ± 3.37 |
| 4 B | 26.28 ± 1.01 i | >50 |
| 5 B | 29.62 ± 2.26 i | >50 |
| 6 B | 35.00 ± 0.40 h | >50 |
| 7 B | 52.71 ± 0.85 f | >50 |
| 8 B | 57.02 ± 36.3 e | >50 |
| 9 B | 74.41 ± 1.02 b | 4.29 ± 0.52 |
| 11 B | 53.98 ± 1.65 e | >50 |
| 12 B | 41.48 ± 1.66 g | >50 |
| 13 B | 69.91 ± 0.59 c | 16.99 ± 2.70 |
| 14 B | 17.49 ± 4.45 j | >50 |
| 17 B | 55.68 ± 0.17 e | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.-L.; Zhang, K.; Zhao, X.; Wang, H.-L.; Han, M.; Li, R.; Zhang, Z.-N.; Zhang, Y.-M. Triterpenoids from Kochiae Fructus: Glucose Uptake in 3T3-L1 Adipocytes and α-Glucosidase Inhibition, In Silico Molecular Docking. Int. J. Mol. Sci. 2023, 24, 2454. https://doi.org/10.3390/ijms24032454
Chen X-L, Zhang K, Zhao X, Wang H-L, Han M, Li R, Zhang Z-N, Zhang Y-M. Triterpenoids from Kochiae Fructus: Glucose Uptake in 3T3-L1 Adipocytes and α-Glucosidase Inhibition, In Silico Molecular Docking. International Journal of Molecular Sciences. 2023; 24(3):2454. https://doi.org/10.3390/ijms24032454
Chicago/Turabian StyleChen, Xue-Lin, Kun Zhang, Xia Zhao, Han-Lei Wang, Mei Han, Ru Li, Zhen-Nan Zhang, and Yu-Mei Zhang. 2023. "Triterpenoids from Kochiae Fructus: Glucose Uptake in 3T3-L1 Adipocytes and α-Glucosidase Inhibition, In Silico Molecular Docking" International Journal of Molecular Sciences 24, no. 3: 2454. https://doi.org/10.3390/ijms24032454
APA StyleChen, X. -L., Zhang, K., Zhao, X., Wang, H. -L., Han, M., Li, R., Zhang, Z. -N., & Zhang, Y. -M. (2023). Triterpenoids from Kochiae Fructus: Glucose Uptake in 3T3-L1 Adipocytes and α-Glucosidase Inhibition, In Silico Molecular Docking. International Journal of Molecular Sciences, 24(3), 2454. https://doi.org/10.3390/ijms24032454