
Glucose 6-P Dehydrogenase Overexpression Improves Aging-Induced Endothelial Dysfunction in Aorta from Mice: Role of Arginase II

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
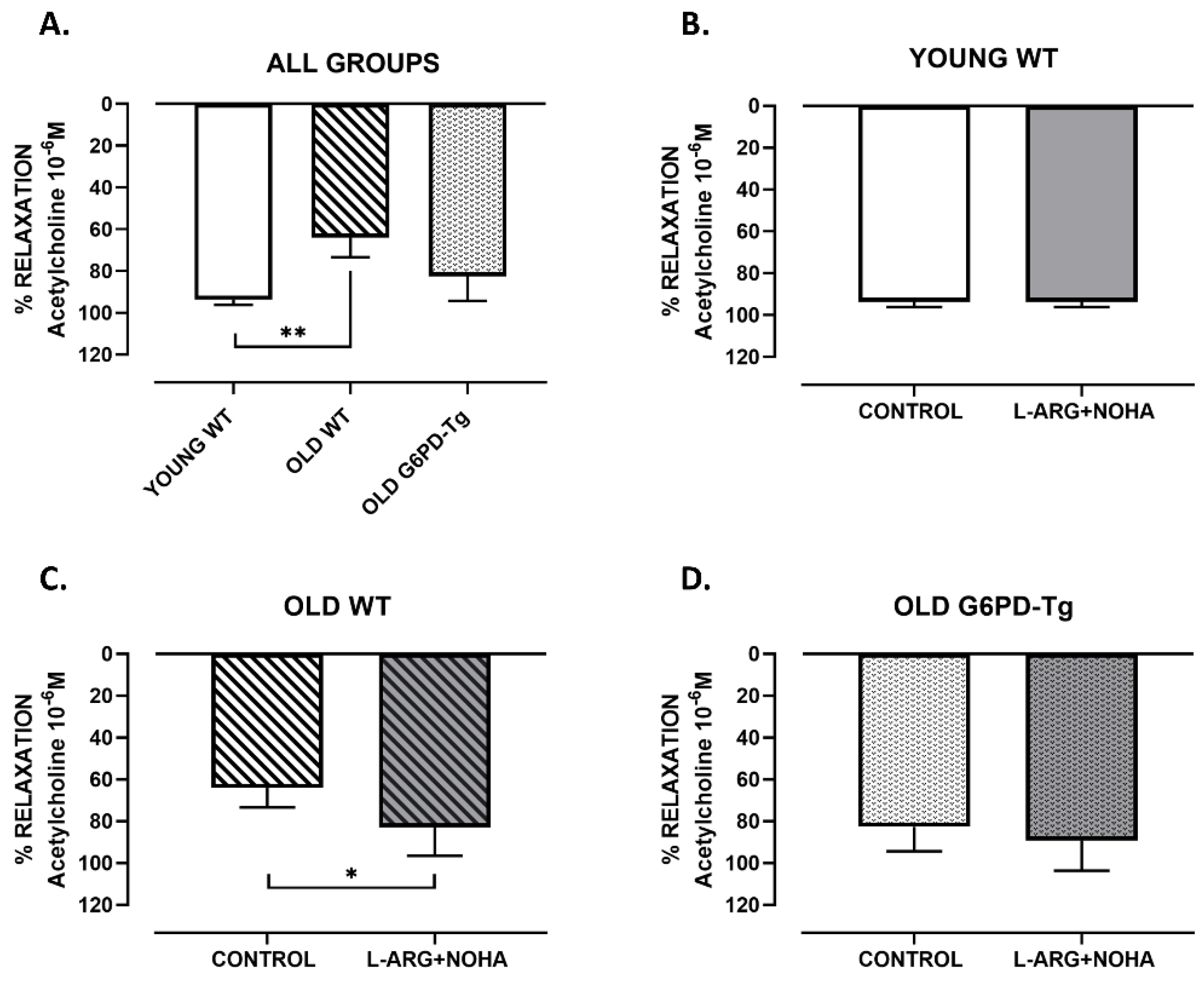
2.1. Vascular Reactivity Study
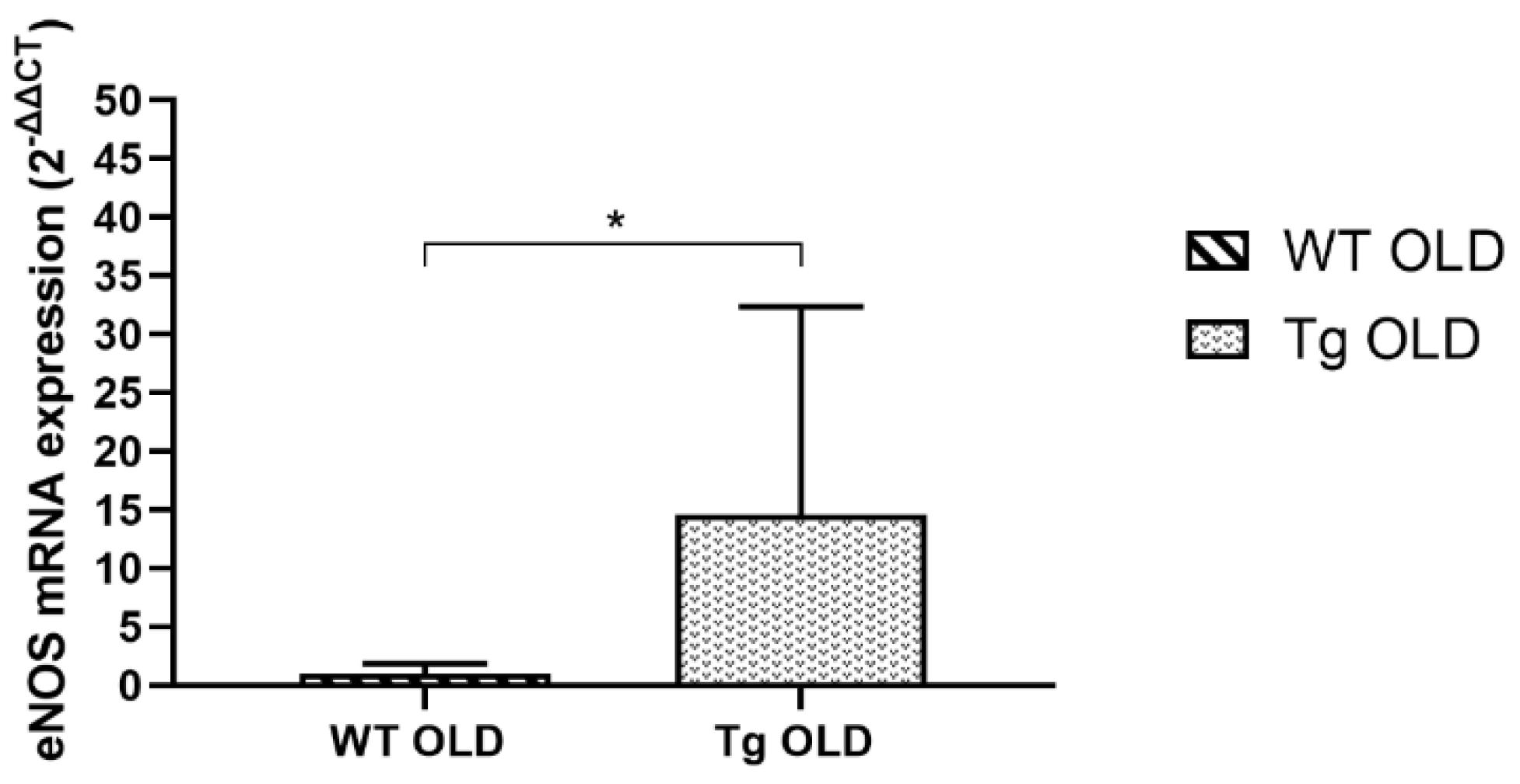
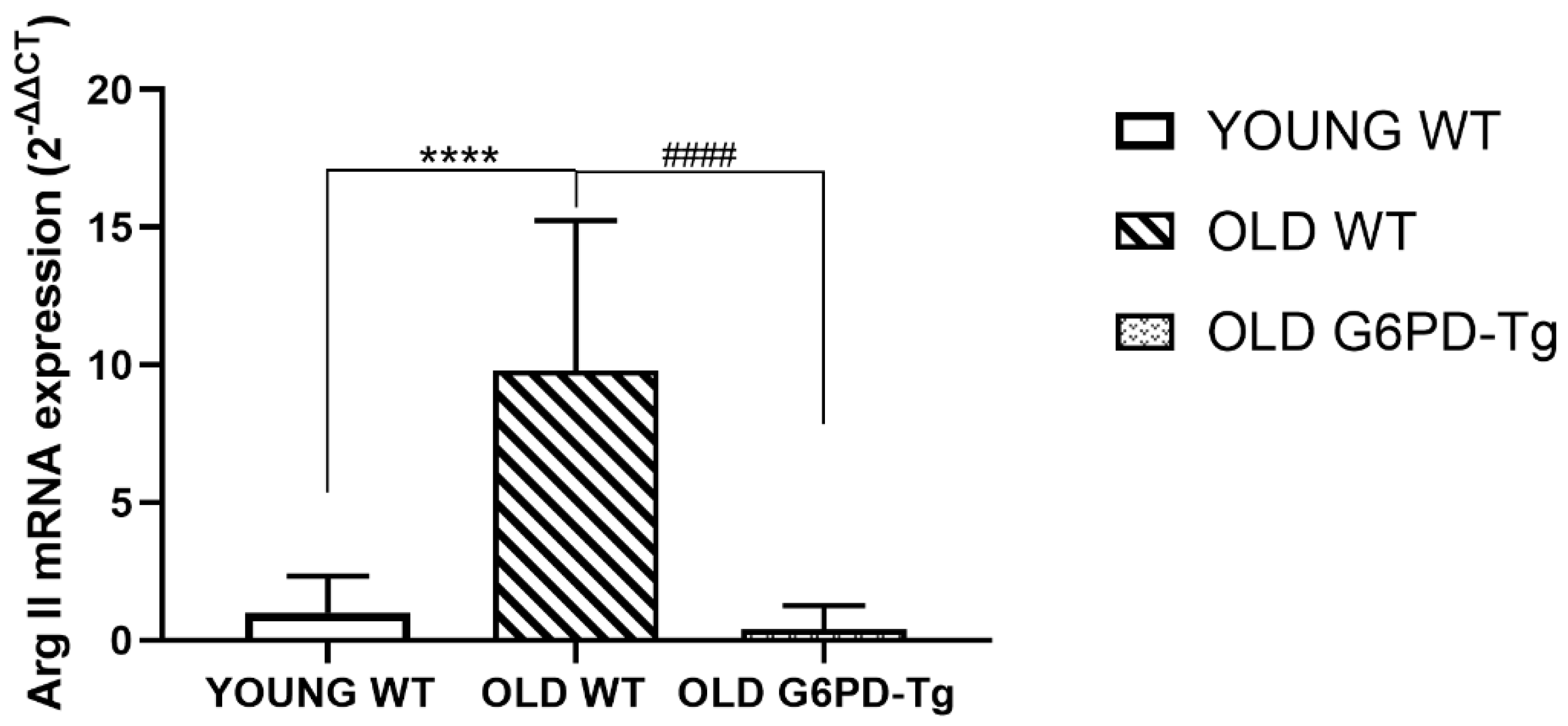
2.2. Gene Expression Analysis
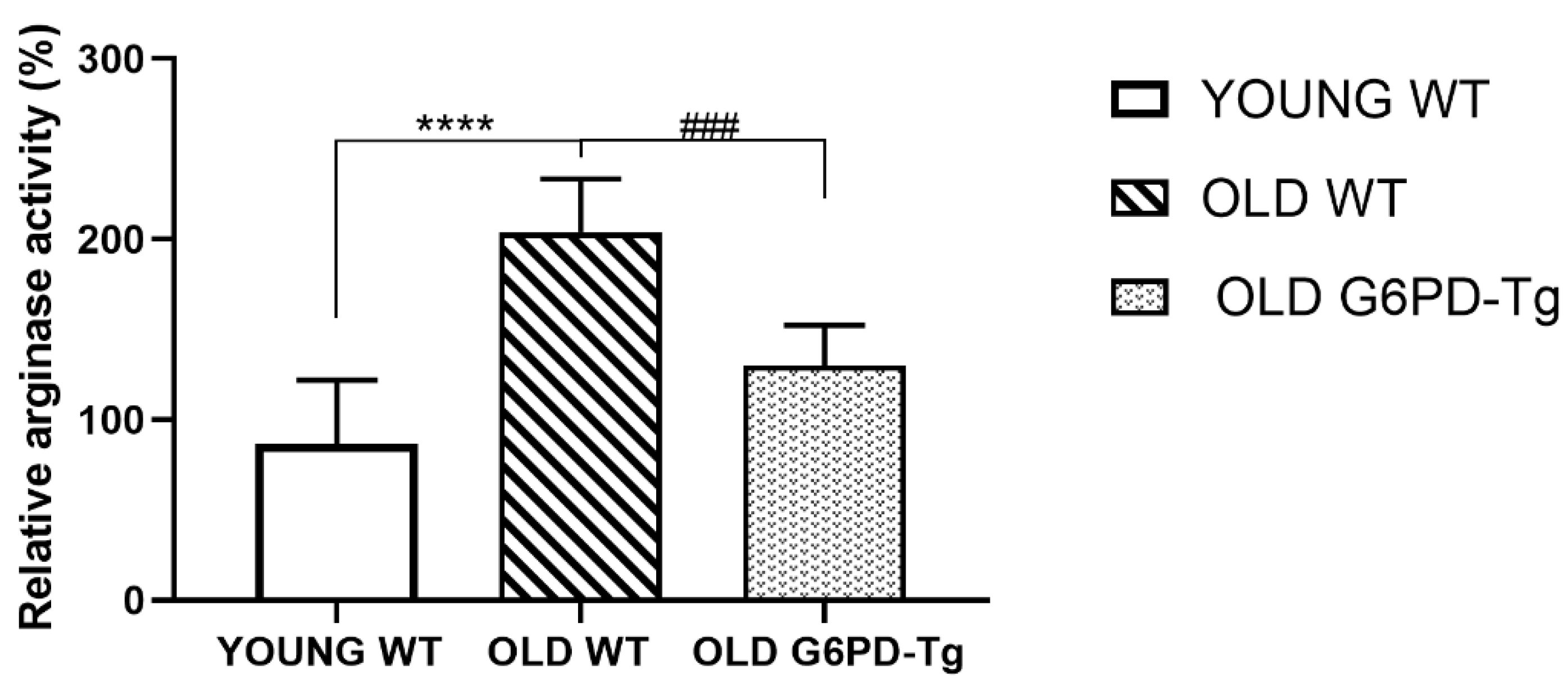
2.3. Arginase Activity
2.4. Histological Study: Arterial Wall Thickness and Collagen Deposition
3. Discussion
4. Materials and Methods
4.1. Animal Model and Study Design
4.2. Vascular Reactivity Studies
4.3. Total RNA Extraction and Real Time Polymerase Chain Reaction (RT-PCR) Studies
4.4. Arginase Activity Studies
4.5. Histological Study
4.6. Statistics
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Flatt, T. A new definition of aging? Front. Genet. 2012, 3, 148. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Park, D.C.; Yeo, S.G. Aging. Korean J. Audiol. 2013, 17, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Rebelo-Marques, A.; De Sousa Lages, A.; Andrade, R.; Ribeiro, C.F.; Mota-Pinto, A.; Carrilho, F.; Espregueira-Mendes, J. Aging hallmarks: The benefits of physical exercise. Front. Endocrinol. 2018, 9, 258. [Google Scholar] [CrossRef]
- Vijg, J.; Wei, J.Y. Understanding the biology of aging: The key to prevention and therapy. J. Am. Geriatr. Soc. 1995, 43, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Garatachea, N.; Pareja-Galeano, H.; Sanchis-Gomar, F.; Santos-Lozano, A.; Fiuza-Luces, C.; Morán, M.; Emanuele, E.; Joyner, M.J.; Lucia, A. Exercise attenuates the major hallmarks of aging. Rejuven. Res. 2015, 18, 57–89. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.S.; Singer, B.D.; Vaughan, D.E. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell 2017, 16, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Pitrez, P.R.; Aires, H.R.; Tomé, I.; Ferreira, R.S.; Ferreira, L. Physiological and pathological vascular aging. In Biophysical Regulation of Vascular Differentiation and Assembly; Gerecht, S., Ed.; Biological and Medical Physics, Biomedical Engineering; Springer International Publishing: Cham, Switzerland, 2018; pp. 51–72. ISBN 978-3-319-99319-5. [Google Scholar]
- Nilsson, P.M.; Boutouyrie, P.; Laurent, S. Vascular aging: A tale of EVA and ADAM in cardiovascular risk assessment and prevention. Hypertension 2009, 54, 3–10. [Google Scholar] [CrossRef]
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef]
- Hamczyk, M.R.; Nevado, R.M.; Barettino, A.; Fuster, V.; Andrés, V. Biological versus chronological aging: JACC focus seminar. J. Am. Coll. Cardiol. 2020, 75, 919–930. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Harrison, D.G.; Figueroa, C.A.; Lacolley, P.; Laurent, S. Central artery stiffness in hypertension and aging: A problem with cause and consequence. Circ. Res. 2016, 118, 379–381. [Google Scholar] [CrossRef]
- Elliott, R.J.; McGrath, L.T. Calcification of the human thoracic aorta during aging. Calcif. Tissue Int. 1994, 54, 268–273. [Google Scholar] [CrossRef]
- Lanzer, P.; Boehm, M.; Sorribas, V.; Thiriet, M.; Janzen, J.; Zeller, T.; St Hilaire, C.; Shanahan, C. Medial vascular calcification revisited: Review and perspectives. Eur. Heart J. 2014, 35, 1515–1525. [Google Scholar] [CrossRef]
- Cooke, J.P.; Dzau, J.; Creager, A. Endothelial dysfunction in hypercholesterolemia is corrected by L-arginine. Basic Res. Cardiol. 1991, 86 (Suppl. 2), 173–181. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.G. Cellular and molecular mechanisms of endothelial cell dysfunction. J. Clin. Investig. 1997, 100, 2153–2157. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Meininger, C.J. Arginine nutrition and cardiovascular function. J. Nutr. 2000, 130, 2626–2629. [Google Scholar] [CrossRef]
- Laina, A.; Stellos, K.; Stamatelopoulos, K. Vascular ageing: Underlying mechanisms and clinical implications. Exp. Gerontol. 2018, 109, 16–30. [Google Scholar] [CrossRef]
- Krebs, H.A.; Henseleit, K. Untersuchungen uber die Harnstoffbildung im Tierkörper. Hoppe-Seyler’S Z. Für Physiol. Chem. 1932, 210, 33–66. [Google Scholar] [CrossRef]
- Berkowitz, D.E.; White, R.; Li, D.; Minhas, K.M.; Cernetich, A.; Kim, S.; Burke, S.; Shoukas, A.A.; Nyhan, D.; Champion, H.C.; et al. Arginase reciprocally regulates nitric oxide synthase activity and contributes to endothelial dysfunction in aging blood vessels. Circulation 2003, 108, 2000–2006. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An old enzyme with new tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A multifaceted enzyme important in health and disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef]
- Durante, W. Role of arginase in vessel wall remodeling. Front. Immunol. 2013, 4, 111. [Google Scholar] [CrossRef]
- Mahdi, A.; Pernow, J.; Kövamees, O. Arginase inhibition improves endothelial function in an age-dependent manner in healthy elderly humans. Rejuven. Res. 2019, 22, 385–389. [Google Scholar] [CrossRef]
- Moretto, J.; Girard, C.; Demougeot, C. The role of arginase in aging: A systematic review. Exp. Gerontol. 2019, 116, 54–73. [Google Scholar] [CrossRef] [PubMed]
- Pandya, C.D.; Lee, B.; Toque, H.A.; Mendhe, B.; Bragg, R.T.; Pandya, B.; Atawia, R.T.; Isales, C.; Hamrick, M.; Caldwell, R.W.; et al. Age-dependent oxidative stress elevates arginase 1 and uncoupled nitric oxide synthesis in skeletal muscle of aged mice. Oxid. Med. Cell. Longev. 2019, 2019, 1704650. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, L.; Christianson, D.W.; Nyhan, D.; Berkowitz, D.E. Arginase and vascular aging. J. Appl. Physiol. 2008, 105, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.S.; Berkowitz, D.E.; Ryoo, S.W. Increased arginase II activity contributes to endothelial dysfunction through endothelial nitric oxide synthase uncoupling in aged mice. Exp. Mol. Med. 2012, 44, 594–602. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine metabolism: Boundaries of our knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Regulation of enzymes of the urea cycle and arginine metabolism. Annu. Rev. Nutr. 2002, 22, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Arginine metabolism revisited. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.M.; Bhamidipati, D.; Kepka-Lenhart, D. Human type II arginase: Sequence analysis and tissue-specific expression. Gene 1997, 193, 157–161. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine metabolism in vascular biology and disease. Vasc. Med. Lond. Engl. 2005, 10 (Suppl. 1), S83–S87. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Recent advances in arginine metabolism: Roles and regulation of the arginases. Br. J. Pharmacol. 2009, 157, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Durante, W.; Liao, L.; Reyna, S.V.; Peyton, K.J.; Schafer, A.I. Transforming growth factor-β1 stimulates L-arginine transport and metabolism in vascular smooth muscle cells: Role in polyamine and collagen synthesis. Circulation 2001, 103, 1121–1127. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Moncada, S.; Higgs, E.A. The discovery of nitric oxide and its role in vascular biology. Br. J. Pharmacol. 2006, 147 (Suppl. 1), S193–S201. [Google Scholar] [CrossRef]
- Bignon, E.; Rizza, S.; Filomeni, G.; Papaleo, E. Use of computational biochemistry for elucidating molecular mechanisms of nitric oxide synthase. Comput. Struct. Biotechnol. J. 2019, 17, 415–429. [Google Scholar] [CrossRef]
- Leopold, J.A.; Cap, A.; Scribner, A.W.; Stanton, R.C.; Loscalzo, J. Glucose-6-phosphate dehydrogenase deficiency promotes endothelial oxidant stress and decreases endothelial nitric oxide bioavailability. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2001, 15, 1771–1773. [Google Scholar] [CrossRef]
- Leopold, J.A.; Zhang, Y.-Y.; Scribner, A.W.; Stanton, R.C.; Loscalzo, J. Glucose-6-phosphate dehydrogenase overexpression decreases endothelial cell oxidant stress and increases bioavailable nitric oxide. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 411–417. [Google Scholar] [CrossRef]
- Herrera, M.D.; Mingorance, C.; Rodríguez-Rodríguez, R.; Alvarez de Sotomayor, M. Endothelial dysfunction and aging: An update. Ageing Res. Rev. 2010, 9, 142–152. [Google Scholar] [CrossRef]
- El Assar, M.; Angulo, J.; Vallejo, S.; Peiró, C.; Sánchez-Ferrer, C.F.; Rodríguez-Mañas, L. Mechanisms involved in the aging-induced vascular dysfunction. Front. Physiol. 2012, 3, 132. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, E.M.; Vasconcelos, A.R.; Degaspari, S.; Böhmer, A.E.; Scavone, C.; Marcourakis, T. Age-related changes in nitric oxide activity, cyclic GMP, and TBARS levels in platelets and erythrocytes reflect the oxidative status in central nervous system. Age Dordr. Neth. 2013, 35, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Bouzón, C.; Carcaillon, L.; García-García, F.J.; Amor-Andrés, M.S.; El Assar, M.; Rodríguez-Mañas, L. Association between endothelial dysfunction and frailty: The Toledo Study for Healthy Aging. Age Dordr. Neth. 2014, 36, 495–505. [Google Scholar] [CrossRef]
- Yepuri, G.; Velagapudi, S.; Xiong, Y.; Rajapakse, A.G.; Montani, J.-P.; Ming, X.-F.; Yang, Z. Positive crosstalk between arginase-II and S6K1 in vascular endothelial inflammation and aging. Aging Cell 2012, 11, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Bugaj, L.J.; Oh, Y.J.; Bivalacqua, T.J.; Ryoo, S.; Soucy, K.G.; Santhanam, L.; Webb, A.; Camara, A.; Sikka, G.; et al. Arginase inhibition restores NOS coupling and reverses endothelial dysfunction and vascular stiffness in old rats. J. Appl. Physiol. Bethesda Md 1985 2009, 107, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Vatner, S.F.; Zhang, J.; Vyzas, C.; Mishra, K.; Graham, R.M.; Vatner, D.E. Vascular stiffness in aging and disease. Front. Physiol. 2021, 12, 762437. [Google Scholar] [CrossRef]
- Nóbrega-Pereira, S.; Fernandez-Marcos, P.J.; Brioche, T.; Gomez-Cabrera, M.C.; Salvador-Pascual, A.; Flores, J.M.; Viña, J.; Serrano, M. G6PD protects from oxidative damage and improves healthspan in mice. Nat. Commun. 2016, 7, 10894. [Google Scholar] [CrossRef]
- Arc-Chagnaud, C.; Salvador-Pascual, A.; Garcia-Dominguez, E.; Olaso-Gonzalez, G.; Correas, A.G.; Serna, E.; Brioche, T.; Chopard, A.; Fernandez-Marcos, P.J.; Serrano, M.; et al. Glucose 6-P dehydrogenase delays the onset of frailty by protecting against muscle damage. J. Cachexia Sarcopenia Muscle 2021, 12, 1879–1896. [Google Scholar] [CrossRef]
- Corcoran, C.M.; Fraser, P.; Martini, G.; Luzzatto, L.; Mason, P.J. High-level regulated expression of the human G6PD gene in transgenic mice. Gene 1996, 173, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, M.D.; Serna, E.; Fernández-Murga, M.L.; Portero, J.; Aldasoro, M.; Valles, S.L.; Sanz, Y.; Vila, J.M. Bifidobacterium pseudocatenulatum CECT 7765 supplementation restores altered vascular function in an experimental model of obese mice. Int. J. Med. Sci. 2017, 14, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segarra, G.; Cortina, B.; Mauricio, M.D.; Novella, S.; Lluch, P.; Navarrete-Navarro, J.; Noguera, I.; Medina, P. Effects of asymmetric dimethylarginine on renal arteries in portal hypertension and cirrhosis. World J. Gastroenterol. 2016, 22, 10545–10556. [Google Scholar] [CrossRef] [PubMed]
- Huynh, N.N.; Harris, E.E.; Chin-Dusting, J.F.P.; Andrews, K.L. The vascular effects of different arginase inhibitors in rat isolated aorta and mesenteric arteries. Br. J. Pharmacol. 2009, 156, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kerr, M.M.; Gourlay, T. Design and numerical simulation for the development of an expandable paediatric heart valve. Int. J. Artif. Organs 2021, 44, 518–524. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YWT | OWT | OTg |
|---|---|---|
| 207.9 | 204.7 | 204.6 |
| 208.5 | 226.6 | 202.9 |
| 211.5 | 237.7 | 205.2 |
| 206.9 | 243.2 | 204.4 |
| 207.0 | 218.6 | 204.6 |
| 209.8 | 212.6 | 208.2 |
| 208.6 ± 1.8 | 223.9 ± 14.8 | 205.0 ± 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serna, E.; Mauricio, M.D.; San-Miguel, T.; Guerra-Ojeda, S.; Verdú, D.; Valls, A.; Arc-Chagnaud, C.; De la Rosa, A.; Viña, J. Glucose 6-P Dehydrogenase Overexpression Improves Aging-Induced Endothelial Dysfunction in Aorta from Mice: Role of Arginase II. Int. J. Mol. Sci. 2023, 24, 3622. https://doi.org/10.3390/ijms24043622
Serna E, Mauricio MD, San-Miguel T, Guerra-Ojeda S, Verdú D, Valls A, Arc-Chagnaud C, De la Rosa A, Viña J. Glucose 6-P Dehydrogenase Overexpression Improves Aging-Induced Endothelial Dysfunction in Aorta from Mice: Role of Arginase II. International Journal of Molecular Sciences. 2023; 24(4):3622. https://doi.org/10.3390/ijms24043622
Chicago/Turabian StyleSerna, Eva, Maria D Mauricio, Teresa San-Miguel, Sol Guerra-Ojeda, David Verdú, Alicia Valls, Coralie Arc-Chagnaud, Adrián De la Rosa, and José Viña. 2023. "Glucose 6-P Dehydrogenase Overexpression Improves Aging-Induced Endothelial Dysfunction in Aorta from Mice: Role of Arginase II" International Journal of Molecular Sciences 24, no. 4: 3622. https://doi.org/10.3390/ijms24043622