

Extracellular Matrix Disorganization and Sarcolemmal Alterations in COL6-Related Myopathy Patients with New Variants of COL6 Genes
 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Clinical Features
2.2. Genetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Patient | Nucleotide Change | Protein Change | Protein Domain | ClinVar | Sift | PolyPhen 2 | Mutation Tester | ACMG Classification | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| COL6A1 NM_001848 | ||||||||||
| 6 | c.628C>G | p.(Arg210Gly) | vWFA1 | _ | Damaging | Probably damaging | Disease causing | Uncertain significance | New | |
| 4.1 | c.930+189C>T | p.Lys310_Gly311 insX [20] | THD | Pathogenic | [21] | |||||
| 1 | c.958-2A>G | p.(Gly320_Lys322del) | THD | Pathogenic | _ | _ | Disease causing | Pathogenic | rs1556425717 | |
| COL6A2 NM_001849 | ||||||||||
| Family 9 | c.1806C>G | p.(Cys602Trp) | Non-helical region | _ | Damaging | Damaging | Disease causing | Uncertain significance | New | |
| Family 9 | c.1832G>A | p.(Cys611Tyr) | Non-helical region | Probably damaging | Deleterious | Probably damaging | Disease causing | Uncertain significance | rs1375051583 | |
| Family 7 | c.2145C>G | p.(Ile715Met) | vWFA2 | _ | Damaging | Probably damaging | Disease causing | Uncertain significance | New | |
| Family 7 | c.2192C>T | p.(Thr731Met) | vWFA2 | Pathogenic | Deleterious | Probably damaging | Disease causing | Likely pathogenic | rs794727419 [22] | |
| Family 8 | c.2423-2A>G | p.(Asp808_Thr820del) | Link | Pathogenic | [23] | |||||
| 2 | c.2503C>T | p.(Arg835Cys) 1 | vWFA3 | Uncertain significance | Deleterious | Possibly damaging | Disease causing | Likely benign | rs534856775 | |
| Family 8 | c.2738_2740del | p.(Ser913del) | vWFA3 | _ | _ | _ | Disease causing | Uncertain significance | New | |
| COL6A3 NM_004369 | ||||||||||
| 2 | c.1688A>G | p.(Asp563Gly) | vWFA3 | Uncertain significance | Deleterious | Probably damaging | Disease causing | Uncertain significance | rs112913396 [24] | |
| 5 | c.2195C>T | p.(Thr732Met) | vWFA4 | Uncertain significance | Deleterious | Possible damaging | Disease causing | Likely benign | rs370719148 | |
| 3 | c.7928C>T | p.(Ala2643Val) | vWFA12 | Uncertain significance | Deleterious | Possible damaging | Polymorphism | Benign | rs111595697 | |
| 3 | c.8572G>A | p.(Val2858Ile) | Link | Uncertain significance | Tolerated | Benign | Polymorphism | Benign | rs111859552 |
2.3. Muscle Biopsy
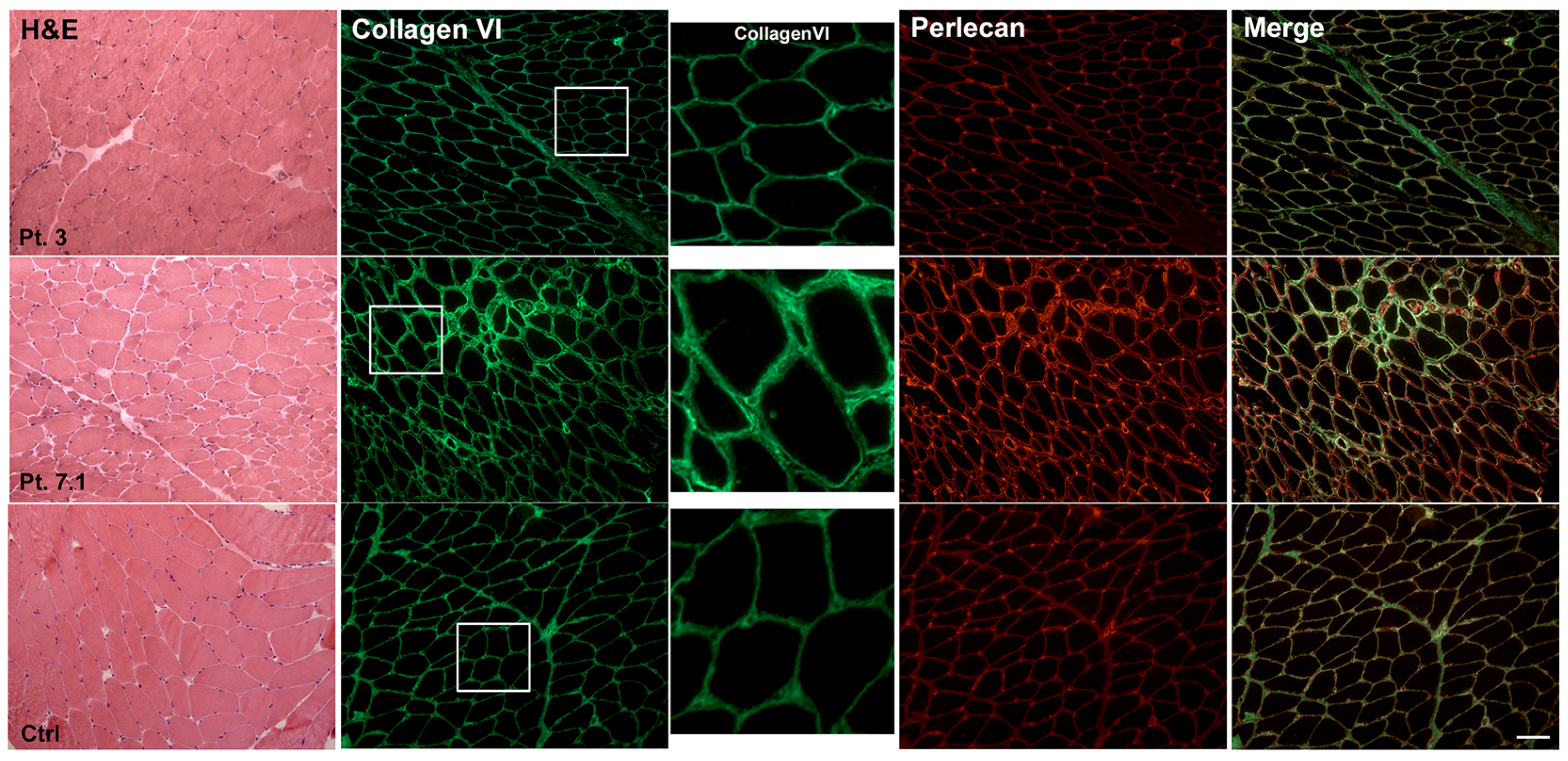
2.4. Muscle Immunohistochemistry and Skin Fibroblasts Immunofluorescence
2.5. Western Blot
2.6. Electron Microscopy Examination
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldock, C.; Sherratt, M.J.; Shuttleworth, C.A.; Kielty, C.M. The Supramolecular Organization of Collagen VI Microfibrils. J. Mol. Biol. 2003, 330, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Petrini, S.; Tessa, A.; Stallcup, W.B.; Sabatelli, P.; Pescatori, M.; Giusti, B.; Carrozzo, R.; Verardo, M.; Bergamin, N.; Columbaro, M.; et al. Altered Expression of the MCSP/NG2 Chondroitin Sulfate Proteoglycan in Collagen VI Deficiency. Mol. Cell Neurosci. 2005, 30, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Nihei, K.; Kamoshita, S.; Atsumi, T. A Case of Ullrich’s Disease (Kongenitale, Atonisch-Sklerotische Muskeldystrophie). Brain Dev. 1979, 1, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Bethlem, J.; Wijngaarden, G.K. Benign Myopathy, with Autosomal Dominant Inheritance. A Report on Three Pedigrees. Brain 1976, 99, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Bönnemann, C.G. The Collagen VI-Related Myopathies: Muscle Meets Its Matrix. Nat. Rev. Neurol. 2011, 7, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Merlini, L.; Martoni, E.; Grumati, P.; Sabatelli, P.; Squarzoni, S.; Urciuolo, A.; Ferlini, A.; Gualandi, F.; Bonaldo, P. Autosomal Recessive Myosclerosis Myopathy Is a Collagen VI Disorder. Neurology 2008, 71, 1245–1253. [Google Scholar] [CrossRef]
- Scacheri, P.C.; Gillanders, E.M.; Subramony, S.H.; Vedanarayanan, V.; Crowe, C.A.; Thakore, N.; Bingler, M.; Hoffman, E.P. Novel Mutations in Collagen VI Genes: Expansion of the Bethlem Myopathy Phenotype. Neurology 2002, 58, 593–602. [Google Scholar] [CrossRef]
- Bushby, K.M.D.; Collins, J.; Hicks, D. Collagen Type VI Myopathies. Adv. Exp. Med. Biol. 2014, 802, 185–199. [Google Scholar] [CrossRef]
- Yonekawa, T.; Nishino, I. Ullrich Congenital Muscular Dystrophy: Clinicopathological Features, Natural History and Pathomechanism(s). J. Neurol. Neurosurg. Psychiatry 2015, 86, 280–287. [Google Scholar] [CrossRef]
- Bersini, S.; Gilardi, M.; Mora, M.; Krol, S.; Arrigoni, C.; Candrian, C.; Zanotti, S.; Moretti, M. Tackling Muscle Fibrosis: From Molecular Mechanisms to next Generation Engineered Models to Predict Drug Delivery. Adv. Drug Deliv. Rev. 2018, 129, 64–77. [Google Scholar] [CrossRef]
- Martinez, F.J.; Collard, H.R.; Pardo, A.; Raghu, G.; Richeldi, L.; Selman, M.; Swigris, J.J.; Taniguchi, H.; Wells, A.U. Idiopathic Pulmonary Fibrosis. Nat. Rev. Dis. Primers 2017, 3, 17074. [Google Scholar] [CrossRef]
- Schuppan, D.; Ashfaq-Khan, M.; Yang, A.T.; Kim, Y.O. Liver Fibrosis: Direct Antifibrotic Agents and Targeted Therapies. Matrix Biol. 2018, 68–69, 435–451. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Q.; Kong, W. Extracellular Matrix Remodeling and Cardiac Fibrosis. Matrix Biol. 2018, 68–69, 490–506. [Google Scholar] [CrossRef]
- Schulz, J.-N.; Plomann, M.; Sengle, G.; Gullberg, D.; Krieg, T.; Eckes, B. New Developments on Skin Fibrosis - Essential Signals Emanating from the Extracellular Matrix for the Control of Myofibroblasts. Matrix Biol. 2018, 68–69, 522–532. [Google Scholar] [CrossRef]
- Mahdy, M.A.A. Skeletal Muscle Fibrosis: An Overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef]
- Smith, L.R.; Barton, E.R. Regulation of Fibrosis in Muscular Dystrophy. Matrix Biol. 2018, 68–69, 602–615. [Google Scholar] [CrossRef]
- Solomon-Degefa, H.; Gebauer, J.M.; Jeffries, C.M.; Freiburg, C.D.; Meckelburg, P.; Bird, L.E.; Baumann, U.; Svergun, D.I.; Owens, R.J.; Werner, J.M.; et al. Structure of a Collagen VI A3 Chain VWA Domain Array: Adaptability and Functional Implications of Myopathy Causing Mutations. J. Biol. Chem. 2020, 295, 12755–12771. [Google Scholar] [CrossRef]
- Engvall, E.; Hessle, H.; Klier, G. Molecular Assembly, Secretion, and Matrix Deposition of Type VI Collagen. J. Cell Biol. 1986, 102, 703–710. [Google Scholar] [CrossRef] [Green Version]
- Specks, U.; Mayer, U.; Nischt, R.; Spissinger, T.; Mann, K.; Timpl, R.; Engel, J.; Chu, M.L. Structure of Recombinant N-Terminal Globule of Type VI Collagen Alpha 3 Chain and Its Binding to Heparin and Hyaluronan. EMBO J. 1992, 11, 4281–4290. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, W.J.; Kim, H.; Choi, S.A.; Lee, J.S.; Cho, A.; Jang, S.S.; Lim, B.C.; Kim, K.J.; Kim, J.-I.; et al. Collagen VI-Related Myopathy: Expanding the Clinical and Genetic Spectrum. Muscle Nerve 2018, 58, 381–388. [Google Scholar] [CrossRef]
- Cummings, B.B.; Marshall, J.L.; Tukiainen, T.; Lek, M.; Donkervoort, S.; Foley, A.R.; Bolduc, V.; Waddell, L.B.; Sandaradura, S.A.; O’Grady, G.L.; et al. Improving Genetic Diagnosis in Mendelian Disease with Transcriptome Sequencing. Sci. Transl. Med. 2017, 9, eaal5209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, A.R.; Quijano-Roy, S.; Collins, J.; Straub, V.; McCallum, M.; Deconinck, N.; Mercuri, E.; Pane, M.; D’Amico, A.; Bertini, E.; et al. Natural History of Pulmonary Function in Collagen VI-Related Myopathies. Brain 2013, 136, 3625–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, A.K.; Dunn, D.M.; von Niederhausern, A.C.; Hamil, C.; Aoyagi, A.; Laval, S.H.; Marie, S.K.; Chu, M.-L.; Swoboda, K.; Muntoni, F.; et al. Automated Genomic Sequence Analysis of the Three Collagen VI Genes: Applications to Ullrich Congenital Muscular Dystrophy and Bethlem Myopathy. J. Med. Genet. 2005, 42, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterfield, R.J.; Foley, A.R.; Dastgir, J.; Asman, S.; Dunn, D.M.; Zou, Y.; Hu, Y.; Donkervoort, S.; Flanigan, K.M.; Swoboda, K.J.; et al. Position of Glycine Substitutions in the Triple Helix of COL6A1, COL6A2, and COL6A3 Is Correlated with Severity and Mode of Inheritance in Collagen VI Myopathies. Hum. Mutat. 2013, 34, 1558–1567. [Google Scholar] [CrossRef] [Green Version]
- Briñas, L.; Richard, P.; Quijano-Roy, S.; Gartioux, C.; Ledeuil, C.; Lacène, E.; Makri, S.; Ferreiro, A.; Maugenre, S.; Topaloglu, H.; et al. Early Onset Collagen VI Myopathies: Genetic and Clinical Correlations. Ann. Neurol. 2010, 68, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.Z.; Zou, Y.; Pan, T.C.; Markova, D.; Fertala, A.; Hu, Y.; Squarzoni, S.; Reed, U.C.; Marie, S.K.N.; Bonnemann, C.G.; et al. Recessive COL6A2 C-Globular Missense Mutations in Ullrich Congenital Muscular Dystrophy: Role of the C2a Splice Variant. J. Biol. Chem. 2010, 285, 10005–10015. [Google Scholar] [CrossRef] [Green Version]
- Telfer, W.R.; Busta, A.S.; Bonnemann, C.G.; Feldman, E.L.; Dowling, J.J. Zebrafish Models of Collagen VI-Related Myopathies. Hum. Mol. Genet. 2010, 19, 2433–2444. [Google Scholar] [CrossRef] [Green Version]
- Irwin, W.A.; Bergamin, N.; Sabatelli, P.; Reggiani, C.; Megighian, A.; Merlini, L.; Braghetta, P.; Columbaro, M.; Volpin, D.; Bressan, G.M.; et al. Mitochondrial Dysfunction and Apoptosis in Myopathic Mice with Collagen VI Deficiency. Nat. Genet. 2003, 35, 367–371. [Google Scholar] [CrossRef]
- Tagliavini, F.; Sardone, F.; Squarzoni, S.; Maraldi, N.M.; Merlini, L.; Faldini, C.; Sabatelli, P. Ultrastructural Changes in Muscle Cells of Patients with Collagen VI-Related Myopathies. Muscles Ligaments Tendons J. 2013, 3, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Sugie, K.; Murayama, K.; Ito, M.; Minami, N.; Nishino, I.; Nonaka, I. Ullrich Disease: Collagen VI Deficiency: EM Suggests a New Basis for Muscular Weakness. Neurology 2002, 59, 920–923. [Google Scholar] [CrossRef]
- Squarzoni, S.; Sabatelli, P.; Bergamin, N.; Guicheney, P.; Demir, E.; Merlini, L.; Lattanzi, G.; Ognibene, A.; Capanni, C.; Mattioli, E.; et al. Ultrastructural Defects of Collagen VI Filaments in an Ullrich Syndrome Patient with Loss of the Alpha3(VI) N10-N7 Domains. J. Cell Physiol. 2006, 206, 160–166. [Google Scholar] [CrossRef]
- Cenacchi, G.; Fanin, M.; De Giorgi, L.B.; Angelini, C. Ultrastructural Changes in Dysferlinopathy Support Defective Membrane Repair Mechanism. J. Clin. Pathol. 2005, 58, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Kubisch, C.; Schoser, B.G.H.; von Düring, M.; Betz, R.C.; Goebel, H.-H.; Zahn, S.; Ehrbrecht, A.; Aasly, J.; Schroers, A.; Popovic, N.; et al. Homozygous Mutations in Caveolin-3 Cause a Severe Form of Rippling Muscle Disease. Ann. Neurol. 2003, 53, 512–520. [Google Scholar] [CrossRef]



| Patient | Sex | Age (Years) | Age of Onset (Years) | First Symptoms | CLINICAL CLUES | Retractions | CPK (U/L) | Respiratory Difficulty | Others |
|---|---|---|---|---|---|---|---|---|---|
| 1 | F | 30 | 4 | Motor development delay Hypotonia at birth | Proximal and distal weakness (4 limbs) Loss of independent ambulation (10 years) | Yes (Multiple surgery) | 300 | Mild restrictive | Cheloids, scoliosis |
| 2 | F | 26 | 4 | Weakness of the lower limbs | Proximal and distal weakness (4 limbs) | Yes | 2568–12,518 | Mild restrictive | _ |
| 3 | F | 34 | 2 | Fingers flexors contractures (2 years) | Proximal and distal weakness (4 limbs) | Yes (Multiple surgery) | Normal | No | Rigid spine, ptosis, bilateral cataract |
| 4.1 | M | 16 | 1.2 | Mild proximal weakness motor milestones delay | Proximal weakness (4 limbs), loss of independent ambulation (5 years) | Yes | Normal-258 | Mild restrictive | Ptosis, mild scoliosis, hyperlaxity |
| 4.2 | M | 48 | _ | Mild right ptosis | Mild proximal weakness at lower limbs | No | 2500 | Mild restrictive | _ |
| 5 | F | 72 | 49 | Proximal weakness | _ | _ | 3000 | _ | _ |
| 6 | F | 27 | _ | _ | _ | _ | _ | _ | _ |
| 7.1 | M | 57 | _ | Proximal weakness | Proximal > distal weakness (upper > lower limbs) | Yes | _ | Obstructive sleep apnoea syndrome | _ |
| 7.2 | M | 47 | _ | Mialgia and cramps | No motor impairment up to 43 years | Yes | 700–800 | No | _ |
| 7.3 | F | 58 | _ | Mild CPK increase | Proximal and distal weakness lower limbs | _ | Mild increase | No | Rigid trunk |
| 7.4 | F | 61 | _ | Mild CPK increase | Mild proximal weakness | _ | _ | _ | _ |
| 8.1 | M | 59 | Early childhood | Mild proximal weakness | Proximal and distal weakness (4 limbs) | No | _ | No | Dysphagia |
| 8.2 | F | 68 | Early childhood | Weakness and fatigue | Proximal and distal weakness (4 limbs) | No | Normal | No | _ |
| 9.1 | M | 56 | 4 | Weakness lower limbs | Proximal and distal weakness (4 limbs) | Yes (moderate) | 203–523 | Severe restrictive | Rigid spine |
| 9.2 | M | 41 | 10 | Weakness lower limbs | Proximal and distal weakness (4 limbs) | Yes (moderate) | 190–425 | Severe restrictive | Rigid spine |
| Patient | Fiber Size Variability | Fiber Type Distribution | Centronuclear Fibers | Fibrosis (%Area) | Muscle Tissue IHC COL6 | Western Blot COL6 | Skin Fibroblast IF COL6 | Ultrastructural Analysis M.E. |
|---|---|---|---|---|---|---|---|---|
| 1 | Marked | Correct typological differentiation and topographic distribution | Rare | 27.72 ± 6.52 | Widespread increase/some fibers with a subtle reduction/absence membrane staining | Slight reduction | Retention | Microvilli-like projections of membrane/Collagen fibrils through the membrane/Increase glycogen content |
| 2 | Marked variability | _ | No | 39.73 ± 4.08 | Increase at perimysium Normal at membrane | Reduction | Partial retention/globular speckles | _ |
| 3 | Mild | Correct typological differentiation Hypotrophic fibers predominantly of type II | No | 11.48 ± 2.71 | Normal | Normal | Patchy distribution | Subsarcolemmal vacuoles, sometimes containing replicated membranes-like |
| 4.1 | Marked | Correct typological differentiation Hypotrophic fibers of both types | Rare | 39.92 ± 8.12 | Marked increase at perimysium Some fibers with irregular sarcolemma staining | Normal | Retention | Microvilli-like projections of membrane/Rare streaming of Z line/Rare dilation of sarcoplasmic reticulum |
| 4.2 | Normal | Correct typological differentiation and topographic distribution | Rare | 12.82 ± 2.86 | Some fibers with a subtle reduction Absence membrane staining | _ | Weak distribution | _ |
| 5 | Discrete | Hypotrophic fibers of both types | Rare | 12.91 ± 0.61 | _ | _ | _ | Sarcolemma extroflection/Basal lamina replication/Collagen fibrils through the membrane/Subsarcolemmal vacuoles |
| 6 | Marked | Hypotrophic fibers predominantly of type I | No | 18.13 ± 5.28 | Discrete increase Some fibers with a subtle reduction/absence membrane staining | _ | _ | Collagen fibrils through the membrane Subsarcolemmal vacuoles, containing membranes-like Basal lamina extroflection and replication Rare streaming of Z line |
| 7.1 | Marked | Hypotrophic fibers predominantly of type II | Rare | 14.30 ± 3.99 | Strong labelling of the basement membrane and endomysium | _ | Partial retention, patchy distribution | Collagen fibrils go through the membrane Increase glycogen content |
| 7.2 | _ | _ | _ | _ | _ | _ | Partial retention patchy distribution | _ |
| 7.3 | _ | _ | _ | _ | _ | _ | Partial retention patchy distribution | _ |
| 9.2 | Discrete | _ | Slightly increase | 14.83 ± 1.09 | Some fibers with a subtle reduction Absence membrane staining | _ | _ | _ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanotti, S.; Magri, F.; Salani, S.; Napoli, L.; Ripolone, M.; Ronchi, D.; Fortunato, F.; Ciscato, P.; Velardo, D.; D’Angelo, M.G.; et al. Extracellular Matrix Disorganization and Sarcolemmal Alterations in COL6-Related Myopathy Patients with New Variants of COL6 Genes. Int. J. Mol. Sci. 2023, 24, 5551. https://doi.org/10.3390/ijms24065551
Zanotti S, Magri F, Salani S, Napoli L, Ripolone M, Ronchi D, Fortunato F, Ciscato P, Velardo D, D’Angelo MG, et al. Extracellular Matrix Disorganization and Sarcolemmal Alterations in COL6-Related Myopathy Patients with New Variants of COL6 Genes. International Journal of Molecular Sciences. 2023; 24(6):5551. https://doi.org/10.3390/ijms24065551
Chicago/Turabian StyleZanotti, Simona, Francesca Magri, Sabrina Salani, Laura Napoli, Michela Ripolone, Dario Ronchi, Francesco Fortunato, Patrizia Ciscato, Daniele Velardo, Maria Grazia D’Angelo, and et al. 2023. "Extracellular Matrix Disorganization and Sarcolemmal Alterations in COL6-Related Myopathy Patients with New Variants of COL6 Genes" International Journal of Molecular Sciences 24, no. 6: 5551. https://doi.org/10.3390/ijms24065551