Abstract
The transcriptional coactivators Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) are the main downstream effectors of the evolutionarily conserved Hippo signaling pathway. YAP/TAZ are implicated in the transcriptional regulation of target genes that are involved in a wide range of key biological processes affecting tissue homeostasis and play dual roles in the aging process, depending on the cellular and tissue context. The aim of the present study was to investigate whether pharmacological inhibitors of Yap/Taz increase the lifespan of Drosophila melanogaster. Real-time qRT-PCR was performed to measure the changes in the expression of Yki (Yorkie, the Drosophila homolog of YAP/TAZ) target genes. We have revealed a lifespan-increasing effect of YAP/TAZ inhibitors that was mostly associated with decreased expression levels of the wg and E2f1 genes. However, further analysis is required to understand the link between the YAP/TAZ pathway and aging.
1. Introduction
With the increase in the average age of the population, preventing premature aging and treating age-related diseases has become a major concern in modern healthcare [,,]. The promising approach to achieve this goal is to influence the major molecular mechanisms associated with aging which is the main risk factor for age-related diseases, in order to suppress pathological processes and activation of the defense systems of the cell and the body as a whole [,,].
Recent studies demonstrate that damage to long-lived macromolecules, including extracellular matrix (ECM) proteins, make a significant contribution to the aging process [,]. The accumulation of non-enzymatic modifications by glycation, oxidation, and crosslinking of collagen and elastin, the major components of the ECM, occurs during aging [,]. The modifications of macromolecules affect the structural and physical properties of the ECM that increase the stiffness of tissues and reduce their viscoelasticity [,,,]. High ECM stiffness promotes activation of Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ), the Hippo pathway effectors [] that play a key role in the regulation of tissue homeostasis []. YAP/TAZ are transcription coactivators without DNA-binding activity that activate or repress target gene expression through interaction with various transcription factors []. ECM stiffness has been shown to regulate YAP/TAZ-driven gene transcription independent of the Hippo pathway [] and make a significant contribution to the deregulation of tissue homeostasis during aging [,]. Due to YAP/TAZ regulating target genes involved in a wide range of key biological processes, such as modulation of nuclear integrity and functions [], stem cell differentiation [], cell proliferation [], regeneration [], innate immune response [,], and tumorigenesis [], YAP/TAZ activity plays dual roles in the aging process depending on cellular and tissue context [,]. For example, reduced expression of connective tissue growth factor (CTGF), an established YAP/TAZ target gene, which is involved in tissue remodeling, has been reported to mediate collagen loss in chronologically-aged human skin []; whereas, persistent activation of CTGF can result in increased deposition of collagen and fibrotic conditions [].
Despite a large amount of experimental data having demonstrated a decrease in YAP/TAZ activity during physiological aging [,,,], a number of studies have shown that several compounds with proven anti-aging properties such as resveratrol [], rapamycin [], metformin [], and simvastatin [] inhibit transcriptional activity of YAP/TAZ [,]. However, the effects of geroprotectors on YAP/TAZ activity in normal tissues have not yet been studied.
Drosophila melanogaster is one of the most studied and genetically tractable model organisms for investigating the mechanisms of aging and anti-aging interventions [,] using evolutionarily conserved aging-related signaling pathways as potential drug targets [,]. Drosophila Yorkie (Yki) is a homolog of mammalian YAP/TAZ [,] that allows the use of fly models in studies of the geroprotective properties of Yap/Taz inhibitors.
The aim of this study was to investigate whether pharmacological inhibitors of Yki/Yap/Taz could improve the survival of D. melanogaster. In this study, we investigated the effects of pharmacological inhibitors of Yki/Yap/Taz on D. melanogaster survival. The following substances, with previously established inhibitory effects on Yap/Taz activity, were used: Verteporfin (VP) [,,], ML-7 hydrochloride (ML7) [], Cytochalasin D (CD) [,,], and AICAR (AI) [].
2. Results
2.1. Expression Levels of YAP/TAZ Target Genes
Real-time qRT-PCR analyses of YAP/TAZ target genes expression provide direct quantitative estimation of YAP/TAZ transcriptional activity [,]. The analyzed targets of Yki included genes that promote cellular growth (myc) [], genes involved in cell cycle progression (CycE and E2F1) [,], inhibitors of apoptosis (Diap1) [], genes encoding ligands for different signaling pathways (wg and vein) [,], and modulators of signaling pathway activity (dally) [].
To study whether the expression levels of Yki target genes changed with aging, qRT-PCR was performed for male and female flies at the ages of 10 and 20 days (Figure 1, Table S1). Two-way ANOVA analysis (gene × age) showed a significant effect of the gene (p < 0.001), a significant effect of the age (p < 0.05), and a significant interaction (p < 0.001) in male and female flies (Figure 1, Table S2). Given that cycle threshold (Ct) values are inversely proportional to mRNA transcript levels, higher differences in Ct values (dCt) between target genes and reference genes represent a lower expression level. A Duncan post hoc test of the 10-day-old flies versus 20-day-old flies within each gene revealed a significant (p < 0.05) age-related decrease in the expression levels of CycE, dally, Diap1 in males and CycE, dally, myc in females, but an age-related increase in the expression level of wg in females (p < 0.01). As most of the statistically significant changes indicated a decrease in the expression level (increase in dCt values) of the analyzed genes, this suggests a possible age-related decrease in the level of Yki transcriptional activity.

Figure 1.
Age-related changes in the expression level of Yki target genes. Estimation of the expression level in males (A) and females (B) at the age of 10 and 20 days. Two-way ANOVA (age × gene) followed by post hoc Duncan test was used to compare differences in expression levels of genes between 10-day-old and 20-day-old flies, * p < 0.05, ** p < 0.01, *** p < 0.001, ns—not significant. The Ct (cycle thresholds) values are inversely proportional to the mRNA transcript levels. The delta Ct (dCt) values were calculated as the differences in Ct values for target genes and reference genes (β-Tubulin and RpL32). Higher dCt values represent a lower expression level. The error bars show standard errors. The experiments were performed in three biological replicates and three technical replicates (n = 3, repeated three times). The samples included 20 males and 10 females.
These results are consistent with previously published data which demonstrated an age-related decrease in the transcriptional activity of YAP/TAZ [,]. However, it should be noted that because of the complex crosstalk between YAP/TAZ and other signaling pathways, such as Wnt [] and Notch [], several of the used genes are not exclusive targets of the Yki pathway. Therefore, for a more convincing conclusion regarding age-related changes in Yki pathway activity, a more direct and specific read-out of Yki activity should be used in further research.
To assess whether treatment with various concentrations of substances was associated with the inhibition of YAP/TAZ transcriptional activity, the expression levels of Yki target genes were compared between treated and control groups of different ages within each inhibitor.
Two-way ANOVA (gene × concentration) showed that there were significant differences in expression levels among the different genes (p < 0.001, source of variation: gene) in 10-day-old male and female flies. The ANOVA also revealed significant differences in gene expression between the control animals and animals treated with inhibitors at different concentrations (p < 0.05, source of variation: concentration), except males and females treated with AI. Additionally, the ANOVA demonstrated that effects of the inhibitor treatment depended on the gene (p < 0.001, source of variation: interaction), except males treated with CD (Figure 2, Table S3).

Figure 2.
Effects of YAP/TAZ inhibitors on the expression level of Yki target genes. Estimation of the expression level in 10 day-old males and females after treatment with VP (A,B); ML7 (C,D); CD (E,F); AI (G,H). Two-way ANOVA (age × gene) followed by post hoc Duncan test was used to compare differences in expression levels of genes between 10-day-old and 20-day-old flies, * p < 0.05, ** p < 0.01, *** p < 0.001. The Ct (cycle thresholds) values are inversely proportional to the mRNA transcript levels. The delta Ct (dCt) values were calculated as the differences in Ct values for target genes and reference genes (β-Tubulin and RpL32). Higher dCt values represent a lower expression level. The error bars show standard errors. The experiments were performed in three biological replicates and three technical replicates (n = 3, repeated three times). The samples included 20 males and 10 females.
Duncan test of the inhibitor-treated 10-day-old male flies versus control age- and sex-matched flies within each substance revealed a significant (p < 0.05) decrease in the expression level of CycE (0.01 μM ML7), Diap1 (1 μM VP and 1 μM CD), E2f1 (0.1–1 μM CD), wg (0.01–10 μM VP; 0.1 μM ML7; 1 μM CD), Yki (1 μM CD), but an increase (p < 0.05) in the expression of CycE (0.01 μM VP; 0.1 μM AI), myc (0.01–10 μM VP), vn (0.01–10 μM VP), wg (1 μM ML7), and Yki (0.01 and 10 μM VP) (Figure 2, Table S4). At the same time, the post hoc pairwise comparison of gene expression level in treated and control 10-day-old females demonstrated a decrease (p < 0.05) in the expression of CycE (0.01 μM VP), vn (1 μM VP), wg (1 μM ML7; 0.1–1 μM CD; 0.1 μM ML7), but an increase (p < 0.05) in the expression of CycE (0.01 μM ML7; 0.1 μM CD), myc (0.1 μM CD), vn (10 μM VP), wg (1–10 μM VP), and Yki (10 μM VP; 0.1–1 μM AI) (Figure 2, Table S5). All inhibitors have shown the ability to suppress the expression of at least one gene. The most characteristic effect of used inhibitors is the suppression of wg gene expression (except for females treated with VP).
The observed differences in the expression levels between different genes in individuals of the same sex and between the same genes in males and females are consistent with the transcriptional data from the FlyAtlas2 database [], and reflect sex differences in gene expression of most of the studied genes, including Yki target genes.
To determine whether 20 days of treatment is more effective than 10 days of treatment for inhibition of YAP/TAZ transcriptional activity, the expression level of Yki target genes was compared between treated and control groups at the age of 20 days.
Two-way ANOVA (gene × concentration) showed that there was a significant effect of the gene (p < 0.001), a significant effect of the concentration (p < 0.05, except males treated with VP, ML7, AI), and a significant interaction (p < 0.05, except males treated with AI and females treated with ML7, CD, AI) in 20-day-old flies (Figure S1, Table S6).
Duncan post-test of the 20-day-old flies versus ones within each gene revealed a significant (p < 0.05) decrease in expression level of the CycE gene in female flies treated with VP (1 μM) and with CD (0.01 μM), but increased in males treated with ML7 (1 μM). The Duncan’s test also demonstrated a decrease (p < 0.05) in expression of the wg gene in males and females treated with VP (0.01 μM and 1 μM, respectively) and with ML7 (0.01 μM), as well as in females treated with CD (0.1 μM). However, wg expression increased in males treated with CD (0.1 μM) and females treated with VP (10 μM) and AI (0.1 μM) (Figure S1, Tables S7 and S8).
Thus, among four Yap/Taz inhibitors, the most pronounced effects on gene expression levels were detected in flies at the age of 10 days. Based on the obtained results, we used the concentration which showed the most significant effect on gene expression to determine the effect of substances on Drosophila melanogaster lifespan.
2.2. Effects on Survival
To address whether the used solvents have potential toxic effects on flies, we compared the lifespan between the control group treated with 100% (v/v) water and control groups that were treated with 100% (v/v) ethanol, 50% (v/v) solution of ethanol in water, and 10% (v/v) solution of Cyrene in water. Pairwise comparisons of the survival curves, median, and maximum lifespan (using Fleming–Harrington test and Fisher’s exact test with Bonferroni correction, respectively) revealed a positive effect of 100% (v/v) ethanol (in females), negative effect of 50% (v/v) ethanol solution (in males), and positive effect of 10% (v/v) Cyrene solution (in males and females) (Figure S2; Table S9). These results indicate that there are no acute toxic effects of the used solvents. A further analysis of the effects of Yap/Taz inhibitors was then conducted using different appropriate control groups.
According to the results of qRT-PCR analysis, the following concentrations of the YAP/TAZ inhibitors were chosen for the survival analysis: 0.01 μM and 0.1 μM—for VP, 0.1 μM and 1 μM—for ML7, CD, and AI. Two independent replicate experiments were completed for each group (Figures S3 and S4, Table S10).
To reduce incidental effects, we pooled the results of two replicates (Figure 3, Table S11). Using Fisher’s Exact test to compare median and maximum lifespan and Fleming–Harrington test to estimate early and later differences between survival curves of treated and control flies, we revealed the lifespan effects of YAP/TAZ inhibitors.
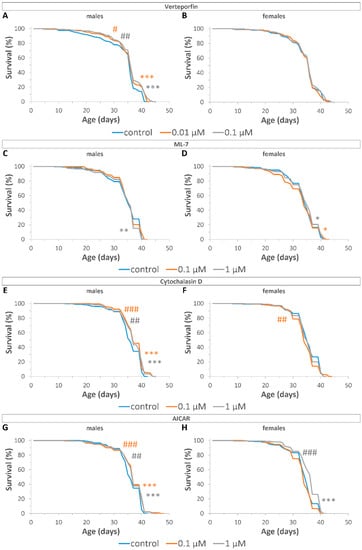
Figure 3.
Effects of YAP/TAZ inhibitors on the lifespan of male (A,C,E,G) and female (B,D,F,H) flies after treatment with VP (A,B); ML7 (C,D); CD (E,F); AI (G,H). Fleming–Harrington test sensitive against early (# p < 0.05, ## p < 0.01, ### p < 0.001) and later (* p < 0.05, ** p < 0.01, *** p < 0.001) differences were used to compare survival curves between inhibitor-treated and control flies. Bonferroni correction was used for multiple comparisons. Two independent experiments were performed: n = 287 (VP control, males); n = 306 (0.01 µM VP, males); n = 293 (0.1 µM VP, males); n = 290 (ML control, males); n = 277 (0.1 µM ML, males); n = 299 (1 µM ML, males); n = 290 (CD control, males); n = 324 (0.1 µM CD, males); n = 303 (1 µM CD, males); n = 293 (AI control, males); n = 321 (0.1 µM AI, males); n = 293 (1 µM AI, males); n = 282 (VP control, females); n = 305 (0.01 µM VP, females); n = 281 (0.1 µM VP, females); n = 300 (ML control, females); n = 274 (0.1 µM ML, females); n = 283 (1 µM ML, females); n = 292 (CD control, females); n = 293 (0.1 µM CD, females); n = 330 (1 µM CD, females); n = 323 (AI control, females); n = 310 (0.1 µM AI, females); n = 330 (1 µM AI, females).
VP at concentrations of 0.01 μM and 0.1 μM increased the maximum lifespan (by 2%, p < 0.001), as well as reduced the rate of early (p < 0.05) and late mortality (p < 0.001) in males, but had no effect (p > 0.05) on the survival of females (Figure 3A,B, Table S11). ML7 at concentration of 1 μM increased late mortality (p < 0.01) in males, but at concentrations of 0.1 μM and 1 μM reduced late mortality (p < 0.05) in females (Figure 3C,D, Table S11). CD at concentrations of 0.1 μM and 1 μM increased median (by 3%, p < 0.05) and maximum (by3%, p < 0.01) lifespan in males, respectively. However, CD at a concentration of 0.1 μM increased the rate of early mortality (p < 0.01) in females (Figure 3E,F, Table S11). AI at concentration of 0.1 μM increased maximum lifespan (by 3%, p < 0.05), and at concentrations of 0.1 μM and 1 μM reduced the rate of early and late mortality (p < 0.001) in males. AI at concentrations of 0.1 μM and 1 μM increased median lifespan in females by 3%, p < 0.01 and by 6%, p < 0.001, respectively, and at a concentration of 1 μM reduced the rate of early (p < 0.001) and late mortality (p < 0.001) in females (Figure 3G,H, Table S11).
Thus, Drosophila treatment with YAP/TAZ inhibitors decreased late mortality, which indicates a geroprotective potential of this intervention.
2.3. Effects on Inflammatory Markers of Aging
Previously, it has been demonstrated that aging in Drosophila is accompanied by a considerable increase in the expression of immune-related genes [,,]. In order to assess the effect of Yap/Taz inhibitors on inflammatory markers of aging, we compared the expression level of antimicrobial genes (Attacin-A (AttA), Diptericin A (DptA), cecropin A1 (CecA1)) between treated and control flies of both sexes at the age of 10 and 20 days.
Three-way ANOVA (gene × concentration × age) showed that there was a significant effect of the gene (p < 0.001), a significant effect of the concentration (p < 0.05, except males treated with AI and females treated with CD), and a significant effect of the age (p < 0.01) of flies (Figure S5, Table S12).
A Duncan test of the 10-day-old control flies versus 20-day-old ones within each substance in male and female flies revealed a significant (p < 0.05) age-related increase in the expression level of AttA (VP and CD treated males and females, AI females), DptA (CD treated males and females, AI females), and CecA1 (CD and AI treated females) (Figure S5, Tables S12–S14).
A Duncan test of the 20-day-old control flies with increased expression of antimicrobial genes versus age- and sex-matched treated flies within each substance revealed a significant (p < 0.05) decrease in the expression level of AttA (0.01 μM VP treated males) and DptA (0.01 μM VP treated males, 0.1 μM VP treated males and females), demonstrating the anti-aging effect of VP (Figure S5, Tables S12–S14). In addition, a Duncan test of the 10-day-old control flies versus age- and sex-matched treated flies within each substance revealed a significant (p < 0.05) anti-inflammatory effect of VP that was manifested in the decreased expression level of AttA (0.1 μM VP treated males) and DptA (0.1 μM VP treated males and females). At the same time, CD treatment significantly increased the expression of AttA (0.1 μM) and DptA (0.1 mM and 1 mM) in 20-day-old males, while AI increased the expression of AttA (0.1 μM) and CecA1 (0.1 μM) in females, suggesting that CD and AI do not cause an anti-inflammatory effect (Figure S5, Tables S12–S14).
Thus, the positive effect of YAP/TAZ inhibitors on lifespan was accompanied by both a decrease (VP) and an increase (CD and AI) in the expression levels of inflammatory markers of aging, suggesting a complex relationship between Yap/Taz signaling and the expression rate of the antimicrobial peptide genes.
2.4. Effects on Wnt Pathway Activity
Previously, wg was shown to repress the expression of decapentaplegic (dpp) morphogen via an Armadillo/dTCF/Brinker complex during Drosophila leg development []. To determine the effect of YAP/TAZ inhibitors on Wnt pathway activity, the expression level of the dpp gene was analyzed.
Three-way ANOVA (gene × concentration × age) followed by a post hoc Duncan test revealed a significant (p < 0.05) increase in the expression level of dpp after ML7 and CD treatment of male flies (Figure S5, Tables S12–S14). The increase in dpp expression level caused by treatment with YAP/TAZ inhibitors was consistent with the decrease in wg expression level. Moreover, post-developmental RNA interference of dpp decreased the mean lifespan of Drosophila, demonstrating its pro-longevity potential [].
3. Discussion
Thus, the effects of pharmacological inhibitors of Yki/Yap/Taz on the survival of D. melanogaster were investigated in this study. The substances with previously established inhibitor effects on Yap/Taz activity, including Verteporfin (VP) [,,], ML-7 hydrochloride (ML7) [], Cytochalasin D (CD) [,,], and AICAR (AI) [] were used.
It has been previously established that evolutionary conservation of both the molecular target and the mechanism of action of a geroprotector increases the probability that the geroprotective effects observed in simpler model organisms will be reproduced in mammals []. The functional conservation of the YAP target was confirmed by early studies showing that human YAP is able to rescue the lethality of flies with the Yki mutation []. Drosophila TEAD/TEF family protein Scalloped (Sc) [] has been shown to be a DNA-binding transcription factor that partners with Yki to mediate the transcriptional output []. The Yki-Sc triggered transcription of target genes has been shown to be conserved in its mammalian homolog [].
VP has been demonstrated to be a suppressor of Yap-TEAD and Yki-Sd interaction in mammalian and fly cells, respectively []. The VP-induced transcriptional changes (decrease in Diap1 mRNA) in Drosophila cells were consistent with the inhibition of the Yki-Sd complex. VP has also been shown to suppress expression of YAP/TAZ transcriptional targets (SOX2, C-MYC, and EGFR) in human patient-derived glioblastoma stem cells and confer significant survival benefit in the Drosophila GBM model [].
Cytoskeletal tension generated through myosin and actin fiber interactions promotes nuclear localization and transcriptional activity of YAP/TAZ [,]. ML7 treatment has been shown to decrease cytoskeletal tension [,] and inhibit YAP/TAZ through myosin light-chain kinase (MLCK) inhibition [,]. Studies in Drosophila have also shown that cytoskeletal tension resulted in increased Yki target gene expression []. Stretchin-Mlck (Strn-MLCK) is the Drosophila ortholog of vertebrate MLCK that shows high sequence homology to the catalytic and regulatory domains of vertebrate MLCK [,] and may be suggested as a potential target for ML7.
Similarly, actin polymerization inhibitor CD has also been demonstrated to inhibit YAP/TAZ nuclear accumulation associated with cytoskeletal tension in mammalian and fly models [,].
AI-mediated AMPK activation has been shown to inhibit YAP through phosphorylation and stabilization angiomotin-like 1 (AMOTL1), an upstream regulator of YAP []. Yki is also regulated by the AMPK in the Drosophila, suggesting its inhibition by AI [].
Thus, the inhibitors used in this study were found to be highly specific for components of the evolutionarily conserved YAP/TAZ pathway.
At the same time, the YAP/TAZ independent effects of these compounds were experimentally confirmed. For example, VP has been demonstrated to promote apoptosis of various tumor cells through increased generation of reactive oxygen radicals, which can result from the inhibition of oxidative phosphorylation [] and binding of free iron [,]. VP can also affect tumor cell proliferation, drug resistance, and tumorigenicity through regulation of the Wnt, PI3K, Ras, mTOR, and NF-κB signaling pathways in various cancer cells []. Remarkably, these effects of VP are specific to tumor cells and do not exhibit any toxicity to normal cells, suggesting that VP may have good geroprotective potential.
Previous studies have demonstrated that activation of MLCK increases permeability of vascular [] and intestinal [] barriers through myosin light chain (MLC) phosphorylation induced contractility of actomyosin and subsequent weakening of endothelial and epithelial cell–cell adhesion, respectively. It was revealed that the maintenance of the barrier’s integrity is one of the hallmarks of health []. Loss of intestinal barrier integrity has been shown to be associated with aging in humans [] and model organisms [,]. At the same time, ML7, a MLCK kinase inhibitor, has been shown to improve vascular endothelial dysfunction in high-fat diet-fed rabbits []. It is worth noting that ML7 treatment prevented an age-dependent increase in the level of expression of the antimicrobial peptide gene DptA (Figure S5C), suggesting the maintenance of intestinal barrier integrity []. It should be taken into account that nonspecific effects of ML7 may also be associated with the inhibition of protein kinase A and protein kinase C, which are involved in a variety of signal transduction pathways [].
The YAP/TAZ independent effects of CD include modulation of the activity calcium channels [], and voltage-dependent sodium channels [], and inhibition of DNA synthesis []. Through inhibition of actin polymerization and induction of actin filaments depolymerization, CD also affects the mechanical properties of the cytoskeleton [] and alters cell motility, adherence, secretion, drug efflux, deformability, morphology, and size [].
The YAP/TAZ independent effects of AI include stimulation of glucose uptake [], increase in fatty acid oxidation [], inhibition of the production of proinflammatory cytokines and mediators [,], and activation of antioxidant effects []. AI has been demonstrated to inhibit autophagy through the regulation of proteasome activity []. Additionally, AI has been found to upregulate autophagy and reduce senescence-associated changes in mesenchymal stromal cells by selectively inhibiting mammalian target of rapamycin complex 1 (mTORC1) in addition to activating AMPK []. AI has been demonstrated to be a naturally occurring metabolic intermediate in the biosynthesis of purine nucleotides [,], which are essential components for the synthesis of biomolecules essential for cellular survival and proliferation, such as DNA, RNA, ATP, and cofactors for a variety of metabolic and signaling enzymes [].
The inhibitory effect of used compounds on the expression level of Yki/Yap/Taz target genes in Drosophila was analyzed using the qRT-PCR test (Figure 2). It should be noted that among the analyzed Yki target genes (Diap1, dally, myc, wg, CycE, vn), not all of them decreased their expression level in response to treatment with inhibitors. This effect may be associated with non-specific activities and tolerance development in response to long-term (10-day) exposure to inhibitors due to possible activation of negative feedback shunts that foster the activation of Yki target genes regardless of Yki inhibition. The decrease in the effect of inhibitors on the expression of target genes after 20 days of treatment also supports the assumption about the development of tolerance to the action of inhibitors.
The set of repressed genes depended on the used inhibitor and on the sex of the fly. Further research is needed to link the change in fly survival to molecular targets. However, according to the obtained results, the increase in survival was most often associated with a decrease in the level of expression of certain genes, namely wg and E2f1, that may be associated with aging and longevity (Table 1).

Table 1.
Overall effects of Drosophila treatment with Yap/Taz inhibitors for 10 days.
For example, the positive effect on survival time in the case of treatment with VP (males: 0.01 μM and 0.1 μM), CD (males: 1 μM), ML7 (females: 1 μM), and AI (males: 1 μM) was accompanied by a decrease in the expression level of the wg gene. On the contrary, an increase in wg expression leads to an increase in mortality in ML7 (1 μM) treated males (Table 1).
The Drosophila wg gene is the structural homolog of vertebrate Wnt genes [], encoding secreted glycoproteins that act as signaling molecules essential for growth, development, and tissue homeostasis [,]. Recently, a new computational approach based on the analysis of networks of co-located loci has identified wg among novel genes associated with longevity in Drosophila []. Experimental data suggest both a positive and a negative role of the Wnt in cell senescence, aging-associated diseases, and organism longevity [,]. For example, reduced Wnt signaling has been shown to protect against mutant Huntingtin toxicity in Drosophila and prolong the lifespan of flies with Huntington’s disease []. In mice, increased age-associated expression of Wnt5a has been correlated with activation of non-canonical Wnt signaling, leading to the aging of hematopoietic stem cells. On the other hand, decreased expression of Wnt5a has been observed to be associated with functional rejuvenation of aged stem-cells []. The opposing roles of Wnt ligands mom-2/Wnt (pro-aging) and lin-44/Wnt (anti-aging) have been revealed in Caenorhabditis elegans []. Meanwhile, studies demonstrating increased Wnt signaling in a Klotho mouse model of accelerated aging [] and ameliorated aging-related tissue fibrosis after treatment with Wnt inhibitors [,] support the pro-aging function of Wnt signaling. Thus, given the dual role of Wnt in aging, the observed mortality-reducing effect may be partly due to its inhibition. We also found that the decrease in wg expression after treatment with YAP/TAZ inhibitors was accompanied by an increase in dpp expression (Figure S5), which demonstrated pro-longevity potential in Drosophila [].
In addition, in CD (0.1 and 1 μM) treated males, a decrease in the level of early and late mortality was accompanied by a decrease in the level of E2f1 expression (Table 1). RNAi mediated knockdown of E2f1 was found to increase lifespan in C. elegans by a FOXO/daf-16 mediated mechanism [].
The process of aging is accompanied by gradual deleterious changes across all levels of biological complexity (molecular, cellular, tissue, and organismal) []. Numerous studies support the idea that age-related death of model organisms and flies in particular could be caused by life-limiting pathologies in critical tissues and organs [,], such as gut [,], fat body [,,,], muscle [], brain [], and neurosecretory cells [,,]. These organs may be considered as potential targets for anti-aging interventions [].
To investigate whether there are tissue and organ-specific effects of YAP/TAZ inhibitors associated with wg and E2f1 suppression, we used the web application FlyAtlas2 []. It should be noted that FlyAtlas2 does not take into account temporal changes in the level of gene expression that occur with age [,,] and therefore has limited application in our study.
It has been found that wg is prominently expressed in the salivary glands and eye of both male and female flies. A direct link between Wnt signaling in imago salivary glands and lifespan has not been established yet. However, wg was found to downregulate Sunspot protein activity, which transactivates E2f1 and proliferating cell nuclear antigen (PCNA) expression for endoreplication in the salivary gland of Drosophila []. It has been previously shown that PCNA, which also participates in DNA damage repair [], is duplicated in the genome of the longest-living mammal bowhead whale (Balaena mysticetus). Further investigations should be conducted to determine the localization of wg gene expression in adult tissues and to elucidate the contribution of PCNA to Drosophila longevity. Thus, the impact of Wnt signaling on the organismal level is likely to be contingent upon a range of variables, such as a tissue-specific environment and genetic background. Nevertheless, the precise role of the Wnt pathway in the positive effect of YAP/TAZ inhibitors has yet to be determined.
The FlyAtlas2 data demonstrate that the E2f1 gene is expressed in all adult tissues, including those which are associated with age-related functional changes, such as the gut, fat body, muscle, and brain []. Overexpression of dFOXO in the adult fat body has been demonstrated to increase Drosophila lifespan [,]. This suggests a potential mechanism of the lifespan-increasing effect of YAP/TAZ inhibition via E2f1-mediated activation of dFOXO in the fat body []. Furthermore, E2F1 has been shown to regulate specific metabolic functions in different organs, contributing to global metabolic homeostasis []. Additionally, E2F1 activity has been found to be augmented during obesity, potentially contributing to some of the associated comorbidities []. Further research is necessary to elucidate the potential role of E2f1 in regulating Drosophila lifespan.
The level of transcription of other genes did not show a clear relationship with the survival of the flies (Table 1). In some groups (males and females treated with 1 μM AI, females treated with 1 μM ML), the decrease in mortality was not associated with changes in the expression level of Yki-target genes, suggesting the presence of additional YAP/TAZ-independent activities of these substances.
Indeed, ML7 is widely used as a myosin light-chain kinase (MLCK) inhibitor [,]. MLCK is activated by numerous physiological factors and inflammatory or angiogenic mediators, inducing actomyosin contraction and causing endothelial hyperpermeability. Aging is considered as a major risk factor for microvascular dysfunction and hyperpermeability []. ML7 has been reported to alleviate advanced glycation end products-induced microvascular hyperpermeability in vivo [], demonstrating the potential to protect against aging. Drosophila suppression of actomyosin contractility via protein kinase A-mediated regulation of MLCK activity has shown to be required for blood–brain barrier integrity [].
AI is one of the most commonly used pharmacological activators of AMP-activated protein kinase (AMPK) []. AMPK is known to control the aging process via an integrated signaling network which affects energy metabolism, autophagic degradation, and stress resistance []. Metformin-induced AMPK activation in C. elegans and mice has been demonstrated to increase lifespan by about 20% and 6%, respectively, in comparison with untreated control animals [].
In addition, aside from Yap/Taz inhibition through preventing YAP/TAZ-TEAD interaction [], VP demonstrates antifibrotic effects by reducing the expression of fibrogenic genes in different models [,].
Contrarily, CD demonstrated pro-aging properties. CD is known as a potent inhibitor of actin cytoskeleton polymerization []. Cytoskeletal integrity was found to be closely related with aging and age-associated diseases [,]. Actin polymerization decreases cell stiffness and leads to cellular aging [], suggesting a possible pro-aging activity of CD.
Despite the finding that Drosophila aging is associated with a highly increased expression of antimicrobial peptide genes [], the results of previous studies on the expression levels of antimicrobial peptide genes after different geroprotective interventions are not unambiguous. While a striking increase in the expression of antimicrobial peptides was observed in glucose-supplemented diet-treated long-lived flies [], selection for longevity reduced the age-dependent increase in Toll signaling-regulated antimicrobial peptide expression []. Knockdown of the Toll receptor [] or knockdown of individual antimicrobial peptides in the Imd pathway has been shown to extend Drosophila lifespan []. The overexpression of antimicrobial peptides may contribute to aging [] or extend lifespan [] of Drosophila, depending on the context of the experiment.
The dual effect of YAP/TAZ inhibitors on the expression of antimicrobial peptide genes (decreased after VP and increased after CD and AI) in long-lived inhibitor-treated flies (Figure S5) demonstrates a complex relationship between antimicrobial peptides and lifespan. In addition, the nonspecific effects of inhibitors on the expression of antimicrobial peptide genes due to the maintenance of intestinal barrier integrity [] should be considered, as well as the involvement of Yki in Toll receptor-mediated Hippo signaling, which is required for the antimicrobial response triggering antimicrobial peptide production in Drosophila [,].
Thus, the results of this study indicate that the administration of Yap/Taz inhibitors increased the lifespan of D. melanogaster, which was dependent on the sex of the fly and the concentration of the substance. Considering the minimal effect of the inhibitors on the expression levels of Yki target genes and the presence of off-target effects, the lifespan effect cannot be uniquely attributed to the Yki pathway. The analysis of changes in gene expression suggests that suppression of the Wnt and E2f1 signaling pathways may be involved in the observed geroprotective effects. Additional research is necessary to elucidate the relationship between the YAP/TAZ pathway and lifespan.
4. Materials and Methods
4.1. Drosophila Strain and Experimental Conditions
Drosophila melanogaster wild-type Canton-S line was obtained from the Drosophila Stock Center at Indiana University (Bloomington, IN, USA). To accelerate the aging, the flies were kept at a temperature of 29 °C, which is a common approach in Drosophila studies []. To maintain constant conditions, Binder KT 115 incubator (Binder, Tuttlingen, Germany) was used. Control end experimental flies were kept on food medium consisting of corn flour—92 g/L, dry yeast—32.1 g/L, agar-agar—5.2 g/L, glucose—136.9 g/L, 8 mL/L—10% solution of Nipagin (methyl 4-hydroxybenzoate, #H5501, Merck, Rahway, NJ, USA) in ethanol, and 5 mL/L of propionic acid (#49685, Merck, Rahway, NJ, USA).
4.2. Treatment with Yap/Taz Inhibitors
Verteporfin (VP, #SML0534, Merck, USA), ML-7 hydrochloride (ML7, #I2764, Merck, USA), Cytochalasin D (CD, #C8273, Merck, USA), AICAR (AI, #A9978, Merck, USA) were used as pharmacological inhibitors of Yap/Taz activity. Depending on the solubility of the substances, water, ethanol 96%, or Cyrene (dihydrolevoglucosenone, #807796, Merck, USA) [] were used as solvents to prepare stock solutions. The stock solutions of VP were prepared with 10% (v/v) solution of Cyrene in water, ML7—with 50% (v/v) solution of ethanol in water, CD—with 100% (v/v) ethanol, and AI—with 100% (v/v) water.
A volume of 30 μL of stock solutions were pipetted onto the medium surface of each experimental vial. In a control vial, 30 μL of corresponding solvent were pipetted. The final concentrations of substances in the food media were estimated using blue food dye (Brilliant Blue FCF, Roha Dyechem Ltd., Mumbai, India) as a tracer of stock solution diffusion []. The obtained results demonstrated a 1:30 dilution of the stock solutions and a final concentration of VP—0.01, 0.1, 1, 10 μM, ML7—0.01, 0.1, 1 μM, CD, and AI—0.1, 1 μM.
Treatment with inhibitors was started from the first day of imago life and continued for 10 days for analysis of survival throughout the lifetime for qRT-PCR assay.
4.3. RNA Isolation and Real-Time Quantitative RT-PCR
The expression level of Yki and its target genes (Death-associated inhibitor of apoptosis 1 (Diap1), division abnormally delayed (dally), Myc (myc), wingless (wg), Cyclin E (CycE), vein (vn), and E2F transcription factor 1 (E2f1)) [,], immune-related genes (Attacin-A (AttA) Cecropin A1 (CecA1), Diptericin A (DptA1) [], and target gene repressed by wg signaling (decapentaplegic (dpp)) [,] was measured by the real-time quantitative reverse transcription-polymerase chain reaction (qRT-PCR). RNA was isolated using an Aurum Total RNA Mini kit (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s instructions. RNA concentration was measured using a Quant-iT RNA Assay Kit (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. cDNA was synthesized according to the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA) from the resulting RNA solution. The reaction mixture for the PCR reaction was prepared based on qPCR mix-HS SYBR (Evrogen, Moscow, Russia) and primers (Table S15). The primer design was performed using the QuantPrime online tool []. The polymerase chain reaction was carried out in a CFX96 amplifier (Bio-Rad, Hercules, CA, USA) using the following program: (1) 95 °C for 30 s, (2) 95 °C for 10 s, (3) 60 °C for 30 s, (4) steps 2–3 were repeated 49 times, (5) DNA melting step.
The expression of the studied genes was calculated relative to the expression of the reference genes β-Tubulin at 56D (β-Tubulin) and Ribosomal protein L32 (RpL32) using threshold cycles (Cts). The delta Ct (dCt) values were calculated as the differences in Ct values for target genes and reference genes. The values of Ct were taken from the CFX Manager 3.1 software (Bio-Rad, Hercules, CA, USA). For each experimental group, 20 males and 10 females were used. The experiments were carried out in three biological replicates with three technical replicates in each.
4.4. Analysis of Survival
Survival was assessed using the concentrations of substances affecting the expression levels of Yki target genes. The emerged imagoes were separated by sex (mated males and non-virgin females) and transferred to control and experimental vials within 24 h. Flies were maintained at 29 °C. The dead individuals were recorded daily. The median and maximum (age of 90% mortality) lifespan were calculated and survival curves were plotted. The experiments were performed separately for each sex in two biological replicates (30 flies per vial, 5 vials, 150 flies per group per replicate).
4.5. Statistical Analysis
To determine the statistical significance of differences in the levels of gene expression, multi-factor analysis of variance (ANOVA) with post hoc Duncan’s multiple-range test were used []. The survival curves were created using the Kaplan–Meier method []. The statistical significance of differences between survival curves was evaluated using the log-rank test []. A Fleming–Harrington test was used to estimate differences between control and experimental groups in earlier or later deaths [] The significance of differences in median maximum lifespan was assessed using Fisher’s exact test []. Bonferroni correction was used to adjust for multiple comparisons. Statistical data analysis was performed using the TIBCO Statistica, version 13.3 (TIBCO Software, Palo Alto, CA, USA) and the online application for survival analysis OASIS 2 [].
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms24066006/s1.
Author Contributions
Conceptualization, A.M. and M.V.S.; data curation, D.A.G., N.V.Z. and A.A.G.; formal analysis, D.A.G. and N.V.Z.; funding acquisition, M.V.S.; investigation, N.V.Z., A.A.G. and D.V.K.; methodology, D.A.G., N.V.Z. and A.A.G.; software, D.A.G., N.V.Z. and M.V.S.; supervision, A.M. and M.V.S.; visualization, D.A.G., N.V.Z. and M.V.S.; writing—original draft, D.A.G. and M.V.S.; writing—review and editing, A.M. and M.V.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Russian Science Foundation grant N 22-24-00961, https://rscf.ru/project/22-24-00961/ (accessed on 15 December 2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nature 2018, 561, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Bischof, E.; Scheibye-Knudsen, M.; Siow, R.; Moskalev, A. Longevity medicine: Upskilling the physicians of tomorrow. Lancet Health Longev. 2021, 2, e187–e188. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-K. Antiaging agents: Safe interventions to slow aging and healthy life span extension. Nat. Prod. Bioprospect. 2022, 12, 18. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Barzilai, N. New horizons in life extension, healthspan extension and exceptional longevity. Age Ageing 2022, 51, afac156. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, A.; Guvatova, Z.; Lopes, I.D.A.; Beckett, C.W.; Kennedy, B.K.; De Magalhaes, J.P.; Makarov, A.A. Targeting aging mechanisms: Pharmacological perspectives. Trends Endocrinol. Metab. 2022, 33, 266–280. [Google Scholar] [CrossRef]
- Proshkina, E.N.; Solovev, I.A.; Shaposhnikov, M.V.; Moskalev, A.A. Key Molecular Mechanisms of Aging, Biomarkers, and Potential Interventions. Mol. Biol. 2020, 54, 883–921. [Google Scholar] [CrossRef]
- Fedintsev, A.; Moskalev, A. Stochastic non-enzymatic modification of long-lived macromolecules—A missing hallmark of aging. Ageing Res. Rev. 2020, 62, 101097. [Google Scholar] [CrossRef]
- Selman, M.; Pardo, A. Fibroageing: An ageing pathological feature driven by dysregulated extracellular matrix-cell mechanobiology. Ageing Res. Rev. 2021, 70, 101393. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Nicklett, E.J.; Ferrucci, L. Does Accumulation of Advanced Glycation End Products Contribute to the Aging Phenotype? J. Gerontol. Ser. A 2010, 65A, 963–975. [Google Scholar] [CrossRef]
- Snedeker, J.G.; Gautieri, A. The role of collagen crosslinks in ageing and diabetes—The good, the bad, and the ugly. Muscles Ligaments Tendons J. 2014, 4, 303–308. [Google Scholar] [CrossRef]
- Gautieri, A.; Passini, F.S.; Silván, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Schoenhuber, H.; Redaelli, A.; Berli, M.; et al. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol. 2017, 59, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Stammers, M.; Ivanova, I.M.; Niewczas, I.S.; Segonds-Pichon, A.; Streeter, M.; Spiegel, D.A.; Clark, J. Age-related changes in the physical properties, cross-linking, and glycation of collagen from mouse tail tendon. J. Biol. Chem. 2020, 295, 10562–10571. [Google Scholar] [CrossRef] [PubMed]
- Nash, A.; Notou, M.; Lopez-Clavijo, A.F.; Bozec, L.; de Leeuw, N.H.; Birch, H.L. Glucosepane is associated with changes to structural and physical properties of collagen fibrils. Matrix Biol. Plus 2019, 4, 100013. [Google Scholar] [CrossRef]
- Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Exp. Cell Res. 2016, 343, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.G.; Kim, H.K.; Hwang, E.S. The essential role of TAZ in normal tissue homeostasis. Arch. Pharmacal Res. 2021, 44, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-K.; Jang, J.-W.; Bae, S.-C. DNA binding partners of YAP/TAZ. BMB Rep. 2018, 51, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, K.H. YAP and TAZ feel the force. Nat. Rev. Mol. Cell Biol. 2011, 12, 404–405. [Google Scholar] [CrossRef]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef]
- Yeung, Y.T.; Guerrero-Castilla, A.; Cano, M.; Muñoz, M.F.; Ayala, A.; Argüelles, S. Dysregulation of the Hippo pathway signaling in aging and cancer. Pharmacol. Res. 2019, 143, 151–165. [Google Scholar] [CrossRef]
- Francisco, J.; Del Re, D.P. YAP/TAZ dull the STING of aging. J. Cardiovasc. Aging 2022, 2, 44. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Zhong, W. Regulation and mechanism of YAP/TAZ in the mechanical microenvironment of stem cells (Review). Mol. Med. Rep. 2021, 24, 506. [Google Scholar] [CrossRef]
- Lei, Q.-Y.; Zhang, H.; Zhao, B.; Zha, Z.-Y.; Bai, F.; Pei, X.-H.; Zhao, S.; Xiong, Y.; Guan, K.-L. TAZ Promotes Cell Proliferation and Epithelial-Mesenchymal Transition and Is Inhibited by the Hippo Pathway. Mol. Cell. Biol. 2008, 28, 2426–2436. [Google Scholar] [CrossRef] [PubMed]
- Elster, D.; von Eyss, B. Hippo signaling in regeneration and aging. Mech. Ageing Dev. 2020, 189, 111280. [Google Scholar] [CrossRef] [PubMed]
- Sladitschek-Martens, H.L.; Guarnieri, A.; Brumana, G.; Zanconato, F.; Battilana, G.; Xiccato, R.L.; Panciera, T.; Forcato, M.; Bicciato, S.; Guzzardo, V.; et al. YAP/TAZ activity in stromal cells prevents ageing by controlling cGAS–STING. Nature 2022, 607, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, Y.; Yin, F.; Yu, J.; Silverman, N.; Pan, D. Toll Receptor-Mediated Hippo Signaling Controls Innate Immunity in Drosophila. Cell 2016, 164, 406–419. [Google Scholar] [CrossRef]
- Jho, E.-H. Dual role of YAP: Oncoprotein and tumor suppressor. J. Thorac. Dis. 2018, 10, S3895–S3898. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced Expression of Connective Tissue Growth Factor (CTGF/CCN2) Mediates Collagen Loss in Chronologically Aged Human Skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, N.; Chu, H.Y.; Yu, Y.; Zhang, Z.-K.; Zhang, G.; Zhang, B.-T. Connective Tissue Growth Factor: From Molecular Understandings to Drug Discovery. Front. Cell Dev. Biol. 2020, 8, 593269. [Google Scholar] [CrossRef]
- Prabhakaran, D.; Perel, P.; Roy, A.; Singh, K.; Raspail, L.; Faria-Neto, J.R.; Gidding, S.S.; Ojji, D.; Hakim, F.; Newby, L.K.; et al. Management of Cardiovascular Disease Patients with Confirmed or Suspected COVID-19 in Limited Resource Settings. Glob. Heart 2020, 15, 44. [Google Scholar] [CrossRef]
- Na Kim, Y.; Choe, S.R.; Cho, K.H.; Cho, D.Y.; Kang, J.; Park, C.G.; Lee, H.Y. Resveratrol suppresses breast cancer cell invasion by inactivating a RhoA/YAP signaling axis. Exp. Mol. Med. 2017, 49, e296. [Google Scholar] [CrossRef]
- Park, Y.; Sohn, B.H.; Johnson, R.L.; Kang, M.; Kim, S.B.; Shim, J.; Mangala, L.S.; Kim, J.H.; Yoo, J.E.; Rodriguez-Aguayo, C.; et al. Yes-associated protein 1 and transcriptional coactivator with PDZ-binding motif activate the mammalian target of rapamycin complex 1 pathway by regulating amino acid transporters in hepatocellular carcinoma. Hepatology 2015, 63, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Tang, B.; Wang, C.; Sun, D.; Zhang, R.; Luo, N.; Han, Z.; Liang, R.; Gao, Z.; Wang, L. Metformin mediates resensitivity to 5-fluorouracil in hepatocellular carcinoma via the suppression of YAP. Oncotarget 2016, 7, 46230–46241. [Google Scholar] [CrossRef] [PubMed]
- Spindler, S.R.; Li, R.; Dhahbi, J.M.; Yamakawa, A.; Mote, P.; Bodmer, R.; Ocorr, K.; Williams, R.T.; Wang, Y.; Ablao, K.P. Statin Treatment Increases Lifespan and Improves Cardiac Health in Drosophila by Decreasing Specific Protein Prenylation. PLoS ONE 2012, 7, e39581. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.M.; Pantano, L.; Pronzati, G.; Grasberger, P.; Probst, C.K.; Black, K.E.; Spinney, J.J.; Hariri, L.P.; Nichols, R.; Lin, Y.; et al. Screening for YAP Inhibitors Identifies Statins as Modulators of Fibrosis. Am. J. Respir. Cell Mol. Biol. 2020, 62, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yolitz, J.; Wang, C.; Spangler, E.; Zhan, M.; Zou, S. Aging Studies in Drosophila Melanogaster. Methods Mol. Biol. 2013, 1048, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Min, K.-J. Drosophila melanogaster as a model system in the study of pharmacological interventions in aging. Transl. Med. Aging 2019, 3, 98–103. [Google Scholar] [CrossRef]
- Ogienko, A.A.; Omelina, E.S.; Bylino, O.V.; Batin, M.A.; Georgiev, P.G.; Pindyurin, A.V. Drosophila as a Model Organism to Study Basic Mechanisms of Longevity. Int. J. Mol. Sci. 2022, 23, 11244. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, H.; Wang, B.; Zhang, Y.; Zheng, X.; Shao, B.; Zhuge, Q.; Jin, K. Key Signaling Pathways in Aging and Potential Interventions for Healthy Aging. Cells 2021, 10, 660. [Google Scholar] [CrossRef]
- Huang, J.; Wu, S.; Barrera, J.; Matthews, K.; Pan, D. The Hippo Signaling Pathway Coordinately Regulates Cell Proliferation and Apoptosis by Inactivating Yorkie, the Drosophila Homolog of YAP. Cell 2005, 122, 421–434. [Google Scholar] [CrossRef]
- Hong, W.; Guan, K.-L. The YAP and TAZ transcription co-activators: Key downstream effectors of the mammalian Hippo pathway. Semin. Cell Dev. Biol. 2012, 23, 785–793. [Google Scholar] [CrossRef]
- Li, Y.; Yang, S.; Yang, S. Verteporfin Inhibits the Progression of Spontaneous Osteosarcoma Caused by Trp53 and Rb1 Deficiency in Ctsk-Expressing Cells via Impeding Hippo Pathway. Cells 2022, 11, 1361. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, X.; Feng, W.; Yu, Y.; Jeong, K.; Guo, W.; Lu, Y.; Mills, G.B. Verteporfin inhibits YAP function through up-regulating 14-3-3σ sequestering YAP in the cytoplasm. Am. J. Cancer Res. 2015, 6, 27–37. [Google Scholar] [PubMed]
- Gibault, F.; Bailly, F.; Corvaisier, M.; Coevoet, M.; Huet, G.; Melnyk, P.; Cotelle, P. Molecular Features of the YAP Inhibitor Verteporfin: Synthesis of Hexasubstituted Dipyrrins as Potential Inhibitors of YAP/TAZ, the Downstream Effectors of the Hippo Pathway. ChemMedChem 2017, 12, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.-I.; Itoga, K.; Okano, T.; Yonemura, S.; Sasaki, H. Hippo pathway regulation by cell morphology and stress fibers. Development 2011, 138, 3907–3914. [Google Scholar] [CrossRef]
- Zhao, B.; Li, L.; Wang, L.; Wang, C.-Y.; Yu, J.; Guan, K.-L. Cell detachment activates the Hippo pathway via cytoskeleton reorganization to induce anoikis. Genes Dev. 2012, 26, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Rausch, V.; Hansen, C.G. The Hippo Pathway, YAP/TAZ, and the Plasma Membrane. Trends Cell Biol. 2020, 30, 32–48. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, J.; Hong, H.; Lee, S.H.; Lee, J.-K.; Jung, E.; Kim, J. Actin remodeling confers BRAF inhibitor resistance to melanoma cells through YAP/TAZ activation. EMBO J. 2016, 35, 462–478. [Google Scholar] [CrossRef]
- Mo, J.-S.; Meng, Z.; Kim, Y.C.; Park, H.W.; Hansen, C.; Kim, S.; Lim, D.-S.; Guan, K.-L. Cellular energy stress induces AMPK-mediated regulation of YAP and the Hippo pathway. Nature 2015, 17, 500–510. [Google Scholar] [CrossRef]
- Pocaterra, A.; Romani, P.; Dupont, S. YAP/TAZ functions and their regulation at a glance. J. Cell Sci. 2020, 133, jcs230425. [Google Scholar] [CrossRef]
- Cao, X.; Zhao, B. Quantitative Real-Time PCR to Measure YAP/TAZ Activity in Human Cells. In The Hippo Pathway: Methods and Protocols; Hergovich, A., Ed.; Springer: New York, NY, USA, 2019; pp. 137–152. [Google Scholar]
- Ziosi, M.; Baena-López, L.A.; Grifoni, D.; Froldi, F.; Pession, A.; Garoia, F.; Trotta, V.; Bellosta, P.; Cavicchi, S.; Pession, A. dMyc Functions Downstream of Yorkie to Promote the Supercompetitive Behavior of Hippo Pathway Mutant Cells. PLoS Genet. 2010, 6, e1001140. [Google Scholar] [CrossRef]
- Tapon, N.; Harvey, K.F.; Bell, D.W.; Wahrer, D.C.; Schiripo, T.A.; Haber, D.A.; Hariharan, I.K. salvador Promotes Both Cell Cycle Exit and Apoptosis in Drosophila and Is Mutated in Human Cancer Cell Lines. Cell 2002, 110, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Goulev, Y.; Fauny, J.D.; Gonzalez-Marti, B.; Flagiello, D.; Silber, J.; Zider, A. SCALLOPED Interacts with YORKIE, the Nuclear Effector of the Hippo Tumor-Suppressor Pathway in Drosophila. Curr. Biol. 2008, 18, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Ohsawa, S.; Igaki, T. Yorkie ensures robust tissue growth in Drosophila ribosomal protein mutants. Development 2021, 148, dev198705. [Google Scholar] [CrossRef] [PubMed]
- Wittkorn, E.; Sarkar, A.; Garcia, K.; Kango-Singh, M.; Singh, A. The Hippo pathway effector Yki downregulates Wg signaling to promote retinal differentiation in the Drosophila eye. Development 2015, 142, 2002–2013. [Google Scholar] [CrossRef]
- Zhang, J.; Ji, J.-Y.; Yu, M.; Overholtzer, M.; Smolen, G.A.; Wang, R.; Brugge, J.S.; Dyson, N.J.; Haber, D.A. YAP-dependent induction of amphiregulin identifies a non-cell-autonomous component of the Hippo pathway. Nature 2009, 11, 1444–1450. [Google Scholar] [CrossRef]
- Rodríguez, I.; Baena-Lopez, L.A.; Baonza, A. Upregulation of Glypicans in Hippo mutants alters the coordinated activity of morphogens. Fly 2008, 2, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, J.; Zhang, C.; Shang, Y.; Lin, J. YAP-mediated crosstalk between the Wnt and Hippo signaling pathways (Review). Mol. Med. Rep. 2020, 22, 4101–4106. [Google Scholar] [CrossRef]
- Totaro, A.; Castellan, M.; Di Biagio, D.; Piccolo, S. Crosstalk between YAP/TAZ and Notch Signaling. Trends Cell Biol. 2018, 28, 560–573. [Google Scholar] [CrossRef]
- Leader, D.P.; Krause, S.A.; Pandit, A.; Davies, S.A.; Dow, J.A.T. FlyAtlas 2: A new version of the Drosophila melanogaster expression atlas with RNA-Seq, miRNA-Seq and sex-specific data. Nucleic Acids Res. 2017, 46, D809–D815. [Google Scholar] [CrossRef]
- Pletcher, S.D.; Macdonald, S.J.; Marguerie, R.; Certa, U.; Stearns, S.C.; Goldstein, D.B.; Partridge, L. Genome-Wide Transcript Profiles in Aging and Calorically Restricted Drosophila melanogaster. Curr. Biol. 2002, 12, 712–723. [Google Scholar] [CrossRef]
- Zerofsky, M.; Harel, E.; Silverman, N.; Tatar, M. Aging of the innate immune response in Drosophila melanogaster. Aging Cell 2005, 4, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Corbally, M.-K.; Regan, J.C. Fly immunity comes of age: The utility of Drosophila as a model for studying variation in immunosenescence. Front. Aging 2022, 3, 1016962. [Google Scholar] [CrossRef] [PubMed]
- Theisen, H.; Syed, A.; Nguyen, B.T.; Lukacsovich, T.; Purcell, J.; Srivastava, G.P.; Iron, D.; Gaudenz, K.; Nie, Q.; Wan, F.Y.; et al. Wingless Directly Represses DPP Morphogen Expression via an Armadillo/TCF/Brinker Complex. PLoS ONE 2007, 2, e142. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Kang, P.; Hernandez, A.M.; Tatar, M. Activin Signaling Targeted by Insulin/dFOXO Regulates Aging and Muscle Proteostasis in Drosophila. PLoS Genet. 2013, 9, e1003941. [Google Scholar] [CrossRef]
- Moskalev, A.; Chernyagina, E.; Tsvetkov, V.; Fedintsev, A.; Shaposhnikov, M.; Krut’Ko, V.; Zhavoronkov, A.; Kennedy, B.K. Developing criteria for evaluation of geroprotectors as a key stage toward translation to the clinic. Aging Cell 2016, 15, 407–415. [Google Scholar] [CrossRef]
- Campbell, S.; Inamdar, M.; Rodrigues, V.; Raghavan, V.; Palazzolo, M.; Chovnick, A. The scalloped gene encodes a novel, evolutionarily conserved transcription factor required for sensory organ differentiation in Drosophila. Genes Dev. 1992, 6, 367–379. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Zheng, Y.; Dong, J.; Pan, D. The TEAD/TEF Family Protein Scalloped Mediates Transcriptional Output of the Hippo Growth-Regulatory Pathway. Dev. Cell 2008, 14, 388–398. [Google Scholar] [CrossRef]
- Liu-Chittenden, Y.; Huang, B.; Shim, J.S.; Chen, Q.; Lee, S.-J.; Anders, R.A.; Liu, J.O.; Pan, D. Genetic and pharmacological disruption of the TEAD-YAP complex suppresses the oncogenic activity of YAP. Genes Dev. 2012, 26, 1300–1305. [Google Scholar] [CrossRef]
- Vigneswaran, K.; Boyd, N.H.; Oh, S.-Y.; Lallani, S.; Boucher, A.; Neill, S.G.; Olson, J.J.; Read, R.D. YAP/TAZ Transcriptional Coactivators Create Therapeutic Vulnerability to Verteporfin in EGFR-mutant Glioblastoma. Clin. Cancer Res. 2021, 27, 1553–1569. [Google Scholar] [CrossRef]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Aragona, M.; Panciera, T.; Manfrin, A.; Giulitti, S.; Michielin, F.; Elvassore, N.; Dupont, S.; Piccolo, S. A Mechanical Checkpoint Controls Multicellular Growth through YAP/TAZ Regulation by Actin-Processing Factors. Cell 2013, 154, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Q.; Kang, S.M.; Eng, G.; Grayson, W.L.; Lu, X.L.; Huo, B.; Gimble, J.; Guo, X.E.; Mow, V.C.; Vunjak-Novakovic, G. Geometric control of human stem cell morphology and differentiation. Integr. Biol. 2010, 2, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, T.; Cosgrove, B.D.; Heo, S.-J.; Shurden, Z.E.; Mauck, R.L. Cytoskeletal to Nuclear Strain Transfer Regulates YAP Signaling in Mesenchymal Stem Cells. Biophys. J. 2015, 108, 2783–2793. [Google Scholar] [CrossRef] [PubMed]
- Rauskolb, C.; Sun, S.; Sun, G.; Pan, Y.; Irvine, K.D. Cytoskeletal Tension Inhibits Hippo Signaling through an Ajuba-Warts Complex. Cell 2014, 158, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.-I.; Mishima, M.; Mabuchi, I.; Hotta, Y. A single Drosophila melanogaster myosin light chain kinase gene produces multiple isoforms whose activities are differently regulated. Genes Cells 1996, 1, 855–871. [Google Scholar] [CrossRef] [PubMed]
- Tohtong, R.; Rodriguez, D.; Maughan, D.; Simcox, A. Analysis of cDNAs encoding Drosophila melanogaster myosin light chain kinase. J. Muscle Res. Cell Motil. 1997, 18, 43–56. [Google Scholar] [CrossRef]
- Sansores-Garcia, L.; Bossuyt, W.; Wada, K.-I.; Yonemura, S.; Tao, C.; Sasaki, H.; Halder, G. Modulating F-actin organization induces organ growth by affecting the Hippo pathway. EMBO J. 2011, 30, 2325–2335. [Google Scholar] [CrossRef]
- DeRan, M.; Yang, J.; Shen, C.-H.; Peters, E.C.; Fitamant, J.; Chan, P.; Hsieh, M.; Zhu, S.; Asara, J.M.; Zheng, B.; et al. Energy Stress Regulates Hippo-YAP Signaling Involving AMPK-Mediated Regulation of Angiomotin-like 1 Protein. Cell Rep. 2014, 9, 495–503. [Google Scholar] [CrossRef]
- Gailite, I.; Aerne, B.L.; Tapon, N. Differential control of Yorkie activity by LKB1/AMPK and the Hippo/Warts cascade in the central nervous system. Proc. Natl. Acad. Sci. USA 2015, 112, E5169–E5178. [Google Scholar] [CrossRef]
- Kuramoto, K.; Yamamoto, M.; Suzuki, S.; Sanomachi, T.; Togashi, K.; Seino, S.; Kitanaka, C.; Okada, M. Verteporfin inhibits oxidative phosphorylation and induces cell death specifically in glioma stem cells. FEBS J. 2020, 287, 2023–2036. [Google Scholar] [CrossRef]
- Delvaux, M.; Hagué, P.; Craciun, L.; Wozniak, A.; Demetter, P.; Schöffski, P.; Erneux, C.; Vanderwinden, J.-M. Ferroptosis Induction and YAP Inhibition as New Therapeutic Targets in Gastrointestinal Stromal Tumors (GISTs). Cancers 2022, 14, 5050. [Google Scholar] [CrossRef]
- Eales, K.L.; Wilkinson, E.A.; Cruickshank, G.; Tucker, J.H.R.; Tennant, D.A. Verteporfin selectively kills hypoxic glioma cells through iron-binding and increased production of reactive oxygen species. Sci. Rep. 2018, 8, 14358. [Google Scholar] [CrossRef]
- Wei, C.; Li, X. The Role of Photoactivated and Non-Photoactivated Verteporfin on Tumor. Front. Pharmacol. 2020, 11, 557429. [Google Scholar] [CrossRef]
- Shen, Q.; Rigor, R.R.; Pivetti, C.D.; Wu, M.H.; Yuan, S.Y. Myosin light chain kinase in microvascular endothelial barrier function. Cardiovasc. Res. 2010, 87, 272–280. [Google Scholar] [CrossRef]
- Jin, Y.; Blikslager, A.T. The Regulation of Intestinal Mucosal Barrier by Myosin Light Chain Kinase/Rho Kinases. Int. J. Mol. Sci. 2020, 21, 3550. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Kroemer, G. Hallmarks of Health. Cell 2020, 184, 33–63. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Meerveld, B.G.-V. Age-Associated Remodeling of the Intestinal Epithelial Barrier. J. Gerontol. Ser. A 2013, 68, 1045–1056. [Google Scholar] [CrossRef]
- Rera, M.; Clark, R.I.; Walker, D.W. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 21528–21533. [Google Scholar] [CrossRef]
- Rera, M.; Azizi, M.J.; Walker, D.W. Organ-specific mediation of lifespan extension: More than a gut feeling? Ageing Res. Rev. 2012, 12, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, X.; Wan, Y.; Zhou, Q.; Zhu, H.; Wang, Y. Myosin light chain kinase inhibitor ML7 improves vascular endothelial dysfunction via tight junction regulation in a rabbit model of atherosclerosis. Mol. Med. Rep. 2015, 12, 4109–4116. [Google Scholar] [CrossRef] [PubMed]
- Krarup, T.; Jakobsen, L.D.; Jensen, B.S.; Hoffmann, E.K. Na+-K+-2Cl− cotransport in Ehrlich cells: Regulation by protein phosphatases and kinases. Am. J. Physiol. 1998, 275, C239–C250. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.D.; Byerly, L. A Cytoskeletal Mechanism for Ca2+ Channel Metabolic Dependence and Inactivation by Intracellular Ca2+. Neuron 1993, 10, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Cantiello, H.F.; Stow, J.L.; Prat, A.G.; Ausiello, D.A. Actin filaments regulate epithelial Na+ channel activity. Am. J. Physiol. 1991, 261, C882–C888. [Google Scholar] [CrossRef] [PubMed]
- Glushankova, N.A. Effect of cytochalasin D on DNA synthesis in cultured cells. Bull. Exp. Biol. Med. 1986, 101, 564–566. [Google Scholar] [CrossRef]
- Goddette, D.W.; Frieden, C. Actin polymerization. The mechanism of action of cytochalasin D. J. Biol. Chem. 1986, 261, 15974–15980. [Google Scholar] [CrossRef]
- Trendowski, M.; Christen, T.D.; Acquafondata, C.; Fondy, T.P. Effects of cytochalasin congeners, microtubule-directed agents, and doxorubicin alone or in combination against human ovarian carcinoma cell lines in vitro. BMC Cancer 2015, 15, 632. [Google Scholar] [CrossRef]
- de Laat, M.; Robinson, M.; Gruntmeir, K.; Liu, Y.; Soma, L.; Lacombe, V. AICAR administration affects glucose metabolism by upregulating the novel glucose transporter, GLUT8, in equine skeletal muscle. Vet. J. 2015, 205, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.; Bruce, C.R.; Dyck, D.J. AMP kinase activation with AICAR further increases fatty acid oxidation and blunts triacylglycerol hydrolysis in contracting rat soleus muscle. J. Physiol. 2005, 565, 547–553. [Google Scholar] [CrossRef]
- Giri, S.; Nath, N.; Smith, B.; Viollet, B.; Singh, A.K.; Singh, I. 5-Aminoimidazole-4-Carboxamide-1-β-4-Ribofuranoside Inhibits Proinflammatory Response in Glial Cells: A Possible Role of AMP-Activated Protein Kinase. J. Neurosci. 2004, 24, 479–487. [Google Scholar] [CrossRef]
- Kirchner, J.; Brüne, B.; Namgaladze, D. AICAR inhibits NFκB DNA binding independently of AMPK to attenuate LPS-triggered inflammatory responses in human macrophages. Sci. Rep. 2018, 8, 7801. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, H.; Lu, C.; Shi, K.; Huang, H.; Zheng, Y.; Wang, Y.; Wang, D.; Wang, H.; Huang, W. AICAR, an AMP-Activated Protein Kinase Activator, Ameliorates Acute Pancreatitis-Associated Liver Injury Partially through Nrf2-Mediated Antioxidant Effects and Inhibition of NLRP3 Inflammasome Activation. Front. Pharmacol. 2021, 12, 724514. [Google Scholar] [CrossRef]
- Viana, R.; Aguado, C.; Esteban, I.; Moreno, D.; Viollet, B.; Knecht, E.; Sanz, P. Role of AMP-activated protein kinase in autophagy and proteasome function. Biochem. Biophys. Res. Commun. 2008, 369, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Khorraminejad-Shirazi, M.; Sani, M.; Talaei-Khozani, T.; Dorvash, M.; Mirzaei, M.; Faghihi, M.A.; Monabati, A.; Attar, A. AICAR and nicotinamide treatment synergistically augment the proliferation and attenuate senescence-associated changes in mesenchymal stromal cells. Stem Cell Res. Ther. 2020, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Boutchueng-Djidjou, M.; Collard-Simard, G.; Fortier, S.; Hébert, S.; Kelly, I.; Landry, C.R.; Faure, R.L. The Last Enzyme of the De Novo Purine Synthesis Pathway 5-aminoimidazole-4-carboxamide Ribonucleotide Formyltransferase/IMP Cyclohydrolase (ATIC) Plays a Central Role in Insulin Signaling and the Golgi/Endosomes Protein Network. Mol. Cell. Proteom. 2015, 14, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Daignan-Fornier, B.; Pinson, B. 5-Aminoimidazole-4-carboxamide-1-beta-D-ribofuranosyl 5’-Monophosphate (AICAR), a Highly Conserved Purine Intermediate with Multiple Effects. Metabolites 2012, 2, 292–302. [Google Scholar] [CrossRef]
- De Vitto, H.; Arachchige, D.B.; Richardson, B.C.; French, J.B. The Intersection of Purine and Mitochondrial Metabolism in Cancer. Cells 2021, 10, 2603. [Google Scholar] [CrossRef]
- Ge, H.; Tollner, T.L.; Hu, Z.; Dai, M.; Li, X.; Guan, H.; Shan, D.; Zhang, X.; Lv, J.; Huang, C.; et al. The importance of mitochondrial metabolic activity and mitochondrial DNA replication during oocyte maturation in vitro on oocyte quality and subsequent embryo developmental competence. Mol. Reprod. Dev. 2012, 79, 392–401. [Google Scholar] [CrossRef]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. WNT Signaling in Disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Langton, P.F.; Kakugawa, S.; Vincent, J.-P. Making, Exporting, and Modulating Wnts. Trends Cell Biol. 2016, 26, 756–765. [Google Scholar] [CrossRef]
- Hall, B.S.; Barnett, Y.A.; Crofts, J.J.; Chuzhanova, N. Identification of novel genes associated with longevity in Drosophila melanogaster—A computational approach. Aging 2019, 11, 11244–11267. [Google Scholar] [CrossRef]
- Gruber, J.; Yee, Z.; Tolwinski, N.S. Developmental Drift and the Role of Wnt Signaling in Aging. Cancers 2016, 8, 73. [Google Scholar] [CrossRef]
- Maiese, K.; Li, F.; Chong, Z.Z.; Shang, Y.C. The Wnt signaling pathway: Aging gracefully as a protectionist? Pharmacol. Ther. 2008, 118, 58–81. [Google Scholar] [CrossRef]
- Dupont, P.; Besson, M.-T.; Devaux, J.; Liévens, J.-C. Reducing canonical Wingless/Wnt signaling pathway confers protection against mutant Huntingtin toxicity in Drosophila. Neurobiol. Dis. 2012, 47, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Florian, M.C.; Nattamai, K.J.; Dörr, K.; Marka, G.; Überle, B.; Vas, V.; Eckl, C.; Andrä, I.; Schiemann, M.; Oostendorp, R.A.J.; et al. A canonical to non-canonical Wnt signalling switch in haematopoietic stem-cell ageing. Nature 2013, 503, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Lezzerini, M.; Budovskaya, Y. A dual role of the Wnt signaling pathway during aging in Caenorhabditis elegans. Aging Cell 2013, 13, 8–18. [Google Scholar] [CrossRef]
- Liu, H.; Fergusson, M.M.; Castilho, R.M.; Liu, J.; Cao, L.; Chen, J.; Malide, D.; Rovira, I.I.; Schimel, D.; Kuo, C.J.; et al. Augmented Wnt Signaling in a Mammalian Model of Accelerated Aging. Science 2007, 317, 803–806. [Google Scholar] [CrossRef]
- Hu, H.-H.; Cao, G.; Wu, X.-Q.; Vaziri, N.D.; Zhao, Y.-Y. Wnt signaling pathway in aging-related tissue fibrosis and therapies. Ageing Res. Rev. 2020, 60, 101063. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt Signaling During Aging Alters Muscle Stem Cell Fate and Increases Fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Xie, Q.; Peng, S.; Tao, L.; Ruan, H.; Yang, Y.; Li, T.-M.; Adams, U.; Meng, S.; Bi, X.; Dong, M.-Q.; et al. E2F Transcription Factor 1 Regulates Cellular and Organismal Senescence by Inhibiting Forkhead Box O Transcription Factors. J. Biol. Chem. 2014, 289, 34205–34213. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, H.-Z.; Liu, D.-P. The Four Layers of Aging. Cell Syst. 2015, 1, 180–186. [Google Scholar] [CrossRef]
- Keshavarz, M.; Xie, K.; Schaaf, K.; Bano, D.; Ehninger, D. Targeting the “hallmarks of aging” to slow aging and treat age-related disease: Fact or fiction? Mol. Psychiatry 2022, 28, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Piper, M.D.; Partridge, L. Drosophila as a model for ageing. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2707–2717. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.I.; Salazar, A.; Yamada, R.; Fitz-Gibbon, S.; Morselli, M.; Alcaraz, J.; Rana, A.; Rera, M.; Pellegrini, M.; Ja, W.W.; et al. Distinct Shifts in Microbiota Composition during Drosophila Aging Impair Intestinal Function and Drive Mortality. Cell Rep. 2015, 12, 1656–1667. [Google Scholar] [CrossRef] [PubMed]
- Gáliková, M.; Klepsatel, P. Obesity and Aging in the Drosophila Model. Int. J. Mol. Sci. 2018, 19, 1896. [Google Scholar] [CrossRef]
- Bai, H.; Kang, P.; Tatar, M. Drosophila insulin-like peptide-6 (dilp6) expression from fat body extends lifespan and represses secretion of Drosophila insulin-like peptide-2 from the brain. Aging Cell 2012, 11, 978–985. [Google Scholar] [CrossRef]
- Hwangbo, D.S.; Gersham, B.; Tu, M.-P.; Palmer, M.; Tatar, M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature 2004, 429, 562–566. [Google Scholar] [CrossRef]
- Giannakou, M.E.; Goss, M.; Jacobson, J.; Vinti, G.; Leevers, S.J.; Partridge, L. Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell 2007, 6, 429–438. [Google Scholar] [CrossRef]
- Augustin, H.; Partridge, L. Invertebrate models of age-related muscle degeneration. Biochim. Biophys. Acta 2009, 1790, 1084–1094. [Google Scholar] [CrossRef]
- Omelyanchuk, L.V.; Shaposhnikov, M.V.; Moskalev, A.A. Drosophila nervous system as a target of aging and anti-aging interventions. Front. Genet. 2015, 6, 89. [Google Scholar] [CrossRef]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of Insulin-Producing Neurons in Flies: Growth and Diabetic Phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef]
- Broughton, S.J.; Piper, M.D.W.; Ikeya, T.; Bass, T.M.; Jacobson, J.; Driege, Y.; Martinez, P.; Hafen, E.; Withers, D.J.; Leevers, S.J.; et al. Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands. Proc. Natl. Acad. Sci. USA 2005, 102, 3105–3110. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Broughton, S.; Nässel, D.R. Behavioral Senescence and Aging-Related Changes in Motor Neurons and Brain Neuromodulator Levels Are Ameliorated by Lifespan-Extending Reproductive Dormancy in Drosophila. Front. Cell. Neurosci. 2017, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Krause, S.A.; Overend, G.; Dow, J.A.T.; Leader, D.P. FlyAtlas 2 in 2022: Enhancements to the Drosophila melanogaster expression atlas. Nucleic Acids Res. 2021, 50, D1010–D1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, J.; Zhang, X.; Wu, B.; Liu, X.; Shen, Z. The Spatial Association of Gene Expression Evolves from Synchrony to Asynchrony and Stochasticity with Age. PLoS ONE 2011, 6, e24076. [Google Scholar] [CrossRef]
- Zhan, M.; Yamaza, H.; Sun, Y.; Sinclair, J.; Li, H.; Zou, S. Temporal and spatial transcriptional profiles of aging in Drosophila melanogaster. Genome Res. 2007, 17, 1236–1243. [Google Scholar] [CrossRef]
- Girardot, F.; Lasbleiz, C.; Monnier, V.; Tricoire, H. Specific age related signatures in Drosophila body parts transcriptome. BMC Genom. 2006, 7, 69. [Google Scholar] [CrossRef]
- Taniue, K.; Nishida, A.; Hamada, F.; Sugie, A.; Oda, T.; Ui-Tei, K.; Tabata, T.; Akiyama, T. Sunspot, a link between Wingless signaling and endoreplication in Drosophila. Development 2010, 137, 1755–1764. [Google Scholar] [CrossRef]
- Essers, J.; Theil, A.F.; Baldeyron, C.; van Cappellen, W.A.; Houtsmuller, A.B.; Kanaar, R.; Vermeulen, W. Nuclear Dynamics of PCNA in DNA Replication and Repair. Mol. Cell. Biol. 2005, 25, 9350–9359. [Google Scholar] [CrossRef]
- Denechaud, P.-D.; Fajas, L.; Giralt, A. E2F1, a Novel Regulator of Metabolism. Front. Endocrinol. 2017, 8, 311. [Google Scholar] [CrossRef]
- Lum, M.; Morona, R. Myosin IIA is essential for Shigella flexneri cell-to-cell spread. Pathog. Dis. 2014, 72, 174–187. [Google Scholar] [CrossRef]
- Antoine, T.E.; Shukla, D. Inhibition of Myosin Light Chain Kinase Can be Targeted for the Development of New Therapies against Herpes Simplex Virus Type-1 Infection. Antivir. Ther. 2014, 19, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Oakley, R.; Tharakan, B. Vascular Hyperpermeability and Aging. Aging Dis. 2014, 5, 114–125. [Google Scholar] [CrossRef]
- Wu, F.; Guo, X.; Xu, J.; Wang, W.; Li, B.; Huang, Q.; Su, L.; Xu, Q. Role of myosin light chain and myosin light chain kinase in advanced glycation end product–induced endothelial hyperpermeability in vitro and in vivo. Diabetes Vasc. Dis. Res. 2016, 13, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fetter, R.; Schwabe, T.; Jung, C.; Liu, L.; Steller, H.; Gaul, U. The cAMP effector PKA mediates Moody GPCR signaling in Drosophila blood–brain barrier formation and maturation. Elife 2021, 10, e68275. [Google Scholar] [CrossRef]
- Višnjić, D.; Lalić, H.; Dembitz, V.; Tomić, B.; Smoljo, T. AICAr, a Widely Used AMPK Activator with Important AMPK-Independent Effects: A Systematic Review. Cells 2021, 10, 1095. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef]
- De Haes, W.; Frooninckx, L.; Van Assche, R.; Smolders, A.; Depuydt, G.; Billen, J.; Braeckman, B.P.; Schoofs, L.; Temmerman, L. Metformin promotes lifespan through mitohormesis via the peroxiredoxin PRDX-2. Proc. Natl. Acad. Sci. USA 2014, 111, E2501–E2509. [Google Scholar] [CrossRef]
- Giraud, J.; Molina-Castro, S.; Seeneevassen, L.; Sifré, E.; Izotte, J.; Tiffon, C.; Staedel, C.; Boeuf, H.; Fernandez, S.; Barthelemy, P.; et al. Verteporfin targeting YAP1/TAZ-TEAD transcriptional activity inhibits the tumorigenic properties of gastric cancer stem cells. Int. J. Cancer 2019, 146, 2255–2267. [Google Scholar] [CrossRef]
- Mohede, D.; Bank, R.; Van Driel, M.; De Jong, I. Mp67-14 Anti-Fibrotic Effects of Verteporfin on Myofibroblasts in Peyronie Disease. J. Urol. 2018, 199, e912–e913. [Google Scholar] [CrossRef]
- Shi-Wen, X.; Racanelli, M.; Ali, A.; Simon, A.; Quesnel, K.; Stratton, R.J.; Leask, A. Verteporfin inhibits the persistent fibrotic phenotype of lesional scleroderma dermal fibroblasts. J. Cell Commun. Signal. 2021, 15, 71–80. [Google Scholar] [CrossRef]
- Shoji, K.; Ohashi, K.; Sampei, K.; Oikawa, M.; Mizuno, K. Cytochalasin D acts as an inhibitor of the actin–cofilin interaction. Biochem. Biophys. Res. Commun. 2012, 424, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Cho, M.J.; Yu, W.D.; Kim, M.J.; Kim, S.Y.; Lee, J.H. Links of Cytoskeletal Integrity with Disease and Aging. Cells 2022, 11, 2896. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.-F.; Wong, W.-T. Roles of the actin cytoskeleton in aging and age-associated diseases. Ageing Res. Rev. 2020, 58, 101021. [Google Scholar] [CrossRef] [PubMed]
- Zahn, J.T.; Louban, I.; Jungbauer, S.; Bissinger, M.; Kaufmann, D.; Kemkemer, R.; Spatz, J.P. Age-Dependent Changes in Microscale Stiffness and Mechanoresponses of Cells. Small 2011, 7, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Galenza, A.; Foley, E. A glucose-supplemented diet enhances gut barrier integrity in Drosophila. Biol. Open 2021, 10, bio056515. [Google Scholar] [CrossRef]
- Fabian, D.K.; Garschall, K.; Klepsatel, P.; Santos-Matos, G.; Sucena, E.; Kapun, M.; Lemaitre, B.; Schlötterer, C.; Arking, R.; Flatt, T. Evolution of longevity improves immunity in Drosophila. Evol. Lett. 2018, 2, 567–579. [Google Scholar] [CrossRef]
- Lin, Y.-R.; Parikh, H.; Park, Y. Stress resistance and lifespan enhanced by downregulation of antimicrobial peptide genes in the Imd pathway. Aging 2018, 10, 622–631. [Google Scholar] [CrossRef]
- Badinloo, M.; Nguyen, E.; Suh, W.; Alzahrani, F.; Castellanos, J.; Klichko, V.I.; Orr, W.C.; Radyuk, S.N. Overexpression of antimicrobial peptides contributes to aging through cytotoxic effects in Drosophila tissues. Arch. Insect Biochem. Physiol. 2018, 98, e21464. [Google Scholar] [CrossRef]
- Loch, G.; Zinke, I.; Mori, T.; Carrera, P.; Schroer, J.; Takeyama, H.; Hoch, M. Antimicrobial peptides extend lifespan in Drosophila. PLoS ONE 2017, 12, e0176689. [Google Scholar] [CrossRef]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef]
- Kang, H.-L.; Benzer, S.; Min, K.-T. Life extension in Drosophila by feeding a drug. Proc. Natl. Acad. Sci. USA 2002, 99, 838–843. [Google Scholar] [CrossRef]
- Camp, J.E.; Nyamini, S.B.; Scott, F.J. Cyrene™ is a green alternative to DMSO as a solvent for antibacterial drug discovery against ESKAPE pathogens. RSC Med. Chem. 2020, 11, 111–117. [Google Scholar] [CrossRef]
- Landis, G.N.; Doherty, D.; Tower, J. Analysis of Drosophila melanogaster Lifespan. Methods Mol. Biol. 2020, 2144, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ikmi, A.; Gaertner, B.; Seidel, C.; Srivastava, M.; Zeitlinger, J.; Gibson, M.C. Molecular Evolution of the Yap/Yorkie Proto-Oncogene and Elucidation of Its Core Transcriptional Program. Mol. Biol. Evol. 2014, 31, 1375–1390. [Google Scholar] [CrossRef]
- Shu, Z.; Deng, W.-M. Differential Regulation of Cyclin E by Yorkie-Scalloped Signaling in Organ Development. G3 Genes Genomes Genet. 2017, 7, 1049–1060. [Google Scholar] [CrossRef]
- Jiang, J.; Struhl, G. Complementary and Mutually Exclusive Activities of Decapentaplegic and Wingless Organize Axial Patterning during Drosophila Leg Development. Cell 1996, 86, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, S.; Kwasniewski, M.; Riaño-Pachón, D.M.; Mueller-Roeber, B. QuantPrime—A flexible tool for reliable high-throughput primer design for quantitative PCR. BMC Bioinform. 2008, 9, 465. [Google Scholar] [CrossRef] [PubMed]
- Allen, M. (Ed.) The SAGE Encyclopedia of Communication Research Methods; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2017. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. In Breakthroughs in Statistics; Kotz, S., Johnson, N.L., Eds.; Springer: New York, NY, USA, 1992. [Google Scholar]
- Mantel, N. Evaluation of survival data and two new rank order statistics arising in its consideration. Cancer Chemother. Rep. 1966, 50, 163–170. [Google Scholar] [PubMed]
- Su, Z.; Zhu, M. Is it time for the weighted log-rank test to play a more important role in confirmatory trials? Contemp. Clin. Trials Commun. 2018, 10, A1–A2. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q.; Redden, D.T.; Weindruch, R.; Allison, D.B. Statistical methods for testing effects on “maximum lifespan”. Mech. Ageing Dev. 2004, 125, 629–632. [Google Scholar] [CrossRef]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.-S.; Lee, S.-J.V.; Kim, S. OASIS 2: Online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).