
Brain-Derived Major Glycoproteins Are Possible Biomarkers for Altered Metabolism of Cerebrospinal Fluid in Neurological Diseases
 , ,
, ,
Abstract
:1. Introduction
2. Results
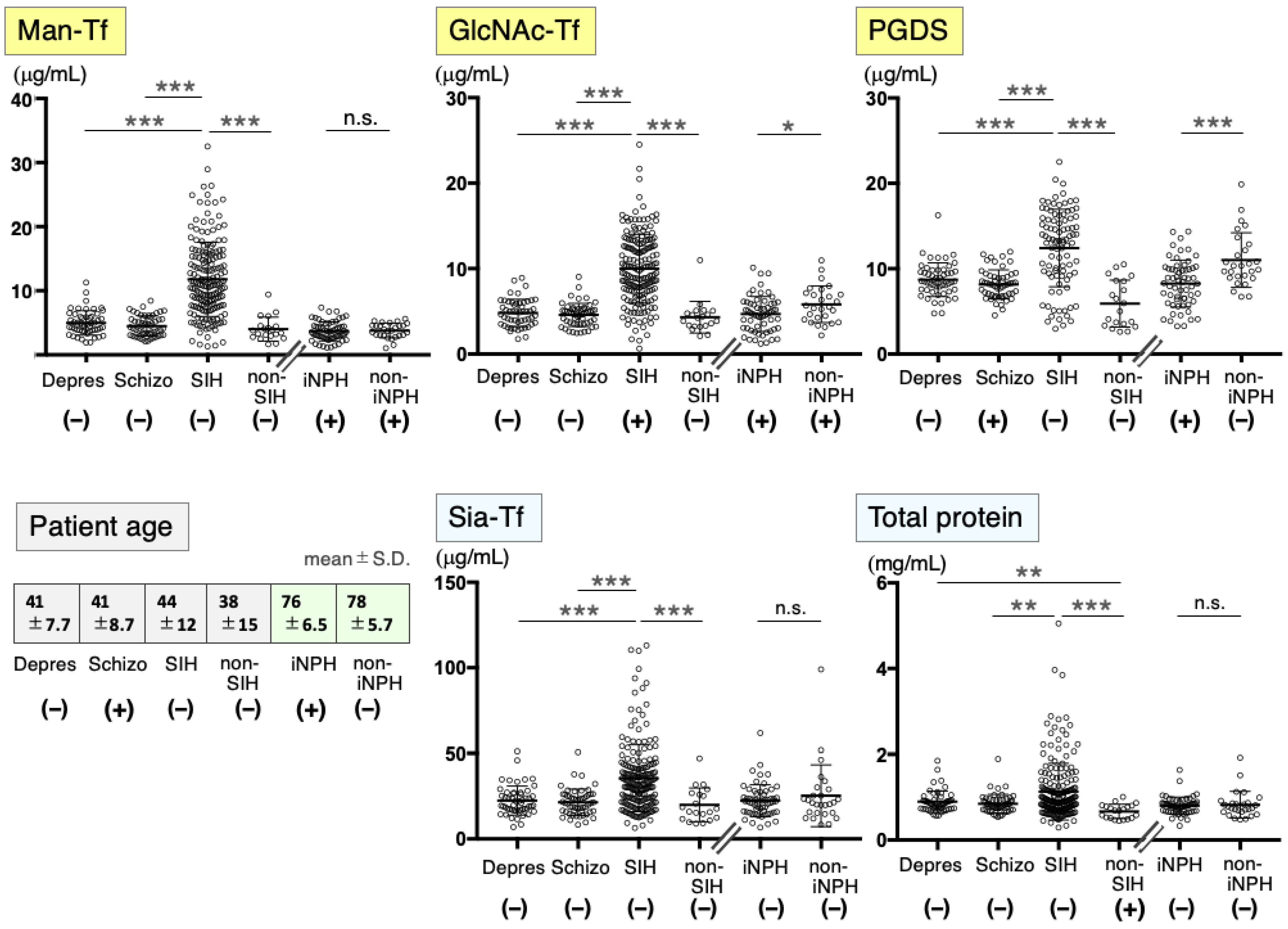
2.1. Marker Levels in CSF from Patients with Neurological Diseases
2.2. CSF Markers in Psychiatric Disease Patients
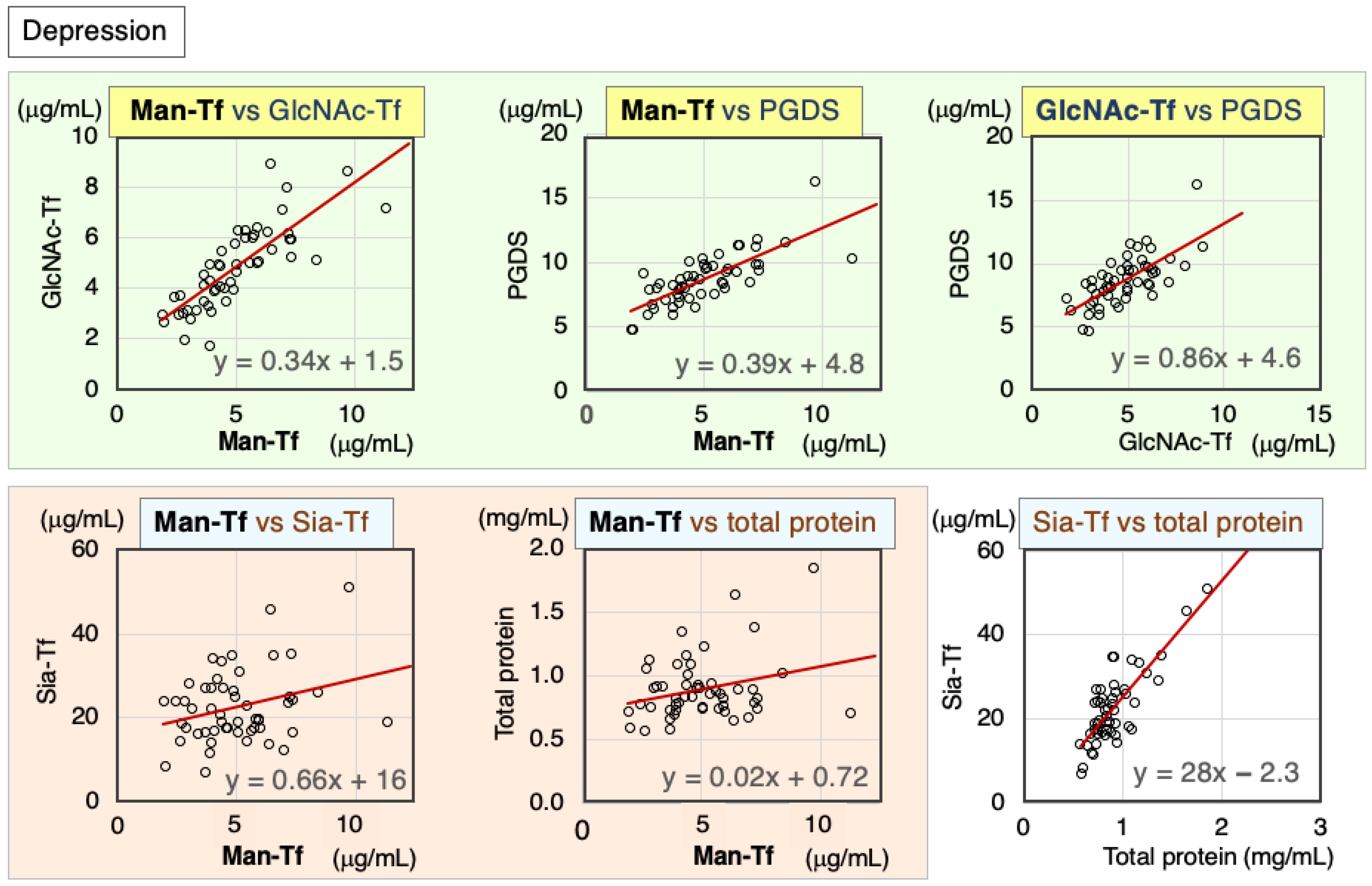
2.2.1. Correlation of CSF Markers in Patients with Depression
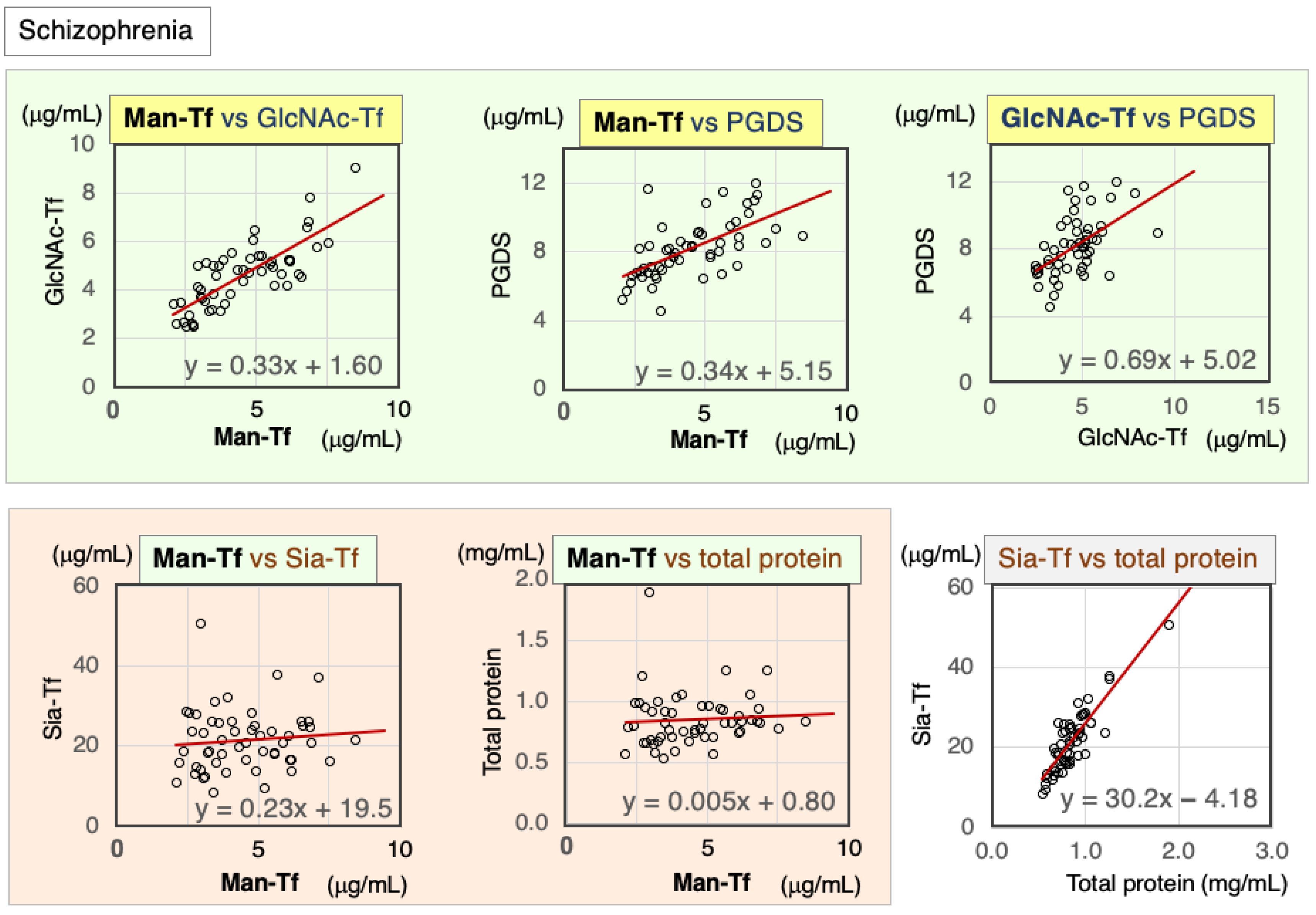
2.2.2. Correlations between CSF Markers in Schizophrenia Patients
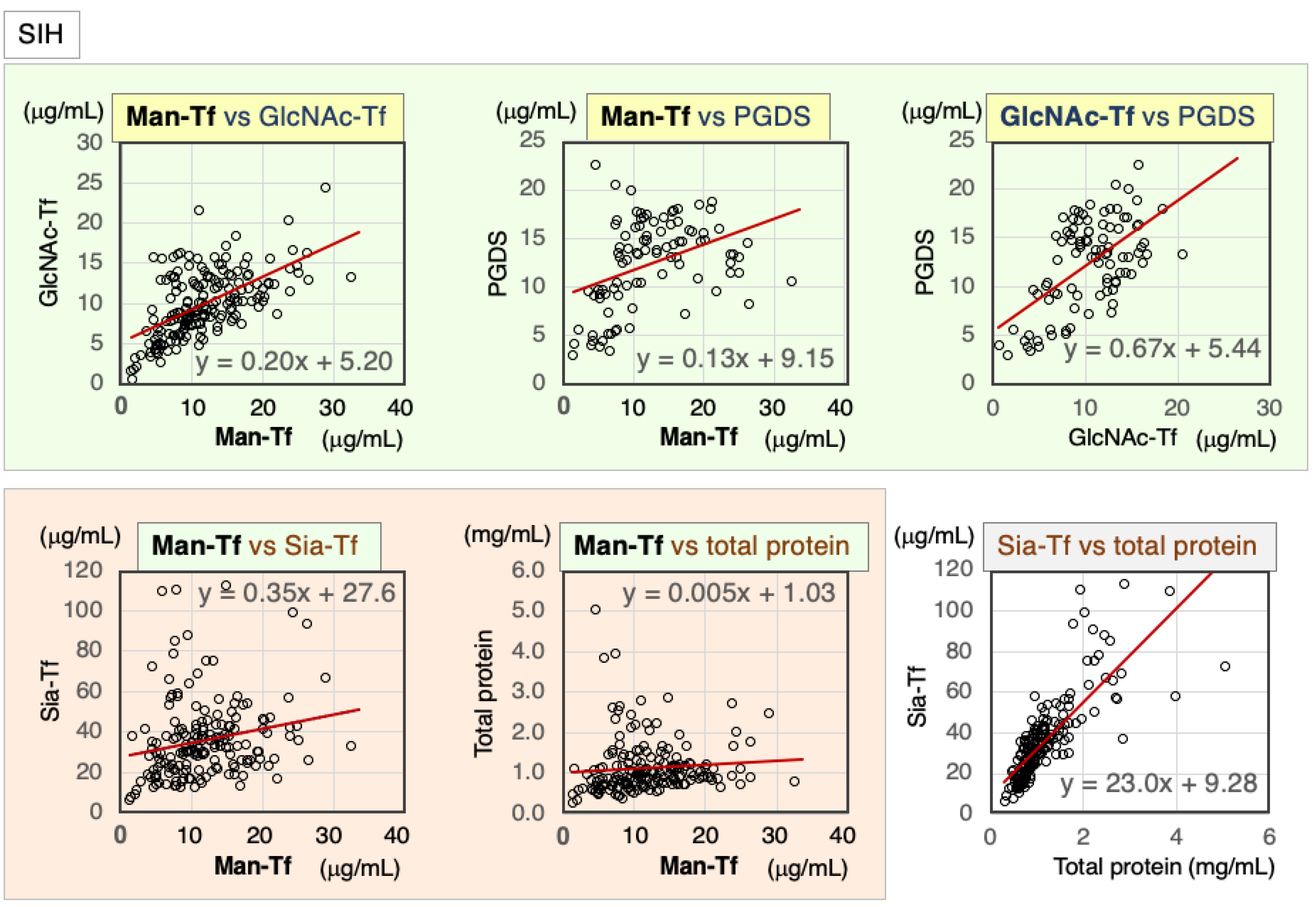
2.3. CSF Markers in SIH
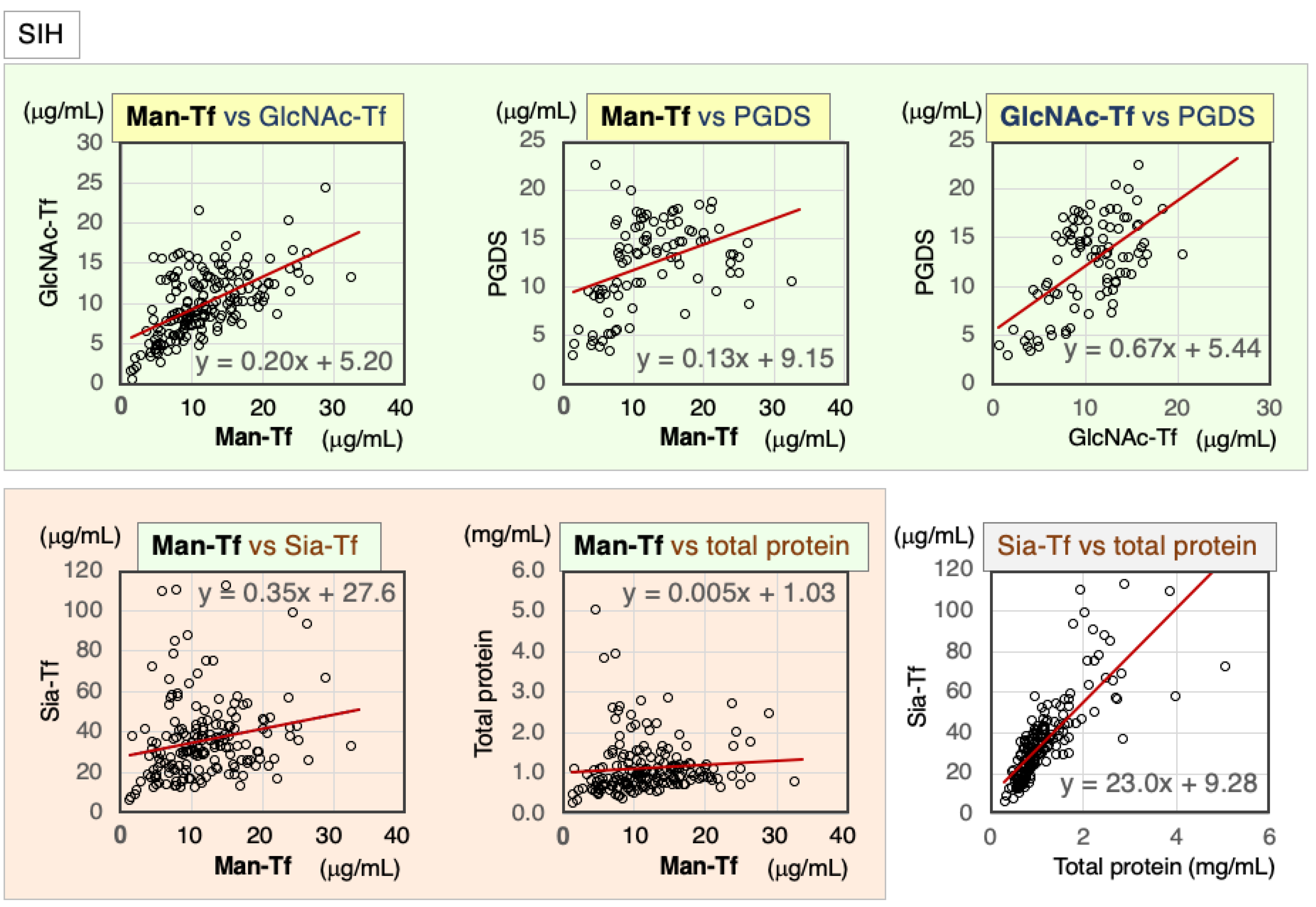
2.3.1. Correlation of CSF Markers in CSF of SIH Patients
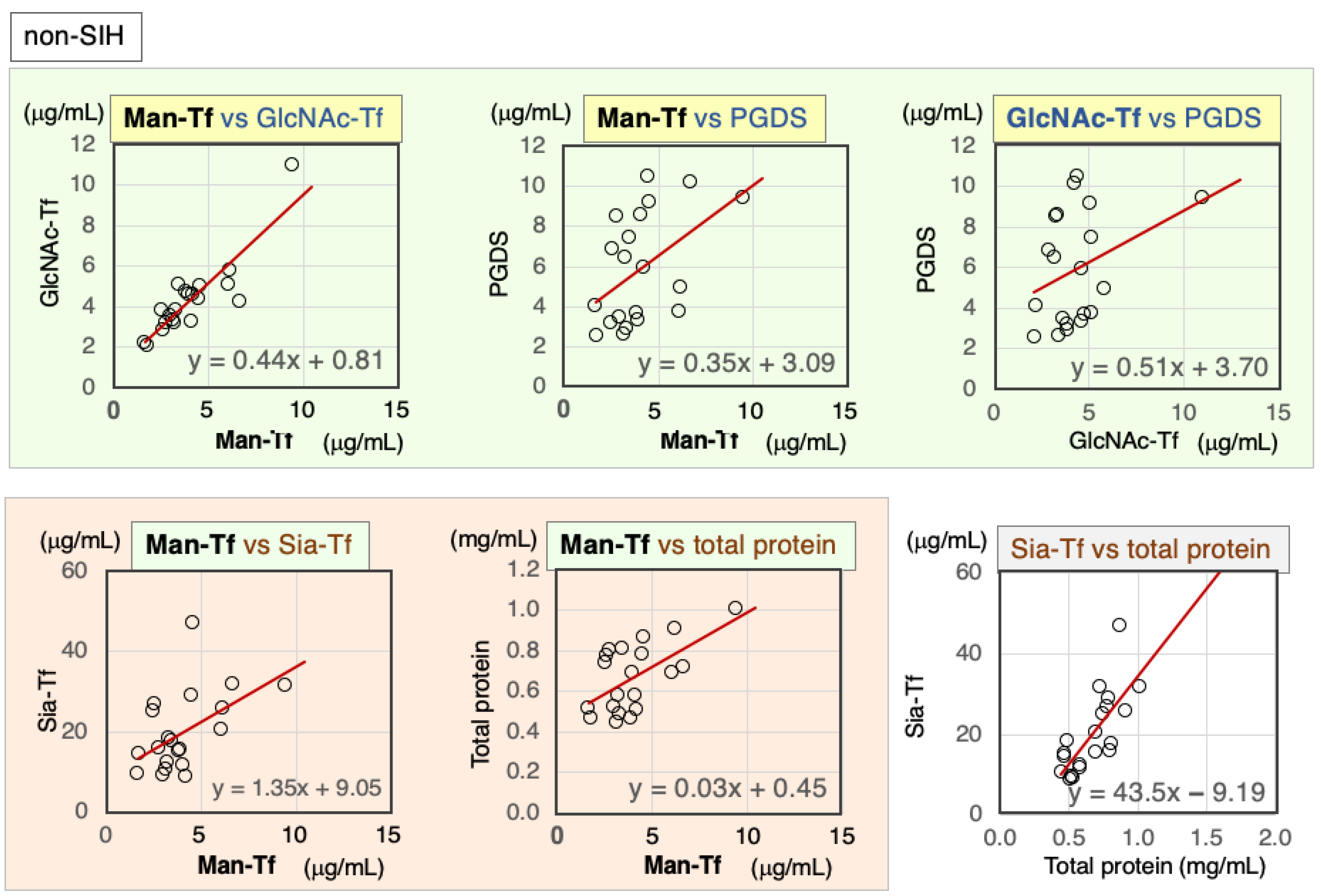
2.3.2. Correlation of Markers in CSF from Non-SIH Patients
2.4. CSF Markers in iNPH
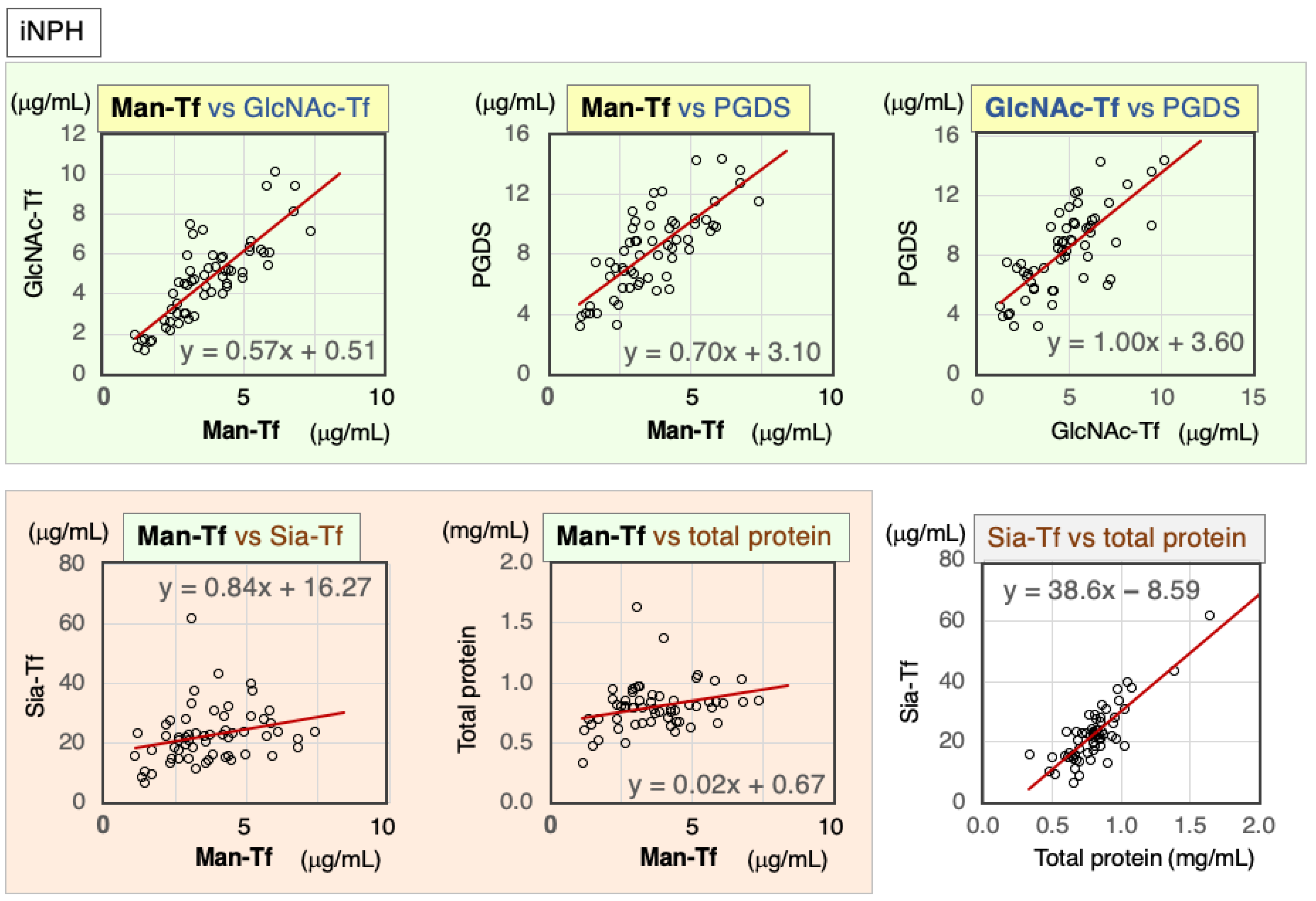
2.4.1. Correlation of Markers in CSF of iNPH Patients
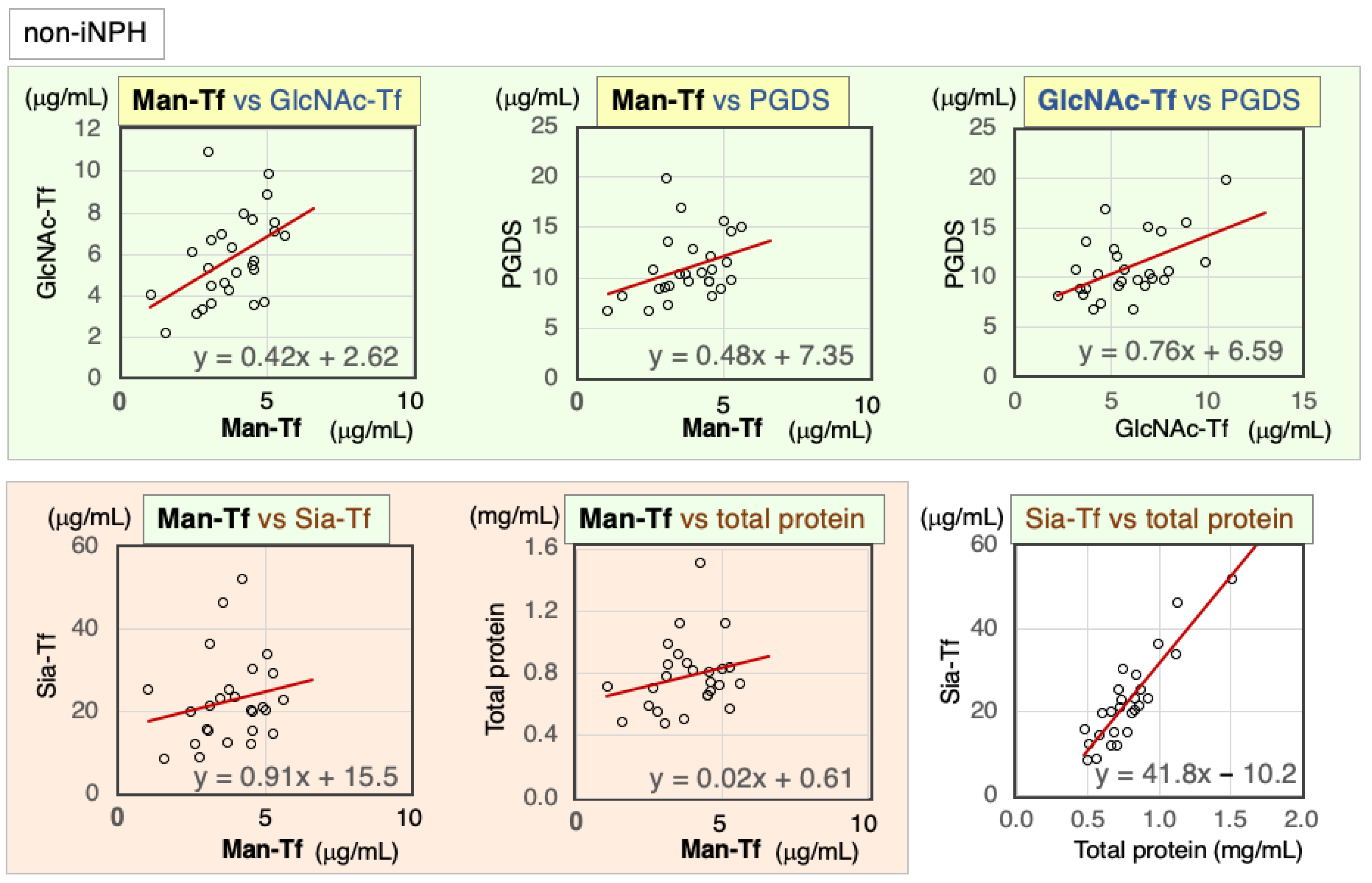
2.4.2. Correlation of CSF Markers in Non-iNPH Patients
2.5. Diagnostic Accuracy of CSF Markers for SIH and iNPH
2.6. CSF Markers after Shunt Operation
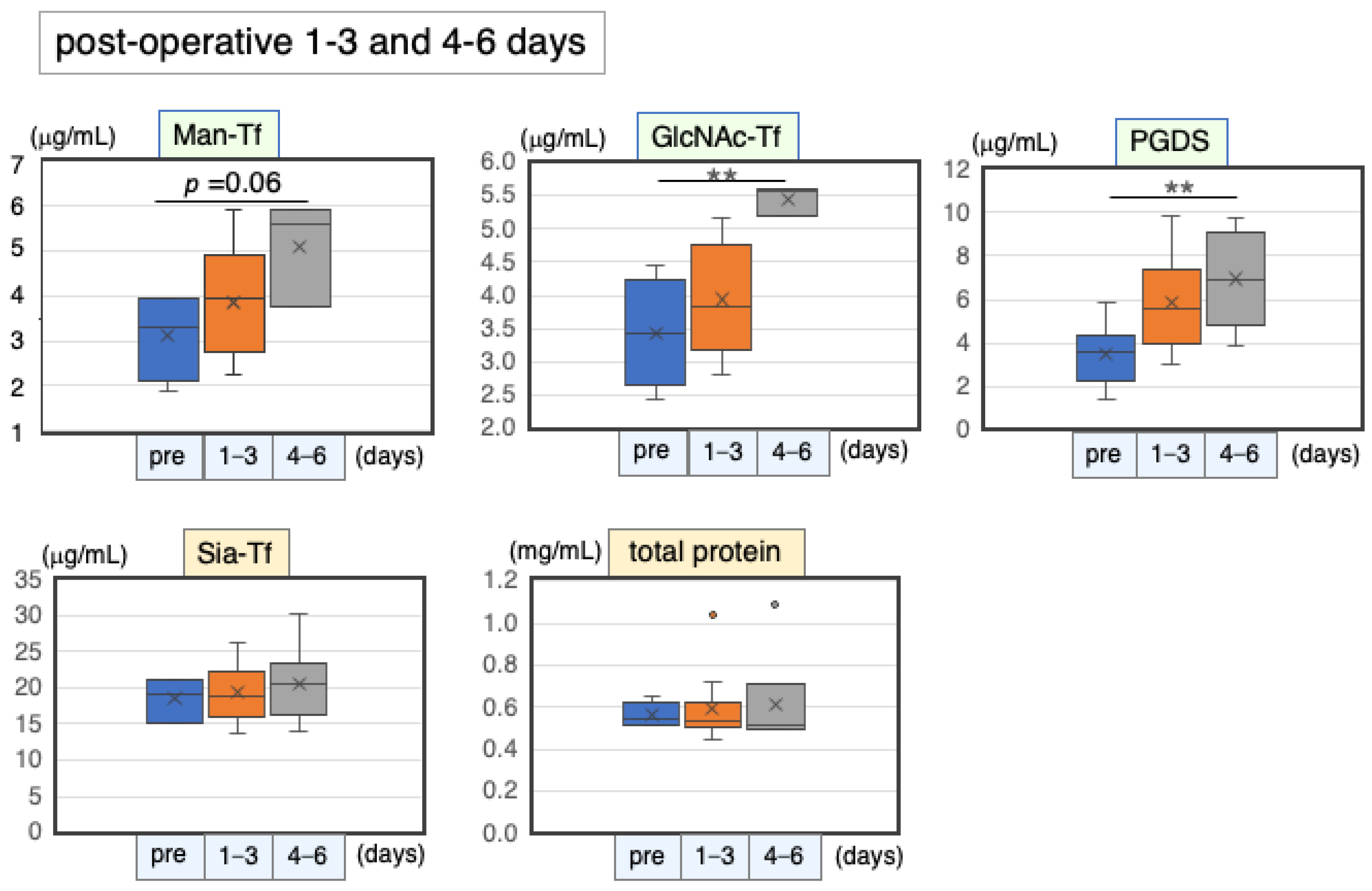
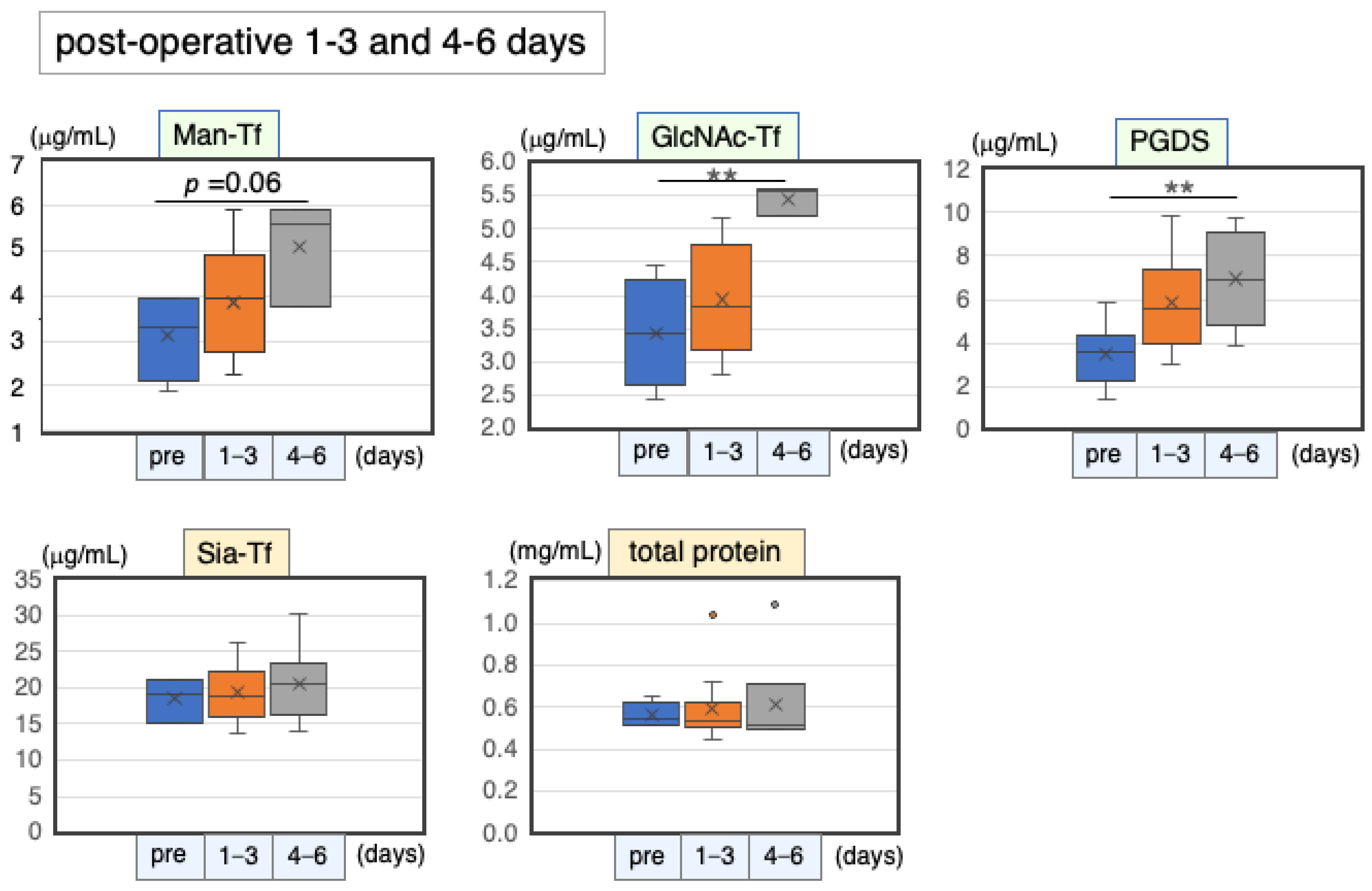
2.6.1. Marker Levels at Post-Operative Days 1–6
2.6.2. Correlation Coefficients of CSF Marker Levels at Postoperative Days 1–6
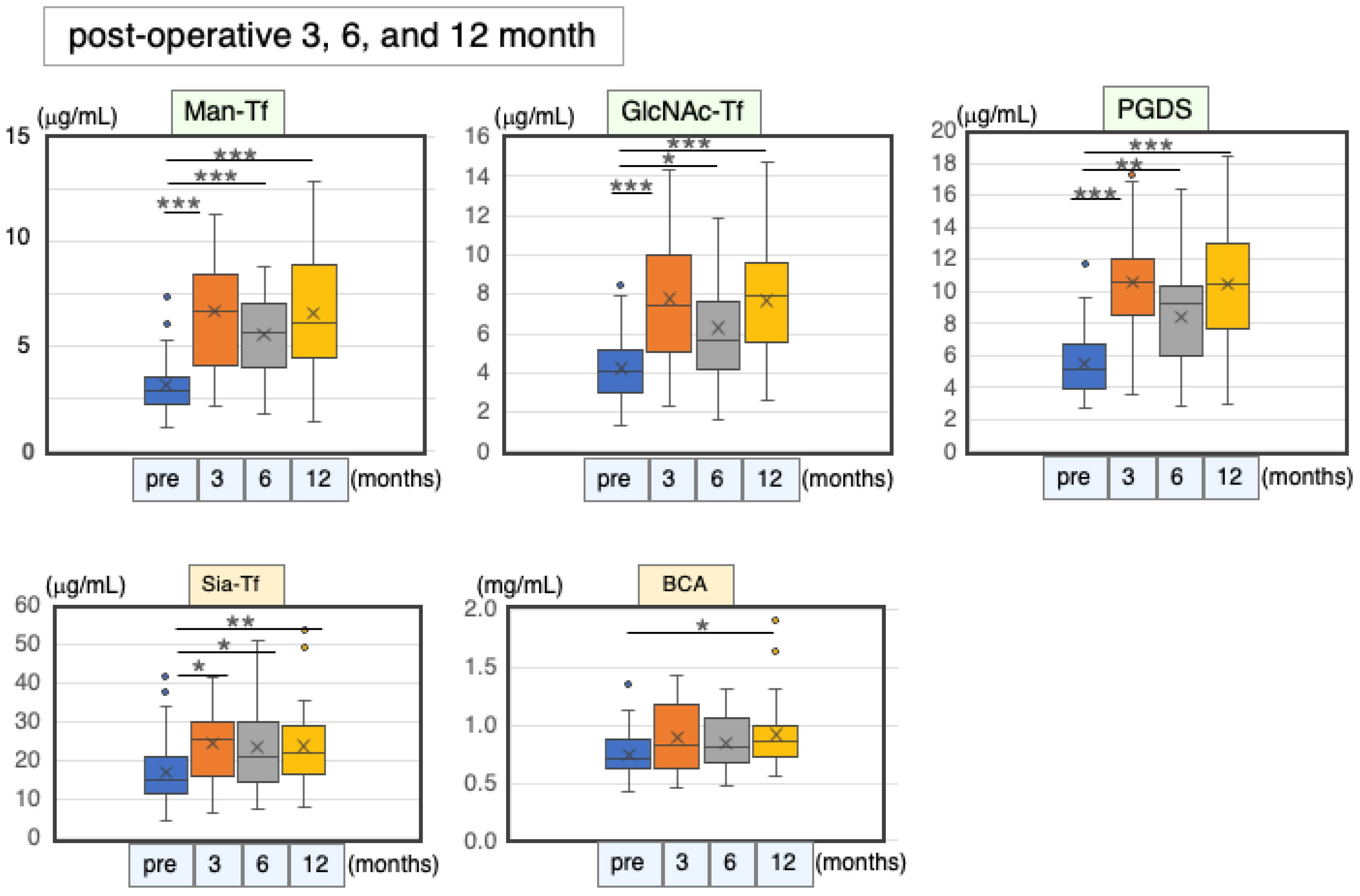
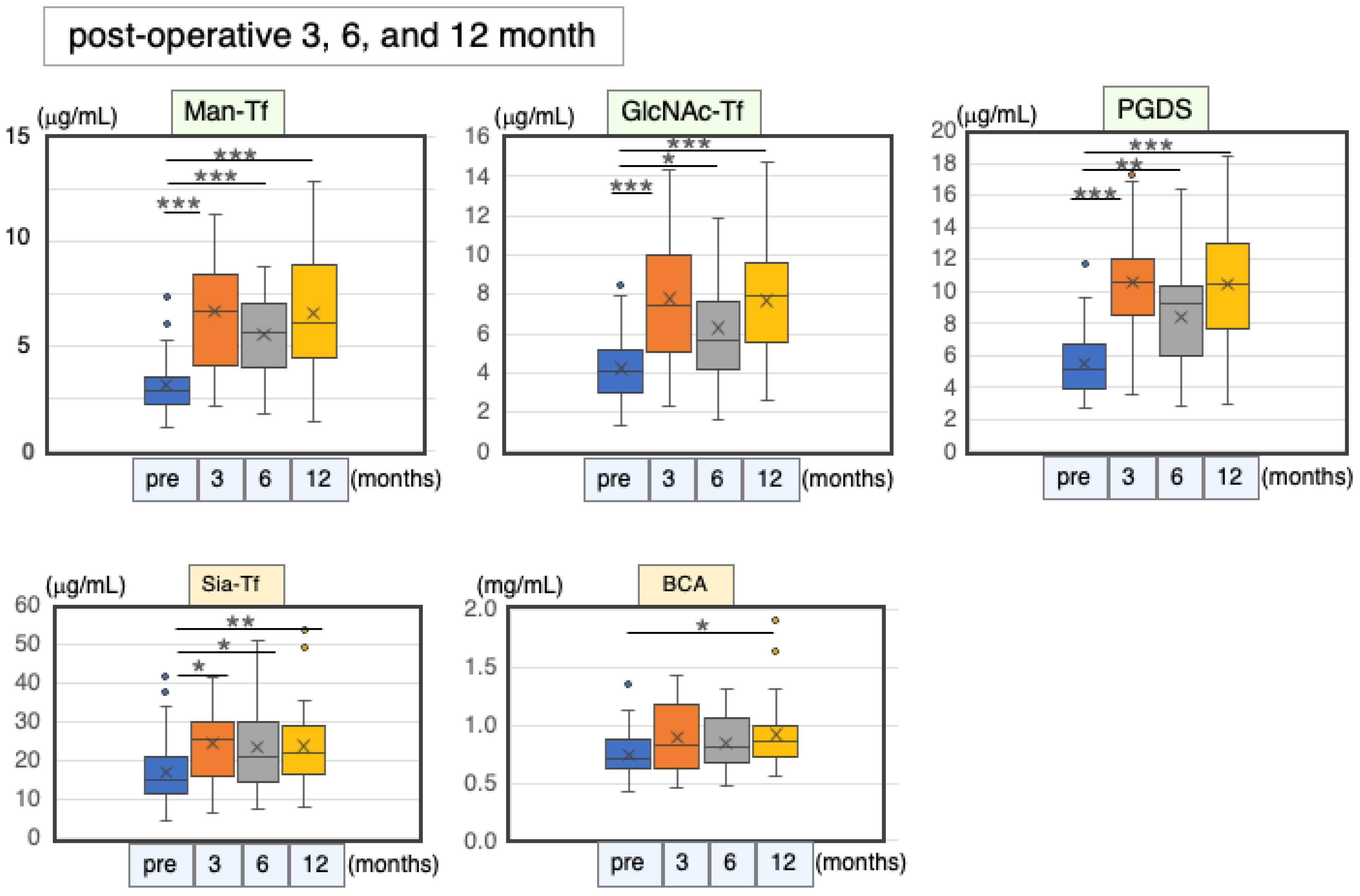
2.6.3. Marker Levels at Post-Operative Months 3, 6 and 12
3. Discussion
4. Materials and Methods
4.1. Patients and CSF Samples
4.2. Quantificatuin of CSF Proteins
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tumani, H.; Huss, A.; Bachhuber, F. The cerebrosinal fluid and barriers—Anatomic and physiologic considerations. Handb. Clin. Neurol. 2017, 146, 21–32. [Google Scholar]
- Cushing, H. The third circulation and its channels. In Harvey Cushing Selected Papers on Neurosurgery; Matson, D.D., Ed.; Yale University Press: New Haven, CT, USA, 1969; pp. 289–319. [Google Scholar]
- Lliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2014, 4, 147ra111. [Google Scholar]
- Jessen, N.A.; Munk, A.S.F.; Lundgaard, I.; Nedergaard, M. The Glymphatic System—A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, Y.; Saito, K.; Ito, H.; Hashimoto, Y. Transferrin isoforms in cerebrospinal fluid and their relation to neurological diseases. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, K.; Kanno, M.; Abe, M.; Murakami, T.; Ugawa, Y.; Goto, A.; Honda, T.; Saito, T.; Saido, T.C.; Yamaguchi, Y.; et al. High correlation among brain-derived major protein levels in cerebrospinal fluid: Implication for amyloid-beta and tau protein changes in Alzheimer’s disease. Metabolites 2022, 12, 355. [Google Scholar] [CrossRef]
- Murakami, Y.; Takahashi, K.; Hoshi, K.; Ito, H.; Kanno, M.; Saito, K.; Nollet, K.; Yamaguchi, Y.; Miyajima, M.; Arai, H.; et al. Spontaneous intracranial hypotension is diagnosed by a combination of lipocalin-type prostaglandin D synthase and brain-type transferrin in cerebrospinal fluid. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.C. Iron homeostasis: Insights from genetics and animal models. Nat. Rev. Genet. 2000, 1, 208–217. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, G.; van Dijk, J.P.; van Eijk, H.G. The biology of transferrin. Clin. Chim. Acta Int. J. Clin. Chem. 1990, 190, 1–46. [Google Scholar] [CrossRef]
- Hoshi, K.; Ito, H.; Abe, E.; Fuwa, T.J.; Kanno, M.; Murakami, Y.; Abe, M.; Murakami, T.; Yoshihara, A.; Ugawa, Y.; et al. Transferrin biosynthesized in the brain is a novel biomarker for Alzheimer’s disease. Metabolites 2021, 11, 616–633. [Google Scholar] [CrossRef]
- Pavone, P.; Pratico, A.D.; Vitaliti, G.; Ruggieri, M.; Rizzo, R.; Parano, E.; Pavone, L.; Pero, G.; Falsaperla, R. Hydranencephaly: Cerebral spinal fluid instead of cerebral mantles. Ital. J. Pediatr. 2014, 40, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, Y.; Matsumoto, Y.; Hoshi, K.; Ito, H.; Fuwa, T.J.; Yamaguchi, Y.; Nakajima, M.; Miyajima, M.; Arai, H.; Nollet, K.; et al. Rapid increase of ‘brain-type’ transferrin in cerebrospinal fluid after shunt surgery for idiopathic normal pressure hydrocephalus: A prognosis marker for cognitive recovery. J. Biochem. 2018, 164, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Urade, Y.; Kitahama, K.; Ohishi, H.; Kaneko, T.; Mizuno, N.; Hayaishi, O. Dominant expression of mRNA for prostaglandin D synthase in leptomeninges, choroid plexus, and oligodendrocytes of the adult rat brain. Proc. Natl. Acad. Sci. USA 1993, 90, 9070–9074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urade, Y. Biochemical and structural characteristics, gene regulation, physiological, pathological and clinical features of lipocalin-type prostaglandin D2 synthase as a multifunctional lipocalin. Front. Physiol. 2021, 12, 718002. [Google Scholar] [CrossRef]
- Schievink, W.I. Spontaneous spinal cerebrospinal fluid leaks and intracranial hypotension. JAMA 2006, 295, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Schievink, W.I.; Dodick, D.W.; Mokri, B.; Silberstein, S.; Bousser, M.G.; Goadsby, P.J. Diagnostic criteria for headache due to spontaneous intracranial hypotension: A perspective. Headache 2011, 51, 1442–1444. [Google Scholar] [CrossRef]
- Olesen, J.; Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211. [Google Scholar]
- Schievink, W.I.; Maya, M.M.; Louy, C.; Moser, F.G.; Tourje, J. Diagnostic criteria for spontaneous spinal CSF leaks and intracranial hypotension. AJNR Am. J. Neuroradiol. 2008, 29, 853–856. [Google Scholar] [CrossRef] [Green Version]
- Monteith, T.S.; Kralik, S.F.; Dillon, W.P.; Hawkins, R.A.; Goadsby, P.J. The utility of radioisotope cisternography in low CSF/volume syndromes compared to myelography. Cephalalgia 2016, 36, 1291–1295. [Google Scholar] [CrossRef]
- Takahashi, K.; Mima, T. Cerebrospinal fluid leakage after radioisotope cisternography is not influenced by needle size at lumbar puncture in patients with intracranial hypotension. Cereb. Fluid Res. 2009, 6, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallia, G.L.; Rigamonti, D.; Williams, M.A. The diagnostic and treatment of idiopathic normal poressure hydrocephalus. Nat. Clin. Pract. Neurol. 2006, 2, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relkin, N.; Marmarou, A.; Klinge, P.; Bergsneider, M.; Black, P.M. Diagnosing idiopathic normal pressure hydrocephalus. Neurosurgery 2005, 57, S4–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goelz, L.; Ruppert, F.-H.; Meier, U.; Lemcke, J. Outcome of modern shunt therapy in patients with idiopathic normal pressure hydrocephalus 6 years postoperatively. J. Neurosug. 2014, 121, 771–775. [Google Scholar] [CrossRef] [Green Version]
- Beuckmann, C.T.; Lazarus, M.; Gerashchenko, D.; Mizoguchi, A.; Nomura, S.; Mohri, I.; Uesugi, A.; Kaneko, T.; Mizuno, N.; Hayaishi, O.; et al. Cellular localization of lipocalin-type prostaglandin D synthase (beta-trace) in the central nervous system of the adult rat. J. Comp. Neurol. 2000, 428, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Futakawa, S.; Nara, K.; Miyajima, M.; Kuno, A.; Ito, H.; Kaji, H.; Shirotani, K.; Honda, T.; Tohyama, Y.; Hoshi, K.; et al. A unique N-glycan on human transferrin in CSF: A possible biomarker for iNPH. Neurobiol. Aging 2012, 33, 1807–1815. [Google Scholar] [CrossRef]
- Solar, P.; Zamani, A.; Kubickova, L.; Dubovy, P.; Joukal, M. Choroid plexus and the blood-cerebrospinal fluid barrier in disease. Fluid Barriers CNS 2020, 17, 35. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, G.D.; Mayo, M.; Saul, T.; Rubenstein, E.; McGuire, D. Alzheimer’s disease, normal-pressure hydrocephalus, and senescent changes in CSF circulatory physiology: A hypothesis. Lancet Neurol. 2003, 2, 506–511. [Google Scholar] [CrossRef]
- Mase, M.; Yamada, K.; Shimazu, N.; Seiki, K.; Oda, H.; Nakau, H.; Inui, T.; Li, W.; Eguchi, N.; Urade, Y. Lipocalin-type prostaglandin D synthase (beta-trace) in cerebrospinal fluid: A useful marker for the diagnosis of normal pressure hydrocephalus. Neurosci. Res. 2003, 47, 455–459. [Google Scholar] [CrossRef]
- Nakajima, M.; Yamada, S.; Miyajima, M.; Ishii, K.; Kuriyama, N.; Kazui, H.; Kanemoto, H.; Suehiro, T.; Yoshiyama, K.; Kameda, M.; et al. Guidelines for management of idiopathic normal pressure hydrocephalus (third edition): Endorsed by the Japanese Society of Normal Pressure Hydrocephalus. Neurol. Med. Chir. 2021, 61, 63–97. [Google Scholar] [CrossRef]
- Shirotani, K.; Futakawa, S.; Nara, K.; Hoshi, K.; Saito, T.; Tohyama, Y.; Kitazume, S.; Yuasa, T.; Miyajima, M.; Arai, H.; et al. High Throughput ELISAs to Measure a Unique Glycan on Transferrin in Cerebrospinal Fluid: A Possible Extension toward Alzheimer’s Disease Biomarker Development. Int. J. Alzheimer’s Dis 2011, 2011, 352787. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depression | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | − | rs | 1.00 | 0.87 ** | 0.77 ** | 0.20 | 0.16 |
| GlcNAc-Tf | + | rs | 0.87 ** | 1.00 | 0.70 ** | 0.19 | 0.13 |
| L-PGDS | − | rs | 0.77 ** | 0.70 ** | 1.00 | 0.44 ** | 0.33 * |
| Sia-Tf | − | rs | 0.20 | 0.19 | 0.44 ** | 1.00 | 0.65 ** |
| Total protein | − | rs | 0.16 | 0.13 | 0.33 * | 0.65 ** | 1.00 |
| Schizophrenia | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | − | rs | 1.00 | 0.76 ** | 0.68 ** | 0.13 | 0.18 |
| GlcNAc-Tf | − | rs | 0.76 ** | 1.00 | 0.55 ** | 0.00 | 0.00 |
| L-PGDS | + | rs | 0.68 ** | 0.55 ** | 1.00 | 0.49 ** | 0.46 ** |
| Sia-Tf | − | rs | 0.13 | 0.00 | 0.49 ** | 1.00 | 0.79 ** |
| Total protein | − | rs | 0.18 | 0.00 | 0.46 ** | 0.79 ** | 1.00 |
| SIH | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | − | rs | 1.00 | 0.57 ** | 0.45 ** | 0.26 ** | 0.27 ** |
| GlcNAc-Tf | + | rs | 0.57 ** | 1.00 | 0.47 ** | 0.70 ** | 0.75 ** |
| L-PGDS | − | rs | 0.45 ** | 0.47 ** | 1.00 | 0.53 ** | 0.57 ** |
| Sia-Tf | − | rs | 0.26 ** | 0.70 ** | 0.53 ** | 1.00 | 0.86 ** |
| Total protein | − | rs | 0.27 ** | 0.75 ** | 0.57 ** | 0.86 ** | 1.00 |
| Non-SIH | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | − | rs | 1.00 | 0.79 ** | 0.55 ** | 0.51 * | 0.44 * |
| GlcNAc-Tf | − | rs | 0.79 ** | 1.00 | 0.27 | 0.46 * | 0.50 * |
| L-PGDS | − | rs | 0.55 * | 0.27 | 1.00 | 0.51 * | 0.67 ** |
| Sia-Tf | + | rs | 0.51 * | 0.46 * | 0.51 * | 1.00 | 0.73 ** |
| Total protein | − | rs | 0.44 * | 0.50 * | 0.67 ** | 0.73 ** | 1.00 |
| iNPH | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | + | rs | 1.00 | 0.82 ** | 0.75 ** | 0.37 ** | 0.29 * |
| GlcNAc-Tf | + | rs | 0.82 ** | 1.00 | 0.73 ** | 0.60 ** | 0.56 ** |
| L-PGDS | + | rs | 0.75 ** | 0.73 ** | 1.00 | 0.46 ** | 0.53 ** |
| Sia-Tf | − | rs | 0.37 ** | 0.60 ** | 0.46 ** | 1.00 | 0.70 ** |
| Total protein | − | rs | 0.29 * | 0.56 ** | 0.53 ** | 0.70 ** | 1.00 |
| Non-iNPH | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | + | rs | 1.00 | 0.48 * | 0.42 ** | 0.25 | 0.25 |
| GlcNAc-Tf | + | rs | 0.48 * | 1.00 | 0.44 * | 0.45 * | 0.43 * |
| L-PGDS | − | rs | 0.42 * | 0.44 * | 1.00 | 0.28 | 0.47 * |
| Sia-Tf | − | rs | 0.25 | 0.45 * | 0.28 | 1.00 | 0.83 ** |
| Total protein | − | rs | 0.25 | 0.43 * | 0.47 * | 0.83 ** | 1.00 |
| Non-SIH vs. SIH | Cutoff (μg/mL) | AUC | Sensitivity (%) | Specificity (%) |
|---|---|---|---|---|
| Man-Tf | 6.4 | 0.98 | 97 | 95 |
| GlcNA-cTf | 9.2 | 0.59 | 77 | 75 |
| L-PGDS | 9.1 | 0.78 | 77 | 75 |
| Sia-Tf | 31.8 | 0.76 | 46 | 95 |
| Total protein | 810 | 0.77 | 62 | 85 |
| Non-iNPH vs. iNPH | Cutoff (μg/mL) | AUC | Sensitivity (%) | Specificity (%) |
|---|---|---|---|---|
| Man-Tf | 6.0 | 0.55 | 62 | 38 |
| GlcNAc-Tf | 3.1 | 0.65 | 72 | 28 |
| L-PGDS | 8.9 | 0.74 | 82 | 57 |
| Sia-Tf | 12.7 | 0.52 | 81 | 19 |
| Total protein | 750 | 0.55 | 46 | 54 |
| Pre-Operation | 1–3 Days | 4–6 Days | ||||||
|---|---|---|---|---|---|---|---|---|
| n | (μg/mL) | n | (μg/mL) | % | n | (μg/mL) | % | |
| Man-Tf | 4 | 6.2 ± 2.0 * | 12 | 7.7 ± 2.3 * | 124 | 3 | 10.2 ± 2.4 * | 163 |
| GlcNAc-Tf | 4 | 3.4 ± 0.8 | 12 | 3.9 ± 0.8 | 115 | 3 | 5.5 ± 0.2 | 159 |
| L-PGDS | 4 | 3.9 ± 1.4 | 12 | 5.8 ± 2.3 | 151 | 8 | 7.0 ± 2.3 | 179 |
| Sia-Tf | 3 | 18.5 ± 3.0 | 12 | 19.4 ± 3.7 | 105 | 6 | 20.6 ± 5.5 | 111 |
| Total protein | 4 | 563 ± 60 | 12 | 593 ± 158 | 105 | 6 | 616 ± 233 | 109 |
| Post-Operative 1–6 Days | Nor. | Correl. Coeff. | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein |
|---|---|---|---|---|---|---|---|
| Man-Tf | + | rs | 1.00 | 0.72 ** | 0.69 ** | 0.18 | −0.19 |
| GlcNAc-Tf | + | rs | 0.72 ** | 1.00 | 0.71 ** | 0.40 | 0.21 |
| L-PGDS | + | rs | 0.69 ** | 0.71 ** | 1.00 | 0.24 | −0.18 |
| Sia-Tf | + | rs | 0.18 | 0.40 | 0.24 | 1.00 | 0.26 |
| Total protein | − | rs | −0.19 | 0.21 | −0.18 | 0.26 | 1.00 |
| Pre-Operation | 3 Months | 6 Months | 12 Months | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | (μg/mL) | n | (μg/mL) | % | n | (μg/mL) | % | n | (μg/mL) | % | |
| Man-Tf | 48 | 6.3 ± 2.5 * | 16 | 13.4 ± 5.4 * | 213 | 26 | 11.1 ± 4.0 * | 176 | 34 | 13.2 ± 5.4 * | 209 |
| GlcNAc-Tf | 48 | 4.2 ± 1.7 | 16 | 7.8 ± 3.4 | 184 | 26 | 6.3 ± 2.8 | 149 | 34 | 7.7 ± 3.1 | 182 |
| L-PGDS | 44 | 5.5 ± 1.9 | 12 | 10.6 ± 3.8 | 193 | 21 | 8.4 ± 3.1 | 154 | 30 | 10.5 ± 3.6 | 192 |
| Sia-Tf | 48 | 17.0 ± 7.5 | 16 | 24.5 ± 9.5 | 144 | 26 | 23.6 ± 11.8 | 139 | 34 | 23.8 ± 9.8 | 140 |
| Total protein | 44 | 750 ± 187 | 13 | 892 ± 313 | 119 | 23 | 840 ± 223 | 112 | 31 | 921 ± 284 | 123 |
| 3 Months | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Protein | |
|---|---|---|---|---|---|---|
| Man-Tf | rs | 1.00 | 0.59 * | 0.64 * | 0.43 | 0.29 |
| GlcNAc-Tf | rs | 0.59 * | 1.00 | 0.62 * | 0.58 * | 0.76 ** |
| L-PGDS | rs | 0.64 * | 0.62 * | 1.00 | 0.32 | 0.57 * |
| Sia-Tf | rs | 0.43 | 0.58 * | 0.32 | 1.00 | 0.64 * |
| Total protein | rs | 0.29 | 0.76 ** | 0.57 * | 0.64 * | 1.00 |
| 6 months | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Pprotein | |
| Man-Tf | r | 1.00 | 0.73 ** | 0.68 ** | 0.51 ** | 0.54 ** |
| GlcNAc-Tf | r | 0.73 ** | 1.00 | 0.70 ** | 0.58 ** | 0.64 ** |
| L-PGDS | r | 0.68 ** | 0.70 ** | 1.00 | 0.42 | 0.39 |
| Sia-Tf | r | 0.51 ** | 0.58 ** | 0.42 | 1.00 | 0.55 ** |
| Total protein | r | 0.54 ** | 0.64 ** | 0.39 | 0.55 ** | 1.00 |
| 12 months | Man-Tf | GlcNAc-Tf | L-PGDS | Sia-Tf | Total Pprotein | |
| Man-Tf | rs | 1.00 | 0.76 ** | 0.74 ** | 0.42 * | 0.23 |
| GlcNAc-Tf | rs | 0.76 ** | 1.00 | 0.76 ** | 0.55 ** | 0.31 |
| L-PGDS | rs | 0.74 ** | 0.76 ** | 1.00 | 0.48 ** | 0.26 |
| Sia-Tf | rs | 0.42 * | 0.55 ** | 0.48 ** | 1.00 | 0.44 * |
| Total protein | rs | 0.23 | 0.31 | 0.26 | 0.44 * | 1.00 |
| Disease | Age * (Years) | Patient Number | Gender (M/F) | Hospital |
|---|---|---|---|---|
| Depression | 41 ± 7.7 | 52 | 24/28 | NCNP |
| Schizophrenia | 41 ± 8.7 | 54 | 27/27 | NCNP |
| SIH: spontaneous intracranial hypotension | 44 ± 12 | 199 | 85/114 | SNO, MTK |
| Non-SIH ** | 33 ± 11 | 20 | 7/13 | SNO, FMU |
| iNPH: idiopathic normal pressure hydrocephalus | 76 ± 6.5 | 60 | 36/24 | JTN, FMU |
| iNPH for time course study | 73 ± 5.7 | 48 | 35/13 | JTN, FMU |
| Non-iNPH | 78 ± 5.7 | 27 | 15/12 | JTN, FMU |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoshi, K.; Kanno, M.; Goto, A.; Ugawa, Y.; Furukawa, K.; Arai, H.; Miyajima, M.; Takahashi, K.; Hattori, K.; Kan, K.; et al. Brain-Derived Major Glycoproteins Are Possible Biomarkers for Altered Metabolism of Cerebrospinal Fluid in Neurological Diseases. Int. J. Mol. Sci. 2023, 24, 6084. https://doi.org/10.3390/ijms24076084
Hoshi K, Kanno M, Goto A, Ugawa Y, Furukawa K, Arai H, Miyajima M, Takahashi K, Hattori K, Kan K, et al. Brain-Derived Major Glycoproteins Are Possible Biomarkers for Altered Metabolism of Cerebrospinal Fluid in Neurological Diseases. International Journal of Molecular Sciences. 2023; 24(7):6084. https://doi.org/10.3390/ijms24076084
Chicago/Turabian StyleHoshi, Kyoka, Mayumi Kanno, Aya Goto, Yoshikazu Ugawa, Katsutoshi Furukawa, Hiroyuki Arai, Masakazu Miyajima, Koichi Takahashi, Kotaro Hattori, Keiichi Kan, and et al. 2023. "Brain-Derived Major Glycoproteins Are Possible Biomarkers for Altered Metabolism of Cerebrospinal Fluid in Neurological Diseases" International Journal of Molecular Sciences 24, no. 7: 6084. https://doi.org/10.3390/ijms24076084