
Molecular Insights into Female Hybrid Sterility in Interspecific Crosses between Drosophila melanogaster and Drosophila simulans
 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Hybrid Ovaries Exhibit Multiple Phenotypic Abnormalities of Oogenesis
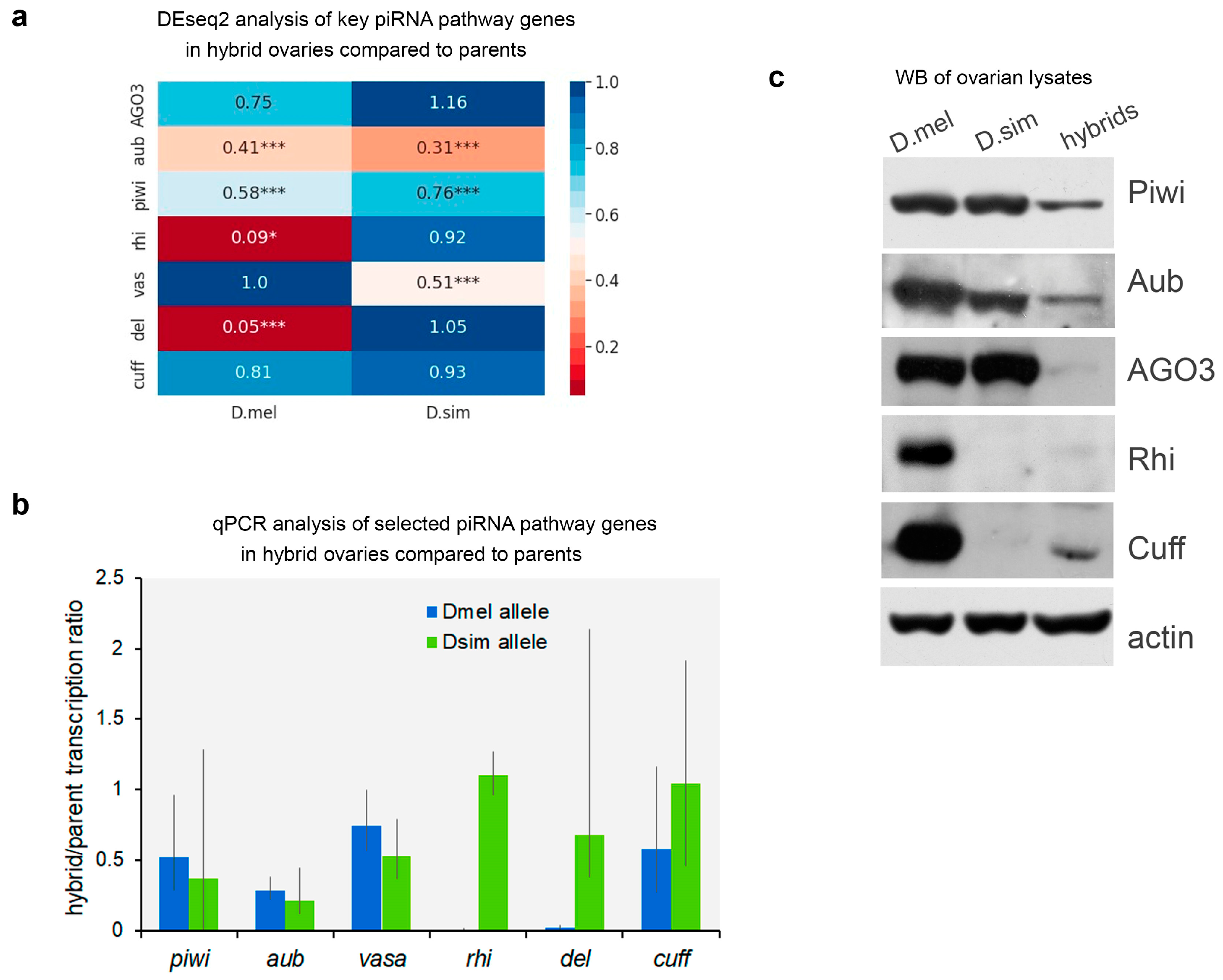
2.2. Differential Gene Expression in Hybrid Ovaries
2.3. Expression of Key piRNA Pathway Factors in Hybrid Ovaries
2.4. RDCsim Complex Acts in Hybrid Ovaries
2.5. Transposon Expression in Hybrid Ovaries
2.6. Biogenesis of TE-Related piRNAs Is Maintained in Hybrid Ovaries
2.7. Vasa Expression in Hybrid Ovaries
3. Discussion
4. Materials and Methods
4.1. Fly Stocks and Genetics
4.2. Immunofluorescence Staining and Confocal Microscopy
4.3. Antibodies
4.4. RT-qPCR Analysis
4.5. RNA Library Preparation
4.6. Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orr, H.A.; Presgraves, D.C. Speciation by postzygotic isolation: Forces, genes and molecules. BioEssays 2000, 22, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004; pp. 1–545. [Google Scholar]
- Sawamura, K.; Yamamoto, M.T. Cytogenetical localization of Zygotic hybrid rescue (Zhr), a Drosophila melanogaster gene that rescues interspecific hybrids from embryonic lethality. Mol. Gen. Genet. 1993, 239, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Ting, C.T.; Tsaur, S.C.; Wu, M.L.; Wu, C.I. A rapidly evolving homeobox at the site of a hybrid sterility gene. Science 1998, 282, 1501–1504. [Google Scholar] [CrossRef]
- Sun, S.; Ting, C.T.; Wu, C.I. The normal function of a speciation gene, Odysseus, and its hybrid sterility effect. Science 2004, 305, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Barbash, D.A.; Roote, J.; Ashburner, M. The Drosophila melanogaster hybrid male rescue gene causes inviability in male and female species hybrids. Genetics 2000, 154, 1747–1771. [Google Scholar] [CrossRef] [PubMed]
- Barbash, D.A.; Ashburner, M. A novel system of fertility rescue in Drosophila hybrids reveals a link between hybrid lethality and female sterility. Genetics 2003, 163, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Phadnis, N.; Orr, H.A. A single gene causes both male sterility and segregation distortion in Drosophila hybrids. Science 2009, 323, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Masly, J.P.; Jones, C.D.; Noor, M.A.; Locke, J.; Orr, H.A. Gene transposition as a cause of hybrid sterility in Drosophila. Science 2006, 313, 1448–1450. [Google Scholar] [CrossRef] [PubMed]
- Araripe, L.O.; Montenegro, H.; Lemos, B.; Hartl, D.L. Fine-scale genetic mapping of a hybrid sterility factor between Drosophila simulans and D. mauritiana: The varied and elusive functions of “speciation genes”. BMC Evol. Biol. 2010, 10, 385. [Google Scholar] [CrossRef]
- Kotov, A.A.; Adashev, V.E.; Godneeva, B.K.; Ninova, M.; Shatskikh, A.S.; Bazylev, S.S.; Aravin, A.A.; Olenina, L.V. piRNA silencing contributes to interspecies hybrid sterility and reproductive isolation in Drosophila melanogaster. Nucleic Acids Res. 2019, 47, 4255–4271. [Google Scholar] [CrossRef]
- Bladen, J.; Cooper, J.C.; Ridges, J.T.; Guo, P.; Phadnis, N. A new hybrid incompatibility locus between Drosophila melanogaster and Drosophila sechellia. Genetics 2024, 226, iyae001. [Google Scholar] [CrossRef] [PubMed]
- Sturtevant, A.H. Genetic Studies on DROSOPHILA SIMULANS. I. Introduction. Hybrids with DROSOPHILA MELANOGASTER. Genetics 1920, 5, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.W.; Roote, J.; Morley, T.; Sawamura, K.; Herrmann, S.; Ashburner, M. Rescue of hybrid sterility in crosses between D. melanogaster and D. simulans. Nature 1996, 380, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Lachaise, D.; Cariou, M.L.; David, J.R.; Lemeunier, F.; Tsacas, L.; Ashburner, M. Historical biogeography of the Drosophila-Melanogaster species subgroup. Evol. Biol. 1988, 22, 159–225. [Google Scholar]
- Russo, C.A.; Takezaki, N.; Nei, M. Molecular phylogeny and divergence times of drosophilid species. Mol. Biol. Evol. 1995, 12, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Subramanian, S.; Kumar, S. Temporal patterns of fruit fly (Drosophila) evolution revealed by mutation clocks. Mol. Biol. Evol. 2004, 21, 36–44. [Google Scholar] [CrossRef]
- Sawamura, K.; Yamamoto, M.T.; Watanabe, T.K. Hybrid lethal systems in the Drosophila melanogaster species complex. II. The Zygotic hybrid rescue (Zhr) gene of D. melanogaster. Genetics 1993, 133, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Ferree, P.M.; Barbash, D.A. Species-specific heterochromatin prevents mitotic chromosome segregation to cause hybrid lethality in Drosophila. PLoS Biol. 2009, 7, e1000234. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, S.; Barbash, D.A. The genetics of hybrid incompatibilities. Annu. Rev. Genet. 2011, 45, 331–355. [Google Scholar] [CrossRef]
- Adashev, V.E.; Kotov, A.A.; Bazylev, S.S.; Shatskikh, A.S.; Aravin, A.A.; Olenina, L.V. Stellate genes and the piRNA pathway in speciation and reproductive isolation of Drosophila melanogaster. Front. Genet. 2021, 11, 610665. [Google Scholar] [CrossRef]
- Presgraves, D.C.; Meiklejohn, C.D. Hybrid Sterility, Genetic Conflict and Complex Speciation: Lessons From the Drosophila simulans Clade Species. Front. Genet. 2021, 12, 669045. [Google Scholar] [CrossRef] [PubMed]
- Orr, H.A. Dobzhansky, Bateson, and the genetics of speciation. Genetics 1996, 144, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Dobzhansky, T. Studies on hybrid sterility. II. Localization of sterility factors in Drosophila Pseudoobscura hybrids. Genetics 1936, 21, 113–135. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. Isolating mechanisms evolution and temperature. Biol. Symp. 1942, 6, 71–125. [Google Scholar]
- Presgraves, D.C. The molecular evolutionary basis of species formation. Nat. Rev. Genet. 2010, 11, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Perez, D.E.; Wu, C.I. Further characterization of the Odysseus locus of hybrid sterility in Drosophila: One gene is not enough. Genetics 1995, 140, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Biémont, C.; Vieira, C. What transposable elements tell us about genome organization and evolution: The case of Drosophila. Cytogenet. Genome Res. 2005, 110, 25–34. [Google Scholar] [CrossRef]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dou, K.; Moon, S.; Tan, F.J.; Zhang, Z.Z. Hijacking oogenesis enables massive propagation of LINE and retroviral transposons. Cell 2018, 174, 1082–1094.e12. [Google Scholar] [CrossRef]
- Kelleher, E.S.; Edelman, N.B.; Barbash, D.A. Drosophila interspecific hybrids phenocopy piRNA-pathway mutants. PLoS Biol. 2012, 10, e1001428. [Google Scholar] [CrossRef]
- Romero-Soriano, V.; Modolo, L.; Lopez-Maestre, H.; Mugat, B.; Pessia, E.; Chambeyron, S.; Vieira, C.; Garcia Guerreiro, M.P. Transposable Element Misregulation Is Linked to the Divergence between Parental piRNA Pathways in Drosophila Hybrids. Genome Biol. Evol. 2017, 9, 1450–1470. [Google Scholar] [CrossRef]
- Banho, C.A.; Oliveira, D.S.; Haudry, A.; Fablet, M.; Vieira, C.; Carareto, C.M.A. Transposable Element Expression and Regulation Profile in Gonads of Interspecific Hybrids of Drosophila arizonae and Drosophila mojavensis wrigleyi. Cells 2021, 10, 3574. [Google Scholar] [CrossRef]
- Castillo, D.M.; Moyle, L.C. Hybrid incompatibility between Drosophila virilis and D. lummei is stronger in the presence of transposable elements. J. Evol. Biol. 2022, 35, 1319–1334. [Google Scholar] [CrossRef]
- Aravin, A.A.; Hannon, G.J.; Brennecke, J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science 2007, 318, 761–764. [Google Scholar] [CrossRef]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Gunawardane, L.S.; Saito, K.; Nishida, K.M.; Miyoshi, K.; Kawamura, Y.; Nagami, T.; Siomi, H.; Siomi, M.C. A slicer-mediated mechanism for repeat-associated siRNA 5’ end formation in Drosophila. Science 2007, 315, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Malone, C.D.; Brennecke, J.; Dus, M.; Stark, A.; McCombie, W.R.; Sachidanandam, R.; Hannon, G.J. Specialized piRNA pathways act in germline and somatic tissues of the Drosophila ovary. Cell 2009, 137, 522–535. [Google Scholar] [CrossRef]
- Gainetdinov, I.; Colpan, C.; Arif, A.; Cecchini, K.; Zamore, P.D. A Single Mechanism of Biogenesis, Initiated and Directed by PIWI Proteins, Explains piRNA Production in Most Animals. Mol. Cell 2018, 71, 775–790.e5. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Welch, J.J.; Kim, K.W.; Jiggins, F.M. Quantifying adaptive evolution in the Drosophila immune system. PLoS Genet. 2009, 5, e1000698. [Google Scholar] [CrossRef]
- Vermaak, D.; Henikoff, S.; Malik, H.S. Positive selection drives the evolution of rhino, a member of the heterochromatin protein 1 family in Drosophila. PLoS Genet. 2005, 1, e9. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.; Wong, A.; Poh, Y.P.; Theurkauf, W.E.; Jensen, J.D. Recurrent and recent selective sweeps in the piRNA pathway. Evolution 2013, 67, 1081–1090. [Google Scholar] [CrossRef]
- Levine, M.T.; Vander Wende, H.M.; Hsieh, E.; Baker, E.P.; Malik, H.S. Recurrent Gene Duplication Diversifies Genome Defense Repertoire in Drosophila. Mol. Biol. Evol. 2016, 33, 1641–1653. [Google Scholar] [CrossRef] [PubMed]
- Rangan, P.; Malone, C.D.; Navarro, C.; Newbold, S.P.; Hayes, P.S.; Sachidanandam, R.; Hannon, G.J.; Lehmann, R. piRNA Production Requires Heterochromatin Formation in Drosophila. Curr. Biol. 2011, 21, 1373–1379. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Schultz, N.; Zhang, F.; Parhad, S.S.; Tu, S.; Vreven, T.; Zamore, P.D.; Weng, Z.; Theurkauf, W.E. The HP1 homolog rhino anchors a nuclear complex that suppresses piRNA precursor splicing. Cell 2014, 157, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Mohn, F.; Sienski, G.; Handler, D.; Brennecke, J. The Rhino-Deadlock-Cutoff Complex Licenses Noncanonical Transcription of Dual-Strand piRNA Clusters in Drosophila. Cell 2014, 157, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.A.; Stuwe, E.; Luo, Y.; Ninova, M.; Le Thomas, A.; Rozhavskaya, E.; Li, S.; Vempati, S.; Laver, J.D.; Patel, D.J.; et al. Cutoff Suppresses RNA Polymerase II Termination to Ensure Expression of piRNA Precursors. Mol. Cell 2016, 63, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Klattenhoff, C.; Xi, H.; Li, C.; Lee, S.; Xu, J.; Khurana, J.S.; Zhang, F.; Schultz, N.; Koppetsch, B.S.; Nowosielska, A.; et al. The Drosophila HP1 homolog Rhino is required for transposon silencing and piRNA production by dual-strand clusters. Cell 2009, 138, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Le Thomas, A.; Stuwe, E.; Li, S.; Du, J.; Marinov, G.; Rozhkov, N.; Chen, Y.C.; Luo, Y.; Sachidanandam, R.; Toth, K.F.; et al. Transgenerationally inherited piRNAs trigger piRNA biogenesis by changing the chromatin of piRNA clusters and inducing precursor processing. Genes Dev. 2014, 28, 1667–1680. [Google Scholar] [CrossRef]
- Parhad, S.S.; Tu, S.; Weng, Z.; Theurkauf, W.E. Adaptive Evolution Leads to Cross-Species Incompatibility in the piRNA Transposon Silencing Machinery. Dev. Cell 2017, 43, 60–70.e5. [Google Scholar] [CrossRef]
- Parhad, S.S.; Theurkauf, W.E. Rapid evolution and conserved function of the piRNA pathway. Open Biol. 2019, 9, 180181. [Google Scholar] [CrossRef] [PubMed]
- Parhad, S.S.; Yu, T.; Zhang, G.; Rice, N.P.; Weng, Z.; Theurkauf, W.E. Adaptive Evolution Targets a piRNA Precursor Transcription Network. Cell Rep. 2020, 30, 2672–2685.e5. [Google Scholar] [CrossRef]
- Lee, Y.C.; Langley, C.H. Long-term and shortterm evolutionary impacts of transposable elements on Drosophila. Genetics 2012, 192, 1411–1432. [Google Scholar] [CrossRef]
- Xie, T.; Spradling, A.C. A niche maintaining germ line stem cells in the Drosophila ovary. Science 2000, 290, 328–330. [Google Scholar] [CrossRef]
- Lobell, A.S.; Kaspari, R.R.; Serrano Negron, Y.L.; Harbison, S.T. The Genetic Architecture of Ovariole Number in Drosophila melanogaster: Genes with Major, Quantitative, and Pleiotropic Effects. G3 2017, 7, 2391–2403. [Google Scholar] [CrossRef] [PubMed]
- de Cuevas, M.; Spradling, A.C. Morphogenesis of the Drosophila fusome and its implications for oocyte specification. Development 1998, 125, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Kirilly, D.; Xie, T. The Drosophila ovary: An active stem cell community. Cell Res. 2007, 17, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.; Gilmutdinov, R.; Wang, L.; Shidlovskii, Y.; Schedl, P. The Drosophila CPEB Protein Orb Specifies Oocyte Fate by a 3’UTR-Dependent Autoregulatory Loop. Genetics 2019, 213, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Mahowald, A.P. Ultrastructural observations on oogenesis in Drosophila. J. Morphol. 1972, 137, 29–48. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Ramat, A.; Simonelig, M. Functions of PIWI Proteins in Gene Regulation: New Arrows Added to the piRNA Quiver. Trends Genet. 2021, 37, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lin, Y.A.; Parhad, S.S.; Jin, Z.; Ma, J.; Theurkauf, W.E.; Zhang, Z.Z.; Huang, Y. Structural insights into Rhino-Deadlock complex for germline piRNA cluster specification. EMBO Rep. 2018, 19, e45418. [Google Scholar] [CrossRef]
- Chen, P.; Kotov, A.A.; Godneeva, B.K.; Bazylev, S.S.; Olenina, L.V.; Aravin, A.A. piRNA-mediated gene regulation and adaptation to sex-specific transposon expression in D. melanogaster male germline. Genes Dev. 2021, 35, 914–935. [Google Scholar] [CrossRef] [PubMed]
- Saint-Leandre, B.; Capy, P.; Hua-Van, A.; Filée, J. piRNA and Transposon Dynamics in Drosophila: A Female Story. Genome Biol. Evol. 2020, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yu, T.; Xu, B.; Hu, Z.; Zhang, X.O.; Theurkauf, W.E.; Weng, Z. Epigenetic and chromosomal features drive transposon insertion in Drosophila melanogaster. Nucleic Acids Res. 2023, 51, 2066–2086. [Google Scholar] [CrossRef] [PubMed]
- Durdevic, Z.; Pillai, R.S.; Ephrussi, A. Transposon silencing in the Drosophila female germline is essential for genome stability in progeny embryos. Life Sci. Alliance 2018, 1, e201800179. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, F.; Kofler, R.; Signor, S. Evolutionary dynamics of piRNA clusters in Drosophila. Mol. Ecol. 2023, 32, 1306–1322. [Google Scholar] [CrossRef] [PubMed]
- Gebert, D.; Neubert, L.K.; Lloyd, C.; Gui, J.; Lehmann, R.; Teixeira, F.K. Large Drosophila germline piRNA clusters are evolutionarily labile and dispensable for transposon regulation. Mol. Cell 2021, 81, 3965–3978.e5. [Google Scholar] [CrossRef]
- Baumgartner, L.; Handler, D.; Platzer, S.W.; Yu, C.; Duchek, P.; Brennecke, J. The Drosophila ZAD zinc finger protein Kipferl guides Rhino to piRNA clusters. eLife 2022, 11, e80067. [Google Scholar] [CrossRef]
- Lerat, E.; Burlet, N.; Biémont, C.; Vieira, C. Comparative analysis of transposable elements in the melanogaster subgroup sequenced genomes. Gene 2011, 473, 100–109. [Google Scholar] [CrossRef]
- Vieira, C.; Fablet, M.; Lerat, E.; Boulesteix, M.; Rebollo, R.; Burlet, N.; Akkouche, A.; Hubert, B.; Mortada, H.; Biémont, C. A comparative analysis of the amounts and dynamics of transposable elements in natural populations of Drosophila melanogaster and Drosophila simulans. J. Environ. Radioact. 2012, 113, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Castillo, D.M.; Barbash, D.A. Moving Speciation Genetics Forward: Modern Techniques Build on Foundational Studies in Drosophila. Genetics 2017, 207, 825–842. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Cassani, M.; Lin, Y.A.; Wang, L.; Dou, K.; Zhang, Z.Z. A Robust Transposon-Endogenizing Response from Germline Stem Cells. Dev. Cell 2018, 47, 660–671.e3. [Google Scholar] [CrossRef] [PubMed]
- Adashev, V.E.; Kotov, A.A.; Olenina, L.V. RNA Helicase Vasa as a Multifunctional Conservative Regulator of Gametogenesis in Eukaryotes. Curr. Issues Mol. Biol. 2023, 45, 5677–5705. [Google Scholar] [CrossRef] [PubMed]
- Durdevic, Z.; Ephrussi, A. Germ cell lineage homeostasis in Drosophila requires the Vasa RNA helicase. Genetics 2019, 213, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Styhler, S.; Nakamura, A.; Swan, A.; Suter, B.; Lasko, P. vasa is required for GURKEN accumulation in the oocyte, and is involved in oocyte differentiation and germline cyst development. Development 1998, 125, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Hutter, P.; Roote, J.; Ashburner, M. A genetic basis for the inviability of hybrids between sibling species of Drosophila. Genetics 1990, 124, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Kibanov, M.V.; Kotov, A.A.; Olenina, L.V. Multicolor fluorescence imaging of whole-mount Drosophila testes for studying spermatogenesis. Anal. Biochem. 2013, 436, 55–64. [Google Scholar] [CrossRef]
- Osouda, S.; Nakamura, Y.; de Saint Phalle, B.; McConnell, M.; Horigome, T.; Sugiyama, S.; Fisher, P.A.; Furukawa, K. Null mutants of Drosophila B-type lamin Dm(0) show aberrant tissue differentiation rather than obvious nuclear shape distortion or specific defects during cell proliferation. Dev. Biol. 2005, 284, 219–232. [Google Scholar] [CrossRef]
- Li, M.A.; Alls, J.D.; Avancini, R.M.; Koo, K.; Godt, D. The large Maf factor Traffic Jam controls gonad morphogenesis in Drosophila. Nat. Cell Biol. 2003, 5, 994–1000. [Google Scholar] [CrossRef]
- Saito, K.; Inagaki, S.; Mituyama, T.; Kawamura, Y.; Ono, Y.; Sakota, E.; Kotani, H.; Asai, K.; Siomi, H.; Siomi, M.C. A regulatory circuit for piwi by the large Maf gene traffic jam in Drosophila. Nature 2009, 461, 1296–1299. [Google Scholar] [CrossRef]
- Nishida, K.M.; Saito, K.; Mori, T.; Kawamura, Y.; Nagami-Okada, T.; Inagaki, S.; Siomi, H.; Siomi, M.C. Gene silencing mechanisms mediated by Aubergine piRNA complexes in Drosophila male gonad. RNA 2007, 13, 1911–1922. [Google Scholar] [CrossRef]
- Radion, E.; Morgunova, V.; Ryazansky, S.; Akulenko, N.; Lavrov, S.; Abramov, Y.; Komarov, P.A.; Glukhov, S.I.; Olovnikov, I.; Kalmykova, A. Key role of piRNAs in telomeric chromatin maintenance and telomere nuclear positioning in Drosophila germline. Epigenetics Chromatin 2018, 11, 40. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Antoniewski, C. Computing siRNA and piRNA overlap signatures. Methods Mol. Biol. 2014, 1173, 135–146. [Google Scholar] [CrossRef]
- Lopez-Delisle, L.; Rabbani, L.; Wolff, J.; Bhardwaj, V.; Backofen, R.; Grüning, B.; Ramírez, F.; Manke, T. pyGenomeTracks: Reproducible plots for multivariate genomic datasets. Bioinformatics 2021, 37, 422–423. [Google Scholar] [CrossRef]
- Waskom, M.L. seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comp. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TE Family | TE-Mapped Antisense piRNAs in D. sim Ovaries, rpm | TE-Mapped Antisense piRNAs in D. mel Ovaries, rpm | TE-Mapped Antisense piRNAs in Hybrid Ovaries, rpm |
|---|---|---|---|
| blood | 5602.2 | 2904.2 | 1112.3 |
| Burdock | 627.6 | 1072.6 | 80.3 |
| gypsy12 | 1574.0 | 1011.6 | 131.7 |
| diver | 1163.3 | 424.4 | 52.6 |
| HMS-Beagle | 1834.4 | 407.8 | 89.5 |
| rover | 51.0 | 1335.5 | 362.7 |
| jockey | 182.3 | 375.3 | 45.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotov, A.A.; Adashev, V.E.; Kombarov, I.A.; Bazylev, S.S.; Shatskikh, A.S.; Olenina, L.V. Molecular Insights into Female Hybrid Sterility in Interspecific Crosses between Drosophila melanogaster and Drosophila simulans. Int. J. Mol. Sci. 2024, 25, 5681. https://doi.org/10.3390/ijms25115681
Kotov AA, Adashev VE, Kombarov IA, Bazylev SS, Shatskikh AS, Olenina LV. Molecular Insights into Female Hybrid Sterility in Interspecific Crosses between Drosophila melanogaster and Drosophila simulans. International Journal of Molecular Sciences. 2024; 25(11):5681. https://doi.org/10.3390/ijms25115681
Chicago/Turabian StyleKotov, Alexei A., Vladimir E. Adashev, Ilia A. Kombarov, Sergei S. Bazylev, Aleksei S. Shatskikh, and Ludmila V. Olenina. 2024. "Molecular Insights into Female Hybrid Sterility in Interspecific Crosses between Drosophila melanogaster and Drosophila simulans" International Journal of Molecular Sciences 25, no. 11: 5681. https://doi.org/10.3390/ijms25115681
APA StyleKotov, A. A., Adashev, V. E., Kombarov, I. A., Bazylev, S. S., Shatskikh, A. S., & Olenina, L. V. (2024). Molecular Insights into Female Hybrid Sterility in Interspecific Crosses between Drosophila melanogaster and Drosophila simulans. International Journal of Molecular Sciences, 25(11), 5681. https://doi.org/10.3390/ijms25115681