
Tumor-Derived Exosomal miR-143-3p Induces Macrophage M2 Polarization to Cause Radiation Resistance in Locally Advanced Esophageal Squamous Cell Carcinoma

 ,
,
Abstract
1. Introduction
2. Results
2.1. Screening Phase
2.2. The Levels of Serum miRNAs Are Associated with Progression-Free Survival of Patients with ESCC Receiving Radiotherapy
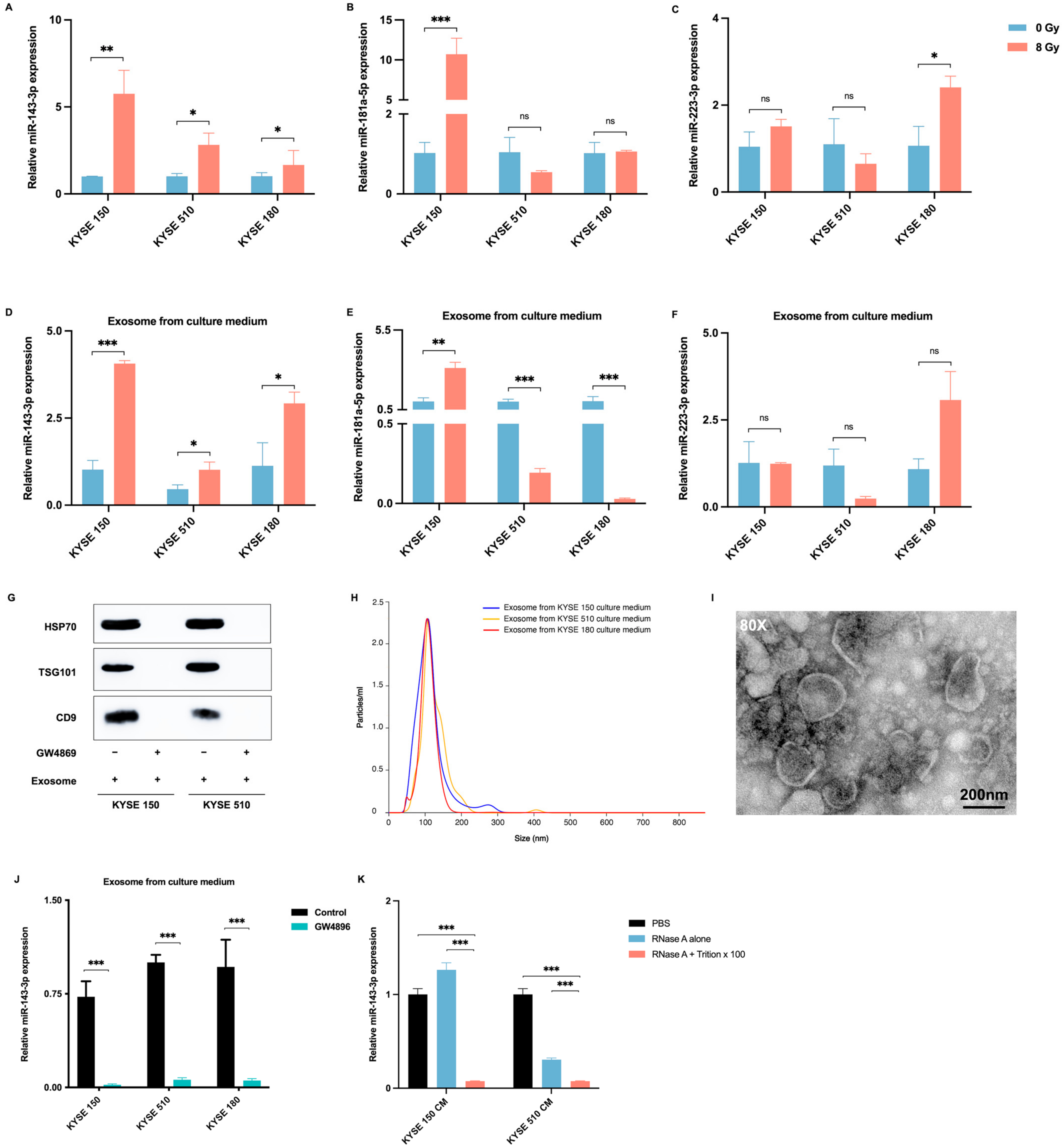
2.3. Radiotherapy Promotes the Delivery of ESCC-Derived Exosomes Carrying miR-143-3p into the Tumor Microenvironment
2.4. MiR-143-3p Is Involved in the ESCC Immune Microenvironment
2.5. ESCC Cell-Derived Exosomal miR-143-3p Induces M2 Polarization of Macrophages
3. Materials and Methods
3.1. Clinical Specimens and Ethical Approval
3.2. Follow-Up
3.3. Cell Culturing, RNA Isolation, and Quantitative Real-Time Reverse-Transcription PCR (qRT-PCR)
3.4. miRNA-Specific qRT-PCR
3.5. RNA Sequencing
3.6. Western Blotting
3.7. Isolation and Analysis of Exosomes
3.8. Retroviral Infection
3.9. Immunofluorescence (IF)
3.10. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, J.; Koulaouzidis, A.; Marlicz, W.; Lok, V.; Chu, C.; Ngai, C.H.; Zhang, L.; Chen, P.; Wang, S.; Yuan, J.; et al. Global Burden, Risk Factors, and Trends of Esophageal Cancer: An Analysis of Cancer Registries from 48 Countries. Cancers 2021, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rice, T.W.; Apperson-Hansen, C.; DiPaola, L.M.; Semple, M.E.; Lerut, T.E.M.R.; Orringer, M.B.; Chen, L.Q.; Hofstetter, W.L.; Smithers, B.M.; Rusch, V.W.; et al. Worldwide Esophageal Cancer Collaboration: Clinical staging data. Dis. Esophagus 2016, 29, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, X.; Wang, L.; Chen, J.; Zhang, W.; Pang, Q.; Zhao, Y.; Sun, X.; Zhang, K.; Li, G.; et al. Clinical practice and outcome of radiotherapy for advanced esophageal squamous cell carcinoma between 2002 and 2018 in China: The multi-center 3JECROG Survey. Acta Oncol. 2021, 60, 627–634. [Google Scholar] [CrossRef]
- NCCN. National Comprehensive Cancer Network: NCCN Clinical Practice Guidelines in Oncology: Esophageal and Esophagogastric Junction Cancers Version 3.2023; NCCN: Plymouth Meeting, PA, USA, 2023. [Google Scholar]
- Herskovic, A.; Martz, K.; Alsarraf, M.; Leichman, L.; Brindle, J.; Vaitkevicius, V.; Cooper, J.; Byhardt, R.; Davis, L.; Emami, B. Combined Chemotherapy and Radiotherapy Compared with Radiotherapy Alone in Patients with Cancer of the Esophagus. N. Engl. J. Med. 1992, 326, 1593–1598. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.S.; Guo, M.D.; Herskovic, A.; Macdonald, J.S.; Martenson, J.A.; Al-Sarraf, M.; Byhardt, R.; Russell, A.H.; Beitler, J.J.; Spencer, S.; et al. Chemoradiotherapy of locally advanced esophageal cancer—Long-term follow-up of a prospective randomized trial (RTOG 85-01). JAMA-J. Am. Med. Assoc. 1999, 281, 1623–1627. [Google Scholar] [CrossRef]
- Kumar, S.; Dimri, K.; Khurana, R.; Rastogi, N.; Das, K.J.M.; Lal, P. A randomised trial of radiotherapy compared with cisplatin chemo-radiotherapy in patients with unresectable squamous cell cancer of the esophagus. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2007, 83, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Toh, Y.; Numasaki, H.; Tachimori, Y.; Uno, T.; Jingu, K.; Nemoto, K.; Matsubara, H. Current status of radiotherapy for patients with thoracic esophageal cancer in Japan, based on the Comprehensive Registry of Esophageal Cancer in Japan from 2009 to 2011 by the Japan Esophageal Society. Esophagus 2020, 17, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Wang, L.; Myles, B.; Thall, P.F.; Hofstetter, W.L.; Swisher, S.G.; Ajani, J.A.; Cox, J.D.; Komaki, R.; Liao, Z. Propensity score-based comparison of long-term outcomes with 3-dimensional conformal radiotherapy vs intensity-modulated radiotherapy for esophageal cancer. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.Y.; Wang, C.; Shi, X.H.; Jiang, G.L.; Wang, Y.; Liu, Y.; Zhao, K.L. Reduced toxicity with three-dimensional conformal radiotherapy or intensity-modulated radiotherapy compared with conventional two-dimensional radiotherapy for esophageal squamous cell carcinoma: A secondary analysis of data from four prospective clinical. Dis. Esophagus 2016, 29, 1121–1127. [Google Scholar] [CrossRef]
- Chen, N.B.; Qiu, B.; Zhang, J.; Qiang, M.Y.; Zhu, Y.J.; Wang, B.; Guo, J.Y.; Cai, L.Z.; Huang, S.M.; Liu, M.Z.; et al. Intensity-Modulated Radiotherapy versus Three-Dimensional Conformal Radiotherapy in Definitive Chemoradiotherapy for Cervical Esophageal Squamous Cell Carcinoma: Comparison of Survival Outcomes and Toxicities. Cancer Res. Treat. 2020, 52, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Li, G.; Li, H.; Jia, F. Comparison of IMRT versus 3D-CRT in the treatment of esophagus cancer. Medicine 2017, 96, e7685. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.; Settle, S.H.; Amini, A.; Xiao, L.; Suzuki, A.; Hayashi, Y.; Hofstetter, W.; Komaki, R.; Liao, Z.; Ajani, J.A. Failure patterns in patients with esophageal cancer treated with definitive chemoradiation. Cancer 2012, 118, 2632–2640. [Google Scholar] [CrossRef] [PubMed]
- Sudo, K.; Xiao, L.; Wadhwa, R.; Shiozaki, H.; Elimova, E.; Taketa, T.; Blum, M.A.; Lee, J.H.; Bhutani, M.S.; Weston, B.; et al. Importance of surveillance and success of salvage strategies after definitive chemoradiation in patients with esophageal cancer. J. Clin. Oncol. 2014, 32, 3400–3405. [Google Scholar] [CrossRef] [PubMed]
- Hulshof, M.C.C.M.; Geijsen, E.D.; Rozema, T.; Oppedijk, V.; Buijsen, J.; Neelis, K.J.; Nuyttens, J.J.M.E.; van der Sangen, M.J.C.; Jeene, P.M.; Reinders, J.G.; et al. Randomized Study on Dose Escalation in Definitive Chemoradiation for Patients with Locally Advanced Esophageal Cancer (ARTDECO Study). J. Clin. Oncol. 2021, 39, 2816–2824. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Isaac, R.; Reis, F.C.G.; Ying, W.; Olefsky, J.M. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021, 33, 1744–1762. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 147. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ning, Z.; Ma, L.; Liu, W.; Shao, C.; Shu, Y.; Shen, H. Exosomal miRNAs and miRNA dysregulation in cancer-associated fibroblasts. Mol. Cancer 2017, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mi, Y.; Guan, B.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Cai, S.; Xu, Y.; Li, X.; et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J. Hematol. Oncol. 2020, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Xie, L.; Lu, C.; Gu, C.; Xia, Y.; Lv, J.; Xuan, Z.; Fang, L.; Yang, J.; Zhang, L.; et al. Gastric cancer-derived exosomal miR-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J. Exp. Clin. Cancer Res. 2022, 41, 296. [Google Scholar] [CrossRef] [PubMed]
- Cassetta, L.; Pollard, J.W. Targeting macrophages: Therapeutic approaches in cancer. Nat. Rev. Drug Discov. 2018, 17, 887–904. [Google Scholar] [CrossRef] [PubMed]
- Doak, G.R.; Schwertfeger, K.L.; Wood, D.K. Distant Relations: Macrophage Functions in the Metastatic Niche. Trends Cancer 2018, 4, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Kong, P.; Liu, Y.; Huang, J.; Xu, E.; Wei, W.; Li, G.; Cheng, X.; Xue, L.; et al. Integrated multi-omics profiling yields a clinically relevant molecular classification for esophageal squamous cell carcinoma. Cancer Cell 2023, 41, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.E.; Pollard, J.W. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Recht, L.; Strober, S. The Promise of Targeting Macrophages in Cancer Therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 3241–3250. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Pioppini, C.; Ozpolat, B.; Calin, G.A. Non-coding RNAs regulation of macrophage polarization in cancer. Mol. Cancer 2021, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Dong, J.; Wang, L.E.; Ma, H.; Liu, J.; Zhao, Y.; Tang, J.; Chen, X.; Dai, J.; Wei, Q.; et al. Serum microRNA profiling and breast cancer risk: The use of miR-484/191 as endogenous controls. Carcinogenesis 2012, 33, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, S.; Zhou, X.; Xue, L.; Quan, L.; Lu, N.; Zhang, G.; Bai, J.; Wang, Y.; Liu, Z.; et al. Overexpression of human pituitary tumor transforming gene (hPTTG), is regulated by beta-catenin/TCF pathway in human esophageal squamous cell carcinoma. Int. J. Cancer 2005, 113, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.K.; Reznik, S.I. Update on Management of Squamous Cell Esophageal Cancer. Curr. Oncol. Rep. 2022, 24, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Forshaw, M.J.; Gossage, J.A.; Chrystal, K.; Cheong, K.; Atkinson, S.; Botha, A.; Harper, P.G.; Mason, R.C. Neoadjuvant chemotherapy for locally advanced carcinoma of the lower oesophagus and oesophago-gastric junction. Eur. J. Surg. Oncol. J. Eur. Soc. Surg. Oncol. Br. Assoc. Surg. Oncol. 2006, 32, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- de Gouw, D.; Klarenbeek, B.R.; Driessen, M.; Bouwense, S.A.W.; van Workum, F.; Futterer, J.J.; Rovers, M.M.; Ten Broek, R.P.G.; Rosman, C. Detecting Pathological Complete Response in Esophageal Cancer after Neoadjuvant Therapy Based on Imaging Techniques: A Diagnostic Systematic Review and Meta-Analysis. J. Thorac. Oncol. 2019, 14, 1156–1171. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhang, W.; Huang, W.; Liao, Y.; Yin, Y.; Shi, M.; Qin, W.; Zhao, L. CT-based radiomics nomogram may predict local recurrence-free survival in esophageal cancer patients receiving definitive chemoradiation or radiotherapy: A multicenter study. Radiother. Oncol. 2022, 174, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Eyck, B.M.; Onstenk, B.D.; Noordman, B.J.; Nieboer, D.; Spaander, M.C.W.; Valkema, R.; Lagarde, S.M.; Wijnhoven, B.P.L.; van Lanschot, J.J.B. Accuracy of Detecting Residual Disease After Neoadjuvant Chemoradiotherapy for Esophageal Cancer: A Systematic Review and Meta-analysis. Ann. Surg. 2020, 271, 245–256. [Google Scholar] [CrossRef] [PubMed]
- van Rossum, P.S.N.; Goense, L.; Meziani, J.; Reitsma, J.B.; Siersema, P.D.; Vleggaar, F.P.; van Vulpen, M.; Meijer, G.J.; Ruurda, J.P.; van Hillegersberg, R. Endoscopic biopsy and EUS for the detection of pathologic complete response after neoadjuvant chemoradiotherapy in esophageal cancer: A systematic review and meta-analysis. Gastrointest. Endosc. 2016, 83, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Goense, L.; van Rossum, P.S.N.; Reitsma, J.B.; Lam, M.G.E.H.; Meijer, G.J.; van Vulpen, M.; Ruurda, J.P.; van Hillegersberg, R. Diagnostic Performance of 18F-FDG PET and PET/CT for the Detection of Recurrent Esophageal Cancer After Treatment with Curative Intent: A Systematic Review and Meta-Analysis. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2015, 56, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Noordman, B.J.; Spaander, M.C.W.; Valkema, R.; Wijnhoven, B.P.L.; van Berge Henegouwen, M.I.; Shapiro, J.; Biermann, K.; van der Gaast, A.; van Hillegersberg, R.; Hulshof, M.C.C.M.; et al. Detection of residual disease after neoadjuvant chemoradiotherapy for oesophageal cancer (preSANO): A prospective multicentre, diagnostic cohort study. Lancet Oncol. 2018, 19, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Nikanjam, M.; Kato, S.; Kurzrock, R. Liquid biopsy: Current technology and clinical applications. J. Hematol. Oncol. 2022, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Shen, S.; Moran, R.; Deng, N.; Marbán, E.; Melmed, S. Pituitary Somatotroph Adenoma-derived Exosomes: Characterization of Nonhormonal Actions. J. Clin. Endocrinol. Metab. 2022, 107, 379–397. [Google Scholar] [CrossRef] [PubMed]
- Hemmatzadeh, M.; Mohammadi, H.; Karimi, M.; Musavishenas, M.H.; Baradaran, B. Differential role of microRNAs in the pathogenesis and treatment of Esophageal cancer. Biomed. Pharmacother. 2016, 82, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Zhao, W.; Wang, D.; Ma, W.; Shang, S.; Feng, C.; Yu, H. MicroRNA expression in esophageal squamous cell carcinoma: Novel diagnostic and prognostic biomarkers. Mol. Med. Rep. 2017, 15, 3833–3839. [Google Scholar] [CrossRef] [PubMed]
- Affo, S.; Yu, L.X.; Schwabe, R.F. The Role of Cancer-Associated Fibroblasts and Fibrosis in Liver Cancer. Annu. Rev. Pathol. 2017, 12, 153–186. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.H.; Hong, J.Y.; Park, H.J.; Lee, S.K. The role of exosomes and miRNAs in drug-resistance of cancer cells. Int. J. Cancer 2017, 141, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Locati, M. Tumor-associated macrophages as a paradigm of macrophage plasticity, diversity, and polarization: Lessons and open questions. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, X.; Si, M.; Yang, J.; Sun, S.; Wu, H.; Cui, S.; Qu, X.; Yu, X. Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages. Cancer Lett. 2020, 474, 36–52. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | % | OS | PFS | |
|---|---|---|---|---|
| p | p | |||
| Age (year) | 0.108 | 0.040 | ||
| ≥65 | 18 | 45.0 | ||
| <65 | 22 | 55.9 | ||
| KPS | 0.019 | 0.014 | ||
| ≥90 | 17 | 42.5 | ||
| <90 | 23 | 57.5 | ||
| Stage (AJCC 6th) | 0.064 | 0.035 | ||
| I–II | 8 | 20.0 | ||
| III–IV | 32 | 80.0 | ||
| Location | 0.392 | 0.128 | ||
| Cervical and upper | 8 | 20.0 | ||
| Middle and lower | 32 | 80.0 | ||
| Length | 0.016 | 0.001 | ||
| ≤5 cm | 18 | 45.0 | ||
| >5 cm | 22 | 55.0 | ||
| Primary tumor volume | 0.019 | 0.007 | ||
| ≤50 cm3 | 18 | 45.0 | ||
| >50 cm3 | 22 | 55.0 | ||
| Therapeutic approach | 0.838 | 0.610 | ||
| RT | 19 | 47.5 | ||
| CRT (Single) | 7 | 17.5 | ||
| CRT (Doublet) | 14 | 35.0 | ||
| Response | 0.566 | 0.758 | ||
| ORR | 20 | 50.0 | ||
| SD | 14 | 35.0 | ||
| PD | 1 | 2.5 | ||
| NA | 5 | 12.5 | ||
| Fold change in miR-143-3p | 0.872 | 0.443 | ||
| ≤1.5 | 27 | 67.5 | ||
| >1.5 | 13 | 32.5 | ||
| Fold change in miR-181a-5p | 0.382 | 0.833 | ||
| ≤1.5 | 19 | 47.5 | ||
| >1.5 | 21 | 52.5 | ||
| Fold change in miR-223-3p | 0.069 | 0.128 | ||
| ≤1.5 | 24 | 60.0 | ||
| >1.5 | 16 | 40.0 | ||
| Fold change in miR-143-3p and miR-223-3p | 0.024 | 0.008 | ||
| Both ≤ 1.5 | 20 | 50.0 | ||
| Any one > 1.5 and both > 1.5 | 20 | 50.0 | ||
| Fold change in miR-143-3p and miR-181a-5p | 0.318 | 0.456 | ||
| Both ≤ 1.5 | 18 | 45.0 | ||
| Any one > 1.5 and both > 1.5 | 22 | 55.0 | ||
| Fold change in miR-181a-5p and miR-223-3p | 0.034 | 0.401 | ||
| Both ≤ 1.5 | 15 | 37.5 | ||
| Any one > 1.5 and both > 1.5 | 25 | 62.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, L.-R.; Zhang, J.; Huang, N.; Deng, W.; Ni, W.; Xiao, Z.; Liu, M. Tumor-Derived Exosomal miR-143-3p Induces Macrophage M2 Polarization to Cause Radiation Resistance in Locally Advanced Esophageal Squamous Cell Carcinoma. Int. J. Mol. Sci. 2024, 25, 6082. https://doi.org/10.3390/ijms25116082
Gao L-R, Zhang J, Huang N, Deng W, Ni W, Xiao Z, Liu M. Tumor-Derived Exosomal miR-143-3p Induces Macrophage M2 Polarization to Cause Radiation Resistance in Locally Advanced Esophageal Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2024; 25(11):6082. https://doi.org/10.3390/ijms25116082
Chicago/Turabian StyleGao, Lin-Rui, Jiajun Zhang, Ning Huang, Wei Deng, Wenjie Ni, Zefen Xiao, and Mei Liu. 2024. "Tumor-Derived Exosomal miR-143-3p Induces Macrophage M2 Polarization to Cause Radiation Resistance in Locally Advanced Esophageal Squamous Cell Carcinoma" International Journal of Molecular Sciences 25, no. 11: 6082. https://doi.org/10.3390/ijms25116082
APA StyleGao, L.-R., Zhang, J., Huang, N., Deng, W., Ni, W., Xiao, Z., & Liu, M. (2024). Tumor-Derived Exosomal miR-143-3p Induces Macrophage M2 Polarization to Cause Radiation Resistance in Locally Advanced Esophageal Squamous Cell Carcinoma. International Journal of Molecular Sciences, 25(11), 6082. https://doi.org/10.3390/ijms25116082