Abstract
Streptococcus gordonii (S. gordonii, Sg) is one of the early colonizing, supragingival commensal bacterium normally associated with oral health in human dental plaque. MicroRNAs (miRNAs) play an important role in the inflammation-mediated pathways and are involved in periodontal disease (PD) pathogenesis. PD is a polymicrobial dysbiotic immune-inflammatory disease initiated by microbes in the gingival sulcus/pockets. The objective of this study is to determine the global miRNA expression kinetics in S. gordonii DL1-infected C57BL/6J mice. All mice were randomly divided into four groups (n = 10 mice/group; 5 males and 5 females). Bacterial infection was performed in mice at 8 weeks and 16 weeks, mice were euthanized, and tissues harvested for analysis. We analyzed differentially expressed (DE) miRNAs in the mandibles of S. gordonii-infected mice. Gingival colonization/infection by S. gordonii and alveolar bone resorption (ABR) was confirmed. All the S. gordonii-infected mice at two specific time points showed bacterial colonization (100%) in the gingival surface, and a significant increase in mandible and maxilla ABR (p < 0.0001). miRNA profiling revealed 191 upregulated miRNAs (miR-375, miR-34b-5p) and 22 downregulated miRNAs (miR-133, miR-1224) in the mandibles of S. gordonii-infected mice at the 8-week mark. Conversely, at 16 weeks post-infection, 10 miRNAs (miR-1902, miR-203) were upregulated and 32 miRNAs (miR-1937c, miR-720) were downregulated. Two miRNAs, miR-210 and miR-423-5p, were commonly upregulated, and miR-2135 and miR-145 were commonly downregulated in both 8- and 16-week-infected mice mandibles. Furthermore, we employed five machine learning (ML) algorithms to assess how the number of miRNA copies correlates with S. gordonii infections in mice. In the ML analyses, miR-22 and miR-30c (8-week), miR-720 and miR-339-5p (16-week), and miR-720, miR-22, and miR-339-5p (combined 8- and 16-week) emerged as the most influential miRNAs.
1. Introduction
Multispecies microbial commensal communities, rather than single pathogens, in oral cavities are critical for the initiation and development of polymicrobial oral diseases (PD and caries). S. gordonii, a member of the oral mitis group of streptococci S. gordonii, is the most abundant first inhabitant of the human oral cavity and dental plaque. Acquired shortly after birth, it plays an important role in the assembly of the oral microbiota [1]. This commensal, non-pathogenic bacterium present in the skin, oral cavity, upper respiratory tract, and intestines has been strongly implicated in infectious complications such as apical PD, bacteremia, infective endocarditis, empyema, perihepatic abscesses, pyogenic spondylitis, or spondylodiscitis [2,3,4,5,6]. S. gordonii is an early initial bacterial colonizer, commensal supragingival Gram-positive bacterium that possesses cell surface proteins that can bind to the cell surface components of host mammalian cells, thereby producing hydrogen peroxide (H2O2) as a by-product of sugar metabolism, rapidly attaching to tooth surfaces and heart valves, and promotes the formation of plaque biofilms [7,8,9]. The cell wall components (lipoteichoic acids, teichoic acids, lipoproteins, serine-rich repeat adhesins, peptidoglycans, and cell wall proteins) are recognized by individual host receptors and are involved in virulence and immunoregulatory processes which cause host inflammatory responses [10]. This bacterium facilitates late bacterial colonizers such as Porphyromonas gingivalis and Fusobacterium nucleatum through a process of coaggregation [11,12]. Periodontitis is a polymicrobial dysbiotic chronic inflammatory disease caused by microbes, including bacteria, viruses, and fungi interacting in the host subgingival sulcus/pockets. A recent meta-analysis encompassing 50 studies comparing the prevalence of microorganisms other than the already-known periodontal pathogens in subgingival plaque and/or saliva samples between subjects with PD and periodontal health that showed 25 bacterial species, including S. gordonii, that were significantly associated with periodontitis [13].
MiRNAs are a class of short, small (21- to 25-nucleotide), non-coding RNAs important for post-transcriptional regulators to the gene expression in metazoans. More than 2000 human microRNAs from the miRBase showed links to predicted and validated targets [14]. These regulatory small miRNAs play critical roles in the regulation of gene expression and have potential as biomarkers [15,16,17,18]. Understanding their cellular functions is fundamentally significant for understanding disease pathogenesis.
Current knowledge is limited regarding how the oral commensal supragingival bacterium S. gordonii affects miRNA expression during PD induced by oral infection. MiRNAs are also strongly considered as diagnostic/genetic novel biomarkers for several diseases such as cardiovascular disease, HIV, diabetes, hypertension, several types of cancers, Alzheimer’s disease (AD), and autoimmune diseases. Moreover, the deregulation of miRNAs has been implicated in a multitude of diseases, and is known to regulate the immune response of hosts infected with bacteria (Treponema pallidum, Helicobacter pylori, Mycobacterium avium, M. tuberculosis, Salmonella, and Listeria monocytogenes). MiRNAs are resistant to degradation by ribonucleases and have been shown to be potential biomarkers for disease prediction and diagnosis. Accordingly, an improved understanding of the biogenesis pattern of miRNAs could potentially lead to the development of novel diagnostic biomarkers for polymicrobial infection-driven inflammatory diseases [19]. S. gordonii increased the expression of miR-4516 and miR-663a in the human oral epithelial cells, modulating validated targets of chemokine-associated pathways [20].
The inflammatory miRNAs miR-21 [21], miR-325-3p [22], miR-27a, miR-34a [23], miR-132 [24,25], miR-146a, miR-155 [26,27], miR-126-3p [28,29] miR-200, and mmu-let-7a [23] have been reported in the initiation and progression of PD. Recently, we characterized alterations in sex-specific microRNA after partial human mouth microbes (PAHMM) in an ecological time-sequential polybacterial periodontal infection (ETSPPI) mouse model and identified seven miRNAs (miR-9, miR-148a, miR-669a, miR-199a-3p, miR-1274a, miR-377, and miR-690) in both sexes that may be implicated in the pathogenesis of PD [19]. The five miRNAs (miR-195, 31, 125b-5p, 15a, and 423-5p) that were DE during the P. gingivalis monobacterial infection [30] and the seven miRs (miR-22, 486, 126-3p, 378, 151a-3p, 423-5p, and 221) during T. denticola monobacterial infection were also DE in chronic PD with diabetes [25,30]. Similarly, we recently identified T. forsythia-specific miRNAs (miR-let-7c, miR-210, miR-146a, miR-423-5p, miR-24, miR-218, miR-26b, and miR-23a-3p) and these miRNAs have also been reported in the gingival tissues and saliva of PD patients [23]. Further, several DE miRNAs that are significantly upregulated (e.g., miR-101b, miR-218, miR-127, and miR-24) are also associated with many systemic diseases such as atherosclerosis, AD, RA, osteoarthritis, diabetes, obesity, and several cancers [23]. These miRNAs are associated in induction of PD and their biogenesis pattern may be bacterial-specific and time dependent [23]. Furthermore, there is a lack of research on miRNA profiling throughout the progression of PD with S. gordonii linked with dental caries and PD. Therefore, to elucidate the changes in miRNA expression associated with S. gordonii-specific bacterial infection and subsequent PD, this study employed S. gordonii to induce experimental PD at two different time points.
A recent literature search revealed an absence of studies employing live S. gordonii to examine miRNA signature patterns under both in vitro and in vivo conditions. Thus, identifying miRNA signature patterns specific to S. gordonii infection is essential to understanding the complex inflammatory pathways initiated by S. gordonii during infection. Furthermore, we employed the ML algorithms XGB, RFC, LR, SVC, and MLP to the S. gordonii infection-induced miRNA data. In a recent study we published, two ML models, XGB and RFC, were employed to analyze periodontal miRNA in response to T. forsythia in the mouse model [23]. This approach provided deeper insights into the specific miRNAs associated with PD infection.
2. Results
2.1. Chronic Infection of S. gordonii Effectively Colonized in Mice Gingival Surface
We previously reported that the oral inoculation of Gram-negative subgingival periodontal bacteria (P. gingivalis, T. denticola, or T. forsythia) promotes bacterial colonization on the gingival surface and induces miRNA expression [19,23,25,30,31]. In the present study, new experiments were conducted designed to examine whether Gram-positive S. gordonii, inhabiting the supragingival region can colonize gingival epithelium and induce PD in mice. Gingival plaque samples of mice showed the presence of 40% S. gordonii DNA after the first infection cycle in both the 8-week and 16-week study groups. All 90% of the mice tested positive for the S. gordonii-specific 16S rRNA gene during the second infection cycle and 100% tested positive after the sixth cycle. This confirmed S. gordonii colonization in both 8 and 16 weeks (Table 1). We estimated S. gordonii-specific IgG antibody kinetics at each time point.

Table 1.
Gingival plaque samples tested positive for S. gordonii gDNA using PCR.
2.2. Higher Alveolar Bone Resorption (ABR) and Bacterial Dissemination to Distal Organs
Osteoclast activation by inflammatory cytokine sensitization and pathogenic factors of periodontal bacteria contributes to ABR in periodontal infection [32]. Microscopic images of mandibles from both sham-infected and S. gordonii-infected are shown in Figure 1B. Mice infected with S. gordonii at both 8 weeks and 16 weeks showed significantly higher ABR in the mandible (lingual) p < 0.05 (Adjusted p-value= 0.0004) for the 16-week group (Figure 1B,C). Similarly, a significantly higher (p < 0.0001) ABR was observed in maxilla palatal for 16-week infected mice. Aggressive gingival infections will permeate bacteria into the bloodstream and can settle on the heart lining and valves [33]. None of the aliquoted internal organs (heart, lungs, brain, liver, kidney, or spleen) tested positive for S. gordonii genomic DNA at both 8 and 16 weeks of infection (Supplementary Table S1).
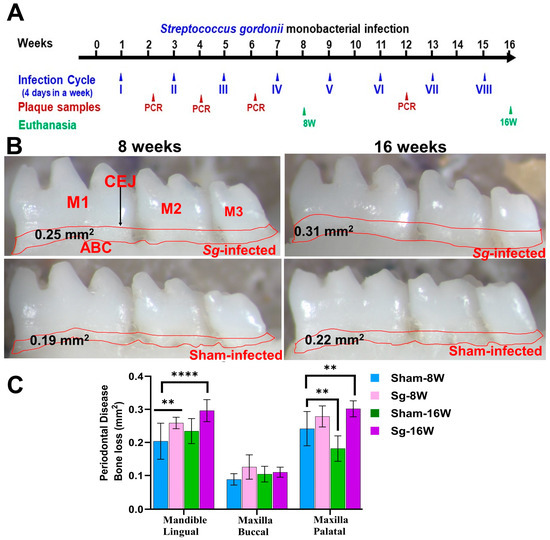
Figure 1.
Intraoral infection of S. gordonii significantly induced ABR. (A) Schematic diagram of the experimental design depicting the monobacterial S. gordonii infection (4 days per week on every alternate week), plaque sampling for PCR, and euthanasia. (B) Representative images showing horizontal ABR (mandible lingual view) of S. gordonii-infected and sham-infected mice with the area of ABR outlined from the alveolar bone crest (ABC) to the cementoenamel junction (CEJ). (C) Morphometric analysis of the mandible and maxillary ABR in mice. A significant increase in ABR was observed in S. gordonii-infected mice compared to sham-infected mice at both 8-week and 16-week infected mice (** p < 0.05; ****, p < 0.0001; adjusted p-value = 0.0004; ordinary two-way ANOVA). Data points and error bars are mean ± SEM (n = 10).
2.3. NanoString Analysis of miRNAs in S. gordonii-Infected Mandibles
The NanoString platform is highly reliable for working with multiple sample types. It is an amplification-free technology that uses molecular barcodes to directly quantify RNA molecules without the need for reverse transcription. The RNA extracted from mandibles was analyzed for global miRNA profiling in mice infected with S. gordonii for 8 and 16 weeks (Table 2). nCounter miRNA expression profiling showed 191 upregulated miRNAs including miR-375, miR-34b-5p, miR-142-5p, miR-135a (Table 3 and Supplementary Table S2), and 22 downregulated miRNAs including miR-133a, miR-1224, miR-2135, miR-499 (Supplementary Table S3) in 8 weeks of S. gordonii-infected mandibles compared to sham-infected mandibles. During 16 weeks of infection, 10 miRNAs (miR-1902, miR-203, miR-98, miR-210) were found to be upregulated (Table 4), and 32 miRNAs (miR-720, miR-1937c, miR-2135, miR-326) were downregulated (Supplementary Table S4). A p-value of <0.05 and a fold change of 1.1 and above was considered for analysis and significance (Table 3). Both upregulated and downregulated DE miRNAs between 8 and 16 weeks are shown in Supplementary Tables S5 and S6.
Table 2.
Differentially expressed (DE) miRNAs during 8- and 16-weeks of S. gordonii infection.
2.4. Identification of Differentially Expressed (DE) miRNAs
A total of 40 samples from the four infection and sham infection groups were analyzed using high-throughput nCounter® miRNA Expression Panels. To determine the DE miRNAs with statistical significance, we performed a volcano plot analysis, where we plotted log2 fold change on the x-axis and the negative log of p-value on the y-axis. We identified 19 downregulated (red) miRNAs and 19 upregulated (green) miRNAs that displayed a fold difference of +1.1 with a p-value of <0.05 in 8-week S. gordonii-infected mice, compared to the 16-week infected group (Figure 2A). The black dots represent miRNAs that do not pass the filter parameters. A total of 191 miRNAs showed higher expression during 8 weeks of infection (e.g., miR-375, miR-34b-5p, miR-142-5p, and miR-135a) and 10 miRNAs (e.g., miR-1902, miR-203, miR-210, miR-876-3p) were showed higher expression during 16 weeks of S. gordonii-infected mandibles (Figure 2B, Table 3 and Table 4). The miRNAs upregulated for 8 weeks are associated with the differentiation of human periodontal ligament stem cells (miR-375), acute inflammation, injury in the lung (miR-34b-5p), inflammation in the brain (miR-142-5p), lowered (miR-135a) in osteoblastic differentiation and higher (miR-323-3p) in obese PD patients. During a 16-week infection of S. gordonii, certain miRNAs were found to increase in response to lipoteichoic acid stimulation (miR-1902), in periodontal diabetic patients with gingivitis (miR-203), in obese PD patients (miR-210), and in PD patients in general (miR-423-5p). The expression of miR-323-3p is consistent across the S. gordonii DE, XGB, and RFA analyses along with the expression of DE-miRNAs miR-142-5p and miR-1968, as identified by XGB and RFA, respectively.
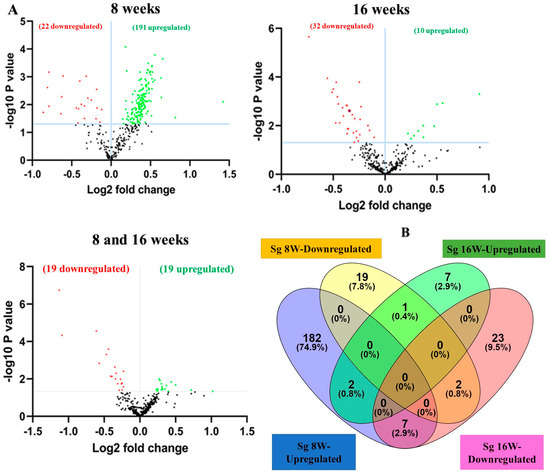
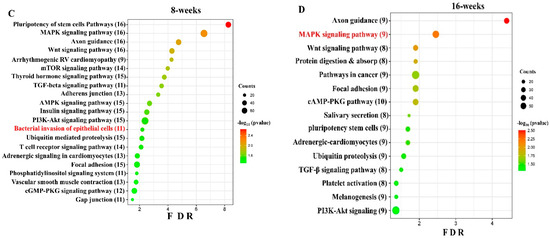
Figure 2.
DE miRNAs in S. gordonii-infected mandibles (8- and 16-weeks). (A) The volcano plot depicts the upregulated (green) and downregulated (red) miRNAs that showed a fold difference of ±1.1 with a p-value of <0.05. The log2 fold change is on the x-axis, and the negative log of the p-value is on the y-axis. The black dots represent the miRNAs that do not pass the filter parameters. A total of 191 significant upregulated miRNAs and 22 downregulated miRNAs were identified in 8 weeks of S. gordonii-infected mice compared to 8 weeks of sham-infected mice (n = 10). Ten significant upregulated miRNAs and 32 downregulated miRs were identified in 16 weeks of S. gordonii-infected mice compared to 16-week sham-infected mice (n = 10). (B) Venn diagram analysis illustrates the distribution of DE miRs in 8-week and 16-week infections with S. gordonii. (C,D) Predicted functional pathway analysis of DE miRNAs from S. gordonii-infected mandibles. Bubble Plot of KEGG analysis on predicted target genes of DE miRNAs in S. gordonii-infected mice at 8- and 16 weeks of infection compared to sham-infected mice. The KEGG pathways are displayed on the y-axis, and the x-axis represents the false discovery rate (FDR), which means the probability of false positives in all tests. The size and color of the dots represent the number of predicted genes and corresponding p-value, respectively. Eleven DE miRNAs were shown to be involved in bacterial invasion of epithelial cells during 8 weeks of infection. Red color highlighted pathways explained in detailed in this manuscript.
Table 3.
S. gordonii infection-induced upregulated miRNAs, reported functions, and target genes.
Table 3.
S. gordonii infection-induced upregulated miRNAs, reported functions, and target genes.
| Upregulated miRNAs in 8-Week S. gordonii Infection | ||||
|---|---|---|---|---|
| miRNA | Fold Change | p-Value | Reported Function | Number of Target Genes |
| miR-375 | 2.68 | 0.0079 | Facilitated the proliferation and osteogenic differentiation capacity in human periodontal ligament stem cells [33]. Potential biomarker in esophageal cancer [15]. Upregulated in the saliva of migraine without aura patients [34]. | 27 (e.g., Akap2, Atrn, Lpin1, Rpsa, Pacsin1) |
| miR-34b-5p | 1.76 | 0.0292 | Upregulated in acute lung injury [35]. Regulates the lung inflammation, and apoptosis in LPS-induced ALI mouse model [36]. Upregulated in the saliva of migraine without aura patients [34]. | 354 (e.g., 2310010M20Rik, Rg9mtd2, Dnase2a, Epb4.2, Fyco1) |
| miR-142-5p | 1.58 | 0.0002 | High levels were reported in the condition of experimental autoimmune encephalomyelitis mice [37]. Promoting brain inflammation through astrocyte activation in traumatic brain injury [38]. | 24 (e.g., Abcg1, Cd28, Sort1, Slc35a1, Cwc25) |
| miR-135a | 1.56 | 0.0056 | Inhibiting the differentiation of osteoblasts [39], and regulating osteoblastic activity [40]. Serving the role in osteoporosis progression by regulating osteogenic differentiation via RUNX2 [41]. | 19 (e.g., Nup133, Adamts14, Cep170, App, Smad5) |
| miR-323-3p | 1.55 | 0.0011 | Upregulated in obese individuals with periodontitis [42]. | 25 (e.g., Fam190a, Eed, Fzd3, Pira1, Tyw1, Ccser1) |
| miR-485 | 1.45 | 0.0015 | up-regulated in osteoporosis patients [43]. Dysregulated miR-485 is associated with AD and Parkinson’s disease [44]. | 14 (Cd93, Pigm, Fasl, Gns) |
| miR-881 | 1.44 | 0.0007 | Upregulated in dental pulp cells induced by 5-AZA-CdR [45]. Upregulated in the NAFLD liver tissue [46]. Downregulated in the chronic alcoholic rats [47]. Upregulated in the acute lung injury mice [48]. | 50 (e.g., Xiap, Stxbp5, Slc31a2, Wbp1l) |
| miR-669e | 1.43 | 0.0004 | Downregulated in the flavonoid-treated mice [49]. | 108 (e.g., Wdr59, Mmp25, Ubac1, Mzt1, Fzd2) |
| miR-187 | 1.43 | 0.0004 | Upregulated in the sepsis-induced myocardial dysfunction- mice supplemented with mesenchymal stem cells [50]. Reported as a diagnostic and predictive factor for oral squamous cell carcinoma patients [51]. Restoring the osteogenic differentiation capacity in bone marrow-derived mesenchymal stem cells [52]. The most abundant miR in the serum of women in different stages of pregnancy or patients with different urothelial cancers [53]. | 14 (e.g., Cd93, Dmpk, Rgs4, Rnaseh2a, Adcy1). |
| miR-1306 | 1.43 | 0.0027 | Mediating the feedback regulation of the TGF-β signaling pathway in granulosa cells [54]. Regulating the human ameloblastoma AM-1 cells differentiation [55]. | 22 (e.g., Rinl, Frmd3, Tpmt, Ptprb, Map3k7). |
| miR-200b | 1.43 | 0.0253 | Variations in the levels of miR-200b were observed in gingival tissue of obese periodontitis subjects [42]. | 55 (e.g., Zeb2, Suz12, Bmi1, Flt1, Mapk14) |
| miR-767 | 1.42 | 0.0076 | Upregulated in hepatocellular carcinoma [56]. Forced upregulation of miR-767-5p may represent a novel therapeutic strategy for glioma patients by targeting SUZ12 [57]. Highly expressed in senescent vascular endothelial cells [58]. | 13 (e.g., Mcl1, Gtpbp1, Klf6, Pigs, Tmem178b) |
| miR-455 | 1.41 | 0.0006 | Associated with cartilage development, adipogenesis, preeclampsia, and cancers in humans [59]. | 31 (e.g., Acvr2b, Smad2, Chrdl1, Fut1, Cd36) |
| miR-2140 | 1.4 | 0.0007 | Predictive markers for the onset of cognitive impairment [60]. Upregulated in the differentiated P19 embryonic carcinoma cells [61]. | ------------ |
| miR-1187 | 1.4 | 0.0027 | Upregulated in hypoxia-induced apoptotic HL-1 cardiomyocytes [62]. Functions as a negative regulator of osteogenesis [63]. Upregulated in high glucose-treated kidney podocytes, and kidney tissue in db/db mice [64]. Regulates hepatocyte apoptosis by targeting caspase-8 [65]. | 148 (e.g., Ado, Lifr, Igf2, Zfp157, Slc39a14) |
| miR-291a-3p | 1.4 | 0.0039 | Improve cell viability and osteogenic differentiation of BMSCs [66]. Decreased expression was reported in Cigarette smoke components exposed rat models [67] and in the lungs of LPS-induced ALI mouse models [68] and in spinal cord ischemia-reperfusion injury [69]. | 73 (e.g., Cdkn1a, Dkk1, Pax6, Myl6b, Zfp568) |
| miR-1968 | 1.4 | 0.0071 | Downregulated in lupus nephritis mice [70]. Associated with hepatic energy metabolism [71]. | 19 (e.g., Snx27, Creb5, Ptpre, Itgav, Cdc42bpa) |
| miR-1957 | 1.39 | 0.0018 | Expressed in the lophotrochozoan taxon [72] | 12 (Sorcs1, Il18r1, Tbc1d24, Pla2r1, Pin1) |
| miR-129-3p | 1.39 | 0.0025 | Downregulated in periodontitis patients [73]. Reported as protective miR from cardiomyocyte hypertrophy [74]. Tumor suppressor miR in the ovarian cancer cells [75]. | 0 |
| miR-1942 | 1.39 | 0.0029 | Upregulated in tumors that evaded the prevention of immunotherapy [76]. | 14 (e.g., Zfp760, Rapgef3, Il17ra, Alkbh1, Slc8a1) |
| miR-2861 | 1.38 | 0.0006 | Promoting osteoblast differentiation [77]. Downregulated in HPV-E6 overexpressed HEK293T cells and HaCaT cells [78]. Upregulated in papillary thyroid carcinoma with lymph node metastasis [79]. | 11 (e.g., Hdac5, Runx2, Hlcs, Plxnc1, Cyb5r1) |
| miR-671-3p | 1.38 | 0.0006 | Involved in the pathogenesis of atherosclerosis, rheumatoid arthritis, and acute myocardial infarction [80]. Reported as a positive regulator of glioma progression and development [81]. | 1 (Pbx1) |
| miR-290-5p | 1.38 | 0.0078 | Reported to suppress the breast cancer progression [82]. | 160 (e.g., Adra2b, Birc5, Rplp0, Bach2, Chic1) |
| miR-509-5p | 1.38 | 0.017 | Has a role in alleviating myocardial infarction damage [83]. Serving as a potential biomarker for evaluating osteosarcoma prognosis [84]. | 43 (e.g., BC016495, Abcb7, Slc7a2, Cflar, Cxcr2) |
| miR-205 | 1.38 | 0.0179 | Downregulated in chronic periodontitis patients [85]. Downregulated in LPS-induced periodontal ligament stem cells [86]. Plays an important role in the physiology of epithelia via regulation of pathways governing differentiation and morphogenesis [87]. Reduces actomyosin contractility and activates hair regeneration in young and old mice [88]. | 44 (e.g., Pten, Lrrk2, Dok4, Atp1a1, Gsk3b) |
| miR-466d-3p | 1.37 | 0.0022 | Downregulated in infarction-exposed fetal and adolescent sheep heart tissue [89]. | 202 (e.g., Rad51l3, Gcfc1, Fam54a, Tmem20, Chic1) |
| miR-34a | 1.37 | 0.0032 | Potential therapeutic anticancer miRNA [90]. Downregulated in adolescent sheep heart infarct samples [89]. | 40 (e.g., Dnase2a, Mapre1, Cd93, Mbd4, Fyco1) |
| miR-2139 | 1.37 | 0.0039 | Upregulated in UV-B-induced tumors [91]. | 1 (Cd4) |
| miR-34b-3p | 1.37 | 0.0075 | Significantly expressed in the mice’s spinal cord development [90]. | 11(e.g., Raph1, Pbx1, Cops2, Sec61b, Ttc19) |
Details of the biological function and target genes were given for the 29 significantly DE miRNAs in 8-week infected mice mandibles out of 191 upregulated miRNAs, and other miRNAs shown in the Supplementary Table S2. miRNAs associated with periodontal disease are marked in red.
Table 4.
S. gordonii infection-induced upregulated miRNAs, reported functions, and target genes.
Table 4.
S. gordonii infection-induced upregulated miRNAs, reported functions, and target genes.
| Upregulated miRNAs in 16-Week S. gordonii Infection | ||||
|---|---|---|---|---|
| miRs | Fold Change | p-Value | Reported Function | Number of Target Genes |
| miR-1902 | 1.88 | 0.0005 | Overexpressed in mice serum and whole blood after intraperitoneal injection of lipoteichoic acid (LTA) [92] | 5 (e.g., Evi2b, Arhgef15, Itga11, Myo9a, Tsn) |
| miR-203 | 1.47 | 0.0012 | Upregulated in the human periodontal disease [27]. Elevated in diabetic periodontal patients [93]. Elevated in primary gingival epithelial cell infection with P. gingivalis 33277 [94]. Regulating the periodontal ligament cells—stimulated by LPS of P. gingivalis bacteria [95]. Master modulator and fine-tuning neuro-inflammation [96]. Decreased expression is reported in periodontitis and is consistent with increased angiogenesis in periodontitis [97]. | 70 (e.g., Stxbp4, Trp63, Zfp281, Snora62, Cav1, Vcan) |
| miR-98 | 1.42 | 0.0013 | Overexpression protects the cardiomyocytes against apoptosis [98]. Critical miRNAs are regarded as a potential biomarker candidate for Crohn’s disease [16] | 39 (e.g., Acvr1b, Mmp11, Il6, Phka1, Poteg) |
| miR-210 | 1.39 | 0.0105 | Elevated in obese periodontitis patients [99]. Human periodontal ligament stem cells had elevated miR-210 in the presence of Endobon-xenograft granules [100]. | 47 (e.g., Tcf7l2, Acvr1b, Ucp2, NFKB1, Bcl2) |
| miR-876-3p | 1.29 | 0.01 | miR-876-3p suppresses the progression of colon cancer and correlates with the prognosis of patients [101]. | 19 (e.g., Sorcs1, Ccsap, Plekha2, Il18r1, Fam161b) |
| mmu-let-7c | 1.29 | 0.0305 | Interfere with critical inflammatory cytokine production viz., IL-1β, IL-6, and TNF-α in human osteoarthritis (OA) and RA [102]. Playing a role in cardiomyogenesis promotion activity [103]. | 37 (e.g., Sall4, Myc, Lin28a, EZH2, Gnl3l) |
| miR-423-5p | 1.25 | 0.0165 | Upregulated expression in severe periodontal disease [104]. Higher expression in obese periodontitis subjects [105]. Identified as a new candidate biomarker in the cross-talk between diabetes mellitus and AD [106]. | 2 (e.g., Map1b, Zmat3) |
| miR-361 | 1.21 | 0.0257 | miR-361-3p/Nfat5 signaling axis controls cementoblast differentiation [107]. | 31 (e.g., Eea1, Dnmt3a, Ctbp2, Vps26a, Zfp120) |
| mmu-let-7a | 1.19 | 0.0343 | A proinflammatory role for let-7 miRNAs in experimental asthma [108]. | |
| miR-101b | 1.16 | 0.021 | Increased expression promotes apoptosis of endothelial cells in acute coronary syndrome [109]. Major mediator of tauopathy and dendritic abnormalities in AD progression [110]. | 44 (e.g., Stc1, Atxn1, Map7d1, Rcor3, Msi2) |
Details of the biological function and target genes were given for the top ten significantly expressed miRNAs in 16-week infected mice mandibles. miRNAs associated with periodontal disease are marked in red.
2.5. DE miRNAs and Functional Pathway Analysis
We conducted functional enrichment analysis to predict the biological function of DE, both in the up-regulated and down-regulated miRNAs. The functional pathways associated with the altered upregulated miRNAs at 8 and 16 weeks of infection were assessed using DIANA-miRPath software (https://diana.e-ce.uth.gr/home). The KEGG pathway analysis revealed that the upregulated genes were significantly enriched in mitogen-activated protein kinase (MAPK) signaling, pathways in cancer, WNT (Wingless and Int-1) signaling pathway, Focal adhesion, TGF-beta signaling pathway, and Ubiquitin mediated proteolysis are commonly identified in DE miRNAs of 8 and 16 weeks (Figure 2C). The other important pathways linked with 8 weeks of infection were bacterial invasion of epithelial cells, B-cell receptor signaling pathway, and T-cell receptor signaling pathways. According to KEGG pathway enrichment analysis, the DE miRNA-target genes were significantly enriched in pathways of bacterial invasion of epithelial cells (BIEC) pathway (8 weeks) and MAPK signaling pathway (16 weeks) S. gordonii-infected mandibles compared with sham-infected controls at p < 0.05. S. gordonii significantly altered 24 gene expressions based on an upregulated miRNA profile in the 8-week infection period (Figure 3). The binding ability of miRNA and its target genes were analyzed using MiRTarBase for both 8- and 16-week infection. It has accumulated more than 300 and 60,000 miRNA-target interactions (MTIs) that were validated by the reported assay, Western blot, microarray, and next-generation sequencing experiments. Each miRNA has a different target gene with a specified MiRTarBase ID (Supplementary Tables S7 and S8).
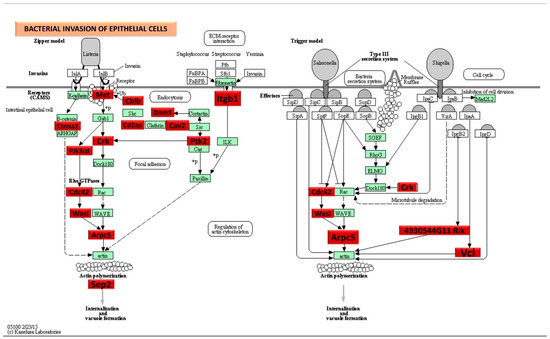
Figure 3.
Significantly, DE genes (identified by KEGG) were involved in the bacterial invasion of epithelial cells signaling pathway during 8-week S. gordonii infection. Red boxes indicate 25 significantly increased expression based on the miRNA profiles from NanoString analysis. Green boxes indicate no change in gene expression. Many pathogenic bacteria can invade phagocytic and non-phagocytic cells and colonize them intracellularly, then become disseminated to other cells. Invasive bacteria induce their uptake by non-phagocytic host cells (e.g., epithelial cells) using two mechanisms referred to as the zipper model and trigger model. Listeria, Staphylococcus, Streptococcus, and Yersinia are examples of bacteria that can enter using the zipper model. These bacteria express proteins on their surfaces that interact with cellular receptors, initiating signaling cascades that result in close apposition of the cellular membrane around the entering bacteria. Shigella and Salmonella are examples of bacteria entering cells using the trigger model. An arrow indicates a molecular interaction and a line without an arrowhead indicates a molecular interaction resulting in inhibition.
2.6. Machine Learning Analysis of miRNA Copies
Our ML analysis results are divided into two groups: tree-based methods consisting of XGB and RFC, and non-tree-based methods consisting of LR, SVC, and MLP. With the 8-week dataset, mmu-miR-30c had the highest impact on the tree-based models (Figure 4A, XGB; Figure 5A, RFC), and mmu-miR-22 had the highest impact on the non-tree-based models (Figure 6A, LR; Figure 7A, SVC; Figure 8A, MLP). For the 16-week dataset, mmu-miR-339-5p had the highest impact on the tree-based models (XGB, RFC; Figure 4B and Figure 5B), while mmu-miR-720 had the most impact on the non-tree-based models (LR, SVC, MLP; Figure 6B, Figure 7B and Figure 8B). Finally, for the combined 8- and 16-week dataset, mmu-miR-339-5p was again the most impactful for the tree-based models (XGB, RFC; Figure 4C and Figure 5C), and mmu-miR-22 and mmu-miR-720 had the highest impacts for the non-tree-based models (LR, SVC, MLP; Figure 6C, Figure 7C and Figure 8C). The results of the SHAP value analysis are summarized in Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8, and a list containing the importance of each miRNA as predicted by the RFC and XGB model is summarized in Table 5. Descriptions of the miRNA functions for the most important agreed-upon features are provided in Table 5. Table 5 provides descriptions for the tree-based models, and Supplementary Table S9 provides descriptions for the non-tree-based models (i.e., LR, SVC, and MLP). The List of miRNAs unique in expression among the DE and 5 ML models depicted in Table S10.
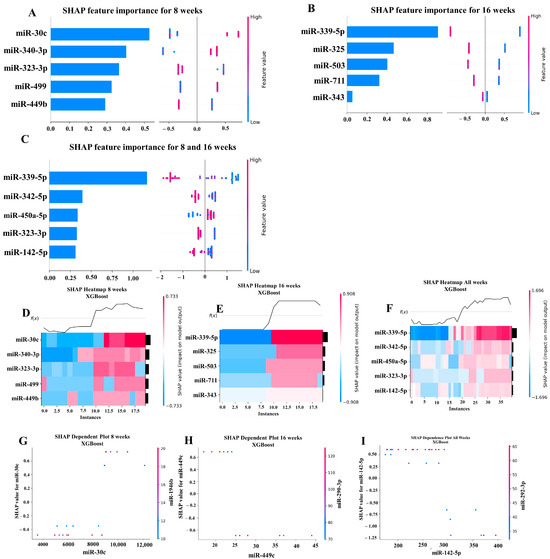
Figure 4.
A summary of the most important features in the XGBoost model using SHAP values. In figures (A–F), feature importance is ranked from the top (the most important) to the bottom (the least important). The x-axis in (A–C) shows the impact that a feature has on the model. The bar charts show the overall impact of a feature whereas the swarm plot shows both the positive and negative impacts. In the swarm plots, each dot represents an instance of a miRNA variable and the color bar shows the value (high to low) of the variable. (A) Mice tested at 8 weeks. (B) Mice tested at 16 weeks are shown, and (C) 8- and 16-week cohorts together. (D–F) The x-axis represents each mouse (instance) in the cohort, and the y-axis is the feature ranking. The color of the cell shows the amount of impact (i.e., the SHAP value) that particular feature inflicted on that feature. The topmost section (i.e., f(x)) of the heatmap shows the predicted infection status for each instance. In all three cohorts, there is a strong correlation between high values for the topmost feature and the model predicting that the mouse was infected. In (G–I), the relation between the topmost feature and the feature it most depends on is shown. Each dot represents a mouse and the x-axis shows the value of the miRNA variable. The left y-axis shows the impact (i.e., the SHAP value) the x-axis variable has on the mode. The right y-axis shows the value of the variable that the x-axis variable interacts with.
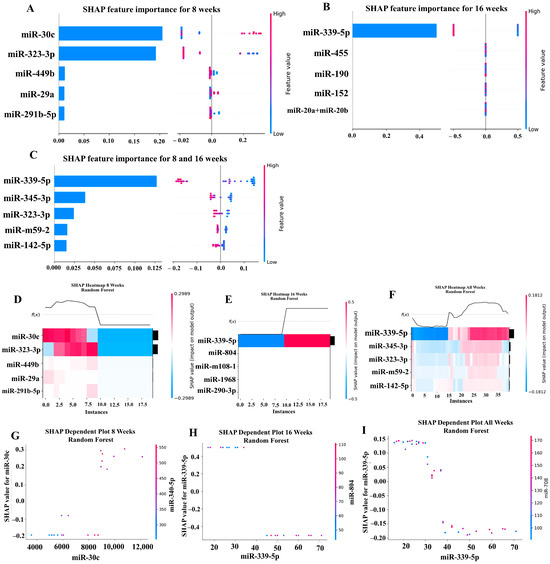
Figure 5.
A summary of the most important features in the random forest classifier model using SHAP values.
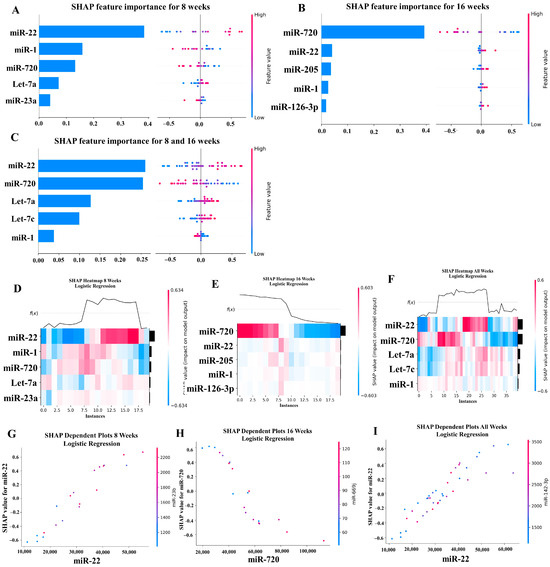
Figure 6.
A summary of the most important features in the logistic regression model using SHAP values.
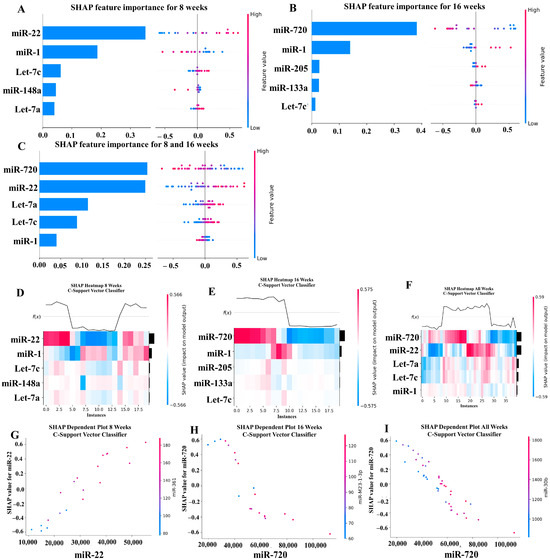
Figure 7.
A summary of the most important features in the C-Support Vector Classifier model using SHAP values.
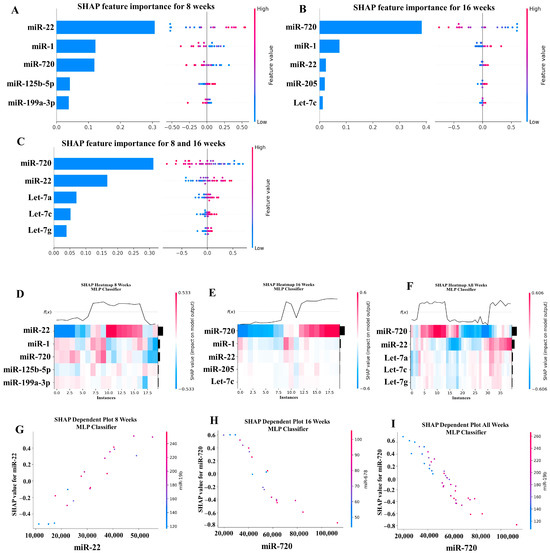
Figure 8.
A summary of the most important features in the multilayer perceptron (Neural Network) model using SHAP values.
In addition to determining which miRNAs were identified by each model, we analyzed which miRNAs had two or more models that agreed for both the miRNA and level of importance (Table 6). The highest agreed-upon miRNAs are consistent with the results mentioned earlier (i.e., in the previous paragraph), but we also found high levels of agreement in the miRNAs ranked 2–5. For the tree-based models, only miR-142-5p was determined as the fifth most important feature in the combined 8- and 16-week datasets (Table 5). For the non-tree-based models, we found more agreement. For the second-ranked feature, LR, SVC, and MLP agreed on miR-1 for the 8-week dataset; SVC and MLP agreed on miR-1 for the 16-week dataset; and LR, SVC and MLP agreed on miR-22 for 8- and 16-week dataset (Table 6). For the third-ranked feature, LR and MLP agreed on miR-720 for the 8-week dataset; LR and SVC agreed on miR-205 for the 16-week dataset; and LR, SVC, and MLP agreed on let-7a for the 8- and 16-week datasets. For the fourth-ranked feature, the only agreement was let-7c in the 8- and 16-week dataset for LR, SVC, and MLP. For the fifth-ranked feature, SVC and MLP agreed on let-7c for the 16-week dataset, and LR and SVC agreed on miR-339-5p for the 8- and 16-week datasets (Table 6). These results are summarized by the cells with the gray background in Table 7.
Lastly, we analyzed which miRNAs we agreed upon by at least two models regardless of rank. These results largely overlap with analysis agreement for both the miRNA and level of agreement, but also include the following miRNAs. For the 8-week dataset, LR (rank 4) and SVC (rank 5) agreed upon let-7a, and RFC (rank 2) and XGB (rank 3) agreed on miR-323-3p (Table 7). For the 8- and 16-week datasets, RFC (rank 3) and XGB (rank 4) agreed on miR-323-3p. We also found agreements of a single model that disagreed with the others for the same miRNA. For the 16-week dataset, LR (rank 4) also agreed with SVC and MLP for miR-1, and MLP (rank 4) also agreed with LR and SVC for miR-205. For the 8- and 16-week dataset, LR (rank 1) agreed with SVC and MLP for miR-22, and LR (rank 2) agreed with SVC and MLP for miR-720. These results are summarized by the cells without a gray background in Table 7.
Table 5.
Random Forest classifier and XGBoost most importantly agreed upon miRNA features and reported miRNA functions.
Table 5.
Random Forest classifier and XGBoost most importantly agreed upon miRNA features and reported miRNA functions.
| miRNA | MIMAT # | Target Functions |
|---|---|---|
| 8-Week Analysis | ||
| miR-30c | MIMAT0000514 | Suppressing the osteogenic differentiation in the human periodontal ligamental stem cells [111]. miR-30 family had a role in the occurrence and development of bone and joint disease [112]. Serving as a non-invasive biomarker for early oral squamous cell carcinoma [17]. Upregulated in multiple system atrophy compared with Parkinson’s disease and healthy subjects [18]. Decreased expression was observed in fasting and post-prandial period of high post-prandial response [113]. |
| miR-323-3p | MIMAT0000551 | Highly expressed in blood samples from patients with coronary heart diseases and rat models of CHD [114]. |
| 16-Week Analysis | ||
| miRNA | MIMAT # | Target Functions |
| miR-339-5p | MIMAT0000584 | Most predictive periodontal miRNA in the T. forsythia-induced mice periodontitis [23]. Downregulated in human atherosclerotic plaques and ox-LDL-induced cells [115]. Downregulated in cardiac tissue of 7-day-old mice [116]. |
| Combined 8 and 16 Weeks | ||
| miRNA | MIMAT # | Target Functions |
| miR-339-5p | MIMAT0000584 | Shown in the 16-week analysis |
| miR-323-3p | MIMAT0000551 | Shown in the 8-week analysis. |
| miR-142-5p | MIMAT0000154 | Upregulated in human gingival crevicular fluids [117]. Regulates inflammation in multiple sclerosis mice models [37]. Decreased expression was reported in breast cancer [118]. Increased expression was reported in macrophages from the tissue samples of patients with liver cirrhosis and idiopathic pulmonary fibrosis [119]. |
miRNAs associated with periodontal disease are marked in red.
Table 6.
Summary of the agreed-upon importance (i.e., rank) and miRNA for each machine learning model.
Table 6.
Summary of the agreed-upon importance (i.e., rank) and miRNA for each machine learning model.
| miRNA Feature Rank | ||||||
|---|---|---|---|---|---|---|
| Dataset | miRNA | 1 | 2 | 3 | 4 | 5 |
| 8 weeks | miR-22 | LR, SVC, MLP | ||||
| miR-1 | LR, SVC, MLP | |||||
| miR-720 | LR, MLP | |||||
| let-7a | LR | SVC | ||||
| miR-23a | LR | |||||
| miR148a | SVC | |||||
| let-7c | SVC | |||||
| miR-125b-5p | MLP | |||||
| miR-199a-3p | MLP | |||||
| mR-30c | RFC, XGB | |||||
| miR-340-3p | XGB | |||||
| miR-323-3p | RFC | XGB | ||||
| miR-499 | XGB | |||||
| miR-449b | RFC | XGB | ||||
| miR-29a | RFC | |||||
| miR-291b-5p | RFC | |||||
| 16 weeks | miR-22 | LR | MLP | |||
| miR-720 | LR, SVC, MLP | |||||
| let-7c | SVC, MLP | |||||
| miR-1 | SVC, MLP | LR | ||||
| miR-205 | LR, SVC | MLP | ||||
| miR-126-3p | LR | |||||
| miR-133a | SVC | |||||
| miR-339-5p | RFC, XGB | |||||
| miR-325 | XGB | |||||
| miR-503 | XGB | |||||
| miR-711 | XGB | |||||
| miR-343 | XGB | |||||
| 8 and 16 weeks | miR-22 | LR | SVC, MLP | |||
| miR-720 | SVC, MLP | LR | ||||
| let-7a | LR, SVC, MLP | |||||
| let-7c | LR, SVC, MLP | |||||
| miR-1 | LR, SVC | |||||
| let-7g | MLP | |||||
| miR-339-5p | RFC, XGB | |||||
| miR-345-3p | RFC | |||||
| miR-323-3p | RFC | XGB | ||||
| miR-m59-2 | RFC | |||||
| miR-142-5p | RFC, XGB | |||||
| miR-342-5p | XGB | |||||
| miR-450a-5p | XGB | |||||
LR—Logistic regression, SVC—C-support vector classifier, MLP—multilayer perceptron, RFC—random forest classifier, and XGB—extreme gradient boosting.
Table 7.
Summary of the agreements for the miRNA and importance (i.e., rank) between each machine learning model.
Table 7.
Summary of the agreements for the miRNA and importance (i.e., rank) between each machine learning model.
| miRNA Feature Rank | ||||||
|---|---|---|---|---|---|---|
| Cohort | miRNA | 1 | 2 | 3 | 4 | 5 |
| 8 weeks | miR-22 | LR, SVC, MLP | ||||
| miR-1 | LR, SVC, MLP | |||||
| miR-720 | LR, MLP | |||||
| let-7a | LR | SVC | ||||
| mR-30c | RFC, XGB | |||||
| miR-323-3p | RFC | XGB | ||||
| 16 weeks | miR-720 | LR, SVC, MLP | ||||
| let-7c | SVC, MLP | |||||
| miR-1 | SVC, MLP | LR | ||||
| miR-205 | LR, SVC | MLP | ||||
| miR-339-5p | RFC, XGB | |||||
| 8 and 16 weeks | miR-22 | LR | SVC, MLP | |||
| miR-720 | SVC, MLP | LR | ||||
| let-7a | LR, SVC, MLP | |||||
| let-7c | LR, SVC, MLP | |||||
| miR-1 | LR, SVC | |||||
| miR-339-5p | RFC, XGB | |||||
| miR-323-3p | RFC | XGB | ||||
| miR-142-5p | RFC, XGB | |||||
Cells with a gray background identify cases in which the two more models agreed on both the importance and miRNA.
3. Discussion
The sex-specific DE miRNA reported in the partial human mouth microbes (PAMHH)- ETSPPI model that utilized five different bacteria including S. gordonii (early bacterial colonizer), F. nucleatum (intermediate bacterial colonizer), P. gingivalis, T. denticola and T. forsythia (late bacterial colonizers) using a time sequential infection [19]. Recently, we reported a polybacterial PD mouse model that induced miRNA expression, but the novelty of studying specific bacterium S. gordonii alone in inducing PD and specific miRNAs DE was not examined. Specific signature DE miRNAs in P. gingivalis [30], T. denticola [25] and T. forsythia [23] monoinfections were reported. We examined S. gordonii bacterium colonization on the mouse gingival surface, horizontal ABR measurements, intravascular dissemination of S. gordonii to distal organs, and global miRNA profiling in S. gordonii mice infected intraorally at 8 weeks and 16 weeks. Mice infected with S. gordonii at both time points indicated 100% bacterial colonization on the gingival surface, which was documented with 16S rRNA gene amplification. Similarly, S. gordonii significantly induced higher ABR in mice at both 8- and 16-week infection time-points. To the best of our knowledge, this is the first report that live S. gordonii a supragingival Gram-positive bacterium, significantly colonized the gingival epithelium, and induced higher ABR during intraoral infection. These results are in accordance with the report that S. gordonii in the periapical lesions of patients with apical periodontitis may contribute to the induction of ABR [3]. Among the 201 DE upregulated miRNAs in 8-week and 16-week S. gordonii-infected mice, eight of the miRNAs were shown to be observed in human periodontitis patients indicating its robust miRNA genesis in induction of PD (miR-323-3p, miR-200b, miR-129-3p, miR-205, miR-203, miR-210, miR-423-5p, and miR-146b). Four of the DE miRNAs, miR-187, miR-671-3p, miR-509-5p, and miR-767 were associated with cardiovascular diseases. Five DE miRNAs, miR-135a, miR-187, miR-1187, miR-291a-3p, and miR-2861 were also reported in the osteocyte regulation. Two of the miRNAs, miR-485 and miR-101b, are associated with Alzheimer’s disease. All of the 39 DE miRNAs were reported to be observed in S. gordonii-induced periodontitis. The reviewed observations of these miRNAs in the human periodontal gingival tissues, S. gordonii-induced PD in mice models highlight the importance of oral bacterial species normally associated with oral health to understand the present reported miRNAs as potential biomarkers for S. gordonii-induced periodontitis.
The inflammatory miRNA (miR-146b) reported in the PD subjects is also upregulated in the 8- and 16-week S. gordonii-infected mice [120]. Thirty-three miRNAs (e.g., miR-375, 148a, 574-3p, 382, and 181c) DE in the polymicrobial infection [19] are also expressed in the present S. gordonii monobacterial infection (Female comparison). Twenty-three miRNAs (e.g., miR-200b, 2141, 202-5p, and 1902) DE in the polymicrobial infection [19] are also expressed in the present S. gordonii monobacterial infection (Male comparison). The seven miRNAs (miR-31, 152, 423-5p, 191, 103, 30d, and 30c) that were DE during the P. gingivalis monobacterial infection [30] are observed in the present S. gordonii monoinfection. The five miRNAs (miR-2135, 30c, 126-5p, 423-5p, and 101b) reported in the T. denticola monobacterial infection [25] are also observed in the present S. gordonii monoinfection. The eight miRNAs (miR-1902, let-7c, 423-5p, 210, let-7a, 98, 876-3p, and 101b) reported in the T. forsythia monoinfection [23] are also induced in the present S. gordonii monoinfection. These data indicate that most of the periodontal-causing bacteria rely on the common miRNA activation pathways and initiate periodontitis.
The DE upregulated miRNAs (out of 191, only 20 miRNAs used in pathway analysis) in the 8-week S. gordonii infection involved in the pluripotency of stem cells pathways, MAPK-signaling pathway, Axon guidance, Wnt signaling pathway, Arrhythmogenic right ventricular cardiomyopathy, mTOR signaling pathway, and thyroid hormone signaling pathway. Among these pathways, bacterial invasion of epithelial cells was highlighted showing the number of genes altered by the associated 11 miRNAs. The DE upregulated miRNAs in the 16-week S. gordonii infection involved in axon guidance pathways, MAPK signaling pathway, Wnt signaling pathway (bone formation), protein digestion and absorption pathways, and focal adhesion pathways.
Interestingly, miRNA-375 is a novel multifunctional regulator miRNA that interacts with a large number of 27 target genes and is involved in the regulation of the differentiation and functioning of cells of the nervous and immune systems, bone and adipose tissue, and development and virus (HPV, HBV, and HIV) replication [121]. Anomalous expressions of miR-375 are found in multitudinous diseases, including PD during polymicrobial infections, T. denticola, T. forsythia, and S. gordonii (8 and 16 weeks) monoinfections, carcinogenesis, inflammation, and autoimmune and cardiovascular diseases. miR-375 is one of the most frequently down-regulated miRNAs in esophageal cancer [122], H. pylori-induced gastric carcinogenesis, and AIDS-KS patients after cART [123]. Gut epithelium-expressed miR-375 has been shown as a key regulator of epithelial properties (gut mucosal immunity) that are necessary for securing epithelium-immune system crosstalk in miR-375-deficient mice [124]. A recent review stated that miR-375 attenuates PD-1/PD-L1 signaling [125], a significant immune checkpoint mentioned in the 2018 Nobel Prize in Physiology or Medicine, indicating that miR-375 can be studied as a latent immune checkpoint modulator in immune investigations [121]. The major limitation of the current study is the inability to use blood and salivary secretions in this microRNA analysis.
For the machine learning analysis, the differences between the tree-based models (RFC, XGB) and non-tree-based models (LR, SVC, MLP) are quite interesting. First, we did not find any overlap between the miRNAs determined to be important for each of these groups of models. Second, both the tree-based and non-tree-based models agreed on their most important miRNAs for the 8-week and 16-week datasets. However, for the 8- and 16-week datasets, we found that LR differed from SVC and MLP. The reasons for these differences need to be investigated.
It is also important to note that we did not include an in-depth discussion of the ML metrics for each model. Each model scored between 99–100% for accuracy, precision, recall, and F1 score. This almost certainly indicates that the models have overfit the data. The reason for this is that due to the small number of mice used in the study, it is very difficult (most likely impossible) to divide the study groups into meaningful subgroups that can be used for training and testing. Instead, we have focused on using ML to gain insights into which miRNAs may be most relevant in identifying infected mice. To meaningfully score our models, we would need to train and test our models on large cohorts of mice.
In a meta-analysis study, Li et al. analyzed and predicted miR-152-3p and miR-34a-5p neurological outcomes in patients with subarachnoid hemorrhage [126]. The studies of Chi et al. predicted 30 miRNA features in pancreatic cancer and Li et al. used RFA in predicting potential miRNAs in diabetes [127,128]. Multilayer perceptron was used to improve the prediction performance in the miRNA and disease associations in lymphoma (e.g., hsa-let-7a), leukemia (e.g., hsa-miR-218), kidney neoplasms (e.g., has-miR-494), colon neoplasms (e.g., has-miR-30e), and breast neoplasms (e.g., has-miR-106a) [129].
4. Materials and Methods
4.1. Animal Models and Ethics Statement
All the animal procedures were performed according to the guidelines (University of Florida Institutional Animal Care and Use Committee protocol number 202200000223). This preclinical mouse study complied with the ARRIVE guidelines (Animal Research: Reporting In Vivo Experiments). Forty 8-week-old wild-type C57BL/6 mice, containing 20 males and 20 females used. The animals housed in a controlled temperature environment with 12 h dark/light cycles and access to mice chow and water ad libitum. The sample size was determined based on our previous studies [19,23,25,30].
4.2. Bacterial Culture and Mice Oral Administration
S. gordonii (Sg) DL1 was grown in Brucella blood agar plates, supplemented with hemin and vitamin K (Hardy Diagnostics, Santa Maria, CA, USA) for three days in Coy anaerobic chamber [19,23,25,30]. A bacterial cell suspension was prepared by combining an equal volume of reduced transport fluid (RTF) and 6% carboxymethyl cellulose (CMC) for intraoral infection in mice [19]. Both genders of mice were divided randomly into four groups (n =10) (Group-I: Sg-infected for 8 weeks; Group II: Sg-infected for 16 weeks; Group-III: sham-infected for 8 weeks; Group-IV: sham-infected for 16 weeks) (Table 1). Before the bacterial infection, mouse oral microflora was suppressed with Kanamycin (500 mg) dissolved in two liters of sterile water for 3 days followed by rinsing with 0.12% of chlorhexidine gluconate ((Peridex: 3M ESPE Dental Products, St. Paul, MN, USA) [19,25,30]. After the kanamycin antibiotic washout period, mice in Groups I and II underwent infection with S. gordonii (108 cells) as described previously [19,23,25,30]. Four bacterial oral infection cycles, as described, were performed for Group I (8 weeks) and eight infection cycles, as described [30], were performed for Group II mice (16 weeks). One infection cycle consists of four days per week of S. gordonii intraoral infection/vehicle control suspension for every alternative week (Table 1 and Figure 1A). An equal volume of RTF and CMC was used as a vehicle control for sham-infected mice (Groups III and IV). These sham-infected mice groups served as common sham-infection controls for 5 different monoinfection mice studies performed simultaneously [23,25,30]. At the end of 8 and 16 weeks, the mice were euthanized by the carbon dioxide (CO2) inhalation method; blood was collected by cardiac puncture and serum was separated. Additionally, distal organs such the brain, heart, liver, lungs, spleen, and kidneys were collected. The left maxilla and mandibles were also collected in RNAlater for miRNA analysis, whereas the right maxilla and mandibles were designated for morphometry analysis of horizontal alveolar bone levels [19,23,25,30].
4.3. Molecular Detection of Bacteria
Two days after each Sg-infection cycle, gingival plaque samples were collected from gingival surfaces of the mice using sterile cotton swabs and the swabs were submerged in Tris EDTA (TE) buffer [19,23,25,30]. Colony PCR was used to investigate the Sg bacteria in gingival plaque samples. Genomic DNA from the heart, lungs, liver, spleen, brain, and kidney were extracted following the standard procedure described in Qiagen DNeasy Blood and Tissue Kit(Qiagen, Germantown, MD, USA)and stored at −20 °C [19,23,25,30]. The presence of S. gordonii genomic DNA was confirmed using PCR with primers specific to the 16S rRNA gene as described previously [19,130]. The forward primer 5′-GTAGCTTGCTACACCATAGA-3′ and reverse primer 5′-CTCACACCCGTTCTTCTCTT-3′ used to amplify the S. gordonii DL1-16S rRNA gene [19]. DNA extracted from S. gordonii DL1 bacterial culture was used as positive control and sterile Milli-Q water was considered negative control in the PCR reaction. The PCR amplified products were run on 1% agarose gel electrophoresis and visualized under UVP GelStudio touch Imaging System (Analytik Jena US LLC, CA, USA).
4.4. Measurement of Alveolar Bone Resorption (ABR)
The impact of S. gordonii infection and sham infection on ABR was quantitatively assessed using histomorphometry, as described previously [19,23,25,30]. Briefly, the jaw specimens (mandibles and maxilla) were autoclaved and defleshed. H2O2 (3%) solution was used to clean the jaw bone and air dried [19]. Two-dimensional alveolar bone imaging was performed using a stereo dissecting microscope (Stereo Discovery V8, Carl Zeiss Microimaging, Inc., Thornwood, NY, USA). A line tool was used to measure the horizontal ABR between cementoenamel junction and alveolar bone crest (AxioVision LE 29A 4.6.3, Thornwood, NY, USA) [19,23,30,130]. Two examiners blinded to the S. gordonii infection and sham infection were measured the ABR.
4.5. Total RNA Isolation and Quality Assessment
Total RNA was extracted from left mandibles using the mirVanaTM miRNA Isolation Kit, following the manufacturer’s instructions, as described previously (Ambion, Austin, TX, USA) [19,23,30,130]. The final RNA yield and purity was assessed using spectrophotometric measurements. RNA samples exhibiting an OD 260/230 ratio of >2 and an OD 260/280 ratio of >2 were considered of sufficient quality for further NanoString (NanoString Technologies, Seattle, WA, USA) analysis.
4.6. NanoString nCounter miRNA Panel Analysis
For transcriptome analysis, total RNA extracted from the left mandibles was analyzed using NanoString nCounter® Mouse miRNA Assay kit v1.5, a high-throughput nCounter® miRNA Expression Panels to identify differential expression (DE) of miRNAs in 8- and 16-week S. gordonii-infected mice and sham-infected mice. Similar NanoString analysis was performed in polymicrobial [19], P. gingivalis [30], T. denticola [25], and T. forsythia-induced periodontal infections [23]. The NanoString nCounter® panel is capable of simultaneously identifying and quantifying up to 577 miRNAs in a single RNA specimen by using molecular barcodes, called nCounter reporter probes. The NanoString capacity to detect miRNAs without requiring reverse transcription or amplification [19]. Briefly, nCounter® miRNA assay panel kit has sample preparation steps of annealing, ligation, and purification. The Master mix preparation and thermal cycler conditions are explained more in our recent publications. Reported probe and miRNA capture probes was added after the purification steps. The strip tubes were incubated at 65 °C for 18 h in the thermal cycler, and the samples were immediately processed for post-hybridization with an nCounter analysis system in the Molecular Pathology Core at the University of Florida. A separate reporter code count (RCC) file for each sample containing the count for each probe was downloaded and used for miRNA data analysis.
4.7. NanoString Data Analysis
Data obtained from the NanoString platform were analyzed using the dedicated software nSolver™ 4.0 (NanoString Technologies, Seattle, WA, USA). The codeset incorporated 577 mouse miRNAs and included 6 ligation controls, and five miRNA reference controls (ACTB, B2M, GAPDH, RPL19, RPLP0). Data normalization was performed using nSolver Analysis Software v4.0 (NanoString Technologies). RCC files were downloaded and imported into nSolver and imaging quality control (QC), binding density QC, positive control linearity QC and positive control limit of detection QC were carried out as recommended in the system QC parameters. The lanes were flagged when the percent field of view (FOV) registration was <75% for imaging QC, binding density was outside the range from 0.1 to 2.25 for binding density QC, and the positive control R2 value was <0.95. All 40 samples passed the QC, and no flag lanes were observed. Raw data were created after passing the QC. To reduce the background signal/noise, the background threshold count value was set to 52 and analyzed by taking an average of eight negative control probe counts from all 40 samples. Codeset content normalization parameters were chosen, and the data normalization was performed based on the top 100 miR genes expressed. The normalized factor was calculated based on the geometric mean values of miR genes expressed in each specimen [19,23,25,30].
4.8. Bioinformatics Analysis
All the miRNA normalized data were analyzed further using ROSALIND® (https://rosalind.bio/ (accessed on 21 September 2023), with a HyperScale architecture developed by ROSALIND®, Inc. (San Diego, CA, USA) [131]. Fold changes (FC) in the miRNA genes were calculated based on the ratio of the difference in the means of the log-transformed normalized data to the square root of the sum of the variances of the specimens in the four groups. The Limma R library [132] was used to calculate FCs. The clustering of miRNA for the final heatmap of DE miRNA was performed using the Partitioning Around Medoids method with the fpc R library. MiRTarBase was used to investigate validated miRNA-target gene interactions (miRTarbase update 2022: an informative resource for experimentally validated miRNA-target interactions) [133]. Venn diagram for higher expression and dysregulated miRNAs in the 8-week and 16-week infection groups was drawn using Venny 2.1 [19,23,25,30].
4.8.1. Kyoto Encyclopedia of Genes and Genomes (KEGG)
KEGG is an integrated database from the National Center for Biotechnology Information RefSeq and GenBank and KEGG orthology [134]. KEGG pathways were plotted using DIANA-miRPath v.3.0 database [135] using the MIMAT accession number with the threshold values of p < 0.05 and false discovery rate (FDR) correction applied to obtain unbiased empirical distribution using the Benjamin and Hochberg method [19,23,25,30]. KEGG interprets the data from high-throughput experimental technologies. KEGG explores the possible key regulatory pathways for the enrichment of DE genes between the sham infection and S. gordonii mice infection.
4.8.2. Multiple Machine Learning (ML) Analysis
To further investigate the impact of S. gordonii on PD, we used ML algorithms to analyze the NanoString data from the infected mice at the 8- and 16-week time points as well as the dataset formed by combining the 8- and 16-week datasets. For our analysis, we used XGBoost version 1.7.6 and the Scikit-learn version 1.3.0 implementations of LR, SVC, MLP, and RFC. The ML models were created and executed using Python version 3.11.4. LR [136], SVC [137] and RFC [138] were chosen because, historically, they have been extensively studied and the underlying mathematics is well understood. MLP was chosen because recent advances in computing power and ML libraries (e.g., Scikit-learn) now permit deep neural networks to be more easily set up and executed [139]. Moreover, the application of the MLP model to novel domains is an active area of research. Finally, XGB was chosen because of its widespread use in biomedical research [140]. For instance, on February 14, 2024, a search on PubMed for ‘XGBoost’ found 3995 articles.
LR uses the natural log of the odds ratio to predict the outcome of a binary variable [136]. Typically, a threshold of 0.5 is to determine whether the outcome is positive (e.g., a value of 1) or negative (e.g., a value of 0). SVC classifies data by calculating a hyperplane that maximally separates the target classes in a high-dimensional space. MLP is a type of neural network that uses multiple interconnected nodes and layers to learn the relations between linear and non-linear data and predict outcomes using the learned relations. RFC is an ensemble ML method that builds multiple decision trees (i.e., a “forest of trees”) and predicts an outcome by polling the decisions of the constructed trees. XBG, like RFC, constructs multiple decision trees, but XBG enhances the RFC approach by incorporating gradient boosting.
To address implicit biases in the data, we randomly shuffled the order of the rows and columns before analysis. We used scikit-learn’s RandomizedGridSearchCV method to determine the hyperparameters for each ML model. The RandomizedGridSearchCV method was executed using between 2 and 11 cross-validation splits of the data, and the performance of each cross-validation split was evaluated using the LeaveOneOut method. A full list of the hyperparameters used for each ML model is available in Appendix S1 (Supplementary). The LeaveOneOut method was chosen because of the small number of samples available in the data. SHAP version 0.42.1 was used to obtain feature importance results [23,140,141]. The code for executing the ML models on the NanoString copy data is available on GitHub at (https://github.com/uflcod/miRNA-periodontal-disease, accessed on 15 December 2023) in the ‘notebooks’ directory. The notebooks for analyzing the S. gordonii NanoString data begin with the prefix ‘sg_’ (e.g., sg_randomforest_miRNA.ipynb, sg_xgboost_miRNA.ipynb).
4.9. Statistical Analysis
All the data in the graphs were presented as mean ± SEM. Ordinary two-way ANOVA with Tukey’s multiple comparison test with a single pooled variance was performed for ABR measurements to identify the statistical significance using Prism 9.4.1 (GraphPad Software, San Diego, CA, USA) [19,23,25,30,142]. A p-value of <0.05 was considered statistically significant. NanoString data were used to obtain the miRNA expression in each specimen as an FC value, and an FC of ±1 was considered significant. Identification of S. gordonii-induced significant differential gene expression was done based on two-tailed t-tests on the log-transformed normalized data that assumed unequal variance. The distribution of the t-statistics was calculated using the Welch–Satterthwaite equation for the degrees of freedom to estimate the 95% confidence intervals for the identified DE of miRNA between infection and sham-infection mice. The volcano plot was drawn using GraphPad Software [19,23,25,30].
5. Conclusions
Over the past few years, critical knowledge of oral microbe-induced miRNAs is developing rapidly during PD pathogenesis as well as from human PD with and without diabetes and obesity. This study’s findings (attachment/colonization, ABR) demonstrates that S. gordonii a commensal oral bacterium can cause full development of PD. Out of several miRs’ anomalous expression (up- and downregulation) during S. gordonii bacterium monoinfection, a few miRNAs such as miR-375, miR-1902, miR-34b-5p are critical novel biomarker miRNAs during PD pathology. Specifically, multi-functional regulator miR-375 altered expression during monoinfection with T. denticola, T. forsythia, S. gordonii and several other diseases and diverse cancers indicating its use in diagnosis and clinical application.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25116217/s1. References [25,50,58,93,99,102,103,108,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175] are cited in the supplementary file.
Author Contributions
C.A. performed all the experiments and analyzed the data. K.M.V. did the molecular analysis of distal organs dissemination. S.J. performed bioinformatic analyses and submission of data at NCBI. W.D.D. performed multiple ML models; L.K. was responsible for the conception, experimental design, analysis, initial drafting, editing, supervision, project administration, interpretation, and funding acquisition. S.O.R. previewed the manuscript. L.K. and S.J. were involved in the final revision and editing the manuscript. L.K. and E.K.L.C. are the Principal Investigators of the NIH (NIDCR) study. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the NIH National Institute of Dental and Craniofacial Research (NIDCR) (R01 DE028536) to L. Kesavalu and E.K.L. Chan. The funders had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
All animal procedures were approved by the University of Florida Institutional Animal Care and Use Committee (IACUC) under protocol number 202200000223.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are openly available in NCBI https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE240502 (accessed on 15 August 2023).
Acknowledgments
The authors acknowledge Andreas Gonzalez for reviewing and editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Abranches, J.; Zeng, L.; Kajfasz, J.K.; Palmer, S.R.; Chakraborty, B.; Wen, Z.T.; Richards, V.P.; Brady, L.J.; Lemos, J.A. Biology of Oral Streptococci. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Okahashi, N.; Nakata, M.; Kuwata, H.; Kawabata, S. Oral mitis group streptococci: A silent majority in our oral cavity. Microbiol. Immunol. 2022, 66, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Chavez de Paz, L.; Svensater, G.; Dahlen, G.; Bergenholtz, G. Streptococci from root canals in teeth with apical periodontitis receiving endodontic treatment. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. Endod. 2005, 100, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Mosailova, N.; Truong, J.; Dietrich, T.; Ashurst, J. Streptococcus gordonii: A Rare Cause of Infective Endocarditis. Case Rep. Infect. Dis. 2019, 2019, 7127848. [Google Scholar] [CrossRef]
- Chang, C.Y.; Gan, Y.L.; Radhakrishnan, A.P.; Ong, E.L.C. Acute abdomen revealed Streptococcus gordonii infective endocarditis with systemic embolism. Oxf. Med. Case Rep. 2022, 2022, omab145. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Hu, H.; Fang, W.; Shi, D.; Liang, C.; Sun, Y.; Gao, G.; Wang, H.; Zhang, Q.; Wang, L.; et al. Detection of pathogens from resected heart valves of patients with infective endocarditis by next-generation sequencing. Int. J. Infect. Dis. 2019, 83, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kilic, A.O.; Tao, L.; Zhang, Y.; Lei, Y.; Khammanivong, A.; Herzberg, M.C. Involvement of Streptococcus gordonii beta-glucoside metabolism systems in adhesion, biofilm formation, and in vivo gene expression. J. Bacteriol. 2004, 186, 4246–4253. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Kreth, J. The role of hydrogen peroxide in environmental adaptation of oral microbial communities. Oxid. Med. Cell Longev. 2012, 2012, 717843. [Google Scholar] [CrossRef]
- Park, O.J.; Kwon, Y.; Park, C.; So, Y.J.; Park, T.H.; Jeong, S.; Im, J.; Yun, C.H.; Han, S.H. Streptococcus gordonii: Pathogenesis and Host Response to Its Cell Wall Components. Microorganisms 2020, 8, 1852. [Google Scholar] [CrossRef]
- Daep, C.A.; Lamont, R.J.; Demuth, D.R. Interaction of Porphyromonas gingivalis with oral streptococci requires a motif that resembles the eukaryotic nuclear receptor box protein-protein interaction domain. Infect. Immun. 2008, 76, 3273–3280. [Google Scholar] [CrossRef]
- Yang, R.; Liu, T.; Pang, C.; Cai, Y.; Lin, Z.; Guo, L.; Wei, X. The Regulatory Effect of Coaggregation Between Fusobacterium nucleatum and Streptococcus gordonii on the Synergistic Virulence to Human Gingival Epithelial Cells. Front. Cell Infect. Microbiol. 2022, 12, 879423. [Google Scholar] [CrossRef]
- Antezack, A.; Etchecopar-Etchart, D.; La Scola, B.; Monnet-Corti, V. New putative periodontopathogens and periodontal health-associated species: A systematic review and meta-analysis. J. Periodontal Res. 2023, 58, 893–906. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Chen, J.; Cai, Z.; Hu, J.; Zhou, L.; Zhang, P.; Xu, X. MicroRNA-375 in extracellular vesicles—Novel marker for esophageal cancer diagnosis. Medicine 2023, 102, e32826. [Google Scholar] [CrossRef]
- Coskun, M.; Bjerrum, J.T.; Seidelin, J.B.; Troelsen, J.T.; Olsen, J.; Nielsen, O.H. miR-20b, miR-98, miR-125b-1*, and let-7e* as new potential diagnostic biomarkers in ulcerative colitis. World J. Gastroenterol. 2013, 19, 4289–4299. [Google Scholar] [CrossRef]
- Mehterov, N.; Vladimirov, B.; Sacconi, A.; Pulito, C.; Rucinski, M.; Blandino, G.; Sarafian, V. Salivary miR-30c-5p as Potential Biomarker for Detection of Oral Squamous Cell Carcinoma. Biomedicines 2021, 9, 1079. [Google Scholar] [CrossRef]
- Vallelunga, A.; Iannitti, T.; Dati, G.; Capece, S.; Maugeri, M.; Tocci, E.; Picillo, M.; Volpe, G.; Cozzolino, A.; Squillante, M.; et al. Serum miR-30c-5p is a potential biomarker for multiple system atrophy. Mol. Biol. Rep. 2019, 46, 1661–1666. [Google Scholar] [CrossRef]
- Aravindraja, C.; Kashef, M.R.; Vekariya, K.M.; Ghanta, R.K.; Karanth, S.; Chan, E.K.L.; Kesavalu, L. Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. Int. J. Mol. Sci. 2022, 23, 5107. [Google Scholar] [CrossRef]
- Tubero Euzebio Alves, V.; Al-Attar, A.; Alimova, Y.; Maynard, M.H.; Kirakodu, S.; Martinez-Porras, A.; Hawk, G.S.; Ebersole, J.L.; Stamm, S.; Gonzalez, O.A. Streptococcus gordonii-Induced miRNAs Regulate CCL20 Responses in Human Oral Epithelial Cells. Infect. Immun. 2022, 90, e0058621. [Google Scholar] [CrossRef]
- Zhou, W.; Su, L.; Duan, X.; Chen, X.; Hays, A.; Upadhyayula, S.; Shivde, J.; Wang, H.; Li, Y.; Huang, D.; et al. MicroRNA-21 down-regulates insflammation and inhibits periodontitis. Mol. Immunol. 2018, 101, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Shao, P.; Wang, L.; Bao, X.; Hu, M. IL1β inhibits differentiation of cementoblasts via microRNA-325-3p. J. Cell Biochem. 2020, 121, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Aravindraja, C.; Jeepipalli, S.; Duncan, W.; Vekariya, K.M.; Bahadekar, S.; Chan, E.K.L.; Kesavalu, L. Unique miRomics Expression Profiles in Tannerella forsythia-Infected Mandibles during Periodontitis Using Machine Learning. Int. J. Mol. Sci. 2023, 24, 16393. [Google Scholar] [CrossRef] [PubMed]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE-/- mice during experimental periodontal disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Aravindraja, C.; Jeepipalli, S.; Vekariya, K.M.; Botello-Escalante, R.; Chan, E.K.L.; Kesavalu, L. Oral Spirochete Treponema denticola Intraoral Infection Reveals Unique miR-133a, miR-486, miR-126-3p, miR-126-5p miRNA Expression Kinetics during Periodontitis. Int. J. Mol. Sci. 2023, 24, 12105. [Google Scholar] [CrossRef] [PubMed]
- Motedayyen, H.; Ghotloo, S.; Saffari, M.; Sattari, M.; Amid, R. Evaluation of MicroRNA-146a and Its Targets in Gingival Tissues of Patients With Chronic Periodontitis. J. Periodontol. 2015, 86, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.F.; Shu, R.; Jiang, S.Y.; Liu, D.L.; Zhang, X.L. Comparison of microRNA profiles of human periodontal diseased and healthy gingival tissues. Int. J. Oral. Sci. 2011, 3, 125–134. [Google Scholar] [CrossRef]
- Amaral, S.A.; Pereira, T.S.F.; Brito, J.A.R.; Cortelli, S.C.; Cortelli, J.R.; Gomez, R.S.; Costa, F.O.; Miranda Cota, L.O. Comparison of miRNA expression profiles in individuals with chronic or aggressive periodontitis. Oral. Dis. 2019, 25, 561–568. [Google Scholar] [CrossRef]
- Nik Mohamed Kamal, N.N.S.; Awang, R.A.R.; Mohamad, S.; Shahidan, W.N.S. Plasma- and Saliva Exosome Profile Reveals a Distinct MicroRNA Signature in Chronic Periodontitis. Front. Physiol. 2020, 11, 587381. [Google Scholar] [CrossRef]
- Aravindraja, C.; Vekariya, K.M.; Botello-Escalante, R.; Rahaman, S.O.; Chan, E.K.L.; Kesavalu, L. Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis. Int. J. Mol. Sci. 2023, 24, 2327. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Rivera, M.F.; Velsko, I.M.; Lee, J.Y.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Invasion of oral and aortic tissues by oral spirochete Treponema denticola in ApoE(-/-) mice causally links periodontal disease and atherosclerosis. Infect. Immun. 2014, 82, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Usui, M.; Onizuka, S.; Sato, T.; Kokabu, S.; Ariyoshi, W.; Nakashima, K. Mechanism of alveolar bone destruction in periodontitis—Periodontal bacteria and inflammation. Jpn. Dent. Sci. Rev. 2021, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Deppe, H.; Reitberger, J.; Behr, A.V.; Vitanova, K.; Lange, R.; Wantia, N.; Wagenpfeil, S.; Sculean, A.; Ritschl, L.M. Oral bacteria in infective endocarditis requiring surgery: A retrospective analysis of 134 patients. Clin. Oral. Investig. 2022, 26, 4977–4985. [Google Scholar] [CrossRef] [PubMed]
- Gallelli, L.; Cione, E.; Peltrone, F.; Siviglia, S.; Verano, A.; Chirchiglia, D.; Zampogna, S.; Guidetti, V.; Sammartino, L.; Montana, A.; et al. Hsa-miR-34a-5p and hsa-miR-375 as Biomarkers for Monitoring the Effects of Drug Treatment for Migraine Pain in Children and Adolescents: A Pilot Study. J. Clin. Med. 2019, 8, 928. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Q.; Wen, Z.; Feng, H.; Deng, X.; Ci, X. Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3beta-Nrf2 signal axis. Redox Biol. 2017, 12, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lu, Q.; Wang, K.; Lu, J.; Gu, X.; Zhu, D.; Liu, F.; Guo, Z. miR-34b-5p inhibition attenuates lung inflammation and apoptosis in an LPS-induced acute lung injury mouse model by targeting progranulin. J. Cell Physiol. 2018, 233, 6615–6631. [Google Scholar] [CrossRef] [PubMed]
- Talebi, F.; Ghorbani, S.; Chan, W.F.; Boghozian, R.; Masoumi, F.; Ghasemi, S.; Vojgani, M.; Power, C.; Noorbakhsh, F. MicroRNA-142 regulates inflammation and T cell differentiation in an animal model of multiple sclerosis. J. Neuroinflamm. 2017, 14, 55. [Google Scholar] [CrossRef]
- Korotkov, A.; Puhakka, N.; Gupta, S.D.; Vuokila, N.; Broekaart, D.W.M.; Anink, J.J.; Heiskanen, M.; Karttunen, J.; van Scheppingen, J.; Huitinga, I.; et al. Increased expression of miR142 and miR155 in glial and immune cells after traumatic brain injury may contribute to neuroinflammation via astrocyte activation. Brain Pathol. 2020, 30, 897–912. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, R.L.; Croce, C.M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. A program of microRNAs controls osteogenic lineage progression by targeting transcription factor Runx2. Proc. Natl. Acad. Sci. USA 2011, 108, 9863–9868. [Google Scholar] [CrossRef]
- Imamura, K.; Tachi, K.; Takayama, T.; Shohara, R.; Kasai, H.; Dai, J.; Yamano, S. Released fibroblast growth factor18 from a collagen membrane induces osteoblastic activity involved with downregulation of miR-133a and miR-135a. J. Biomater. Appl. 2018, 32, 1382–1391. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, Z. MicroRNA-135a-5p is involved in osteoporosis progression through regulation of osteogenic differentiation by targeting RUNX2. Exp. Ther. Med. 2019, 18, 2393–2400. [Google Scholar] [CrossRef]
- Kalea, A.Z.; Hoteit, R.; Suvan, J.; Lovering, R.C.; Palmen, J.; Cooper, J.A.; Khodiyar, V.K.; Harrington, Z.; Humphries, S.E.; D’Aiuto, F. Upregulation of gingival tissue miR-200b in obese periodontitis subjects. J. Dent. Res. 2015, 94, 59S–69S. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Gao, F.; Peng, C.G.; Zheng, C.J.; Wu, M.F. miR-485-5p promotes osteoporosis via targeting Osterix. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4792–4799. [Google Scholar] [CrossRef] [PubMed]
- Ryu, I.S.; Kim, D.H.; Cho, H.J.; Ryu, J.H. The role of microRNA-485 in neurodegenerative diseases. Rev. Neurosci. 2023, 34, 49–62. [Google Scholar] [CrossRef]
- Kearney, M.; Cooper, P.R.; Smith, A.J.; Duncan, H.F. Characterisation of miRNA Expression in Dental Pulp Cells during Epigenetically-Driven Reparative Processes. Int. J. Mol. Sci. 2023, 24, 8631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiang, D.; Hu, X.; Ruan, Q.; Wang, L.; Bao, Z. Identification and study of differentially expressed miRNAs in aged NAFLD rats based on high-throughput sequencing. Ann. Hepatol. 2020, 19, 302–312. [Google Scholar] [CrossRef]
- Choi, M.R.; Han, J.S.; Jin, Y.B.; Lee, S.R.; Choi, I.Y.; Lee, H.; Cho, H.; Kim, D.J. Differential expression of microRNAs in the hippocampi of male and female rodents after chronic alcohol administration. Biol. Sex. Differ. 2020, 11, 65. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, W.; Xia, Q.; Lin, J.; Xu, J.; Zhang, S.; Tian, L.; Han, X. Construction of a potential microRNA and messenger RNA regulatory network of acute lung injury in mice. Sci. Rep. 2022, 12, 777. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Marugame, Y.; Yamada, S.; Lin, I.; Yamashita, S.; Fujimura, Y.; Tachibana, H. Circulating miRNA profiles in mice plasma following flavonoid intake. Mol. Biol. Rep. 2022, 49, 10399–10407. [Google Scholar] [CrossRef]
- Ektesabi, A.M.; Mori, K.; Tsoporis, J.N.; Vaswani, C.M.; Gupta, S.; Walsh, C.; Varkouhi, A.K.; Mei, S.H.J.; Stewart, D.J.; Liles, W.C.; et al. Mesenchymal Stem/Stromal Cells Increase Cardiac miR-187-3p Expression in a Polymicrobial Animal Model of Sepsis. Shock 2021, 56, 133–141. [Google Scholar] [CrossRef]
- Sinha, A.; Bhattacharjee, R.; Bhattacharya, B.; Nandi, A.; Shekhar, R.; Jana, A.; Saha, K.; Kumar, L.; Patro, S.; Panda, P.K.; et al. The paradigm of miRNA and siRNA influence in Oral-biome. Biomed. Pharmacother. 2023, 159, 114269. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Yang, K.; Yu, R.; Hui, H.; Wu, W. circ-Iqsec1 induces bone marrow-derived mesenchymal stem cell (BMSC) osteogenic differentiation through the miR-187-3p/Satb2 signaling pathway. Arthritis Res. Ther. 2022, 24, 273. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Du, X.; Liu, L.; Cao, Q.; Pan, Z.; Li, Q. miR-1306 Mediates the Feedback Regulation of the TGF-beta/SMAD Signaling Pathway in Granulosa Cells. Cells 2019, 8, 298. [Google Scholar] [CrossRef]
- Yoshioka, H.; Wang, Y.Y.; Suzuki, A.; Shayegh, M.; Gajera, M.V.; Zhao, Z.; Iwata, J. Overexpression of miR-1306-5p, miR-3195, and miR-3914 Inhibits Ameloblast Differentiation through Suppression of Genes Associated with Human Amelogenesis Imperfecta. Int. J. Mol. Sci. 2021, 22, 2202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Geng, Z.; Wan, Y.; Meng, F.; Meng, X.; Wang, L. Functional analysis of miR-767-5p during the progression of hepatocellular carcinoma and the clinical relevance of its dysregulation. Histochem. Cell Biol. 2020, 154, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, S.; Xu, J.; Li, Y.; Zhang, J.; Zhang, J.; Lu, X. miR-767-5p inhibits glioma proliferation and metastasis by targeting SUZ12. Oncol. Rep. 2019, 42, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tan, J.; Song, Q.; Yang, X.; Zhang, X.; Qin, H.; Huang, G.; Su, X.; Li, J. Exosomal miR-767 from senescent endothelial-derived accelerating skin fibroblasts aging via inhibiting TAB1. J. Mol. Histol. 2023, 54, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Reddy, P.H. A New Discovery of MicroRNA-455-3p in Alzheimer’s Disease. J. Alzheimers Dis. 2019, 72, S117–S130. [Google Scholar] [CrossRef]
- Chum, P.P.; Hakim, M.A.; Behringer, E.J. Cerebrovascular microRNA Expression Profile During Early Development of Alzheimer’s Disease in a Mouse Model. J. Alzheimers Dis. 2022, 85, 91–113. [Google Scholar] [CrossRef]
- Hu, D.L.; Liu, Y.Q.; Chen, F.K.; Sheng, Y.H.; Yang, R.; Kong, X.Q.; Cao, K.J.; Zhang, J.S.; Qian, L.M. Differential expression of microRNAs in cardiac myocytes compared to undifferentiated P19 cells. Int. J. Mol. Med. 2011, 28, 59–64. [Google Scholar] [CrossRef]
- Hu, S.; Huang, M.; Li, Z.; Jia, F.; Ghosh, Z.; Lijkwan, M.A.; Fasanaro, P.; Sun, N.; Wang, X.; Martelli, F.; et al. MicroRNA-210 as a novel therapy for treatment of ischemic heart disease. Circulation 2010, 122, S124–S131. [Google Scholar] [CrossRef]
- John, A.A.; Prakash, R.; Kureel, J.; Singh, D. Identification of novel microRNA inhibiting actin cytoskeletal rearrangement thereby suppressing osteoblast differentiation. J. Mol. Med. 2018, 96, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; He, Q. miR-1187 induces podocyte injury and diabetic nephropathy through autophagy. Diab. Vasc. Dis. Res. 2023, 20, 14791641231172139. [Google Scholar] [CrossRef]
- Yu, D.S.; An, F.M.; Gong, B.D.; Xiang, X.G.; Lin, L.Y.; Wang, H.; Xie, Q. The regulatory role of microRNA-1187 in TNF-alpha-mediated hepatocyte apoptosis in acute liver failure. Int. J. Mol. Med. 2012, 29, 663–668. [Google Scholar] [CrossRef]
- Li, Z.H.; Hu, H.; Zhang, X.Y.; Liu, G.D.; Ran, B.; Zhang, P.G.; Liao, M.M.; Wu, Y.C. MiR-291a-3p regulates the BMSCs differentiation via targeting DKK1 in dexamethasone-induced osteoporosis. Kaohsiung J. Med. Sci. 2020, 36, 35–42. [Google Scholar] [CrossRef]
- Ge, J.; Xu, W.J.; Chen, H.F.; Dong, Z.H.; Liu, W.; Nian, F.Z.; Liu, J. Induction mechanism of cigarette smoke components (CSCs) on dyslipidemia and hepatic steatosis in rats. Lipids Health Dis. 2022, 21, 117. [Google Scholar] [CrossRef]
- Cao, J.; Kuang, D.; Luo, M.; Wang, S.; Fu, C. Targeting circNCLN/miR-291a-3p/TSLP signaling axis alleviates lipopolysaccharide-induced acute lung injury. Biochem. Biophys. Res. Commun. 2022, 617, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Han, J.; Wang, D. Identification of key microRNAs and the underlying molecular mechanism in spinal cord ischemia-reperfusion injury in rats. PeerJ 2021, 9, e11454. [Google Scholar] [CrossRef]
- Li, S.; Wang, X.; Zhu, X.; Xue, Y.; Zhang, J.; Tan, X.; Deng, J.; Li, C.; Kuang, W.; Li, C. miR-1968-5p is involved in the pathogenesis of lupus nephritis of NZBWF1 mice by targeting csf1. Clin. Exp. Nephrol. 2021, 25, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Hochreuter, M.Y.; Altintas, A.; Garde, C.; Emanuelli, B.; Kahn, C.R.; Zierath, J.R.; Vienberg, S.; Barres, R. Identification of two microRNA nodes as potential cooperative modulators of liver metabolism. Hepatol. Res. 2019, 49, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Kenny, N.J.; Namigai, E.K.; Marletaz, F.; Hui, J.H.; Shimeld, S.M. Draft genome assemblies and predicted microRNA complements of the intertidal lophotrochozoans Patella vulgata (Mollusca, Patellogastropoda) and Spirobranchus (Pomatoceros) lamarcki (Annelida, Serpulida). Mar. Genom. 2015, 24 Pt 2, 139–146. [Google Scholar] [CrossRef]
- Cheng, L.; Fan, Y.; Cheng, J.; Wang, J.; Liu, Q.; Feng, Z. Long non-coding RNA ZFY-AS1 represses periodontitis tissue inflammation and oxidative damage via modulating microRNA-129-5p/DEAD-Box helicase 3 X-linked axis. Bioengineered 2022, 13, 12691–12705. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Xu, G.; Zhang, D.; Wang, R. The protective effects of the miR-129-5p/keap-1/Nrf2 axis on Ang II-induced cardiomyocyte hypertrophy. Ann. Transl. Med. 2021, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhao, H.; Gong, L.; Yao, L.; Li, Y.; Zhang, W. MicroRNA-129-3p functions as a tumor suppressor in serous ovarian cancer by targeting BZW1. Int. J. Clin. Exp. Pathol. 2018, 11, 5901–5908. [Google Scholar] [PubMed]
- Birkhauser, F.D.; Koya, R.C.; Neufeld, C.; Rampersaud, E.N.; Lu, X.; Micewicz, E.D.; Chodon, T.; Atefi, M.; Kroeger, N.; Chandramouli, G.V.; et al. Dendritic cell-based immunotherapy in prevention and treatment of renal cell carcinoma: Efficacy, safety, and activity of Ad-GM.CAIX in immunocompetent mouse models. J. Immunother. 2013, 36, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Liu, W.; Li, H.; Yang, L.; Chen, C.; Xia, Z.Y.; Guo, L.J.; Xie, H.; Zhou, H.D.; Wu, X.P.; et al. A Runx2/miR-3960/miR-2861 regulatory feedback loop during mouse osteoblast differentiation. J. Biol. Chem. 2011, 286, 12328–12339. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wan, X.; Chen, X.; Fang, Y.; Cheng, X.; Xie, X.; Lu, W. miR-2861 acts as a tumor suppressor via targeting EGFR/AKT2/CCND1 pathway in cervical cancer induced by human papillomavirus virus 16 E6. Sci. Rep. 2016, 6, 28968. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, H.; Zhang, P.; Li, J.; Shan, Z.; Teng, W. Upregulation of miR-2861 and miR-451 expression in papillary thyroid carcinoma with lymph node metastasis. Med. Oncol. 2013, 30, 577. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Askari, A.; Hussen, B.M.; Rasul, M.F.; Hatamian, S.; Taheri, M.; Kiani, A. A review on the role of miR-671 in human disorders. Front. Mol. Biosci. 2022, 9, 1077968. [Google Scholar] [CrossRef]
- Lu, G.F.; You, C.Y.; Chen, Y.S.; Jiang, H.; Zheng, X.; Tang, W.W.; Wang, X.Y.; Xu, H.Y.; Geng, F. MicroRNA-671-3p promotes proliferation and migration of glioma cells via targeting CKAP4. OncoTargets Ther. 2018, 11, 6217–6226. [Google Scholar] [CrossRef] [PubMed]
- Goldberger, N.; Walker, R.C.; Kim, C.H.; Winter, S.; Hunter, K.W. Inherited variation in miR-290 expression suppresses breast cancer progression by targeting the metastasis susceptibility gene Arid4b. Cancer Res. 2013, 73, 2671–2681. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.T.; Sun, H.X.; Zhou, X.H.; Tang, B.P. Long non-coding RNA DANCR alleviates acute myocardial infarction damage via regulating microRNA-509-5p/KLF transcription factor 13 pathway. Kaohsiung J. Med. Sci. 2023, 39, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Fang, X.; Zhou, J.; Zeng, L.; Yu, B. Identification and validation of miR-509-5p as a prognosticator for favorable survival in osteosarcoma. Medicine 2022, 101, e29705. [Google Scholar] [CrossRef]
- Jiang, F.; Zhou, Y.; Zhang, R.; Wen, Y. miR-205 and HMGB1 expressions in chronic periodontitis patients and their associations with the inflammatory factors. Am. J. Transl. Res. 2021, 13, 9224–9232. [Google Scholar] [PubMed]
- Kang, L.; Miao, Y.; Jin, Y.; Shen, S.; Lin, X. Exosomal miR-205-5p derived from periodontal ligament stem cells attenuates the inflammation of chronic periodontitis via targeting XBP1. Immun. Inflamm. Dis. 2023, 11, e743. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Gandellini, P. Unveiling the ups and downs of miR-205 in physiology and cancer: Transcriptional and post-transcriptional mechanisms. Cell Death Dis. 2020, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, Y.; Huang, W.; Biswas, R.; Banerjee, A.; Broussard, J.A.; Zhao, Z.; Wang, D.; Bjerke, G.; Raghavan, S.; et al. MicroRNA-205 promotes hair regeneration by modulating mechanical properties of hair follicle stem cells. Proc. Natl. Acad. Sci. USA 2023, 120, e2220635120. [Google Scholar] [CrossRef] [PubMed]
- Lock, M.C.; Tellam, R.L.; Darby, J.R.T.; Soo, J.Y.; Brooks, D.A.; Seed, M.; Selvanayagam, J.B.; Morrison, J.L. Identification of Novel miRNAs Involved in Cardiac Repair Following Infarction in Fetal and Adolescent Sheep Hearts. Front. Physiol. 2020, 11, 614. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Su, Y.C.; Chang, M.; Chen, J.A. MicroRNAs mediate precise control of spinal interneuron populations to exert delicate sensory-to-motor outputs. eLife 2021, 10, e63768. [Google Scholar] [CrossRef]
- Agarwal, A.; Kansal, V.; Farooqi, H.; Singh, V.K.; Prasad, R. Differentially deregulated microRNAs contribute to ultraviolet radiation-induced photocarcinogenesis through immunomodulation: An-analysis of microRNAs expression profiling. bioRxiv 2023. [CrossRef] [PubMed]
- Hsieh, C.H.; Yang, J.C.; Jeng, J.C.; Chen, Y.C.; Lu, T.H.; Tzeng, S.L.; Wu, Y.C.; Wu, C.J.; Rau, C.S. Circulating microRNA signatures in mice exposed to lipoteichoic acid. J. Biomed. Sci. 2013, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Al-Rawi, N.H.; Al-Marzooq, F.; Al-Nuaimi, A.S.; Hachim, M.Y.; Hamoudi, R. Salivary microRNA 155, 146a/b and 203: A pilot study for potentially non-invasive diagnostic biomarkers of periodontitis and diabetes mellitus. PLoS ONE 2020, 15, e0237004. [Google Scholar] [CrossRef]
- Moffatt, C.E.; Lamont, R.J. Porphyromonas gingivalis induction of microRNA-203 expression controls suppressor of cytokine signaling 3 in gingival epithelial cells. Infect. Immun. 2011, 79, 2632–2637. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ren, D.; Zhao, D.; Zhang, B.; Ye, R. MicroRNA-203 mediates Porphyromonas gingivalis LPS-induced inflammation and differentiation of periodontal ligament cells. Oral. Dis. 2023, 29, 1715–1725. [Google Scholar] [CrossRef]
- Li, S.; Li, L.; Li, J.; Liang, X.; Song, C.; Zou, Y. miR-203, fine-tunning neuroinflammation by juggling different components of NF-kappaB signaling. J. Neuroinflamm. 2022, 19, 84. [Google Scholar] [CrossRef]
- Friedman, J.M.; Jones, P.A. MicroRNAs: Critical mediators of differentiation, development and disease. Swiss Med. Wkly. 2009, 139, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Liu, H.; Guo, J.; Yu, Y.; Yang, D.; He, F.; Du, Z. MicroRNA-98 negatively regulates myocardial infarction-induced apoptosis by down-regulating Fas and caspase-3. Sci. Rep. 2017, 7, 7460. [Google Scholar] [CrossRef]
- Perri, R.; Nares, S.; Zhang, S.; Barros, S.P.; Offenbacher, S. MicroRNA modulation in obesity and periodontitis. J. Dent. Res. 2012, 91, 33–38. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Cavalcanti, M.; Trubiani, O.; Diomede, F. MicroRNA 210 Mediates VEGF Upregulation in Human Periodontal Ligament Stem Cells Cultured on 3DHydroxyapatite Ceramic Scaffold. Int. J. Mol. Sci. 2018, 19, 3916. [Google Scholar] [CrossRef]
- Ma, H.; Li, M.; Jia, Z.; Chen, X.; Bu, N. miR-876-3p suppresses the progression of colon cancer and correlates the prognosis of patients. Exp. Mol. Pathol. 2021, 122, 104682. [Google Scholar] [CrossRef]
- Law, Y.Y.; Lee, W.F.; Hsu, C.J.; Lin, Y.Y.; Tsai, C.H.; Huang, C.C.; Wu, M.H.; Tang, C.H.; Liu, J.F. miR-let-7c-5p and miR-149-5p inhibit proinflammatory cytokine production in osteoarthritis and rheumatoid arthritis synovial fibroblasts. Aging 2021, 13, 17227–17236. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Romito, A.; Borel, C.; Gehrig, C.; Gagnebin, M.; Falconnet, E.; Izzo, A.; Altucci, L.; Banfi, S.; Antonarakis, S.E.; et al. Cardiomyogenesis is controlled by the miR-99a/let-7c cluster and epigenetic modifications. Stem Cell Res. 2014, 12, 323–337. [Google Scholar] [CrossRef]
- Costantini, E.; Sinjari, B.; Di Giovanni, P.; Aielli, L.; Caputi, S.; Muraro, R.; Murmura, G.; Reale, M. TNFalpha, IL-6, miR-103a-3p, miR-423-5p, miR-23a-3p, miR-15a-5p and miR-223-3p in the crevicular fluid of periodontopathic patients correlate with each other and at different stages of the disease. Sci. Rep. 2023, 13, 126. [Google Scholar] [CrossRef]
- Naqvi, A.R.; Brambila, M.F.; Martinez, G.; Chapa, G.; Nares, S. Dysregulation of human miRNAs and increased prevalence of HHV miRNAs in obese periodontitis subjects. J. Clin. Periodontol. 2019, 46, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Ghiam, S.; Eslahchi, C.; Shahpasand, K.; Habibi-Rezaei, M.; Gharaghani, S. Exploring the role of non-coding RNAs as potential candidate biomarkers in the cross-talk between diabetes mellitus and Alzheimer’s disease. Front. Aging Neurosci. 2022, 14, 955461. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.Q.; Liu, H.; Sun, H.L.; Xiang, J.B.; Wang, X.X.; Jiang, C.X.; Ma, L.; Cao, Z.G. MiR-361-3p/Nfat5 Signaling Axis Controls Cementoblast Differentiation. J. Dent. Res. 2019, 98, 1131–1139. [Google Scholar] [CrossRef]
- Polikepahad, S.; Knight, J.M.; Naghavi, A.O.; Oplt, T.; Creighton, C.J.; Shaw, C.; Benham, A.L.; Kim, J.; Soibam, B.; Harris, R.A.; et al. Proinflammatory role for let-7 microRNAS in experimental asthma. J. Biol. Chem. 2010, 285, 30139–30149. [Google Scholar] [CrossRef]
- Cao, S.; Li, L.; Geng, X.; Ma, Y.; Huang, X.; Kang, X. The upregulation of miR-101 promotes vascular endothelial cell apoptosis and suppresses cell migration in acute coronary syndrome by targeting CDH5. Int. J. Clin. Exp. Pathol. 2019, 12, 3320–3328. [Google Scholar]
- Liu, D.; Tang, H.; Li, X.Y.; Deng, M.F.; Wei, N.; Wang, X.; Zhou, Y.F.; Wang, D.Q.; Fu, P.; Wang, J.Z.; et al. Targeting the HDAC2/HNF-4A/miR-101b/AMPK Pathway Rescues Tauopathy and Dendritic Abnormalities in Alzheimer’s Disease. Mol. Ther. 2017, 25, 752–764. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, H.; Wang, C.; Zhu, Z. Long non-coding RNA Linc01133 promotes osteogenic differentiation of human periodontal ligament stem cells via microRNA-30c / bone gamma-carboxyglutamate protein axis. Bioengineered 2022, 13, 9602–9612. [Google Scholar] [CrossRef]
- Huang, J.; Li, Y.; Zhu, S.; Wang, L.; Yang, L.; He, C. MiR-30 Family: A Novel Avenue for Treating Bone and Joint Diseases? Int. J. Med. Sci. 2023, 20, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Yaman, S.O.; Orem, A.; Yucesan, F.B.; Kural, B.V.; Orem, C. Evaluation of circulating miR-122, miR-30c and miR-33a levels and their association with lipids, lipoproteins in postprandial lipemia. Life Sci. 2021, 264, 118585. [Google Scholar] [CrossRef]
- Du, S.; Shen, S.; Ding, S.; Wang, L. Suppression of microRNA-323-3p restrains vascular endothelial cell apoptosis via promoting sirtuin-1 expression in coronary heart disease. Life Sci. 2021, 270, 119065. [Google Scholar] [CrossRef]
- Huang, T.; Zhao, H.Y.; Zhang, X.B.; Gao, X.L.; Peng, W.P.; Zhou, Y.; Zhao, W.H.; Yang, H.F. LncRNA ANRIL regulates cell proliferation and migration via sponging miR-339-5p and regulating FRS2 expression in atherosclerosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1956–1969. [Google Scholar] [CrossRef]
- Liu, H.L.; Zhu, J.G.; Liu, Y.Q.; Fan, Z.G.; Zhu, C.; Qian, L.M. Identification of the microRNA expression profile in the regenerative neonatal mouse heart by deep sequencing. Cell Biochem. Biophys. 2014, 70, 635–642. [Google Scholar] [CrossRef]
- Saito, A.; Horie, M.; Ejiri, K.; Aoki, A.; Katagiri, S.; Maekawa, S.; Suzuki, S.; Kong, S.; Yamauchi, T.; Yamaguchi, Y.; et al. MicroRNA profiling in gingival crevicular fluid of periodontitis-a pilot study. FEBS Open Bio. 2017, 7, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, H.H.; Chen, Q.; Wang, Y.Y.; Fan, C.C.; Duan, Y.Y.; Huang, Y.; Zhang, H.M.; Li, J.P.; Zhang, X.Y.; et al. miR-142-5p Inhibits Cell Invasion and Migration by Targeting DNMT1 in Breast Cancer. Oncol. Res. 2022, 28, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Zhao, Q.; He, C.; Huang, D.; Liu, J.; Chen, F.; Chen, J.; Liao, J.Y.; Cui, X.; Zeng, Y.; et al. miR-142-5p and miR-130a-3p are regulated by IL-4 and IL-13 and control profibrogenic macrophage program. Nat. Commun. 2015, 6, 8523. [Google Scholar] [CrossRef]
- Guzeldemir-Akcakanat, E.; Sunnetci-Akkoyunlu, D.; Balta-Uysal, V.M.; Ozer, T.; Isik, E.B.; Cine, N. Differentially expressed miRNAs associated with generalized aggressive periodontitis. Clin. Oral. Investig. 2023, 28, 7. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Wen, J.; Wu, Y.; Man, C. MiR-375: A novel multifunctional regulator. Life Sci. 2021, 275, 119323. [Google Scholar] [CrossRef]
- Ren, S.; Tan, X.; Fu, M.Z.; Ren, S.; Wu, X.; Chen, T.; Latham, P.S.; Lin, P.; Man, Y.G.; Fu, S.W. Downregulation of miR-375 contributes to ERBB2-mediated VEGFA overexpression in esophageal cancer. J. Cancer 2021, 12, 7138–7146. [Google Scholar] [CrossRef] [PubMed]
- Piano, M.A.; Gianesello, L.; Grassi, A.; Del Bianco, P.; Mattiolo, A.; Cattelan, A.M.; Sasset, L.; Zanovello, P.; Calabro, M.L. Circulating miRNA-375 as a potential novel biomarker for active Kaposi’s sarcoma in AIDS patients. J. Cell Mol. Med. 2019, 23, 1486–1494. [Google Scholar] [CrossRef]
- Biton, M.; Levin, A.; Slyper, M.; Alkalay, I.; Horwitz, E.; Mor, H.; Kredo-Russo, S.; Avnit-Sagi, T.; Cojocaru, G.; Zreik, F.; et al. Epithelial microRNAs regulate gut mucosal immunity via epithelium-T cell crosstalk. Nat. Immunol. 2011, 12, 239–246. [Google Scholar] [CrossRef]
- Shafiee, M.; Aleyasin, S.A.; Mowla, S.J.; Vasei, M.; Yazdanparast, S.A. The Effect of MicroRNA-375 Overexpression, an Inhibitor of Helicobacter pylori-Induced Carcinogenesis, on lncRNA SOX2OT. Jundishapur J. Microbiol. 2016, 9, e23464. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Anniwaer, A.; Li, B.; Chen, Y.; Yu, Z.; Yu, X. The Role of MicroRNAs in Predicting the Neurological Outcome of Patients with Subarachnoid Hemorrhage: A Meta-analysis. Cell Mol. Neurobiol. 2023, 43, 2883–2893. [Google Scholar] [CrossRef]
- Li, X.; Dai, A.; Tran, R.; Wang, J. Text mining-based identification of promising miRNA biomarkers for diabetes mellitus. Front. Endocrinol. 2023, 14, 1195145. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Chen, H.; Wang, R.; Zhang, J.; Jiang, L.; Zhang, S.; Jiang, C.; Huang, J.; Quan, X.; Liu, Y.; et al. Proposing new early detection indicators for pancreatic cancer: Combining machine learning and neural networks for serum miRNA-based diagnostic model. Front. Oncol. 2023, 13, 1244578. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhong, C. Prediction of miRNA-Disease Association Using Deep Collaborative Filtering. Biomed. Res. Int. 2021, 2021, 6652948. [Google Scholar] [CrossRef]
- Rivera, M.F.; Lee, J.Y.; Aneja, M.; Goswami, V.; Liu, L.; Velsko, I.M.; Chukkapalli, S.S.; Bhattacharyya, I.; Chen, H.; Lucas, A.R.; et al. Polymicrobial infection with major periodontal pathogens induced periodontal disease and aortic atherosclerosis in hyperlipidemic ApoE(null) mice. PLoS ONE 2013, 8, e57178. [Google Scholar] [CrossRef]
- Cioce, M.; Rutigliano, D.; Puglielli, A.; Fazio, V.M. Butein-instigated miR-186-5p-dependent modulation of TWIST1 affects resistance to cisplatin and bioenergetics of Malignant Pleural Mesothelioma cells. Cancer Drug Resist. 2022, 5, 814–828. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Lin, Y.C.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: An informative resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015; 43, W460–W466. [Google Scholar] [CrossRef]
- Cox, D.R. The Regression Analysis of Binary Sequences. J. R. Stat. Soc. Ser. B 1958, 20, 215–242. [Google Scholar] [CrossRef]
- Cortes, C.; Vapnik, V. Support-vector networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Ho, T.K. Random decision forests. In Proceedings of the 3rd International Conference on Document Analysis and Recognition, Montreal, QC, Canada, 14–16 August 1995; pp. 278–282. [Google Scholar]
- Haykin, S. Neural Networks: A Comprehensive Foundation; Prentice Hall PTR: Hoboken, NJ, USA, 1998. [Google Scholar]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, New York, NY, USA, 13–17 August 2016; pp. 785–794. [Google Scholar]
- Gillies S, o. Shapely: Manipulation and analysis of geometric objects. 2007. Available online: https://github.com/Toblerity/Shapely (accessed on 29 April 2024).
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Global TLR2 and 4 deficiency in mice impacts bone resorption, inflammatory markers and atherosclerosis to polymicrobial infection. Mol. Oral. Microbiol. 2017, 32, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Siow, M.Y.; Ng, L.P.; Vincent-Chong, V.K.; Jamaludin, M.; Abraham, M.T.; Abdul Rahman, Z.A.; Kallarakkal, T.G.; Yang, Y.H.; Cheong, S.C.; Zain, R.B. Dysregulation of miR-31 and miR-375 expression is associated with clinical outcomes in oral carcinoma. Oral Dis. 2014, 20, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Mitani, Y.; Roberts, D.B.; Fatani, H.; Weber, R.S.; Kies, M.S.; Lippman, S.M.; El-Naggar, A.K. MicroRNA profiling of salivary adenoid cystic carcinoma: Association of miR-17-92 upregulation with poor outcome. PLoS ONE 2013, 8, e66778. [Google Scholar] [CrossRef]
- Kawasaki, H.; Amano, H. Anti-inflammatory role of microRNA-429 in human gingival epithelial cells-inhibition of IL-8 production through direct binding to IKKbeta mRNA. Mol. Med. Rep. 2021, 24, 12220. [Google Scholar] [CrossRef]
- Zhang, L.; Yan, H.; Wang, H.; Wang, L.; Bai, B.; Ma, Y.; Tie, Y.; Xi, Z. MicroRNA (miR)-429 Promotes Inflammatory Injury by Targeting Kruppel-like Factor 4 (KLF4) in Neonatal Pneumonia. Curr. Neurovasc. Res. 2020, 17, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Cao, H.; Zhuang, J.; Wan, J.; Guan, M.; Yu, B.; Li, X.; Zhang, W. Identification of miR-130a, miR-27b and miR-210 as serum biomarkers for atherosclerosis obliterans. Clin. Chim. Acta 2011, 412, 66–70. [Google Scholar] [CrossRef]
- Krongbaramee, T.; Zhu, M.; Qian, Q.; Zhang, Z.; Eliason, S.; Shu, Y.; Qian, F.; Akkouch, A.; Su, D.; Amendt, B.A.; et al. Plasmid encoding microRNA-200c ameliorates periodontitis and systemic inflammation in obese mice. Mol. Ther. Nucleic Acids 2021, 23, 1204–1216. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Qu, M.; Li, Y.; Wang, L.; Zhang, L.; Wang, Y.; Tang, Y.; Tian, H.L.; Zhang, Z.; Yang, G.Y. MicroRNA-126-3p/-5p Overexpression Attenuates Blood-Brain Barrier Disruption in a Mouse Model of Middle Cerebral Artery Occlusion. Stroke 2020, 51, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Bostjancic, E.; Zidar, N.; Glavac, D. MicroRNA microarray expression profiling in human myocardial infarction. Dis. Markers 2009, 27, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Zhang, L.; Cheng, T.; Wang, J.; Zhou, J.; Zhao, L.; Hua, C.; Xia, Q. MiR-590-5p inhibits pathological hypertrophy mediated heart failure by targeting RTN4. J. Mol. Histol. 2021, 52, 955–964. [Google Scholar] [CrossRef]
- Lesizza, P.; Prosdocimo, G.; Martinelli, V.; Sinagra, G.; Zacchigna, S.; Giacca, M. Single-Dose Intracardiac Injection of Pro-Regenerative MicroRNAs Improves Cardiac Function After Myocardial Infarction. Circ. Res. 2017, 120, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Purohit, P.; Roy, D.; Dwivedi, S.; Nebhinani, N.; Sharma, P. Association of miR-155, miR-187 and Inflammatory Cytokines IL-6, IL-10 and TNF-alpha in Chronic Opium Abusers. Inflammation 2022, 45, 554–566. [Google Scholar] [CrossRef]
- Deng, J.; Xiao, J.; Ma, P.; Gao, B.; Gong, F.; Lv, L.; Zhang, Y.; Xu, J. Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection. Viruses 2017, 9, 118. [Google Scholar] [CrossRef]
- Xu, F.; Yao, F.; Ning, Y. MicroRNA-202-5p-dependent inhibition of Bcl-2 contributes to macrophage apoptosis and atherosclerotic plaque formation. Gene 2023, 867, 147366. [Google Scholar] [CrossRef]
- Yu, G.; Sun, W.; Wang, W.; Le, C.; Liang, D.; Shuai, L. Overexpression of microRNA-202-3p in bone marrow mesenchymal stem cells improves cerebral ischemia-reperfusion injury by promoting angiogenesis and inhibiting inflammation. Aging 2021, 13, 11877–11888. [Google Scholar] [CrossRef] [PubMed]
- Benakanakere, M.R.; Li, Q.; Eskan, M.A.; Singh, A.V.; Zhao, J.; Galicia, J.C.; Stathopoulou, P.; Knudsen, T.B.; Kinane, D.F. Modulation of TLR2 protein expression by miR-105 in human oral keratinocytes. J. Biol. Chem. 2009, 284, 23107–23115. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Choi, J.W.; Moon, H.; Lee, C.Y.; Park, J.H.; Lee, J.; Seo, H.H.; Han, G.; Lim, S.; Lee, S.; et al. Simultaneous Suppression of Multiple Programmed Cell Death Pathways by miRNA-105 in Cardiac Ischemic Injury. Mol. Ther. Nucleic Acids 2019, 14, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Ongoz Dede, F.; Gokmenoglu, C.; Turkmen, E.; Bozkurt Dogan, S.; Ayhan, B.S.; Yildirim, K. Six miRNA expressions in the saliva of smokers and non-smokers with periodontal disease. J. Periodontal Res. 2023, 58, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, E.; Tuddenham, L.; Macquin, C.; Alsaleh, G.; Schreiber-Becker, J.; Tanguy, M.; Bahram, S.; Pfeffer, S.; Georgel, P. Deregulation of type I IFN-dependent genes correlates with increased susceptibility to cytomegalovirus acute infection of dicer mutant mice. PLoS ONE 2012, 7, e43744. [Google Scholar] [CrossRef] [PubMed]
- Kaid, C.; Jordan, D.; Bueno, H.M.S.; Araujo, B.H.S.; Assoni, A.; Okamoto, O.K. miR-367 as a therapeutic target in stem-like cells from embryonal central nervous system tumors. Mol. Oncol. 2019, 13, 2574–2587. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, Y.; Wang, T.; Zhou, N.; Kong, J.; Chen, L.; Snitow, M.; Morley, M.; Li, D.; Petrenko, N.; et al. A microRNA-Hippo pathway that promotes cardiomyocyte proliferation and cardiac regeneration in mice. Sci. Transl. Med. 2015, 7, 279ra238. [Google Scholar] [CrossRef]
- Yuan, Y.S.; Fei, M.; Yang, Y.X.; Cai, W.W. MiR-201-5p alleviates lipopolysaccharide-induced renal cell dysfunction by targeting NOTCH3. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5592–5603. [Google Scholar] [CrossRef]
- Moghaddam, A.S.; Afshari, J.T.; Esmaeili, S.A.; Saburi, E.; Joneidi, Z.; Momtazi-Borojeni, A.A. Cardioprotective microRNAs: Lessons from stem cell-derived exosomal microRNAs to treat cardiovascular disease. Atherosclerosis 2019, 285, 1–9. [Google Scholar] [CrossRef]
- Ramirez, C.M.; Goedeke, L.; Rotllan, N.; Yoon, J.H.; Cirera-Salinas, D.; Mattison, J.A.; Suarez, Y.; de Cabo, R.; Gorospe, M.; Fernandez-Hernando, C. MicroRNA 33 regulates glucose metabolism. Mol. Cell. Biol. 2013, 33, 2891–2902. [Google Scholar] [CrossRef]
- Horie, T.; Nakao, T.; Miyasaka, Y.; Nishino, T.; Matsumura, S.; Nakazeki, F.; Ide, Y.; Kimura, M.; Tsuji, S.; Rodriguez, R.R.; et al. microRNA-33 maintains adaptive thermogenesis via enhanced sympathetic nerve activity. Nat. Commun. 2021, 12, 843. [Google Scholar] [CrossRef]
- Price, N.L.; Goedeke, L.; Suarez, Y.; Fernandez-Hernando, C. miR-33 in cardiometabolic diseases: Lessons learned from novel animal models and approaches. EMBO Mol. Med. 2021, 13, e12606. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Zhang, P.; Wen, L.; Jia, S.; Wu, Y.; Zhang, Z.; Guan, L.; Yu, Z.; Zhao, L. miR-22 promotes stem cell traits via activating Wnt/beta-catenin signaling in cutaneous squamous cell carcinoma. Oncogene 2021, 40, 5799–5813. [Google Scholar] [CrossRef]
- Lv, L.; Zheng, N.; Zhang, L.; Li, R.; Li, Y.; Yang, R.; Li, C.; Fang, R.; Shabanova, A.; Li, X.; et al. Metformin ameliorates cardiac conduction delay by regulating microRNA-1 in mice. Eur. J. Pharmacol. 2020, 881, 173131. [Google Scholar] [CrossRef]
- Grabmaier, U.; Clauss, S.; Gross, L.; Klier, I.; Franz, W.M.; Steinbeck, G.; Wakili, R.; Theiss, H.D.; Brenner, C. Diagnostic and prognostic value of miR-1 and miR-29b on adverse ventricular remodeling after acute myocardial infarction—The SITAGRAMI-miR analysis. Int. J. Cardiol. 2017, 244, 30–36. [Google Scholar] [CrossRef]
- Safa, A.; Bahroudi, Z.; Shoorei, H.; Majidpoor, J.; Abak, A.; Taheri, M.; Ghafouri-Fard, S. miR-1: A comprehensive review of its role in normal development and diverse disorders. Biomed. Pharmacother. 2020, 132, 110903. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Jiang, P.; Wang, H.; Huang, L.; Wu, G.; Che, X.; Wang, C.; Li, P.; Duan, Q.; Guo, X.; et al. MiR-1 is a critical regulator of chondrocyte proliferation and hypertrophy by inhibiting Indian hedgehog pathway during postnatal endochondral ossification in miR-1 overexpression transgenic mice. Bone 2022, 165, 116566. [Google Scholar] [CrossRef]
- Gu, H.; Shi, S.; Xiao, F.; Huang, Z.; Xu, J.; Chen, G.; Zhou, K.; Lu, L.; Yin, X. MiR-1-3p regulates the differentiation of mesenchymal stem cells to prevent osteoporosis by targeting secreted frizzled-related protein 1. Bone 2020, 137, 115444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.G.; Zhao, X.G.; Wang, X.L.; Liu, M.X.; Wan, W. Low expression of miR-1 promotes osteogenic repair of bone marrow mesenchymal stem cells by targeting TLR1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3492–3500. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, K.; Zhi, T.; Xu, X. miR-720 is a key regulator of glioma migration and invasion by controlling TARSL2 expression. Hum. Cell 2021, 34, 1504–1516. [Google Scholar] [CrossRef]
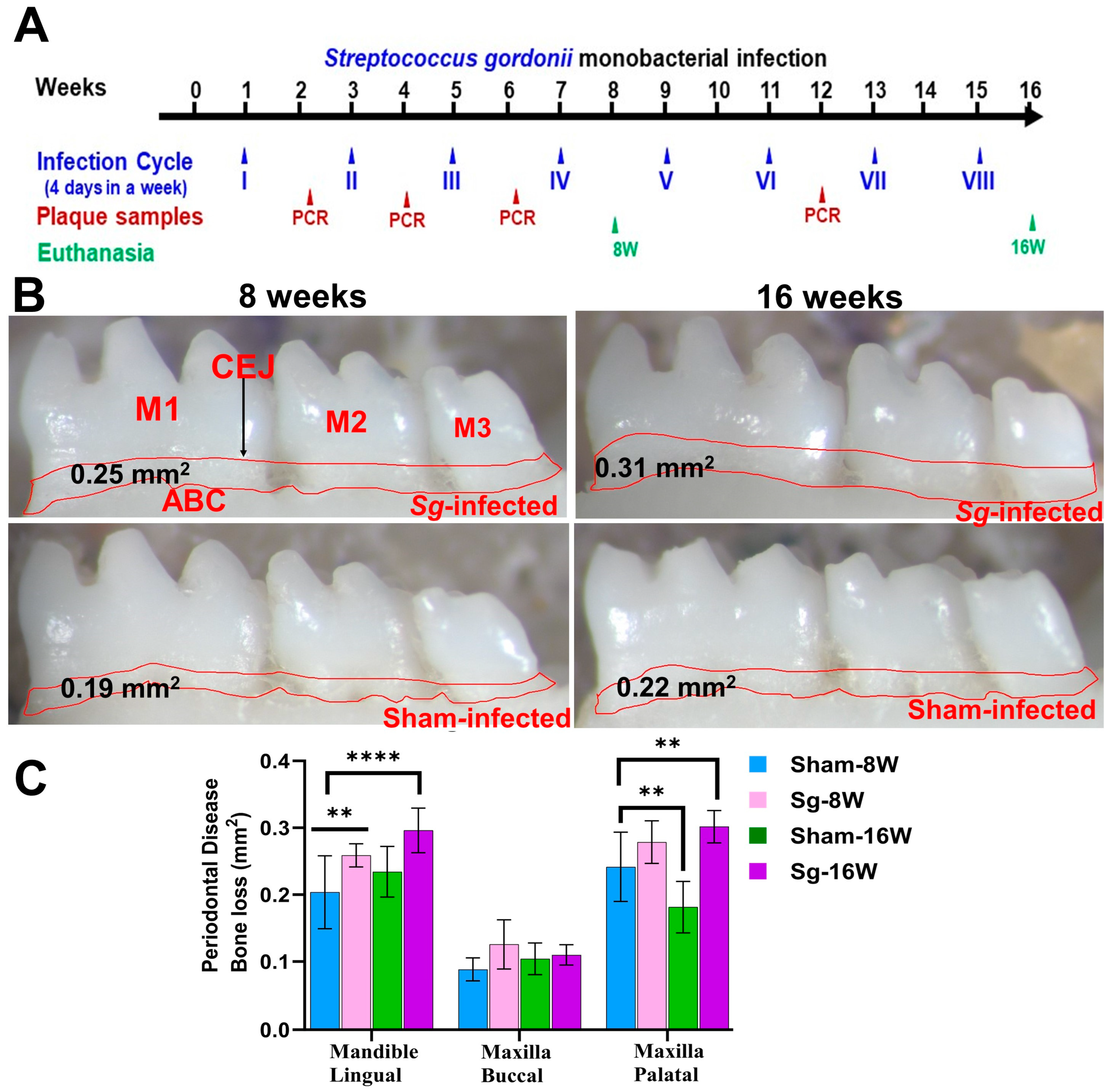
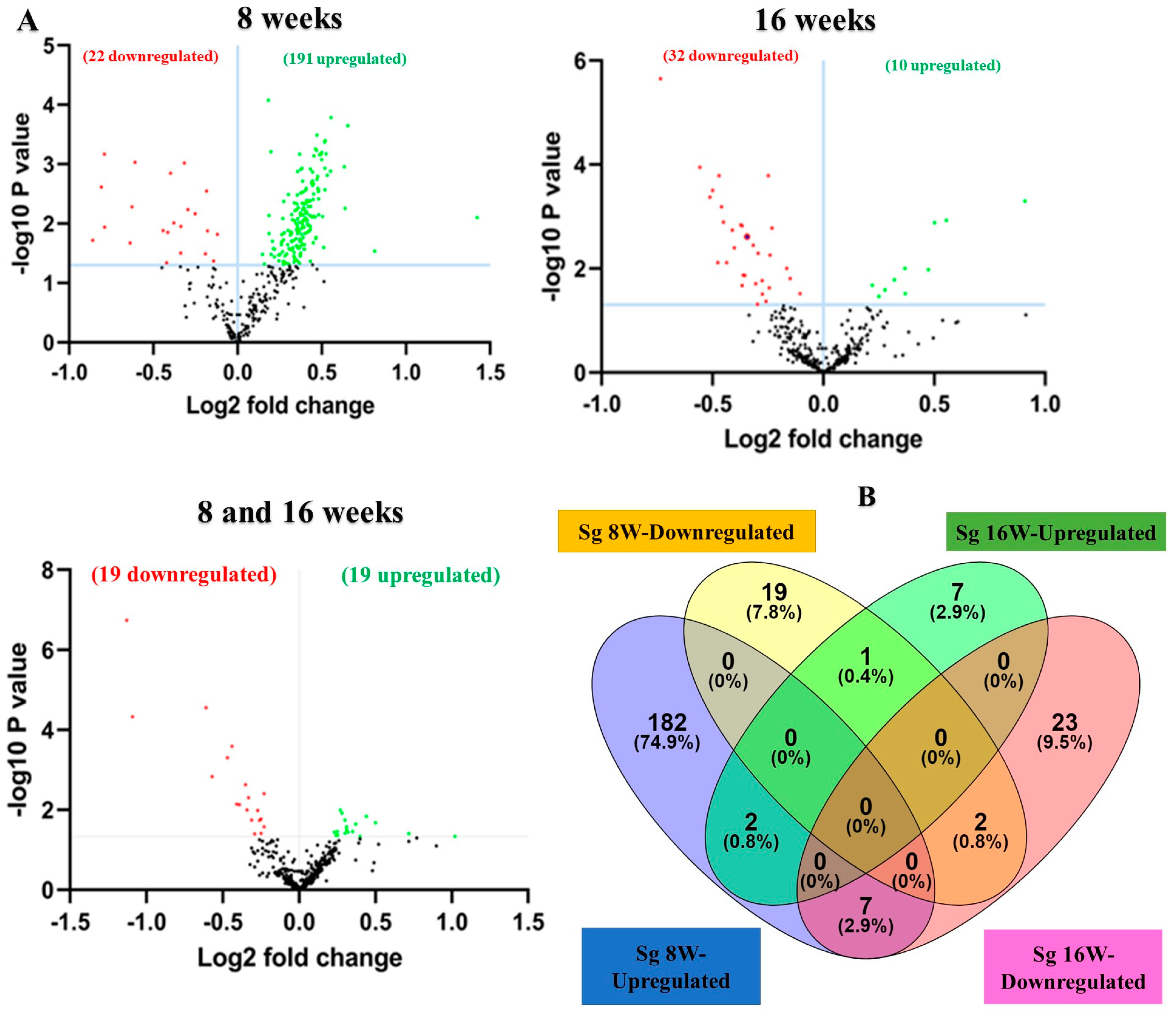
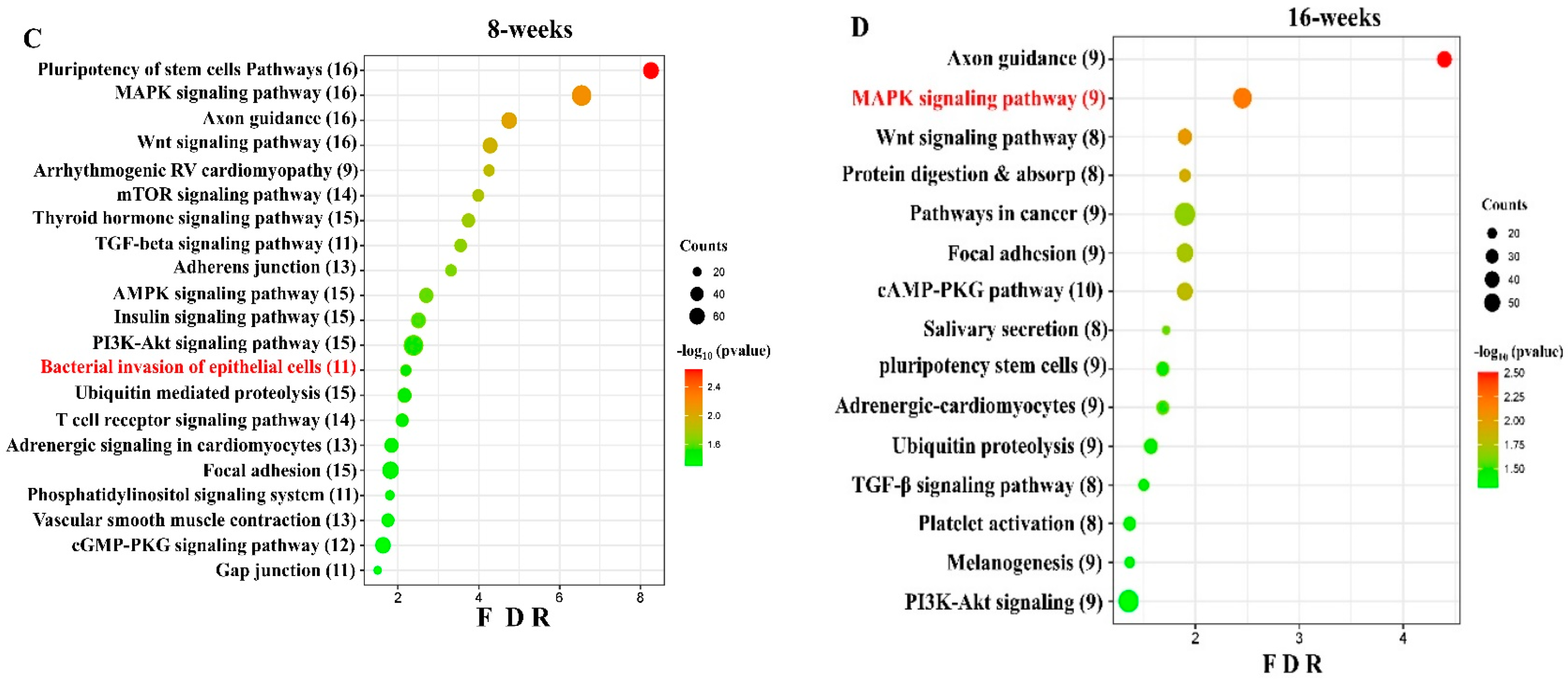
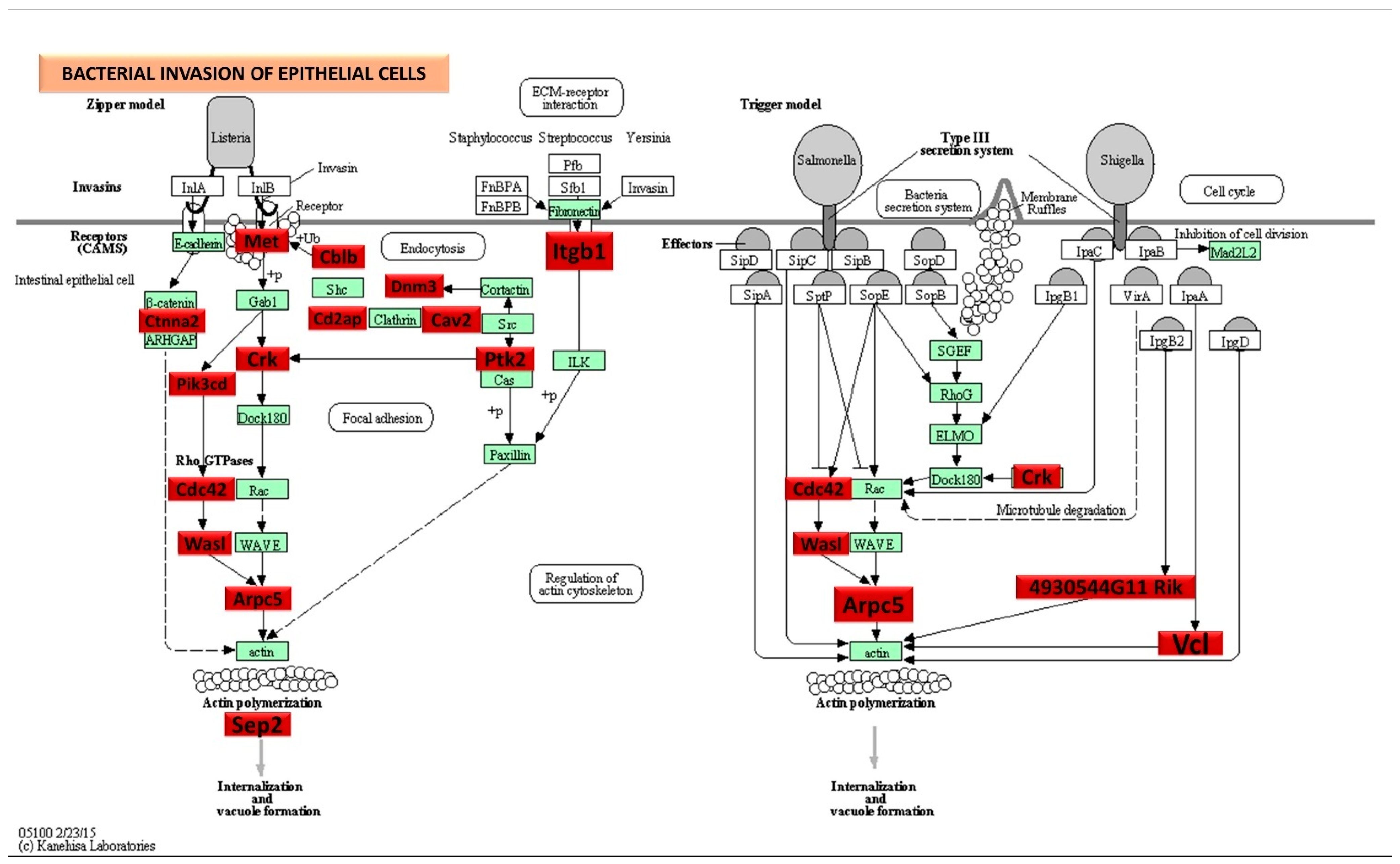
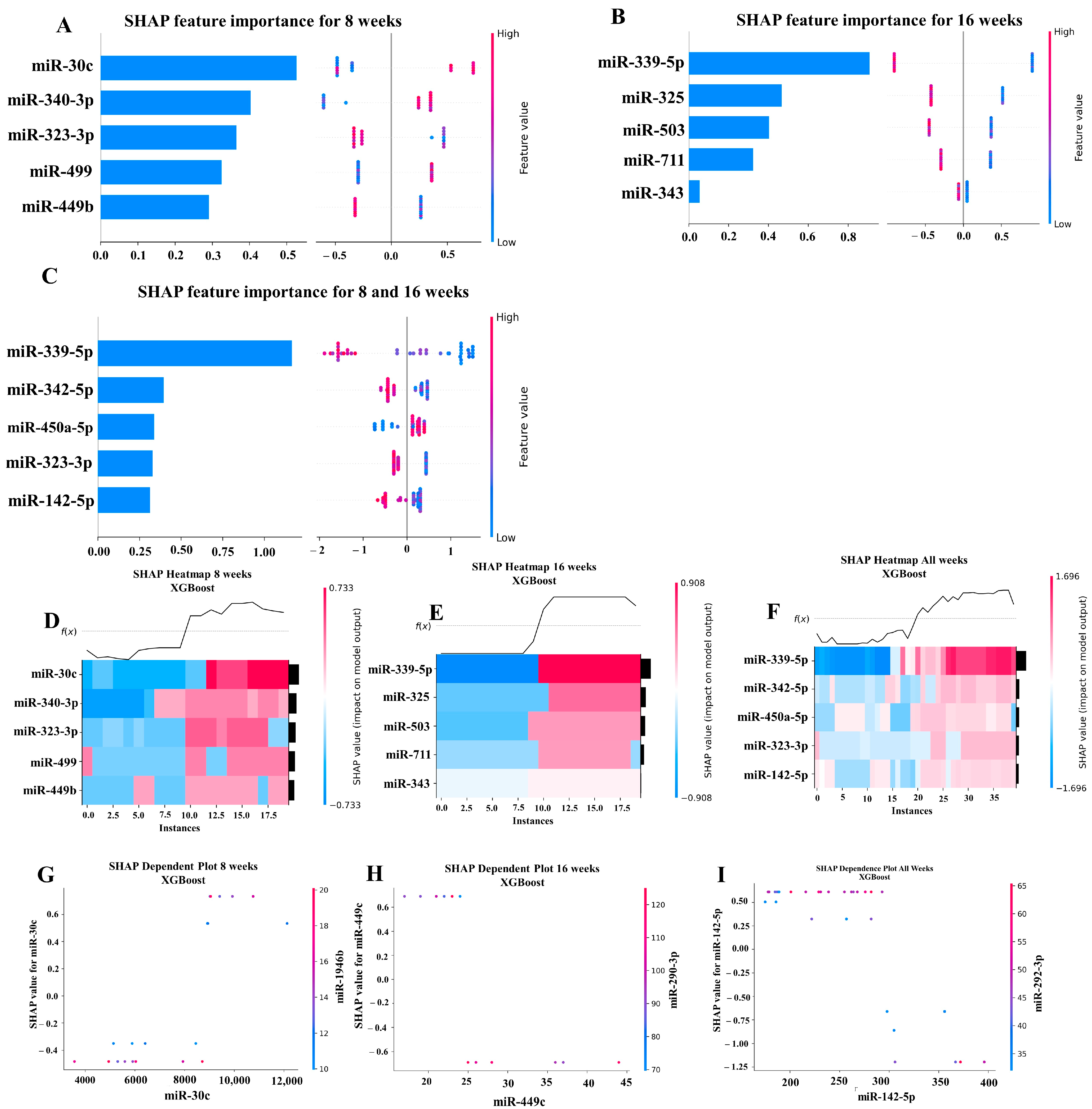
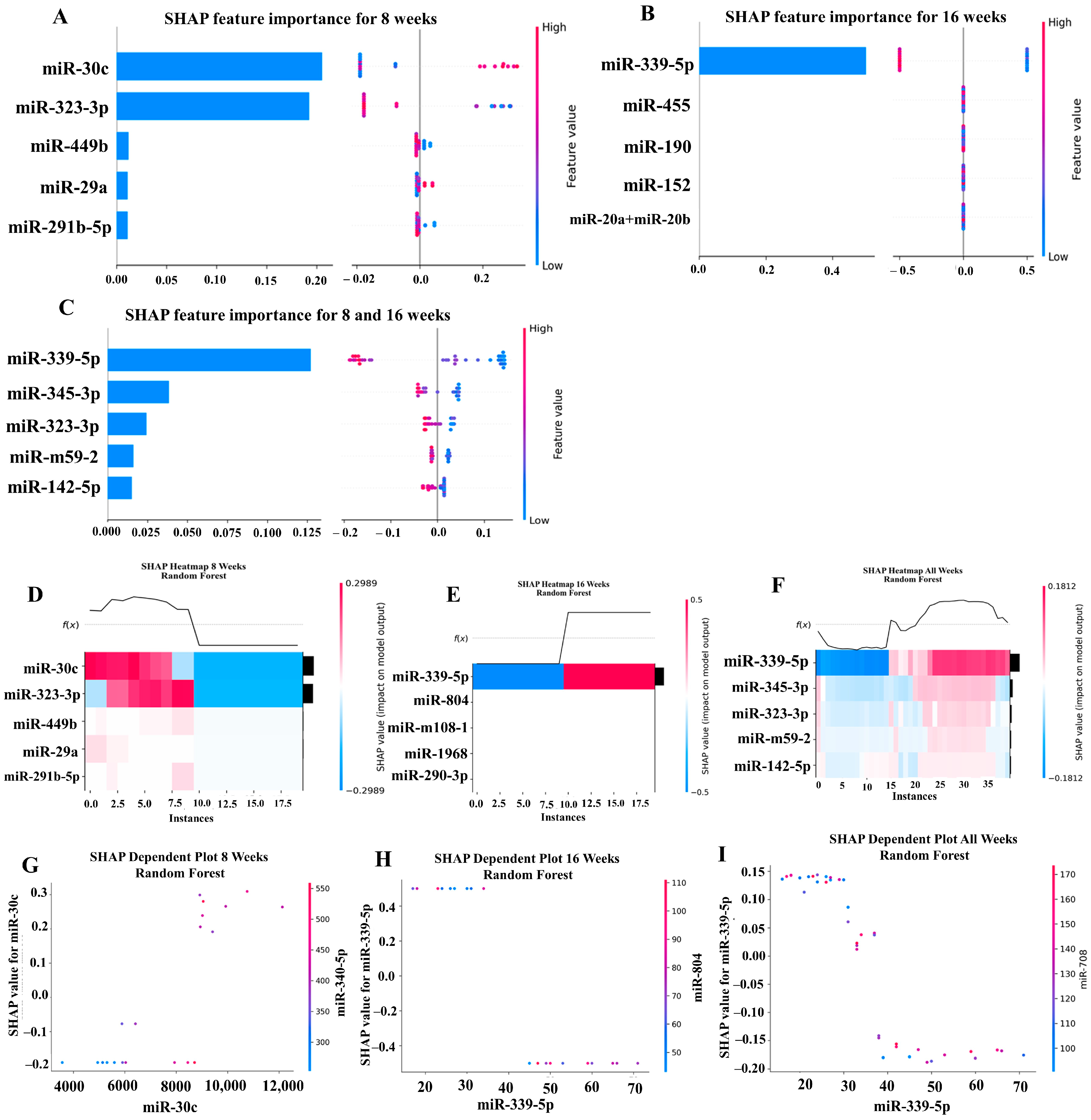
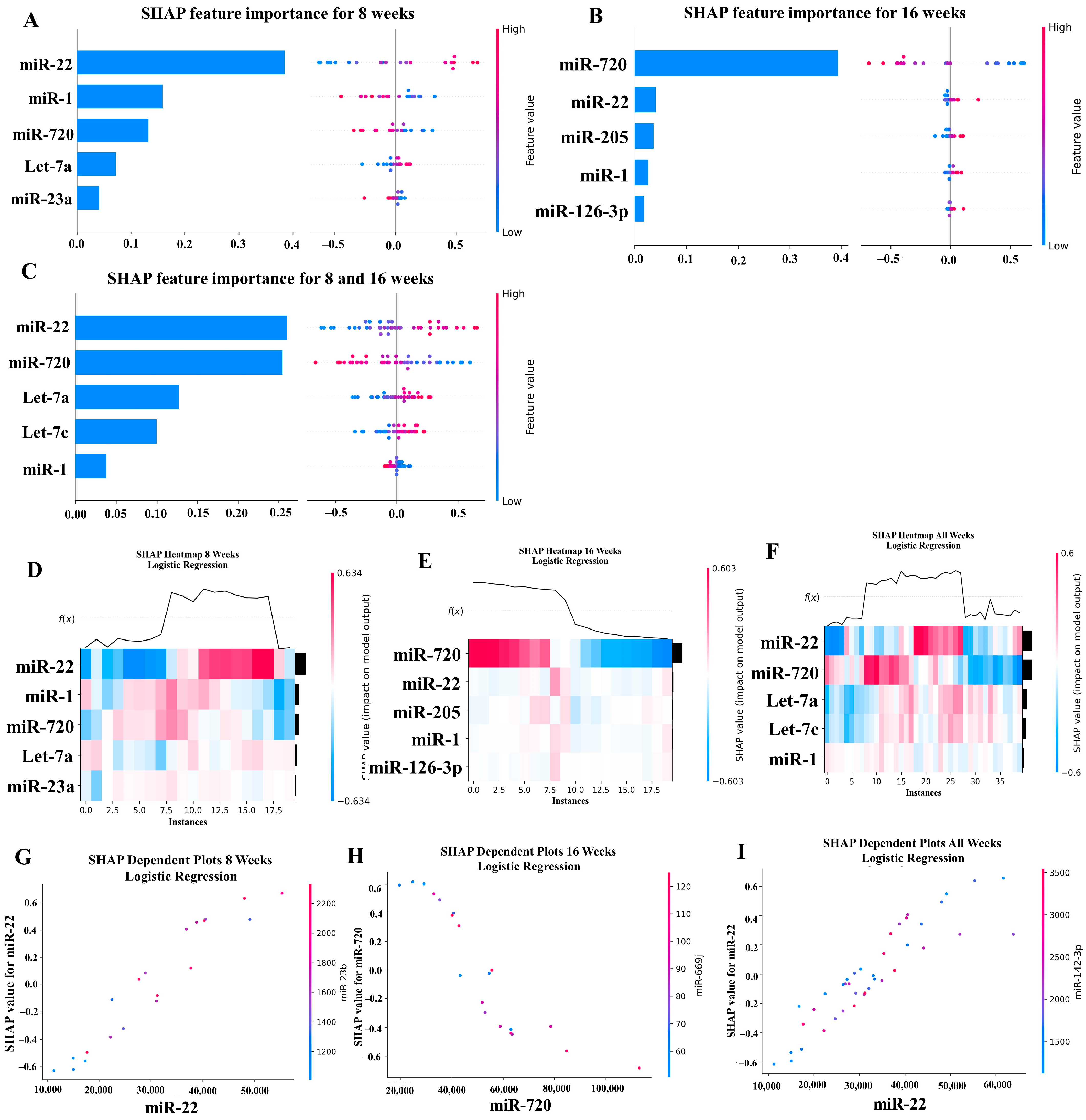
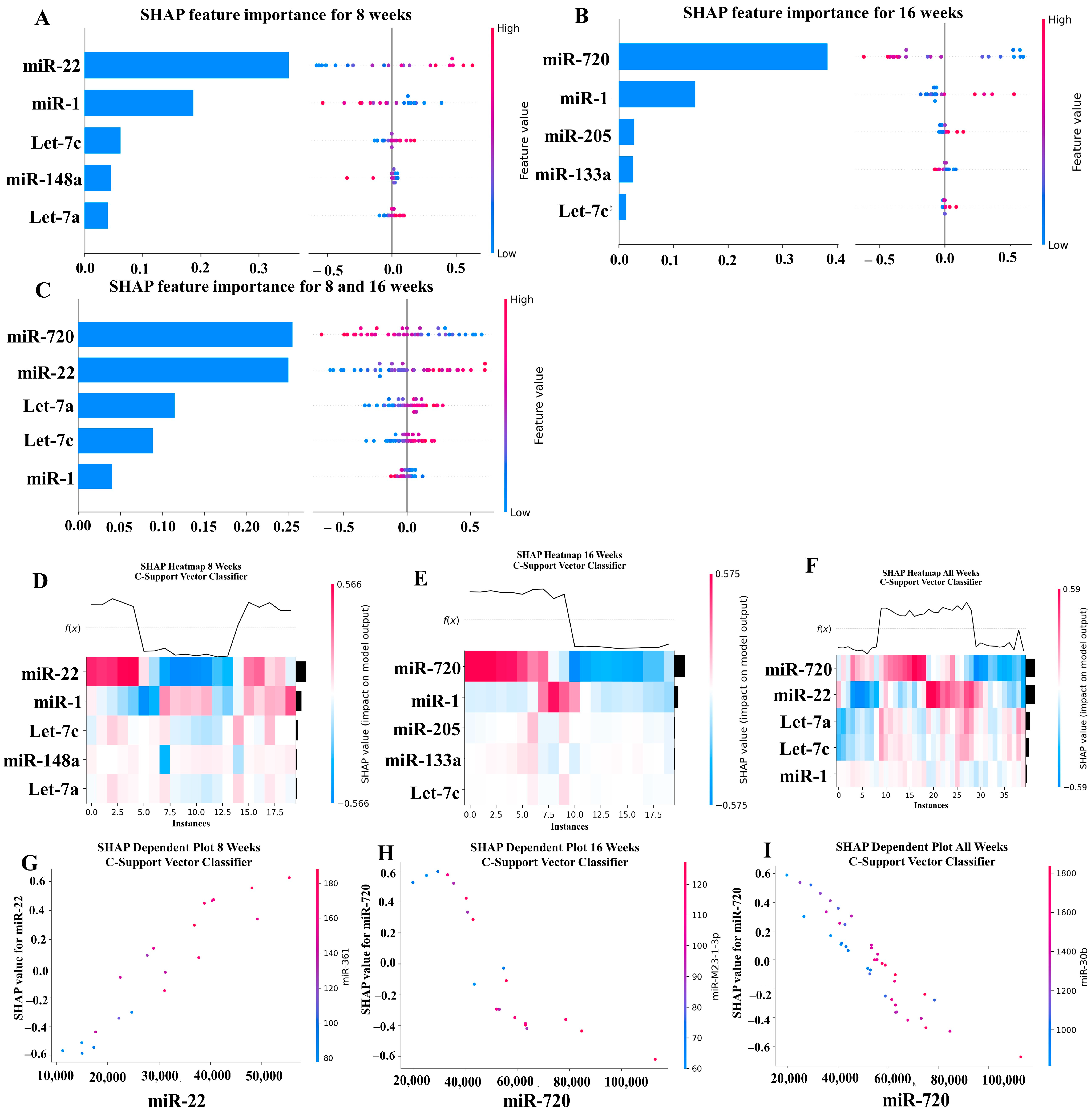
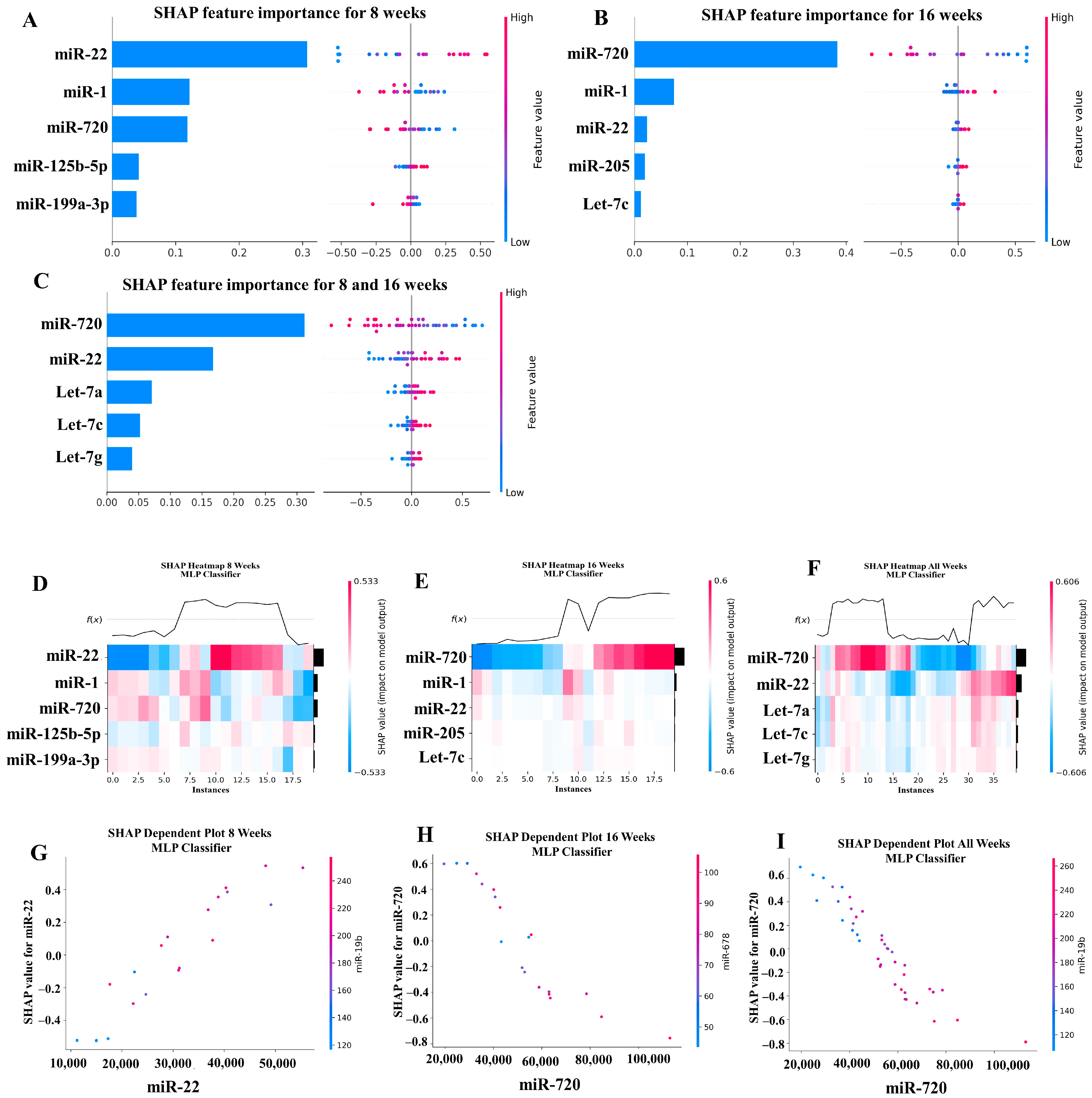
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).