
ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Expression Profile of ZmNF-YA1 in Maize Heat Stress Response
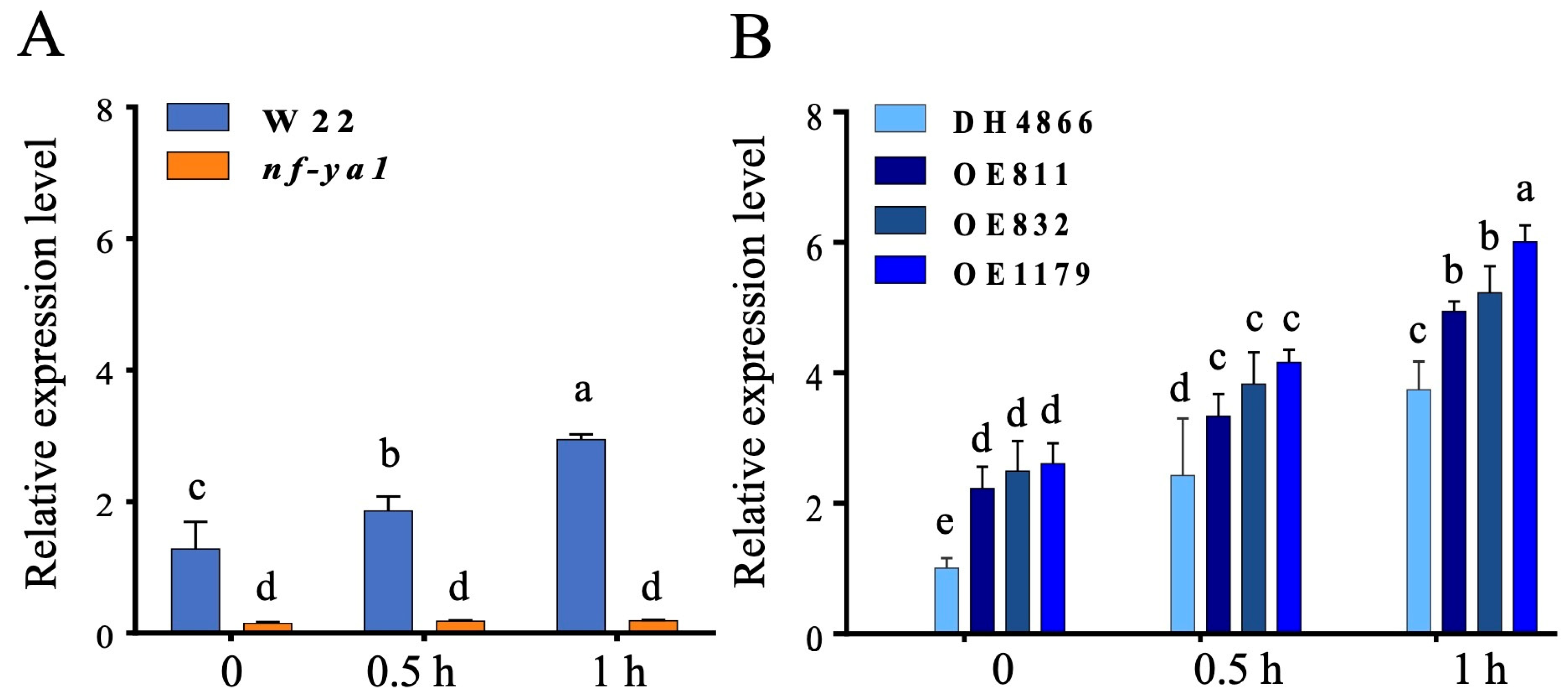
2.2. Expression of ZmNF-YA1 in the Mutant and Overexpression Lines
2.3. ZmNF-YA1 Functions as a Positive Regulator in Maize Thermotolerance
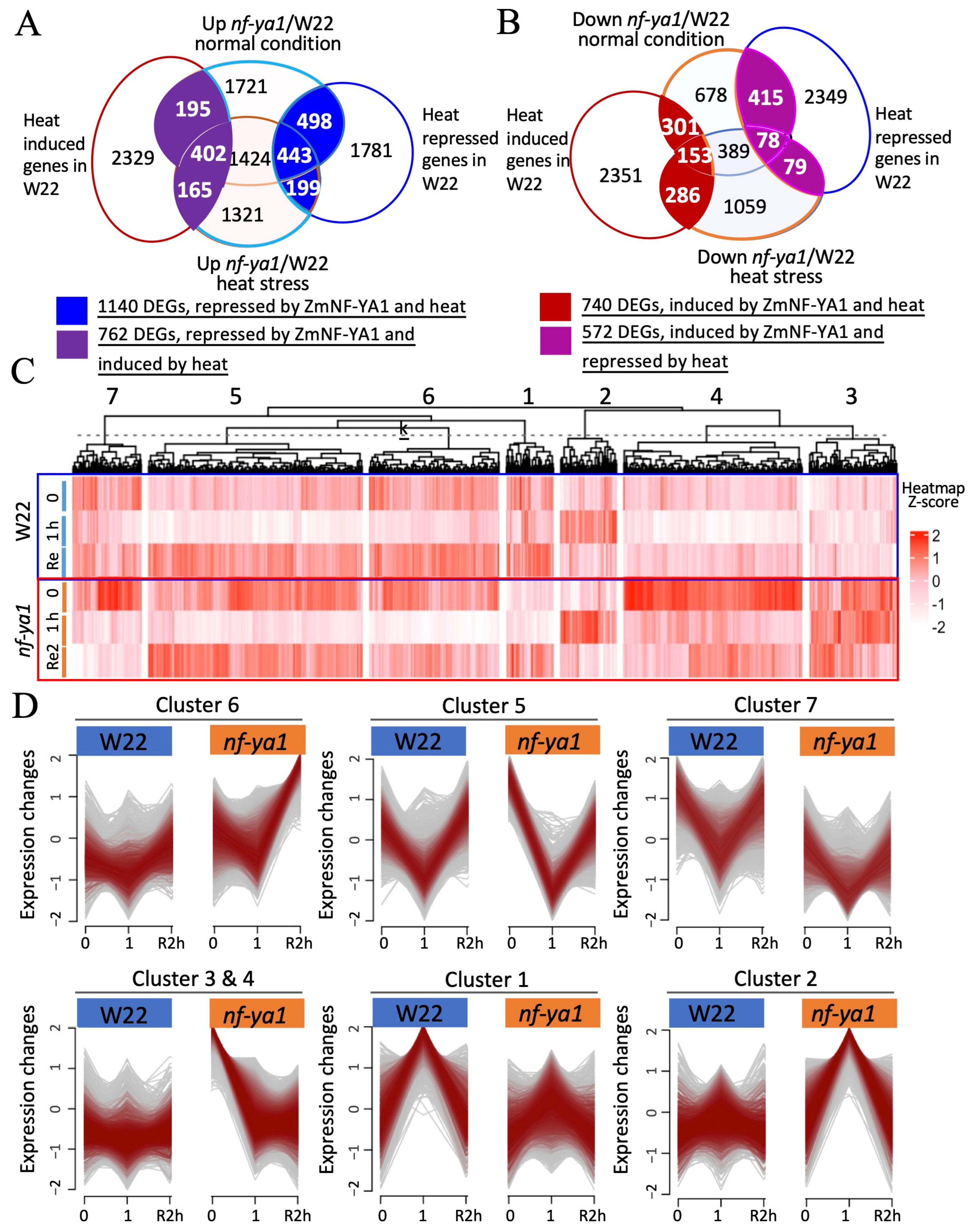
2.4. Comparative Transcriptome Analysis of Maize nf-ya1 and W22 in Heat Stress Response
2.5. ZmNF-YA1 Contributes to the Transcriptional Regulation in Maize Heat Stress Response
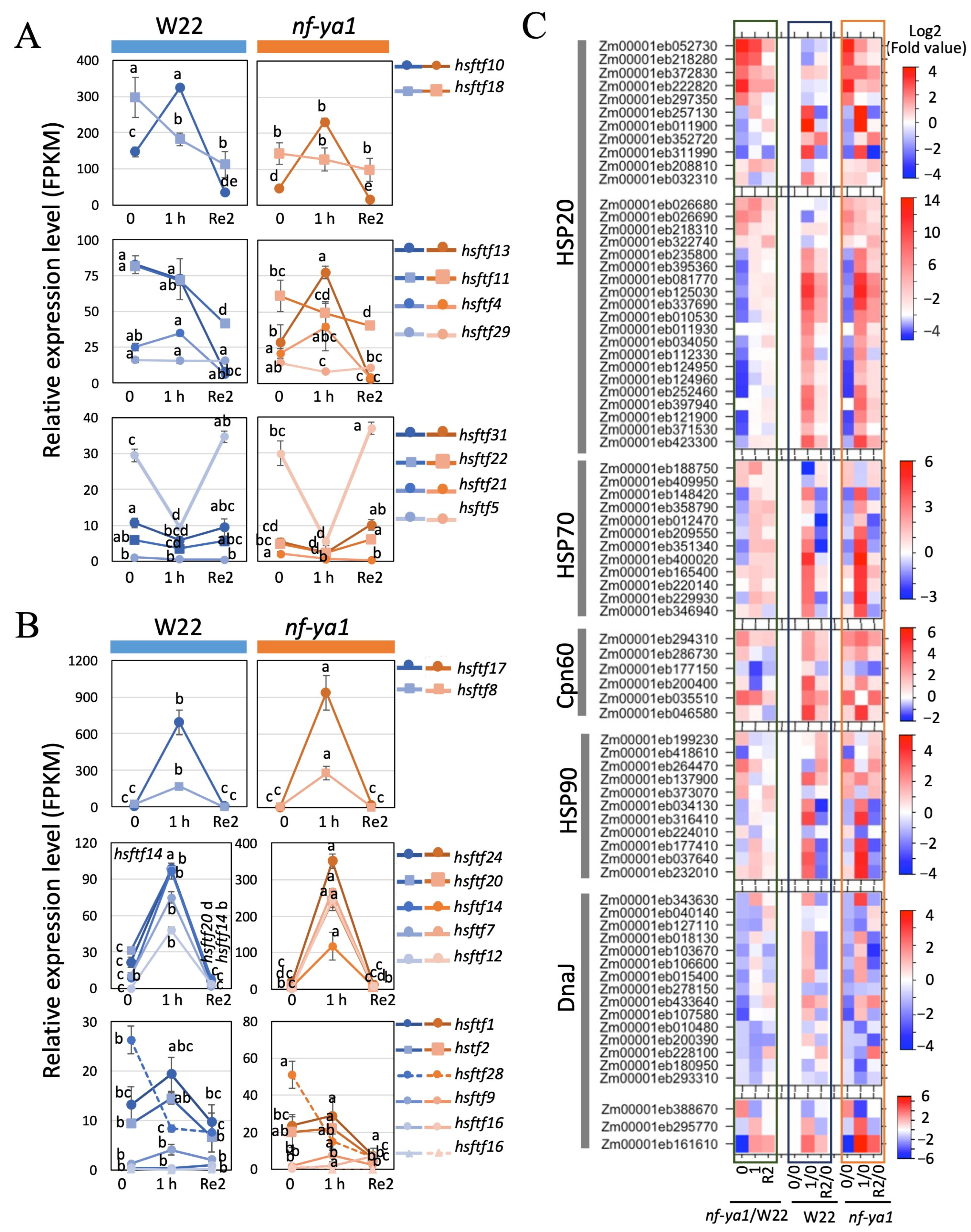
2.6. Effect of ZmNF-YA1 on Maize’s Heat Stress Response
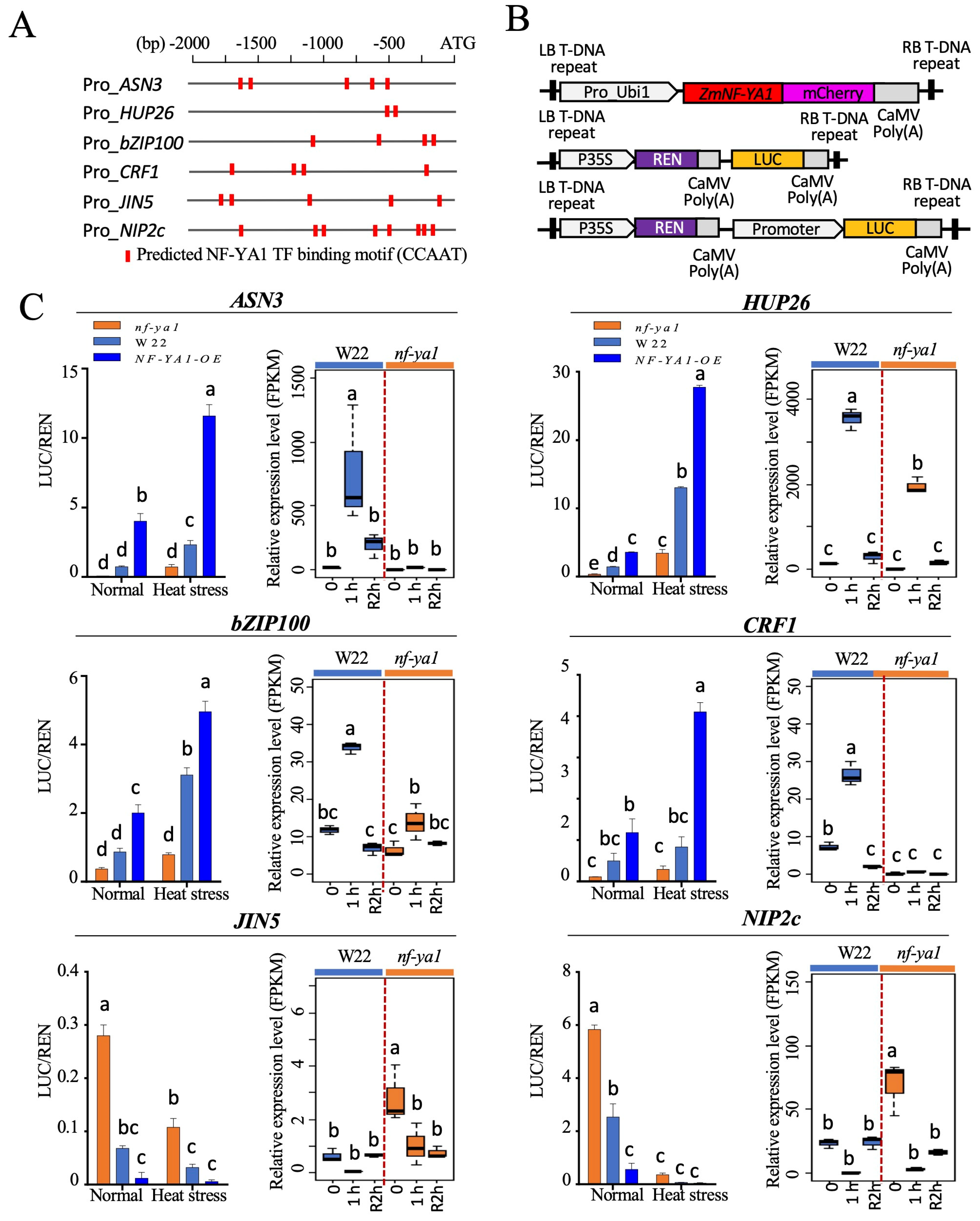
2.7. Validation of the NF-YA1 Function in the Expression of Candidate Target Genes
3. Discussion
3.1. ZmNF-YA1 Is a Genetic Manipulation Target for Improving Thermotolerance and Drought Tolerance
3.2. Failure to Launch Full-Fledged Heat Stress Response in nf-ya1 in Response to Heat Stress
3.3. ZmNF-YA1 Promotes Protein Refolding and Stability When Subjected to Heat Stress Treatment
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Growth and Stress Treatment
4.3. RNA Extraction, qRT-PCR Analysis, and RNA-seq Analysis
4.4. Chlorophyll and MDA Content Analysis, as Well as Cell Staining
4.5. Sequence Analysis
4.6. Dual Luciferase Reporter Assay
4.7. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prasad, P.V.V.; Bheemanahalli, R.; Jagadish, S.V.K. Field Crops and the Fear of Heat Stress—Opportunities, Challenges and Future Directions. Field Crops Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature Increase Reduces Global Yields of Major Crops in Four Independent Estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Schoper, J.B.; Lambert, R.J.; Vasilas, B.L.; Westgate, M.E. Plant Factors Controlling Seed Set in Maize. Plant Physiol. 1987, 83, 121–125. [Google Scholar] [CrossRef]
- Wilhelm, E.P.; Mullen, R.E.; Keeling, P.L.; Singletary, G.W. Heat Stress during Grain Filling in Maize: Effects on Kernel Growth and Metabolism. Crop Sci. 1999, 39, 1733–1741. [Google Scholar] [CrossRef]
- Lizaso, J.I.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.A.; Lorite, I.J.; Sánchez, D.; García, E.; Rodríguez, A. Impact of High Temperatures in Maize: Phenology and Yield Components. Field Crops Res. 2018, 216, 129–140. [Google Scholar] [CrossRef]
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (Eds.)]; Arias, P., Bustamante, M., Elgizouli, I., Flato, G., Howden, M., Méndez-Vallejo, C., Pereira, J.J., Pichs-Madruga, R., Rose, S.K., Saheb, Y., et al., Eds.; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Dauphin, L. NASA Announces Summer 2023 Hottest on Record; NASA: Washington, DC, USA, 2023.
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Huang, S.S.C.; Wise, A.; Castanoz, R.; Nery, J.R.; Chen, H.; Watanabe, M.; Thomas, J.; Bar-Joseph, Z.; Ecker, J.R. A Transcription Factor Hierarchy Defines an Environmental Stress Response Network. Science (1979) 2016, 354, aag1550. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative Response of Maize and Rice Genotypes to Heat Stress: Status of Oxidative Stress and Antioxidants. Acta Physiol. Plant 2012, 34, 75–86. [Google Scholar] [CrossRef]
- Sinha, S.; Maity, S.N.; de Crombrugghe, B. Recombinant Rat CBF-C, the Third Subunit of CBFNFY, Allows Formation of a 622 Protein-DNA Complex with CBF-A and CBF-B and with yeast HAP2 and HAP3. Proc. Natl. Acad. Sci. USA 1995, 28, 1624–1632. [Google Scholar] [CrossRef]
- Petroni, K.; Kumimoto, R.W.; Gnesutta, N.; Calvenzani, V.; Fornari, M.; Tonelli, C.; Holt, B.F.; Mantovani, R. The Promiscuous Life of Plant NUCLEAR FACTOR Y Transcription Factors. Plant Cell 2013, 24, 4777–4792. [Google Scholar] [CrossRef]
- Myers, Z.A.; Holt, B.F. NUCLEAR FACTOR-Y: Still Complex after All These Years? Curr. Opin. Plant Biol. 2018, 45, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; De Mita, S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT-Box Binding Transcription Factors in Plants: Y so Many? Trends Plant Sci. 2013, 18, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Gnesutta, N.; Donati, G.; Gatta, R.; Forni, C.; Fossati, A.; Vonrhein, C.; Moras, D.; Romier, C.; Bolognesi, M.; et al. Sequence-Specific Transcription Factor NF-Y Displays Histone-like DNA Binding and H2B-like Ubiquitination. Cell 2013, 152, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Gnesutta, N.; Kumimoto, R.W.; Swain, S.; Chiara, M.; Siriwardana, C.; Horner, D.S.; Holt, B.F.; Mantovani, R. CONSTANS Imparts DNA Sequence Specificity to the Histone Fold NF-YB/NF-YC Dimer. Plant Cell 2017, 29, 1516–1532. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Ma, C.; Ye, F.; Pang, Y.; Wang, G.; Fahim, A.M.; Lu, X. Genome-Wide Identification of NF-Y Gene Family in Maize (Zea mays L.) and the Positive Role of ZmNF-YC12 in Drought Resistance and Recovery Ability. Front. Plant Sci. 2023, 14, 1159955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.; Zhang, C.; Zou, H.; Wu, Z. Isolation, Structural Analysis, and Expression Characteristics of the Maize Nuclear Factor Y Gene Families. Biochem. Biophys. Res. Commun. 2016, 478, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Ballif, J.; Endo, S.; Kotani, M.; MacAdam, J.; Wu, Y. Over-Expression of HAP3b Enhances Primary Root Elongation in Arabidopsis. Plant Physiol. Biochem. 2011, 49, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Ben-Naim, O.; Eshed, R.; Parnis, A.; Teper-Bamnolker, P.; Shalit, A.; Coupland, G.; Samach, A.; Lifschitz, E. The CCAAT Binding Factor Can Mediate Interactions between CONSTANS-like Proteins and DNA. Plant J. 2006, 46, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Ballif, J.; Endo, S.; Davis, E.; Liang, M.; Chen, D.; Dewald, D.; Kreps, J.; Zhu, T.; Wu, Y. A Putative CCAAT-Binding Transcription Factor Is a Regulator of Flowering Timing in Arabidopsis. Plant Physiol. 2007, 145, 98–105. [Google Scholar] [CrossRef]
- Fornari, M.; Calvenzani, V.; Masiero, S.; Tonelli, C.; Petroni, K. The Arabidopsis NF-YA3 and NF-YA8 Genes Are Functionally Redundant and Are Required in Early Embryogenesis. PLoS ONE 2013, 8, e82043. [Google Scholar] [CrossRef]
- Mu, J.; Tan, H.; Hong, S.; Liang, Y.; Zuo, J. Arabidopsis Transcription Factor Genes NF-YA1, 5, 6, and 9 Play Redundant Roles in Male Gametogenesis, Embryogenesis, and Seed Development. Mol. Plant 2013, 6, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Sorin, C.; Declerck, M.; Christ, A.; Blein, T.; Ma, L.; Lelandais-Brière, C.; Njo, M.F.; Beeckman, T.; Crespi, M.; Hartmann, C. A MiR169 Isoform Regulates Specific NF-YA Targets and Root Architecture in Arabidopsis. New Phytol. 2014, 202, 1197–1211. [Google Scholar] [CrossRef]
- Liu, J.X.; Howell, S.H. BZIP28 and NF-Y Transcription Factors Are Activated by ER Stress and Assemble into a Transcriptional Complex to Regulate Stress Response Genes in Arabidopsis. Plant Cell 2010, 22, 782–796. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Mizoi, J.; Tanaka, H.; Maruyama, K.; Qin, F.; Osakabe, Y.; Morimoto, K.; Ohori, T.; Kusakabe, K.; Nagata, M.; et al. Arabidopsis DPB3-1, A DREB2A Interactor, Specifically Enhances Heat Stress-Induced Gene Expression by Forming a Heat Stress-Specific Transcriptional Complex with NF-Y Subunitsc. Plant Cell 2014, 26, 4954–4973. [Google Scholar] [CrossRef]
- Liu, X.; Hu, P.; Huang, M.; Tang, Y.; Li, Y.; Li, L.; Hou, X. The NF-YC-RGL2 Module Integrates GA and ABA Signalling to Regulate Seed Germination in Arabidopsis. Nat. Commun. 2016, 7, 12768. [Google Scholar] [CrossRef]
- Bello, B.K.; Hou, Y.; Zhao, J.; Jiao, G.; Wu, Y.; Li, Z.; Wang, Y.; Tong, X.; Wang, W.; Yuan, W.; et al. NF-YB1-YC12-BHLH144 Complex Directly Activates Wx to Regulate Grain Quality in Rice (Oryza sativa L.). Plant Biotechnol. J. 2019, 17, 1222–1235. [Google Scholar] [CrossRef]
- Xiong, Y.; Ren, Y.; Li, W.; Wu, F.; Yang, W.; Huang, X.; Yao, J. NF-YC12 Is a Key Multi-Functional Regulator of Accumulation of Seed Storage Substances in Rice. J. Exp. Bot. 2019, 70, 3765–3780. [Google Scholar] [CrossRef]
- Das, S.; Parida, S.K.; Agarwal, P.; Tyagi, A.K. Transcription Factor OsNF-YB9 Regulates Reproductive Growth and Development in Rice. Planta 2019, 250, 1849–1865. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Zhang, Z.; Zhang, J.; Zhou, Y.; Chen, C. The Rice LEC1-like Transcription Factor OsNF-YB9 Interacts with SPK, an Endosperm-Specific Sucrose Synthase Protein Kinase, and Functions in Seed Development. Plant J. 2021, 106, 1233–1246. [Google Scholar] [CrossRef]
- Xu, J.J.; Zhang, X.F.; Xue, H.W. Rice Aleurone Layer Specific OsNF-YB1 Regulates Grain Filling and Endosperm Development by Interacting with an ERF Transcription Factor. J. Exp. Bot. 2016, 67, 6399–6411. [Google Scholar] [CrossRef]
- Su, H.; Chen, Z.; Dong, Y.; Ku, L.; Abou-Elwafa, S.F.; Ren, Z.; Cao, Y.; Dou, D.; Liu, Z.; Liu, H.; et al. Identification of ZmNF-YC2 and Its Regulatory Network for Maize Flowering Time. J. Exp. Bot. 2021, 72, 7792–7807. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Park, H.Y.; Jang, Y.H.; Lee, K.C.; Chung, Y.S.; Lee, J.H.; Kim, J.K. OsNF-YC2 and OsNF-YC4 Proteins Inhibit Flowering under Long-Day Conditions in Rice. Planta 2016, 243, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Xiong, Y.; Xiao, P.; Wang, X.; Yao, J. OsNF-YC10, a Seed Preferentially Expressed Gene Regulates Grain Width by Affecting Cell Proliferation in Rice. Plant Sci. 2019, 280, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Han, M.; Li, S.; Liang, Z.; Ouyang, J.; Wang, X.; Liao, P. A Member of NF-Y Family, OsNF-YC5 Negatively Regulates Salt Tolerance in Rice. Gene 2024, 892, 147868. [Google Scholar] [CrossRef] [PubMed]
- Manimaran, P.; Venkata Reddy, S.; Moin, M.; Raghurami Reddy, M.; Yugandhar, P.; Mohanraj, S.S.; Balachandran, S.M.; Kirti, P.B. Activation-Tagging in Indica Rice Identifies a Novel Transcription Factor Subunit, NF-YC13 Associated with Salt Tolerance. Sci. Rep. 2017, 7, 9341. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Kim, H.I.; Jang, G.; Chung, P.J.; Jeong, J.S.; Kim, Y.S.; Bang, S.W.; Jung, H.; Choi, Y.D.; Kim, J.K. The NF-YA Transcription Factor OsNF-YA7 Confers Drought Stress Tolerance of Rice in an Abscisic Acid Independent Manner. Plant Sci. 2015, 241, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Tanaka, T.; Nakamura, H.; Ichikawa, H.; Kobayashi, K.; Yaeno, T.; Yamaoka, N.; Shimomoto, K.; Takayama, K.; Nishina, H.; et al. Overexpression of a Rice Heme Activator Protein Gene (OsHAP2E) Confers Resistance to Pathogens, Salinity and Drought, and Increases Photosynthesis and Tiller Number. Plant Biotechnol. J. 2015, 13, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Luan, M.; Xu, M.; Lu, Y.; Zhang, Q.; Zhang, L.; Zhang, C.; Fan, Y.; Lang, Z.; Wang, L. Family-Wide Survey of MiR169s and NF-YAs and Their Expression Profiles Response to Abiotic Stress in Maize Roots. PLoS ONE 2014, 9, e91396. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Cao, H.; Wang, X.; Zhang, K.; Si, H.; Zang, J.; Xing, J.; Dong, J. Identification and Expression Analysis of Maize NF-YA Subunit Genes. PeerJ 2022, 10, e14306. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.E.; Repetti, P.P.; Adams, T.R.; Creelman, R.A.; Wu, J.; Warner, D.C.; Anstrom, D.C.; Bensen, R.J.; Castiglioni, P.P.; Donnarummo, M.G.; et al. Plant Nuclear Factor Y (NF-Y) B Subunits Confer Drought Tolerance and Lead to Improved Corn Yields on Water-Limited Acres. Proc. Natl. Acad. Sci. USA 2007, 104, 16450–16455. [Google Scholar] [CrossRef]
- Mei, X.; Liu, C.; Yu, T.; Liu, X.; Xu, D.; Wang, J.; Wang, G.; Cai, Y. Identification and Characterization of Paternal-Preferentially Expressed Gene NF-YC8 in Maize Endosperm. Mol. Genet. Genom. 2015, 290, 1819–1831. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Nan, J.; Zhao, Z.; Yao, S.; Wang, W.; Yang, Y.; Bai, Y.; Dong, E.; Liu, C.; Cai, Y. Maize Transcription Factor ZmNF-YC13 Regulates Plant Architecture. J. Exp. Bot. 2021, 72, 4757–4772. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Yang, Q.; Gan, Y.; Li, M.; Zhao, Z.; Dong, E.; Li, C.; He, D.; Mei, X.; Cai, Y. The ZmNF-YC1–ZmAPRG Pathway Modulates Low Phosphorus Tolerance in Maize. J. Exp. Bot. 2024, 75, 2867–2881. [Google Scholar] [CrossRef]
- Wang, B.; Li, Z.; Ran, Q.; Li, P.; Peng, Z.; Zhang, J. ZmNF-YB16 Overexpression Improves Drought Resistance and Yield by Enhancing Photosynthesis and the Antioxidant Capacity of Maize Plants. Front. Plant Sci. 2018, 9, 709. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, B.; Wang, J.; He, C.; Zhang, D.; Li, P.; Zhang, J.; Li, Z. Transcription Factors ZmNF-YA1 and ZmNF-YB16 Regulate Plant Growth and Drought Tolerance in Maize. Plant Physiol. 2022, 190, 1506–1525. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A Novel Cis-Acting Element in an Arabidopsis Gene Is Involved in Responsiveness to Drought, Low-Temperature, or High-Salt Stress. Plant Cell 1994, 6, 251–264. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Manoli, A.; Sturaro, A.; Trevisan, S.; Quaggiotti, S.; Nonis, A. Evaluation of Candidate Reference Genes for QPCR in Maize. J. Plant Physiol. 2012, 169, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Howell, S.H. Heat Stress Responses and Thermotolerance in Maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the Heat Stress Response in Plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Ni, Z.; Hu, Z.; Jiang, Q.; Zhang, H. GmNFYA3, a Target Gene of MiR169, Is a Positive Regulator of Plant Tolerance to Drought Stress. Plant Mol. Biol. 2013, 82, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Schöffl, F.; Prändl, R.; Reindl, A. Regulation of the Heat-Shock Response. Plant Physiol. 1998, 117, 1135–1141. [Google Scholar] [CrossRef]
- Schlesinger, M.J. Heat Shock Proteins. J. Biol. Chem. 1990, 267, 95–98. [Google Scholar] [CrossRef]
- Hilton, G.R.; Lioe, H.; Stengel, F.; Baldwin, A.J.; Benesch, J.L.P. Small Heat-Shock Proteins: Paramedics of the Cell. Plant Physiol. 2013, 328, 69–98. [Google Scholar]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The Plant Heat Stress Transcription Factor (Hsf) Family: Structure, Function and Evolution. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Nosaka, R.; Hayashi, H.; Tainaka, H.; Maruta, T.; Tamoi, M.; Ikeda, M.; Ohme-Takagi, M.; Yoshimura, K.; Yabuta, Y.; et al. HsfA1d and HsfA1e Involved in the Transcriptional Regulation of Hsfa2 Function as Key Regulators for the Hsf Signaling Network in Response to Environmental Stress. Plant Cell Physiol. 2011, 52, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-H.; Schö Ffl, F. Interaction between Arabidopsis Heat Shock Transcription Factor 1 and 70 KDa Heat Shock Proteins. J. Exp. Bot. 2002, 53, 371–375. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, S.; Zeng, Y.; Wang, X.; Qin, Y.; Ren, Q.; Xiang, S.; Wang, Y.; Xiao, J.; Sun, Y. Reversible Phase Separation of HSF1 Is Required for an Acute Transcriptional Response during Heat Shock. Nat. Cell Biol. 2022, 24, 340–352. [Google Scholar] [CrossRef]
- Charng, Y.Y.; Liu, H.C.; Liu, N.Y.; Chi, W.T.; Wang, C.N.; Chang, S.H.; Wang, T.T. A Heat-Inducible Transcription Factor, HsfA2, Is Required for Extension of Acquired Thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef]
- Schramm, F.; Ganguli, A.; Kiehlmann, E.; Englich, G.; Walch, D.; Von Koskull-Döring, P. The Heat Stress Transcription Factor HsfA2 Serves as a Regulatory Amplifier of a Subset of Genes in the Heat Stress Response in Arabidopsis. Plant Mol. Biol. 2006, 60, 759–772. [Google Scholar] [CrossRef]
- Huang, Y.C.; Niu, C.Y.; Yang, C.R.; Jinn, T.L. The Heat Stress Factor HSFA6b Connects ABA Signaling and ABA-Mediated Heat Responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant Heat Stress: Concepts Directing Future Research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef] [PubMed]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of Photosynthesis in a C4 Plant, Maize, to Heat Stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Yang, C.; Xu, J.; Lu, H.P.; Liu, J.X. The Hot Science in Rice Research: How Rice Plants Cope with Heat Stress. Plant Cell Environ. 2023, 46, 1087–1103. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, I.; Kirat, K.; Pati, P. Detection of Reactive Oxygen Species in Oryza sativa L. (Rice). Bio Protoc. 2016, 6, e2061. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis Mesophyll Protoplasts: A Versatile Cell System for Transient Gene Expression Analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Xu, X.H.; Chen, H.; Sang, Y.L. Identification of genes specifically or preferentially expressed in maize silk reveals similarity and diversity in transcript abundance of different dry stigmas. BMC Genom. 2012, 13, 294. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Sartor, R.C.; Shen, Z. Integration of omic networks in a developmental atlas of maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Wang, F.; Tan, S.J.; Wang, M.X.; Sui, N.; Zhang, X.S. Transcript profiles of maize embryo sacs and preliminary identification of genes involved in the embryo sac–pollen tube interaction. Front Plant Sci. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Swart, V.; Crampton, B.G.; Ridenour, J.B. Complementation of ctb7 in the maize pathogen cercospora zeina overcomes the lack of in vitro cercosporin production. Mol. Plant-Microbe Interact 2017, 30, 710–724. [Google Scholar] [CrossRef]
- Makarevitch, I.; Waters, A.J.; West, P.T. Transposable Elements Contribute to Activation of Maize Genes in Response to Abiotic Stress. PLoS Genet. 2015, 11, e1004915. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Li, Z.; Zhang, J. ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response. Int. J. Mol. Sci. 2024, 25, 6275. https://doi.org/10.3390/ijms25116275
Yang Y, Li Z, Zhang J. ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response. International Journal of Molecular Sciences. 2024; 25(11):6275. https://doi.org/10.3390/ijms25116275
Chicago/Turabian StyleYang, Yaling, Zhaoxia Li, and Juren Zhang. 2024. "ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response" International Journal of Molecular Sciences 25, no. 11: 6275. https://doi.org/10.3390/ijms25116275
APA StyleYang, Y., Li, Z., & Zhang, J. (2024). ZmNF-YA1 Contributes to Maize Thermotolerance by Regulating Heat Shock Response. International Journal of Molecular Sciences, 25(11), 6275. https://doi.org/10.3390/ijms25116275