
Altered Morpho-Functional Features of Neurogenesis in Zebrafish Embryos Exposed to Non-Combustion-Derived Magnetite
 , and
, and
Abstract
:1. Introduction
2. Results
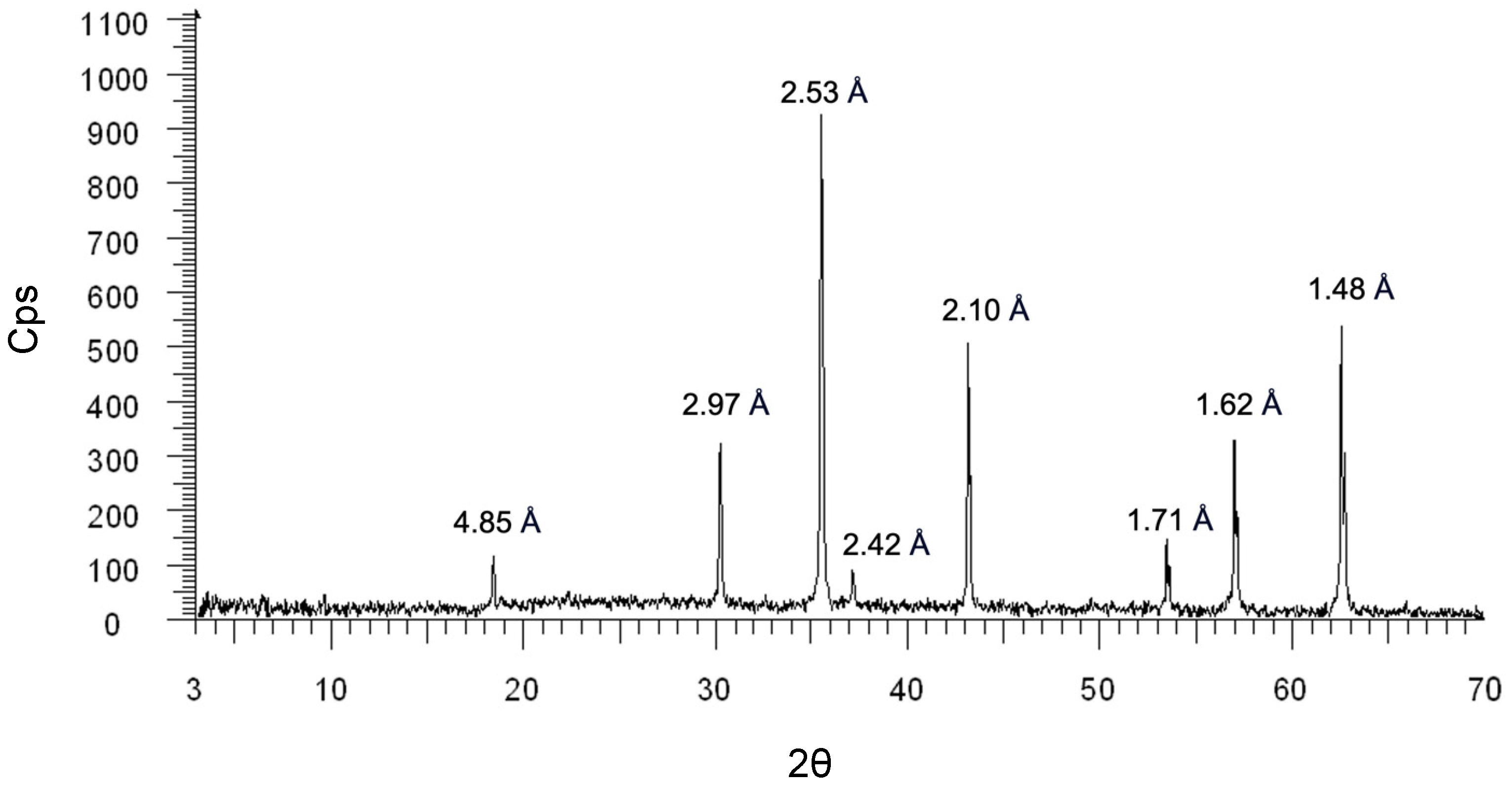
2.1. Mineralogical Characterization of Magnetite
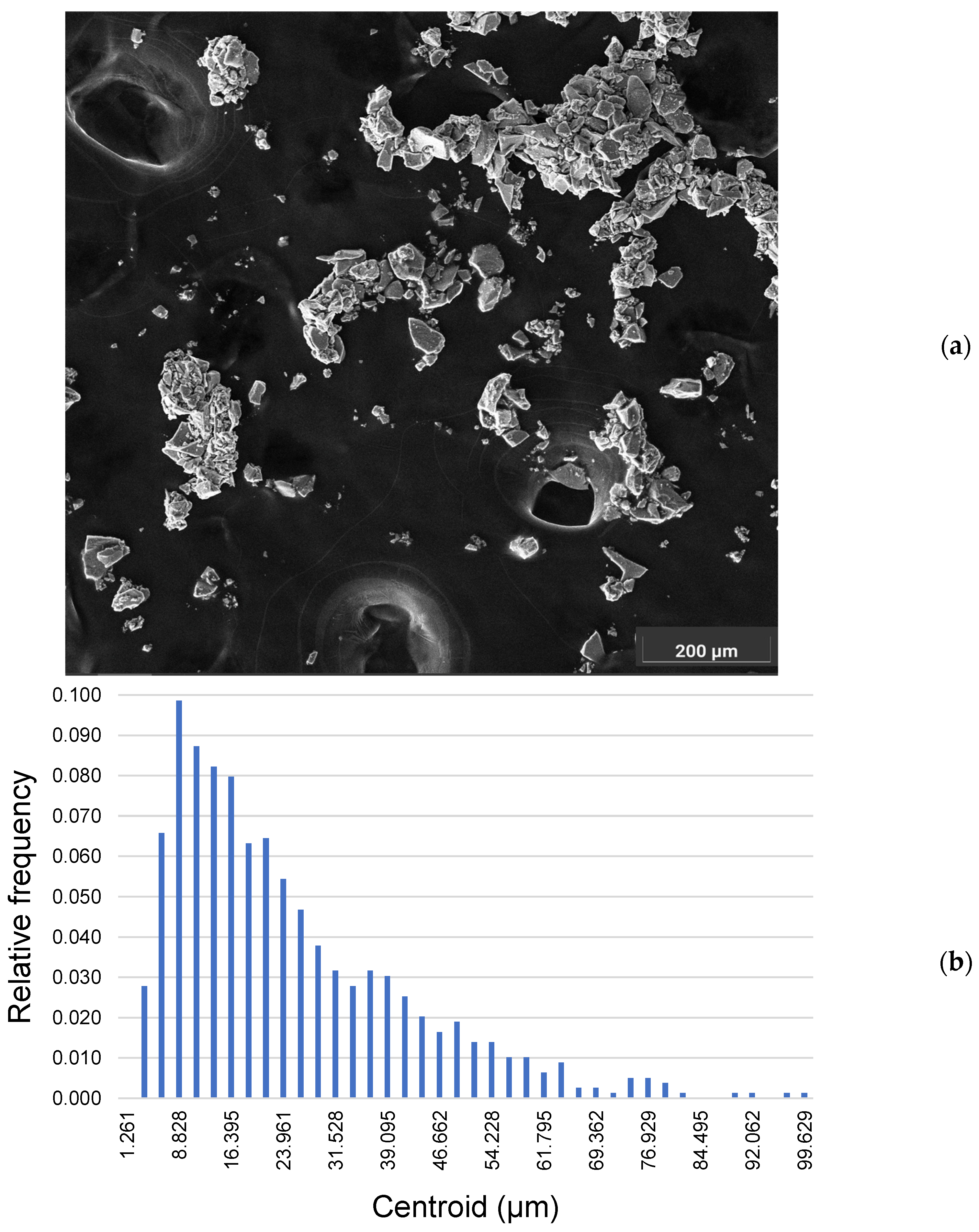
2.2. Morphometric Evaluation of Magnetite
2.3. Atomic Resolution Microscopy and Dual-EELS on Magnetite Nanoparticles and Cleavage Fragments
2.4. Toxicological Monitoring of Zebrafish Embryos
2.5. Magnetite Exposure Increases Oxidative Stress and Apoptosis in Zebrafish Embryo Brain
2.6. Magnetite Treatment Decreases Neural Progenitors in Zebrafish Embryo
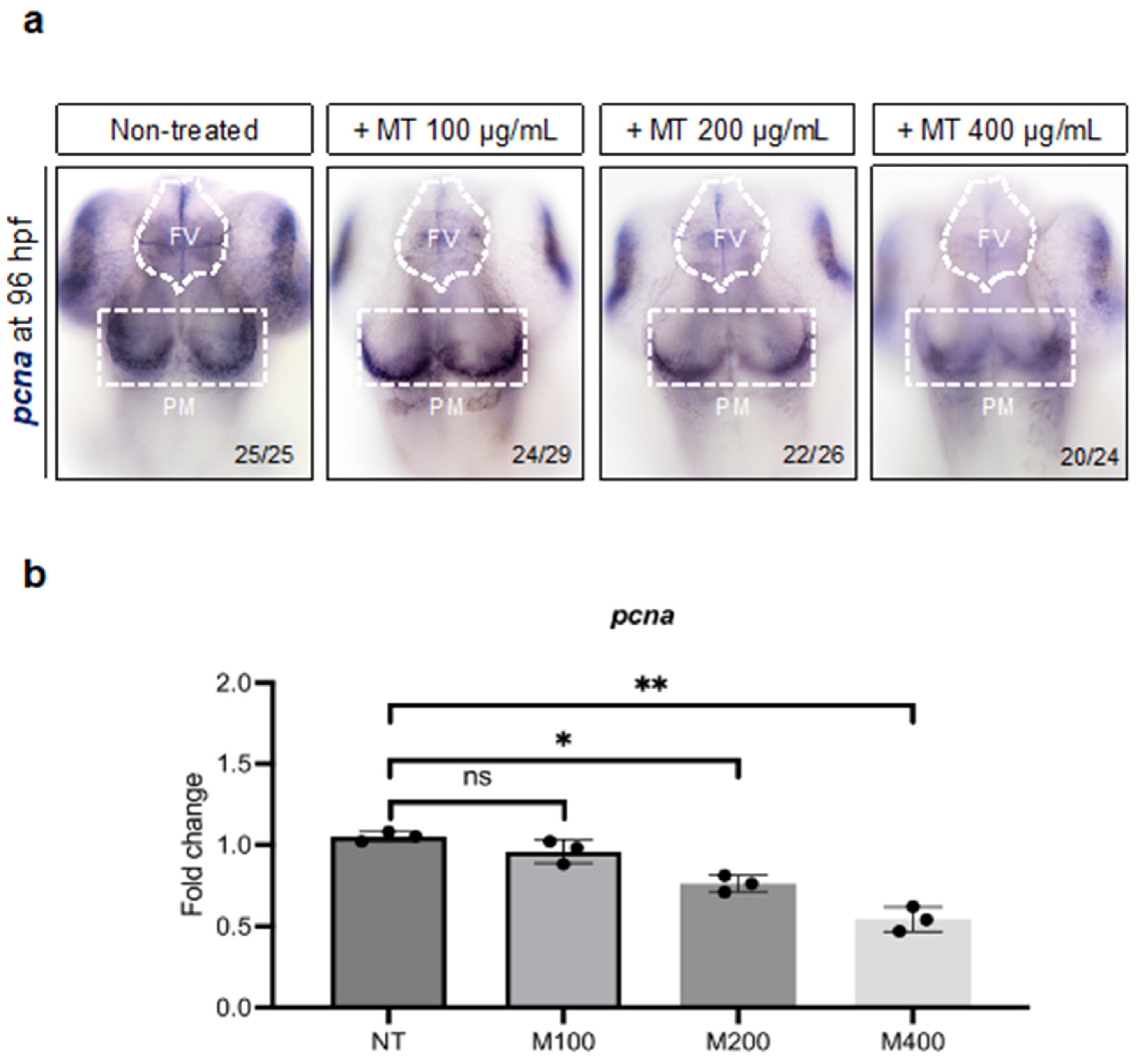
2.7. Magnetite Treatment Affects Neural Cell Proliferation in Embryo Zebrafish Brain
2.8. Magnetite Exposure Increases Inflammation and Microglia Cells in Embryo Zebrafish Brain
2.9. Impact of Magnetite Exposure on Embryo Neuron Specification
3. Discussion
4. Material and Methods
4.1. Powder X-ray Diffraction (PXRD)
4.2. Scanning Electron Microscopy
4.3. Atomic Resolution Scanning Electron Microscopy (ARM) and Dual-Electron Energy-Loss Spectroscopy (Dual-EELS)
4.4. Zebrafish Embryos
4.5. Reactive Oxygen Species (ROS) Detection and TUNEL Assay
4.6. Whole-Mount In Situ Hybridization (WISH)
4.7. RNA Extraction and Reverse Transcription
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9. Statistical Procedures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Accogli, A.; Addour-Boudrahem, N.; Srour, M. Neurogenesis, neuronal migration, and axon guidance. Handb. Clin. Neurol. 2020, 173, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Altbuerger, C.; Rath, M.; Armbruster, D.; Driever, W. Neurog1 and Olig2 integrate patterning and neurogenesis signals in development of zebrafish dopaminergic and glutamatergic dual transmitter neurons. Dev. Biol. 2024, 505, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Allende, M.L.; Weinberg, E.S. The Expression Pattern of 2 Zebrafish Achaete-Scute Homolog (Ash) Genes Is Altered in the Embryonic Brain of the Cyclops Mutant. Dev. Biol. 1994, 166, 509–530. [Google Scholar] [CrossRef] [PubMed]
- Spassky, N.; Aguilar, A. Shh regulates neurogenesis through primary cilia. Med. Sci. 2008, 24, 790–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.K.; Emelyanov, A.; Korzh, V.; Gong, Z.Y. Zebrafish homologue is expressed during neurogenesis in embryonic development. Dev. Dynam 2003, 227, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Samarut, E.; Bekri, A.; Drapeau, P. Transcriptomic Analysis of Purified Embryonic Neural Stem Cells from Zebrafish Embryos Reveals Signaling Pathways Involved in Glycine-Dependent Neurogenesis. Front. Mol. Neurosci. 2016, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Valero, J.; Paris, I.; Sierra, A. Lifestyle Shapes the Dialogue between Environment, Microglia, and Adult Neurogenesis. ACS Chem. Neurosci. 2016, 7, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Briones, T.L. Environment, physical activity, and neurogenesis: Implications for prevention and treatment of Alzhemier’s disease. Curr. Alzheimer Res. 2006, 3, 49–54. [Google Scholar] [CrossRef]
- Costa, L.G.; Cole, T.B.; Coburn, J.; Chang, Y.C.; Dao, K.; Roque, P.J. Neurotoxicity of traffic-related air pollution. Neurotoxicology 2017, 59, 133–139. [Google Scholar] [CrossRef]
- Boda, E.; Rigamonti, A.E.; Bollati, V. Understanding the effects of air pollution on neurogenesis and gliogenesis in the growing and adult brain. Curr. Opin. Pharmacol. 2020, 50, 61–66. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef]
- Liu, X.Q.; Huang, J.; Song, C.; Zhang, T.L.; Liu, Y.P.; Yu, L. Neurodevelopmental toxicity induced by PM2.5 Exposure and its possible role in Neurodegenerative and mental disorders. Hum. Exp. Toxicol. 2023, 42, 9603271231191436. [Google Scholar] [CrossRef]
- Waidyatillake, N.T.; Campbell, P.T.; Vicendese, D.; Dharmage, S.C.; Curto, A.; Stevenson, M. Particulate Matter and Premature Mortality: A Bayesian Meta-Analysis. Int. J. Environ. Res. Public Health 2021, 18, 7655. [Google Scholar] [CrossRef]
- Liu, C.; Chen, R.; Sera, F.; Vicedo-Cabrera, A.M.; Guo, Y.; Tong, S.; Coelho, M.; Saldiva, P.H.N.; Lavigne, E.; Matus, P.; et al. Ambient Particulate Air Pollution and Daily Mortality in 652 Cities. N. Engl. J. Med. 2019, 381, 705–715. [Google Scholar] [CrossRef]
- Fan, J.X.; Yang, J.X.; Cheng, F.L.; Zhang, S.K. The Source, Distribution, and Environmental Effects of Suspended Particulate Matter in the Yangtze River System. Water 2023, 15, 3429. [Google Scholar] [CrossRef]
- Zhao, Z.L.; Wang, L.; Shi, W.H.; Li, C.; Wei, G.Z.J. Motion Adsorption Characteristics of Particulate Matter in Water Supply Network. Water 2022, 14, 3550. [Google Scholar] [CrossRef]
- Collingwood, J.F. Brain iron dysregulation in neurodegenerative disease: Insights from systems modelling and synchrotron spectromicroscopy. Febs. Open Biol. 2018, 8, 33. [Google Scholar]
- Pankhurst, Q.; Hautot, D.; Khan, N.; Dobson, J. Increased levels of magnetic iron compounds in Alzheimer’s disease. J. Alzheimers Dis. 2008, 13, 49–52. [Google Scholar] [CrossRef]
- Giere, R. Magnetite in the human body: Biogenic vs. anthropogenic. Proc. Natl. Acad. Sci. USA 2016, 113, 11986–11987. [Google Scholar] [CrossRef] [PubMed]
- Tabner, B.J.; Mayes, J.; Allsop, D. Hypothesis: Soluble abeta oligomers in association with redox-active metal ions are the optimal generators of reactive oxygen species in Alzheimer’s disease. Int. J. Alzheimers Dis. 2010, 2011, 546380. [Google Scholar] [CrossRef] [PubMed]
- Maher, B.A.; Ahmed, I.A.; Karloukovski, V.; MacLaren, D.A.; Foulds, P.G.; Allsop, D.; Mann, D.M.; Torres-Jardon, R.; Calderon-Garciduenas, L. Magnetite pollution nanoparticles in the human brain. Proc. Natl. Acad. Sci. USA 2016, 113, 10797–10801. [Google Scholar] [CrossRef]
- Hautot, D.; Pankhurst, Q.A.; Khan, N.; Dobson, J. Preliminary evaluation of nanoscale biogenic magnetite in Alzheimer’s disease brain tissue. Proc. Biol. Sci. 2003, 270 (Suppl. S1), S62–S64. [Google Scholar] [CrossRef] [PubMed]
- Hussain, R.; Graham, U.; Elder, A.; Nedergaard, M. Air pollution, glymphatic impairment, and Alzheimer’s disease. Trends Neurosci. 2023, 46, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Cole-Hunter, T.; Zhang, J.; So, R.; Samoli, E.; Liu, S.; Chen, J.; Strak, M.; Wolf, K.; Weinmayr, G.; Rodopolou, S.; et al. Long-term air pollution exposure and Parkinson’s disease mortality in a large pooled European cohort: An ELAPSE study. Environ. Int. 2023, 171, 107667. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.M.; Miyashita, L.; Maher, B.A.; McPhail, G.; Jones, C.J.P.; Barratt, B.; Thangaratinam, S.; Karloukovski, V.; Ahmed, I.A.; Aslam, Z.; et al. Evidence for the presence of air pollution nanoparticles in placental tissue cells. Sci. Total Environ. 2021, 751, 142235. [Google Scholar] [CrossRef] [PubMed]
- Coccini, T.; Pignatti, P.; Spinillo, A.; De Simone, U. Developmental Neurotoxicity Screening for Nanoparticles Using Neuron-Like Cells of Human Umbilical Cord Mesenchymal Stem Cells: Example with Magnetite Nanoparticles. Nanomaterials 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Basaki, M.; Keykavusi, K.; Sahraiy, N.; Ali Shahbazfar, A. Maternal exposure to iron oxide nanoparticles is associated with ferroptosis in the brain: A chicken embryo model analysis. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Semeano, A.T.; Tofoli, F.A.; Correa-Velloso, J.C.; de Jesus Santos, A.P.; Oliveira-Giacomelli, A.; Cardoso, R.R.; Pessoa, M.A.; da Rocha, E.L.; Ribeiro, G.; Ferrari, M.F.R.; et al. Effects of Magnetite Nanoparticles and Static Magnetic Field on Neural Differentiation of Pluripotent Stem Cells. Stem Cell Rev. Rep. 2022, 18, 1337–1354. [Google Scholar] [CrossRef] [PubMed]
- Engert, F.; Wilson, S.W. Zebrafish neurobiology: From development to circuit function and behaviour. Dev. Neurobiol. 2012, 72, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Eisen, J.S. Developmental neurobiology of the zebrafish. J. Neurosci. 1991, 11, 311–317. [Google Scholar] [CrossRef]
- Adams, M.M.; Kafaligonul, H. Zebrafish-A Model Organism for Studying the Neurobiological Mechanisms Underlying Cognitive Brain Aging and Use of Potential Interventions. Front. Cell Dev. Biol. 2018, 6, 135. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Strahle, U.; Scholpp, S. Neurogenesis in zebrafish—From embryo to adult. Neural Dev. 2013, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, V.; Ravichandran, P.; Copeland, C.L.; Gopikrishnan, R.; Biradar, S.; Goornavar, V.; Ramesh, G.T.; Hall, J.C. Magnetite induces oxidative stress and apoptosis in lung epithelial cells. Mol. Cell Biochem. 2012, 363, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, S.; Asokan, N.; Bludau, O.; Chae, J.; Kuscha, V.; Kaslin, J.; Hans, S. Neurogenesis in the inner ear: The zebrafish statoacoustic ganglion provides new neurons from a Neurod/Nestin-positive progenitor pool well into adulthood. Development 2020, 147, dev.176750. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Mueller, T. Identification and morphogenesis of the eminentia thalami in the zebrafish. J. Comp. Neurol. 2004, 471, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.L.; Jorgensen, A.L. Structural and functional characterization of the zebrafish gene for glial fibrillary acidic protein, GFAP. Gene 2003, 310, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Ricci, S.; Frabetti, F.; Ferrando, S.; Franceschini, V. Exposure of Zebrafish Embryos to Urea Affects Gene Expression in Neuronal Cells. Environments 2024, 11, 41. [Google Scholar] [CrossRef]
- Rocha, J.M.V.; de Souza, V.B.; Panunto, P.C.; Nicolosi, J.S.; da Silva, E.D.N.; Cadore, S.; Londono, O.M.; Muraca, D.; Tancredi, P.; de Brot, M.; et al. In vitro and in vivo acute toxicity of a novel citrate-coated magnetite nanoparticle. PLoS ONE 2022, 17, e0277396. [Google Scholar] [CrossRef]
- Schoonen, M.A.A.; Cohn, C.A.; Roemer, E.; Laffers, R.; Simon, S.R.; O’Riordan, T. Mineral-induced formation of reactive oxygen species. Rev. Miner. Geochem. 2006, 64, 179–221. [Google Scholar] [CrossRef]
- Niklison-Chirou, M.V.; Agostini, M.; Amelio, I.; Melino, G. Regulation of Adult Neurogenesis in Mammalian Brain. Int. J. Mol. Sci. 2020, 21, 4869. [Google Scholar] [CrossRef]
- Bolton, J.L.; Marinero, S.; Hassanzadeh, T.; Natesan, D.; Le, D.; Belliveau, C.; Mason, S.N.; Auten, R.L.; Bilbo, S.D. Gestational Exposure to Air Pollution Alters Cortical Volume, Microglial Morphology, and Microglia-Neuron Interactions in a Sex-Specific Manner. Front. Synaptic Neurosci. 2017, 9, 10. [Google Scholar] [CrossRef]
- Gonet, T.; Maher, B.A.; Kukutschova, J. Source apportionment of magnetite particles in roadside airborne particulate matter. Sci. Total Environ. 2021, 752, 141828. [Google Scholar] [CrossRef]
- Maher, B.A. Airborne Magnetite- and Iron-Rich Pollution Nanoparticles: Potential Neurotoxicants and Environmental Risk Factors for Neurodegenerative Disease, Including Alzheimer’s Disease. J. Alzheimers Dis. 2019, 71, 361–375. [Google Scholar] [CrossRef]
- Vigliaturo, R.; Jamnik, M.; Drazic, G.; Podobnik, M.; Znidaric, M.T.; Ventura, G.D.; Redhammer, G.J.; Znidarsic, N.; Caserman, S.; Giere, R. Nanoscale transformations of amphiboles within human alveolar epithelial cells. Sci. Rep. 2022, 12, 1782. [Google Scholar] [CrossRef]
- Moore, D.M.; Reynolds, R.C. X-ray Diffraction and the Identification and Analysis of Clay Minerals; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Vigliaturo, R.; Pollastri, S.; Gieré, R.; Gualtieri, A.F.; Drazic, G. Experimental quantification of the Fe-valence state at amosite-asbestos boundaries using acSTEM dual-electron energy-loss spectroscopy. Am. Mineral. 2019, 104, 1820–1828. [Google Scholar] [CrossRef]
- Cacialli, P.; Dogan, S.; Linnerz, T.; Pasche, C.; Bertrand, J.Y. Minichromosome maintenance protein 10 (mcm10) regulates hematopoietic stem cell emergence in the zebrafish embryo. Stem Cell Rep. 2023, 18, 1534–1546. [Google Scholar] [CrossRef]
- Cacialli, P.; Mailhe, M.P.; Wagner, I.; Merkler, D.; Golub, R.; Bertrand, J.Y. Synergistic prostaglandin E synthesis by myeloid and endothelial cells promotes fetal hematopoietic stem cell expansion in vertebrates. EMBO J. 2022, 41, e108536. [Google Scholar] [CrossRef]
- Jurewicz, A.; Ilyas, S.; Uppal, J.K.; Ivandic, I.; Korsching, S.; Mathur, S. Evaluation of Magnetite Nanoparticle-Based Toxicity on Embryo-Larvae Stages of Zebrafish (Danio rerio). ACS Appl. Nano Mater. 2020, 3, 1621–1629. [Google Scholar] [CrossRef]
- Cacialli, P.; Mahony, C.B.; Petzold, T.; Bordignon, P.; Rougemont, A.L.; Bertrand, J.Y. A connexin/ifi30 pathway bridges HSCs with their niche to dampen oxidative stress. Nat. Commun. 2021, 12, 4484. [Google Scholar] [CrossRef]
- Mahony, C.B.; Cacialli, P.; Pasche, C.; Monteiro, R.; Savvides, S.N.; Bertrand, J.Y. Hapln1b, a central organizer of the ECM, modulates kit signaling to control developmental hematopoiesis in zebrafish. Blood Adv. 2021, 5, 4935–4948. [Google Scholar] [CrossRef]
- Cacialli, P. Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish. Vet. Sci. 2022, 9, 225. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Treated | Magnetite 100 µg | Magnetite 200 µg | Magnetite 400 µg | |
|---|---|---|---|---|
| Nestin | +++ | +++ | ++ | + |
| sox2 | +++ | +++ | ++ | + |
| Pcna | +++ | +++ | ++ | + |
| elavl3 | +++ | +++ | ++ | + |
| Gfap | +++ | +++ | ++++ | +++++ |
| Apoeb | +++ | +++ | ++++ | +++++ |
| il1b | +++ | +++ | ++++ | +++++ |
| Nestin | F: 5′-GCAGCCAACAACTATCAGAAAC-3′ | R: 5′-CATCGAGGTACTGCTTGGT-3′ |
| Pcna | F: 5′-CCTTAAGAAGGTCCTGGAG-3′ | R: 5′-CCACACAACTGTATTCCTGCTC-3′ |
| elavl3 | F: 5′-CCATGGAAACTCAGGTGTC-3′ | R: 5′-GTCAGCTGCTCCTAGT-3′ |
| gfap | F: 5′-CATCTATCAGGAGGAGCTG-3′ | R: 5′-CTCAGCTGGCGCTCCA-3′ |
| sox2 | F: 5′-CCCTGATGAAGAAGGACAAGT-3′ | R: 5′-GTTGTGCGCGTTCAAACTC-3′ |
| apoeb | F: 5′-CACAAACTGACGGCATGGT-3′ | R: 5′-CGGTTCTTCACGTCATCTG-3′ |
| nestin | F: 5′-CTTCAACATCTTCAGGCCCAAG-3′ | R:5′-GTGTTGGTCTGTCGATTCTCAG-3′ |
| pcna | F: 5′-CAAGGAGGATGAAGCGGTAACA-3′ | R: 5′-CTGCGGACATGCTAAGTGTG-3′ |
| elavl3 | F: 5′-GCCAGCTACGGAGTCAAGAG-3′ | R: 5′-CATGGTGACGAAGCCAAAGC-3′ |
| gfap | F: 5′-ACCCGTGACGGAGAGATCAT-3′ | R: 5′-GCCAGTGTCTGAGCCTCATT-3′ |
| sod2 | F: 5′-CAGCAAGCACCATGCAACAT-3′ | R: 5′-CAGCTCACCCTGTGGTTCTC-3′ |
| cat | F: 5′-TGAGGCTGGGTCATCAGATA-3′ | R: 5′-AAAGACGGAAACAGAAGCGT-3′ |
| gsr | F: 5′-CTCCTTGGTCGCAGCATGGCT-3′ | R: 5′-GGCAGTGGTGGCACCGAGTTC-3′ |
| nrf2 | F: 5′-TGTTGGTTCGGAGGCTCTTAA-3′ | R: 5′-AGGCCATGTCCACACGTACA-3′ |
| il1b | F: 5′-ATGGCGAACGTCATCCAAGA-3′ | R: 5′-GAGACCCGCTGATCTCCTTG-3′ |
| ef1a | F: 5′-CCTGGGAGTGAAACAGCTG-3′ | R: 5′-GCCTCCAGCATGTTGTCAC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacialli, P.; Ricci, S.; Servetto, G.P.; Franceschini, V.; Ruiz-Zepeda, F.; Vigliaturo, R. Altered Morpho-Functional Features of Neurogenesis in Zebrafish Embryos Exposed to Non-Combustion-Derived Magnetite. Int. J. Mol. Sci. 2024, 25, 6459. https://doi.org/10.3390/ijms25126459
Cacialli P, Ricci S, Servetto GP, Franceschini V, Ruiz-Zepeda F, Vigliaturo R. Altered Morpho-Functional Features of Neurogenesis in Zebrafish Embryos Exposed to Non-Combustion-Derived Magnetite. International Journal of Molecular Sciences. 2024; 25(12):6459. https://doi.org/10.3390/ijms25126459
Chicago/Turabian StyleCacialli, Pietro, Serena Ricci, Giulia Pia Servetto, Valeria Franceschini, Francisco Ruiz-Zepeda, and Ruggero Vigliaturo. 2024. "Altered Morpho-Functional Features of Neurogenesis in Zebrafish Embryos Exposed to Non-Combustion-Derived Magnetite" International Journal of Molecular Sciences 25, no. 12: 6459. https://doi.org/10.3390/ijms25126459