


Rare Filaggrin Variants Are Associated with Pustular Skin Diseases in Asians

 ,
,  , , , , , , ,
, , , , , , ,  , , and
, , and
Abstract
1. Introduction
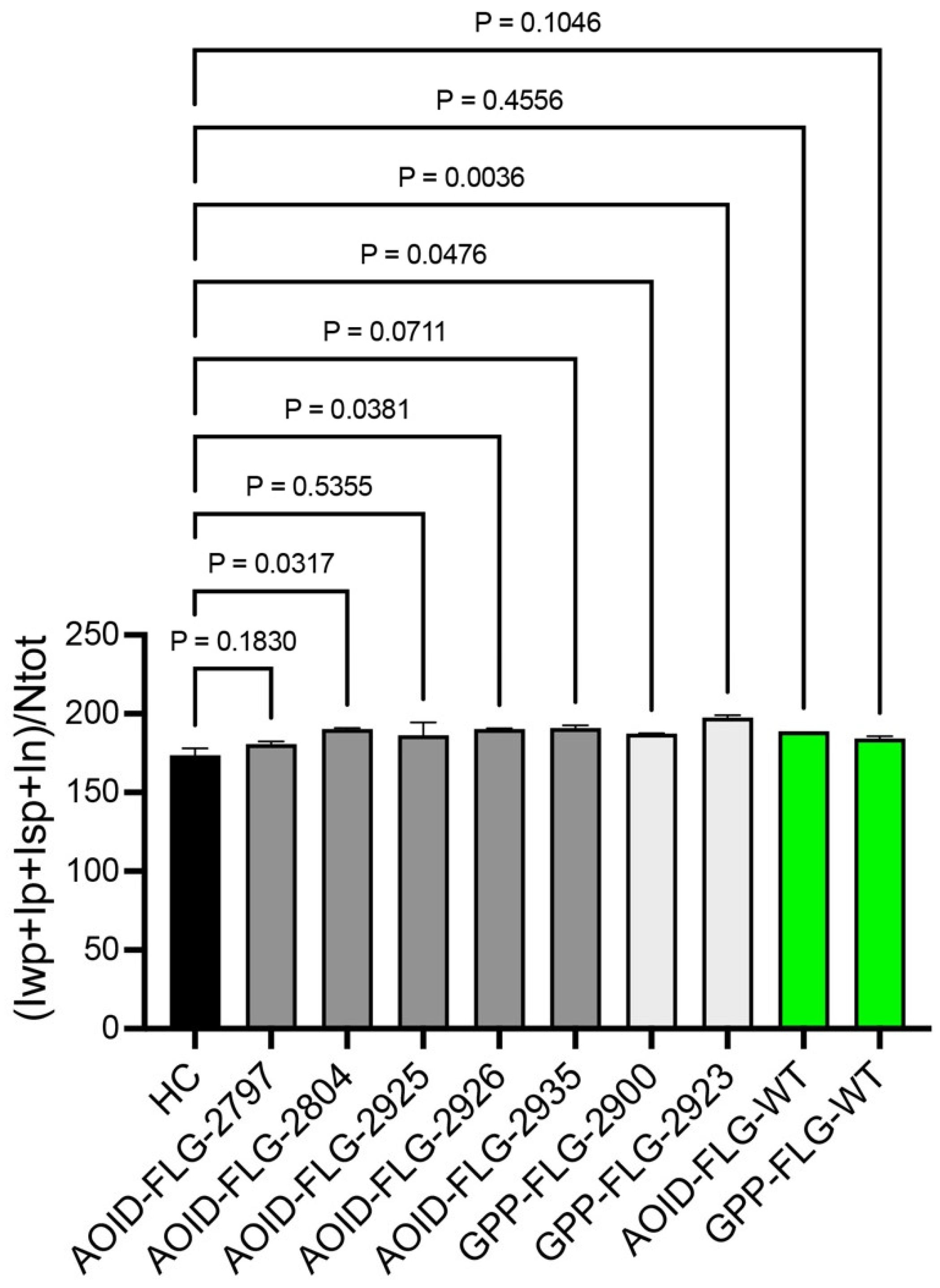
2. Results and Discussion
2.1. Demographic Data
2.2. Several Rare Unreported Genetic Variants of Filaggrin Are Associated with RPEs
2.3. The Newly Identified Variants Have an Impact on FLG Expression
3. Patients and Methods
3.1. Study Subjects
3.2. Molecular Analysis and Assessment of Rare Variants
3.3. Immunohistochemistry
3.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Jutivorakool, K.; Sittiwattanawong, P.; Kantikosum, K.; Hurst, C.P.; Kumtornrut, C.; Asawanonda, P.; Klaewsongkram, J.; Rerknimitr, P. Skin Manifestations in Patients with Adult-onset Immunodeficiency due to Anti-interferon-gamma Autoantibody: A Relationship with Systemic Infections. Acta Derm. Venereol. 2018, 98, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Kantaputra, P.; Daroontum, T.; Chuamanochan, M.; Chaowattanapanit, S.; Kiratikanon, S.; Choonhakarn, C.; Intachai, W.; Olsen, B.; Tongsima, S.; Ngamphiw, C.; et al. SERPINB3, Adult-Onset Immunodeficiency, and Generalized Pustular Psoriasis. Genes 2023, 14, 266. [Google Scholar] [CrossRef] [PubMed]
- Uppala, R.; Tsoi, L.C.; Harms, P.W.; Wang, B.; Billi, A.C.; Maverakis, E.; Michelle Kahlenberg, J.; Ward, N.L.; Gudjonsson, J.E. “Autoinflammatory psoriasis”-genetics and biology of pustular psoriasis. Cell Mol. Immunol. 2021, 18, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Kantaputra, P.; Chaowattanapanit, S.; Kiratikanon, S.; Chaiwarith, R.; Choonhakarn, C.; Intachai, W.; Quarto, N.; Tongsima, S.; Ketudat Cairns, J.R.; Ngamphiw, C.; et al. SERPINA1, generalized pustular psoriasis, and adult-onset immunodeficiency. J. Dermatol. 2021, 48, 1597–1601. [Google Scholar] [CrossRef] [PubMed]
- Kantaputra, P.N.; Chuamanochan, M.; Kiratikanon, S.; Chiewchanvit, S.; Chaiwarith, R.; Intachai, W.; Quarto, N.; Tongsima, S.; McGrath, J.A.; Ngamphiw, C. A truncating variant in SERPINA3, skin pustules and adult-onset immunodeficiency. J. Dermatol. 2021, 48, e370–e371. [Google Scholar] [CrossRef] [PubMed]
- Marrakchi, S.; Puig, L. Pathophysiology of Generalized Pustular Psoriasis. Am. J. Clin. Dermatol. 2022, 23 (Suppl. S1), 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.L.; Chi, C.Y.; von Bernuth, H.; Doffinger, R. Autoantibodies against cytokines: Phenocopies of primary immunodeficiencies? Hum. Genet. 2020, 139, 783–794. [Google Scholar] [CrossRef]
- Kantaputra, P.; Daroontum, T.; Chuamanochan, M.; Chaowattanapanit, S.; Intachai, W.; Olsen, B.; Sastraruji, T.; Tongsima, S.; Ngamphiw, C.; Kampuansai, J.; et al. Loss of Function TGFBR2 Variant as a Contributing Factor in Generalized Pustular Psoriasis and Adult-Onset Immunodeficiency. Genes 2022, 14, 103. [Google Scholar] [CrossRef] [PubMed]
- Pithukpakorn, M.; Roothumnong, E.; Angkasekwinai, N.; Suktitipat, B.; Assawamakin, A.; Luangwedchakarn, V.; Umrod, P.; Thongnoppakhun, W.; Foongladda, S.; Suputtamongkol, Y. HLA-DRB1 and HLA-DQB1 Are Associated with Adult-Onset Immunodeficiency with Acquired Anti-Interferon-Gamma Autoantibodies. PLoS ONE 2015, 10, e0128481. [Google Scholar] [CrossRef] [PubMed]
- Kuechle, M.K.; Presland, R.B.; Lewis, S.P.; Fleckman, P.; Dale, B.A. Inducible expression of filaggrin increases keratinocyte susceptibility to apoptotic cell death. Cell Death Differ. 2000, 7, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; McLean, W.H. One remarkable molecule: Filaggrin. J. Investig. Dermatol. 2012, 132 Pt 2, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122 Pt 9, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.K.; Akiyama, M.; Nemoto-Hasebe, I.; Nomura, T.; Sandilands, A.; Chao, S.C.; Lee, J.Y.; Sheu, H.M.; McLean, W.H.; Shimizu, H. Analysis of Taiwanese ichthyosis vulgaris families further demonstrates differences in FLG mutations between European and Asian populations. Br. J. Dermatol. 2009, 161, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Common, J.E.; Haines, R.L.; Balakrishnan, A.; Brown, S.J.; Goh, C.S.; Cordell, H.J.; Sandilands, A.; Campbell, L.E.; Kroboth, K.; et al. Wide spectrum of filaggrin-null mutations in atopic dermatitis highlights differences between Singaporean Chinese and European populations. Br. J. Dermatol. 2011, 165, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.J.; Gupta, J.; Apter, A.J.; Hoffstad, O.; Papadopoulos, M.; Rebbeck, T.R.; Wubbenhorst, B.; Mitra, N. Exome sequencing of filaggrin and related genes in African-American children with atopic dermatitis. J. Investig. Dermatol. 2014, 134, 2272–2274. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Margolis, D.J. Filaggrin gene mutations with special reference to atopic dermatitis. Curr. Treat. Options Allergy 2020, 7, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.J.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Eaaswarkhanth, M.; Xu, D.; Flanagan, C.; Rzhetskaya, M.; Hayes, M.G.; Blekhman, R.; Jablonski, N.G.; Gokcumen, O. Atopic Dermatitis Susceptibility Variants in Filaggrin Hitchhike Hornerin Selective Sweep. Genome Biol. Evol. 2016, 8, 3240–3255. [Google Scholar] [CrossRef] [PubMed]
- Shotelersuk, V.; Wichadakul, D.; Ngamphiw, C.; Srichomthong, C.; Phokaew, C.; Wilantho, A.; Pakchuen, S.; Nakhonsri, V.; Shaw, P.J.; Wasitthankasem, R.; et al. The Thai reference exome (T-REx) variant database. Clin. Genet. 2021, 100, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Noe, M.H.; Anadkat, M.J.; Armstrong, A.; Bhutani, T.; Farberg, A.; Han, G.; Hawkes, J.E.; Lebwohl, M.; Rekhtman, S.; et al. Development of the Generalized Pustular Psoriasis Clinical Assessment Tool. J. Am. Acad. Dermatol. 2024, 90, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.P.; Brown, S.B.; Griffiths, C.E.; Weller, R.B.; Gibbs, N.K. Feeding filaggrin: Effects of l-histidine supplementation in atopic dermatitis. Clin. Cosmet. Investig. Dermatol. 2017, 10, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, B.; Riedel, R.; Jorss, K.; Loddenkemper, C.; Steinmeyer, A.; Zugel, U.; Babina, M.; Radbruch, A.; Worm, M. Vitamin D receptor activation improves allergen-triggered eczema in mice. J. Investig. Dermatol. 2012, 132, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Navarini, A.A.; Burden, A.D.; Capon, F.; Mrowietz, U.; Puig, L.; Koks, S.; Kingo, K.; Smith, C.; Barker, J.N.; Network, E. European consensus statement on phenotypes of pustular psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1792–1799. [Google Scholar] [CrossRef] [PubMed]
- Poplin, R.; Ruano-Rubio, V.; DePristo, M.A.; Fennell, T.J.; Carneiro, M.O.; Van der Auwera, G.A.; Kling, D.E.; Gauthier, L.D.; Levy-Moonshine, A.; Roazen, D.; et al. Scaling accurate genetic variant discovery to tens of thousands of samples. BioRxiv 2017, 201178. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, C.; Mou, C.; Dong, Y.; Tu, Y. dbNSFP v4: A comprehensive database of transcript-specific functional predictions and annotations for human nonsynonymous and splice-site SNVs. Genome Med. 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Condition | FLG | FLG3 |
|---|---|---|---|
| 2787 | AOID-PR | p.Asn1397Asp (rs112252908), p.Val1395Ala (rs527894804), p.Ser1390Tyr (rs139061200) | n.d. |
| 2797 | AOID-PR | p.Arg1474Gln (rs200551704) | p.Leu994TrpfsTer57 (rs749162569) |
| 2803 | AOID-PR | n.d. | p.Ser1779Asn (rs768542886) |
| 2804 | AOID-PR | p.Ser2706Ter (rs542799026) | n.d. |
| 2898 | AOID-PR | n.d. | p.Ser967Pro (rs200674313) |
| 2904 | AOID-PR | n.d. | p.Ser2830PhefsTer3 |
| 2911 | AOID-PR | p.Arg941His (rs547196696) | n.d. |
| 2914 | AOID-PR | n.d. | p.Arg843Ter (rs141263661), p.Asp668Tyr (rs765957331) |
| 2925 | AOID-PR | p.Arg3829His (rs145079750) | n.d. |
| 2926 | AOID-PR | p.Ser860Trp (rs201661720) | n.d. |
| 2935 | AOID-PR | p.Gly3903Ter | n.d. |
| 2938 | AOID-PR | n.d. | p.Arg843Ter (rs141263661) |
| 2951 | AOID-PR | p.Ala2919Val (rs533740963) | n.d. |
| 2877 | PP | n.d. | p.Gly1853Ser (rs750289349) |
| 2882 | PP | p.Arg1469His (rs145675213) | n.d. |
| 2886 | PP | p.Ala3750Val (rs769188915) | n.d. |
| 2895 | PP | n.d. | p.Glu1796Lys (rs748246449) |
| 2900 | PP | p.Gly2440Glu (rs555272052) | n.d. |
| 2905 | PP | n.d. | p.Thr380Arg (rs566122590) |
| 2907 | PP | p.Asp2136Asn (rs780793108) | n.d. |
| 2910 | PP | p.Ser1188Tyr (rs532746197) | n.d. |
| 2916 | PP | p.Lys679Ile (rs369325094) | n.d. |
| 2921 | PP | n.d. | p.Arg547Gln (rs769962080) |
| 2922 | PP | p.Ala974Val (rs143643121) | p.Gly1259Ser (rs200844006), p.Ser2346Thr (rs753301905) |
| 2923 | PP | p.Glu2133Asp (rs142983961) | n.d. |
| 2933 | PP | n.d. | p.Arg1834Gly (rs755669550) |
| 2937 | PP | n.d. | p.His286Arg (rs368737363) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Piccolo, L.; Wongkummool, W.; Jantaree, P.; Daroontum, T.; Chaowattanapanit, S.; Choonhakarn, C.; Amornpinyo, W.; Chaiwarith, R.; Kiratikanon, S.; Rujiwetpongstorn, R.; et al. Rare Filaggrin Variants Are Associated with Pustular Skin Diseases in Asians. Int. J. Mol. Sci. 2024, 25, 6466. https://doi.org/10.3390/ijms25126466
Lo Piccolo L, Wongkummool W, Jantaree P, Daroontum T, Chaowattanapanit S, Choonhakarn C, Amornpinyo W, Chaiwarith R, Kiratikanon S, Rujiwetpongstorn R, et al. Rare Filaggrin Variants Are Associated with Pustular Skin Diseases in Asians. International Journal of Molecular Sciences. 2024; 25(12):6466. https://doi.org/10.3390/ijms25126466
Chicago/Turabian StyleLo Piccolo, Luca, Wasinee Wongkummool, Phatcharida Jantaree, Teerada Daroontum, Suteeraporn Chaowattanapanit, Charoen Choonhakarn, Warayuwadee Amornpinyo, Romanee Chaiwarith, Salin Kiratikanon, Rujira Rujiwetpongstorn, and et al. 2024. "Rare Filaggrin Variants Are Associated with Pustular Skin Diseases in Asians" International Journal of Molecular Sciences 25, no. 12: 6466. https://doi.org/10.3390/ijms25126466
APA StyleLo Piccolo, L., Wongkummool, W., Jantaree, P., Daroontum, T., Chaowattanapanit, S., Choonhakarn, C., Amornpinyo, W., Chaiwarith, R., Kiratikanon, S., Rujiwetpongstorn, R., Tovanabutra, N., Chiewchanvit, S., Ngamphiw, C., Intachai, W., Kantaputra, P., & Chuamanochan, M. (2024). Rare Filaggrin Variants Are Associated with Pustular Skin Diseases in Asians. International Journal of Molecular Sciences, 25(12), 6466. https://doi.org/10.3390/ijms25126466