
Amelioration of Fibrosis via S1P Inhibition Is Regulated by Inactivation of TGF-β and SPL Pathways in the Human Cornea




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Latent Transforming Growth Factor Beta Binding Proteins (LTBPs)
2.2. Transforming Growth Factor Beta Receptors (TGF-βRs)
2.3. Canonical Downstream SMAD Pathway
2.4. Sphingolipid Members Pathway
2.5. Fibrosis Markers
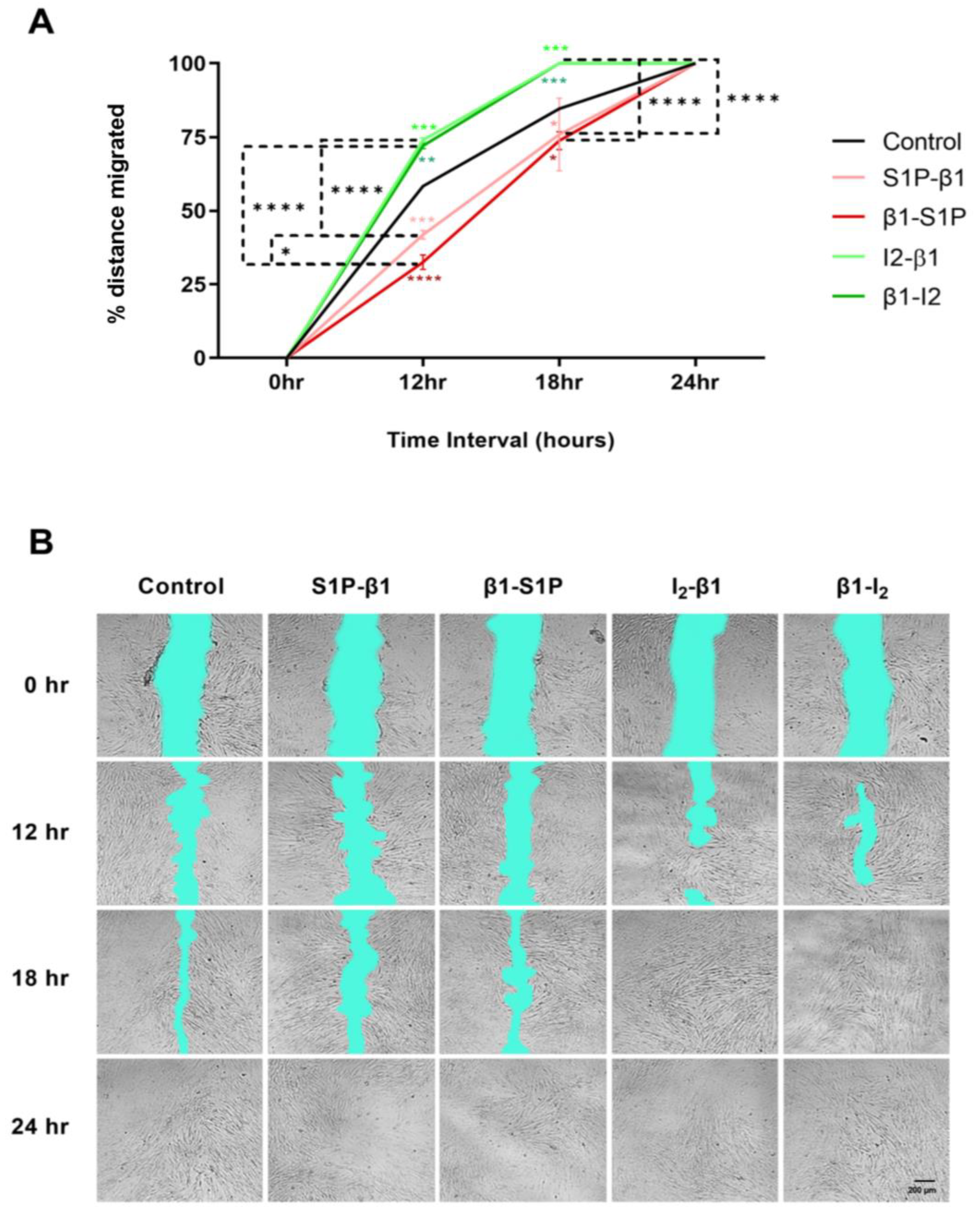
2.6. Effects of S1P and TGF-β Treatment Groups on HCF Cellular Migration
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Human Corneal Fibroblast Cell Isolation and 3D In Vitro Model Cultures
4.3. Western Blot Analysis
4.4. 2D Scratch Assay—Cell Migration
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barrientez, B.; Nicholas, S.E.; Whelchel, A.; Sharif, R.; Hjortdal, J.; Karamichos, D. Corneal injury: Clinical and molecular aspects. Exp. Eye Res. 2019, 186, 107709. [Google Scholar] [CrossRef]
- Yang, C.J.; Nguyen, D.D.; Lai, J.Y. Poly(l-Histidine)-Mediated On-Demand Therapeutic Delivery of Roughened Ceria Nanocages for Treatment of Chemical Eye Injury. Adv. Sci. 2023, 10, e2302174. [Google Scholar] [CrossRef]
- Funderburgh, J.L.; Funderburgh, M.L.; Mann, M.M.; Corpuz, L.; Roth, M.R. Proteoglycan expression during transforming growth factor beta -induced keratocyte-myofibroblast transdifferentiation. J. Biol. Chem. 2001, 276, 44173–44178. [Google Scholar] [CrossRef]
- Mohan, R.R.; Kempuraj, D.; D’Souza, S.; Ghosh, A. Corneal stromal repair and regeneration. Prog. Retin. Eye Res. 2022, 91, 101090. [Google Scholar] [CrossRef]
- Torricelli, A.A.; Wilson, S.E. Cellular and extracellular matrix modulation of corneal stromal opacity. Exp. Eye Res. 2014, 129, 151–160. [Google Scholar] [CrossRef]
- Pouw, A.E.; Greiner, M.A.; Coussa, R.G.; Jiao, C.; Han, I.C.; Skeie, J.M.; Fingert, J.H.; Mullins, R.F.; Sohn, E.H. Cell-Matrix Interactions in the Eye: From Cornea to Choroid. Cells 2021, 10, 687. [Google Scholar] [CrossRef]
- Noom, A.; Sawitzki, B.; Knaus, P.; Duda, G.N. A two-way street–cellular metabolism and myofibroblast contraction. NPJ Regen. Med. 2024, 9, 15. [Google Scholar] [CrossRef]
- Li, R.; Frangogiannis, N.G. Integrins in cardiac fibrosis. J. Mol. Cell Cardiol. 2022, 172, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Fan, W.; Link, F.; Wang, S.; Dooley, S. Transforming growth factor beta latency: A mechanism of cytokine storage and signalling regulation in liver homeostasis and disease. JHEP Rep. 2022, 4, 100397. [Google Scholar] [CrossRef]
- Ortiz-Zapater, E.; Signes-Costa, J.; Montero, P.; Roger, I. Lung Fibrosis and Fibrosis in the Lungs: Is It All about Myofibroblasts? Biomedicines 2022, 10, 1423. [Google Scholar] [CrossRef]
- Stolfi, C.; Troncone, E.; Marafini, I.; Monteleone, G. Role of TGF-Beta and Smad7 in Gut Inflammation, Fibrosis and Cancer. Biomolecules 2020, 11, 17. [Google Scholar] [CrossRef]
- Moretti, L.; Stalfort, J.; Barker, T.H.; Abebayehu, D. The interplay of fibroblasts, the extracellular matrix, and inflammation in scar formation. J. Biol. Chem. 2022, 298, 101530. [Google Scholar] [CrossRef]
- Gimeno, L.I.; Benito-Jardon, M.; Guerrero-Barbera, G.; Burday, N.; Costell, M. The Role of the Fibronectin Synergy Site for Skin Wound Healing. Cells 2022, 11, 2100. [Google Scholar] [CrossRef]
- Karamichos, D.; Hutcheon, A.E.; Zieske, J.D. Transforming growth factor-beta3 regulates assembly of a non-fibrotic matrix in a 3D corneal model. J. Tissue Eng. Regen. Med. 2011, 5, e228–e238. [Google Scholar] [CrossRef]
- Pot, S.A.; Lin, Z.; Shiu, J.; Benn, M.C.; Vogel, V. Growth factors and mechano-regulated reciprocal crosstalk with extracellular matrix tune the keratocyte-fibroblast/myofibroblast transition. Sci. Rep. 2023, 13, 11350. [Google Scholar] [CrossRef]
- Acosta, A.C.; Joud, H.; Sun, M.; Avila, M.Y.; Margo, C.E.; Espana, E.M. Keratocyte-Derived Myofibroblasts: Functional Differences With Their Fibroblast Precursors. Investig. Ophthalmol. Vis. Sci. 2023, 64, 9. [Google Scholar] [CrossRef]
- Guo, X.; Hutcheon, A.E.K.; Zieske, J.D. Molecular insights on the effect of TGF-beta1/-beta3 in human corneal fibroblasts. Exp. Eye Res. 2016, 146, 233–241. [Google Scholar] [CrossRef]
- Kamil, S.; Mohan, R.R. Corneal stromal wound healing: Major regulators and therapeutic targets. Ocul. Surf. 2021, 19, 290–306. [Google Scholar] [CrossRef]
- Karamichos, D.; Hutcheon, A.E.; Zieske, J.D. Reversal of fibrosis by TGF-beta3 in a 3D in vitro model. Exp. Eye Res. 2014, 124, 31–36. [Google Scholar] [CrossRef]
- Meng, N.; Wu, J.; Chen, J.; Luo, Y.; Xu, L.; Li, X. Basement membrane regeneration and TGF-beta1 expression in rabbits with corneal perforating injury. Mol. Vis. 2023, 29, 58–67. [Google Scholar]
- Shah, M.; Foreman, D.M.; Ferguson, M.W. Neutralisation of TGF-beta 1 and TGF-beta 2 or exogenous addition of TGF-beta 3 to cutaneous rat wounds reduces scarring. J. Cell Sci. 1995, 108 Pt 3, 985–1002. [Google Scholar] [CrossRef]
- Tsai, C.H.; Liu, E.; Phan, A.; Lu, K.L.; Mei, H. NBL1 Reduces Corneal Fibrosis and Scar Formation after Wounding. Biomolecules 2023, 13, 1570. [Google Scholar] [CrossRef]
- Wilson, S.E. Corneal myofibroblasts and fibrosis. Exp. Eye Res. 2020, 201, 108272. [Google Scholar] [CrossRef]
- Yeung, V.; Sriram, S.; Tran, J.A.; Guo, X.; Hutcheon, A.E.K.; Zieske, J.D.; Karamichos, D.; Ciolino, J.B. FAK Inhibition Attenuates Corneal Fibroblast Differentiation In Vitro. Biomolecules 2021, 11, 1682. [Google Scholar] [CrossRef]
- Vander Ark, A.; Cao, J.; Li, X. TGF-beta receptors: In and beyond TGF-beta signaling. Cell Signal 2018, 52, 112–120. [Google Scholar] [CrossRef]
- Nicholas, S.E.; Choi, A.J.; Lam, T.N.; Basu, S.K.; Mandal, N.; Karamichos, D. Potentiation of Sphingolipids and TGF-beta in the human corneal stroma reveals intricate signaling pathway crosstalks. Exp. Eye Res. 2023, 231, 109487. [Google Scholar] [CrossRef]
- Priyadarsini, S.; McKay, T.B.; Sarker-Nag, A.; Karamichos, D. Keratoconus in vitro and the key players of the TGF-beta pathway. Mol. Vis. 2015, 21, 577–588. [Google Scholar]
- Klingberg, F.; Hinz, B.; White, E.S. The myofibroblast matrix: Implications for tissue repair and fibrosis. J. Pathol. 2013, 229, 298–309. [Google Scholar] [CrossRef]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef]
- Rifkin, D.; Sachan, N.; Singh, K.; Sauber, E.; Tellides, G.; Ramirez, F. The role of LTBPs in TGF beta signaling. Dev. Dyn. 2022, 251, 95–104. [Google Scholar] [CrossRef]
- Robertson, I.B.; Horiguchi, M.; Zilberberg, L.; Dabovic, B.; Hadjiolova, K.; Rifkin, D.B. Latent TGF-beta-binding proteins. Matrix Biol. 2015, 47, 44–53. [Google Scholar] [CrossRef]
- Shea, B.S.; Tager, A.M. Sphingolipid regulation of tissue fibrosis. Open Rheumatol. J. 2012, 6, 123–129. [Google Scholar] [CrossRef]
- Swaney, J.S.; Moreno, K.M.; Gentile, A.M.; Sabbadini, R.A.; Stoller, G.L. Sphingosine-1-phosphate (S1P) is a novel fibrotic mediator in the eye. Exp. Eye Res. 2008, 87, 367–375. [Google Scholar] [CrossRef]
- Takuwa, Y.; Ikeda, H.; Okamoto, Y.; Takuwa, N.; Yoshioka, K. Sphingosine-1-phosphate as a mediator involved in development of fibrotic diseases. Biochim. Biophys. Acta 2013, 1831, 185–192. [Google Scholar] [CrossRef]
- Watterson, K.R.; Lanning, D.A.; Diegelmann, R.F.; Spiegel, S. Regulation of fibroblast functions by lysophospholipid mediators: Potential roles in wound healing. Wound Repair. Regen. 2007, 15, 607–616. [Google Scholar] [CrossRef]
- Borodzicz, S.; Rudnicka, L.; Mirowska-Guzel, D.; Cudnoch-Jedrzejewska, A. The role of epidermal sphingolipids in dermatologic diseases. Lipids Health Dis. 2016, 15, 13. [Google Scholar] [CrossRef]
- Masuda-Kuroki, K.; Di Nardo, A. Sphingosine 1-Phosphate Signaling at the Skin Barrier Interface. Biology 2022, 11, 809. [Google Scholar] [CrossRef]
- Park, W.J.; Song, J.H.; Kim, G.T.; Park, T.S. Ceramide and Sphingosine 1-Phosphate in Liver Diseases. Mol. Cells 2020, 43, 419–430. [Google Scholar] [CrossRef]
- Ishay, Y.; Nachman, D.; Khoury, T.; Ilan, Y. The role of the sphingolipid pathway in liver fibrosis: An emerging new potential target for novel therapies. Am. J. Physiol. Cell Physiol. 2020, 318, C1055–C1064. [Google Scholar] [CrossRef]
- Choi, R.H.; Tatum, S.M.; Symons, J.D.; Summers, S.A.; Holland, W.L. Ceramides and other sphingolipids as drivers of cardiovascular disease. Nat. Rev. Cardiol. 2021, 18, 701–711. [Google Scholar] [CrossRef]
- Cirillo, F.; Piccoli, M.; Ghiroldi, A.; Monasky, M.M.; Rota, P.; La Rocca, P.; Tarantino, A.; D’Imperio, S.; Signorelli, P.; Pappone, C.; et al. The antithetic role of ceramide and sphingosine-1-phosphate in cardiac dysfunction. J. Cell Physiol. 2021, 236, 4857–4873. [Google Scholar] [CrossRef]
- Sharma, L.; Prakash, H. Sphingolipids Are Dual Specific Drug Targets for the Management of Pulmonary Infections: Perspective. Front. Immunol. 2017, 8, 378. [Google Scholar] [CrossRef]
- Tibboel, J.; Reiss, I.; de Jongste, J.C.; Post, M. Sphingolipids in lung growth and repair. Chest 2014, 145, 120–128. [Google Scholar] [CrossRef]
- Jayant, G.; Kuperberg, S.; Somnay, K.; Wadgaonkar, R. The Role of Sphingolipids in Regulating Vascular Permeability in Idiopathic Pulmonary Fibrosis. Biomedicines 2023, 11, 1728. [Google Scholar] [CrossRef]
- Wang, N.; Li, J.Y.; Zeng, B.; Chen, G.L. Sphingosine-1-Phosphate Signaling in Cardiovascular Diseases. Biomolecules 2023, 13, 818. [Google Scholar] [CrossRef]
- Nojima, H.; Shimizu, H.; Murakami, T.; Shuto, K.; Koda, K. Critical Roles of the Sphingolipid Metabolic Pathway in Liver Regeneration, Hepatocellular Carcinoma Progression and Therapy. Cancers 2024, 16, 850. [Google Scholar] [CrossRef] [PubMed]
- Ueda, N. A Rheostat of Ceramide and Sphingosine-1-Phosphate as a Determinant of Oxidative Stress-Mediated Kidney Injury. Int. J. Mol. Sci. 2022, 23, 4010. [Google Scholar] [CrossRef]
- Kleuser, B.; Baumer, W. Sphingosine 1-Phosphate as Essential Signaling Molecule in Inflammatory Skin Diseases. Int. J. Mol. Sci. 2023, 24, 1456. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Sarker-Nag, A.; Allegood, J.; Chalfant, C.; Karamichos, D. Description of the sphingolipid content and subspecies in the diabetic cornea. Curr. Eye Res. 2015, 40, 1204–1210. [Google Scholar] [CrossRef]
- Qi, H.; Priyadarsini, S.; Nicholas, S.E.; Sarker-Nag, A.; Allegood, J.; Chalfant, C.E.; Mandal, N.A.; Karamichos, D. Analysis of sphingolipids in human corneal fibroblasts from normal and keratoconus patients. J. Lipid Res. 2017, 58, 636–648. [Google Scholar] [CrossRef]
- Nicholas, S.E.; Rowsey, T.G.; Priyadarsini, S.; Mandal, N.A.; Karamichos, D. Unravelling the interplay of sphingolipids and TGF-beta signaling in the human corneal stroma. PLoS ONE 2017, 12, e0182390. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Watsky, M.A. LPA and S1P increase corneal epithelial and endothelial cell transcellular resistance. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1927–1933. [Google Scholar] [CrossRef]
- Wang, D.A.; Du, H.; Jaggar, J.H.; Brindley, D.N.; Tigyi, G.J.; Watsky, M.A. Injury-elicited differential transcriptional regulation of phospholipid growth factor receptors in the cornea. Am. J. Physiol. Cell Physiol. 2002, 283, C1646–C1654. [Google Scholar] [CrossRef]
- Wang, J.; Carbone, L.D.; Watsky, M.A. Receptor-mediated activation of a Cl(-) current by LPA and S1P in cultured corneal keratocytes. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3202–3208. [Google Scholar]
- Wilkerson, J.L.; Basu, S.K.; Stiles, M.A.; Prislovsky, A.; Grambergs, R.C.; Nicholas, S.E.; Karamichos, D.; Allegood, J.C.; Proia, R.L.; Mandal, N. Ablation of Sphingosine Kinase 1 Protects Cornea from Neovascularization in a Mouse Corneal Injury Model. Cells 2022, 11, 2914. [Google Scholar] [CrossRef]
- Basu, S.K.; Mandal, N. Image-Based Longitudinal Characterization of Corneal Wound to Understand the Role of Sphingosine-1-Phosphate. Methods Mol. Biol. 2023, 2625, 337–345. [Google Scholar] [CrossRef]
- Garcia Garcia, J.M.; Vannuzzi, V.; Donati, C.; Bernacchioni, C.; Bruni, P.; Petraglia, F. Endometriosis: Cellular and Molecular Mechanisms Leading to Fibrosis. Reprod. Sci. 2023, 30, 1453–1461. [Google Scholar] [CrossRef]
- Li, J.; Xiao, Y.; Zhang, Y.; Li, S.; Zhao, M.; Xia, T.; Meng, H. Pulmonary Delivery of Specialized Pro-Resolving Mediators-Based Nanotherapeutics Attenuates Pulmonary Fibrosis in Preclinical Animal Models. ACS Nano 2023, 17, 15354–15370. [Google Scholar] [CrossRef]
- Riemma, M.A.; Cerqua, I.; Romano, B.; Irollo, E.; Bertolino, A.; Camerlingo, R.; Granato, E.; Rea, G.; Scala, S.; Terlizzi, M.; et al. Sphingosine-1-phosphate/TGF-beta axis drives epithelial mesenchymal transition in asthma-like disease. Br. J. Pharmacol. 2022, 179, 1753–1768. [Google Scholar] [CrossRef]
- Lin, Q.; Long, C.; Wang, Z.; Wang, R.; Shi, W.; Qiu, J.; Mo, J.; Xie, Y. Hirudin, a thrombin inhibitor, attenuates TGF-beta-induced fibrosis in renal proximal tubular epithelial cells by inhibition of protease-activated receptor 1 expression via S1P/S1PR2/S1PR3 signaling. Exp. Ther. Med. 2022, 23, 3. [Google Scholar] [CrossRef]
- Cui, L.; Wang, X.; Zhao, X.; Sun, B.; Xia, T.; Hu, S. CeO(2) nanoparticles induce pulmonary fibrosis via activating S1P pathway as revealed by metabolomics. Nano Today 2022, 45, 101559. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Uribe, A.P.; Gomez-Sierra, T.; Aparicio-Trejo, O.E.; Orozco-Ibarra, M.; Pedraza-Chaverri, J. Backstage players of fibrosis: NOX4, mTOR, HDAC, and S1P; companions of TGF-beta. Cell Signal 2021, 87, 110123. [Google Scholar] [CrossRef] [PubMed]
- Gellings Lowe, N.; Swaney, J.S.; Moreno, K.M.; Sabbadini, R.A. Sphingosine-1-phosphate and sphingosine kinase are critical for transforming growth factor-beta-stimulated collagen production by cardiac fibroblasts. Cardiovasc. Res. 2009, 82, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Lebman, D.A.; Spiegel, S. Cross-talk at the crossroads of sphingosine-1-phosphate, growth factors, and cytokine signaling. J. Lipid Res. 2008, 49, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Donati, C.; Cencetti, F.; Bernacchioni, C.; Vannuzzi, V.; Bruni, P. Role of sphingosine 1-phosphate signalling in tissue fibrosis. Cell Signal 2021, 78, 109861. [Google Scholar] [CrossRef]
- Wilson, S.E. TGF beta-1, -2 and -3 in the modulation of fibrosis in the cornea and other organs. Exp. Eye Res. 2021, 207, 108594. [Google Scholar] [CrossRef] [PubMed]
- Donovan, C.; Cogswell, D.; Sun, M.; Adams, S.; Avila, M.Y.; Margo, C.E.; Koch, M.; Espana, E.M. Collagen XII regulates stromal wound closure. Exp. Eye Res. 2023, 230, 109456. [Google Scholar] [CrossRef]
- Guo, D.; Li, S.; Xiao, X.; Jiang, Y.; Wang, Y.; Jin, G.; Wang, J.; Ouyang, J.; Jia, X.; Sun, W.; et al. Clinical and Genetic Landscape of Ectopia Lentis Based on a Cohort of Patients From 156 Families. Investig. Ophthalmol. Vis. Sci. 2024, 65, 20. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, T.; Gan, J.; Mao, Y.; Zhao, L.; Jiao, X.; Fan, M.; Wang, T.; Zhang, D.; Xu, M.; et al. Zonule-Associated Gene Variants in Isolated Ectopia Lentis and Glaucoma. J. Glaucoma 2023, 32, e80–e89. [Google Scholar] [CrossRef]
- Lewis, C.J.; Hedberg-Buenz, A.; DeLuca, A.P.; Stone, E.M.; Alward, W.L.M.; Fingert, J.H. Primary congenital and developmental glaucomas. Hum. Mol. Genet. 2017, 26, R28–R36. [Google Scholar] [CrossRef]
- Gupta, V.; Birla, S.; Varshney, T.; Somarajan, B.I.; Gupta, S.; Gupta, M.; Panigrahi, A.; Singh, A.; Gupta, D. In vivo identification of angle dysgenesis and its relation to genetic markers associated with glaucoma using artificial intelligence. Indian J. Ophthalmol. 2024, 72, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Bergman, Z.; Anderson, K.; Kuchtey, R.W. Compound Heterozygous LTBP2 Mutations Associated With Juvenile-Onset Open-Angle Glaucoma and Marfan-Like Phenotype. JAMA Ophthalmol. 2023, 141, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Duvvari, M.R.; Prabhakaran, V.C.; Shetty, J.S.; Murthy, G.J.; Blanton, S.H. A homozygous mutation in LTBP2 causes isolated microspherophakia. Hum. Genet. 2010, 128, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; McKibbin, M.; Booth, A.; Parry, D.A.; Jain, P.; Riazuddin, S.A.; Hejtmancik, J.F.; Khan, S.N.; Firasat, S.; Shires, M.; et al. Null mutations in LTBP2 cause primary congenital glaucoma. Am. J. Hum. Genet. 2009, 84, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Narooie-Nejad, M.; Paylakhi, S.H.; Shojaee, S.; Fazlali, Z.; Rezaei Kanavi, M.; Nilforushan, N.; Yazdani, S.; Babrzadeh, F.; Suri, F.; Ronaghi, M.; et al. Loss of function mutations in the gene encoding latent transforming growth factor beta binding protein 2, LTBP2, cause primary congenital glaucoma. Hum. Mol. Genet. 2009, 18, 3969–3977. [Google Scholar] [CrossRef] [PubMed]
- Azmanov, D.N.; Dimitrova, S.; Florez, L.; Cherninkova, S.; Draganov, D.; Morar, B.; Saat, R.; Juan, M.; Arostegui, J.I.; Ganguly, S.; et al. LTBP2 and CYP1B1 mutations and associated ocular phenotypes in the Roma/Gypsy founder population. Eur. J. Hum. Genet. 2011, 19, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Ohbayashi, T.; Fujikawa, Y.; Yoshida, H.; Akama, T.O.; Noda, K.; Horiguchi, M.; Kameyama, K.; Hata, Y.; Takahashi, K.; et al. Latent TGF-beta binding protein-2 is essential for the development of ciliary zonule microfibrils. Hum. Mol. Genet. 2014, 23, 5672–5682. [Google Scholar] [CrossRef] [PubMed]
- Schlotzer-Schrehardt, U.; Zenkel, M.; Kuchle, M.; Sakai, L.Y.; Naumann, G.O. Role of transforming growth factor-beta1 and its latent form binding protein in pseudoexfoliation syndrome. Exp. Eye Res. 2001, 73, 765–780. [Google Scholar] [CrossRef] [PubMed]
- Zenkel, M.; Poschl, E.; von der Mark, K.; Hofmann-Rummelt, C.; Naumann, G.O.; Kruse, F.E.; Schlotzer-Schrehardt, U. Differential gene expression in pseudoexfoliation syndrome. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3742–3752. [Google Scholar] [CrossRef]
- De Maria, A.; Zientek, K.D.; David, L.L.; Wilmarth, P.A.; Bhorade, A.M.; Harocopos, G.J.; Huang, A.J.W.; Hong, A.R.; Siegfried, C.J.; Tsai, L.M.; et al. Compositional Analysis of Extracellular Aggregates in the Eyes of Patients With Exfoliation Syndrome and Exfoliation Glaucoma. Investig. Ophthalmol. Vis. Sci. 2021, 62, 27. [Google Scholar] [CrossRef]
- Su, C.T.; Urban, Z. LTBP4 in Health and Disease. Genes 2021, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, Y.; Yoshida, H.; Inoue, T.; Ohbayashi, T.; Noda, K.; von Melchner, H.; Iwasaka, T.; Shiojima, I.; Akama, T.O.; Nakamura, T. Latent TGF-beta binding protein 2 and 4 have essential overlapping functions in microfibril development. Sci. Rep. 2017, 7, 43714. [Google Scholar] [CrossRef]
- Ravel, J.M.; Comel, M.; Wandzel, M.; Bronner, M.; Tatopoulos, A.; Renaud, M.; Lambert, L.; Bursztejn, A.C.; Bonnet, C. First report of a short in-frame biallelic deletion removing part of the EGF-like domain calcium-binding motif in LTBP4 and causing autosomal recessive cutis laxa type 1C. Am. J. Med. Genet. A 2022, 188, 3343–3349. [Google Scholar] [CrossRef]
- Callewaert, B.L.; Urban, Z. LTBP4-Related Cutis Laxa. In GeneReviews®; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Mazaheri, M.; Jahantigh, H.R.; Yavari, M.; Mirjalili, S.R.; Vahidnezhad, H. Autosomal recessive cutis laxa type 1C with a homozygous LTBP4 splicing variant: A case report and update of literature. Mol. Biol. Rep. 2022, 49, 4135–4140. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Q.; Wang, L.; Tu, W.; Chu, H.; Ding, W.; Jiang, S.; Ma, Y.; Shi, X.; Pu, W.; et al. Increased expression of latent TGF-beta-binding protein 4 affects the fibrotic process in scleroderma by TGF-beta/SMAD signaling. Lab. Investig. 2017, 97, 1121. [Google Scholar] [CrossRef]
- Park, H.J.; Choi, Y.J.; Park, C.H.; Kim, T.H.; Lee, S.S.; Moon, D.H.; Lee, K.A.; Lee, S.E.; Park, M.S.; Kim, S.Y.; et al. Outstanding Characteristics of Birt-Hogg-Dube Syndrome in Korea. Diagnostics 2023, 13, 2047. [Google Scholar] [CrossRef]
- Koloko Ngassie, M.L.; De Vries, M.; Borghuis, T.; Timens, W.; Sin, D.D.; Nickle, D.; Joubert, P.; Horvatovich, P.; Marko-Varga, G.; Teske, J.J.; et al. Age-associated differences in the human lung extracellular matrix. Am. J. Physiol. Lung Cell Mol. Physiol. 2023, 324, L799–L814. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Tomita, B.; Salinger, A.; Tilvawala, R.R.; Li, L.; Hakami, H.; Liu, T.; Tsoyi, K.; Rosas, I.O.; Reinhardt, D.P.; et al. PAD2-mediated citrullination of Fibulin-5 promotes elastogenesis. Matrix Biol. 2021, 102, 70–84. [Google Scholar] [CrossRef]
- Liu, Y.N.; Lv, X.; Chen, X.; Yan, M.; Guo, L.C.; Liu, G.; Yao, L.; Jiang, H.F. Specific Overexpression of YAP in Vascular Smooth Muscle Attenuated Abdominal Aortic Aneurysm Formation by Activating Elastic Fiber Assembly via LTBP4. J. Cardiovasc. Transl. Res. 2023, 16, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, X.; Tian, X.; Cao, Z.; Zhen, X.; Zhao, W.; Luo, B.; Gao, Y. Novel indel variation of LTBP4 gene associates with risk of sudden cardiac death in Chinese populations with coronary artery disease. Leg. Med. 2024, 69, 102437. [Google Scholar] [CrossRef] [PubMed]
- Bello, L.; Sabbatini, D.; Fusto, A.; Gorgoglione, D.; Borin, G.U.; Penzo, M.; Riguzzi, P.; Villa, M.; Vianello, S.; Calore, C.; et al. The IAAM LTBP4 Haplotype is Protective Against Dystrophin-Deficient Cardiomyopathy. J. Neuromuscul. Dis. 2024, 11, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, D.; Wu, T.; Sun, F. Disruption of LTBP4 Inhibition-Induced TGFbeta1 Activation Promoted Cell Proliferation and Metastasis in Skin Melanoma by Inhibiting the Activation of the Hippo-YAP1 Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 673904. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.N.; Moon, S.J.; Jung, K.C.; Kim, S.W.; Kim, H.; Han, D.; Kim, J.H. Mass Spectrometry-Based Proteomic Discovery of Prognostic Biomarkers in Adrenal Cortical Carcinoma. Cancers 2021, 13, 3890. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; He, X.; Zeng, M. The Role of S1P and the Related Signaling Pathway in the Development of Tissue Fibrosis. Front. Pharmacol. 2018, 9, 1504. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.M.; Cian, A.A.; Weng, T.H.; Liang, C.M.; Pao, S.I.; Chen, Y.J. Beneficial Effects of Hypercapnic Acidosis on the Inhibition of Transforming Growth Factor beta-1-induced Corneal Fibrosis in Vitro. Curr. Eye Res. 2021, 46, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.R.; Tripathi, R.; Balne, P.K.; Suleiman, L.; Simkins, K.; Chaurasia, S.S.; Mohan, R.R. Mustard Gas Exposure Actuates SMAD2/3 Signaling to Promote Myofibroblast Generation in the Cornea. Cells 2023, 12, 1533. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, R.; Zhang, X.; Guo, Z.; Geng, X.; Chu, D.; Cui, H.; Zhang, A.; Li, W.; Zhu, L.; et al. An injectable thermoresponsive-hydrogel for lamellar keratoplasty: In-situ releases celastrol and hampers corneal scars. J. Control Release 2024, 369, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, L.; Liu, Z.; Tang, J.; Zhang, L.; Li, Z.; Zhou, D.; Sun, Z.; Xi, K.; Lu, P.; et al. A responsive hydrogel modulates innate immune cascade fibrosis to promote ocular surface reconstruction after chemical injury. J. Control Release 2024, 365, 1124–1138. [Google Scholar] [CrossRef]
- Nuwormegbe, S.; Park, N.Y.; Kim, S.W. Lobeglitazone attenuates fibrosis in corneal fibroblasts by interrupting TGF-beta-mediated Smad signaling. Graefes Arch. Clin. Exp. Ophthalmol. 2022, 260, 149–162. [Google Scholar] [CrossRef]
- Tang, Y.; Du, E.; Wang, G.; Qin, F.; Meng, Z.; Dai, L.; Wang, Y.; Ren, S. A negative feedback loop centered on SMAD3 expression in transforming growth factor beta1-induced corneal myofibroblast differentiation. Exp. Eye Res. 2023, 236, 109654. [Google Scholar] [CrossRef]
- Kanemitsu, E.; Zhao, X.; Iwaisako, K.; Inoue, A.; Takeuchi, A.; Yagi, S.; Masumoto, H.; Ohara, H.; Hosokawa, M.; Awaya, T.; et al. Antagonist of sphingosine 1-phosphate receptor 3 reduces cold injury of rat donor hearts for transplantation. Transl. Res. 2023, 255, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Bernacchioni, C.; Rossi, M.; Vannuzzi, V.; Prisinzano, M.; Seidita, I.; Raeispour, M.; Muccilli, A.; Castiglione, F.; Bruni, P.; Petraglia, F.; et al. Sphingosine-1-phosphate receptor 3 is a non-hormonal target to counteract endometriosis-associated fibrosis. Fertil. Steril. 2024, 121, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Sumioka, T.; Iwanishi, H.; Okada, Y.; Miyajima, M.; Ichikawa, K.; Reinach, P.S.; Saika, S. Loss of sphingosine 1-phosphate receptor 3 gene function impairs injury-induced stromal angiogenesis in mouse cornea. Lab. Investig. 2021, 101, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.V.; Vera, M.S.; Tenconi, P.E.; Soto, T.; Prado Spalm, F.H.; Torlaschi, C.; Mateos, M.V.; Rotstein, N.P. Sphingosine-1-phosphate and ceramide-1-phosphate promote migration, pro-inflammatory and pro-fibrotic responses in retinal pigment epithelium cells. Exp. Eye Res. 2022, 224, 109222. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kwon, S.; Jeon, S.; Jung, B.J.; Kim, K.S. Sphingosine-1-phosphate expression in human epiretinal membranes. PLoS ONE 2022, 17, e0273674. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Tang, H.; Zhou, Q.; Li, H.; Song, E. FTY720 Suppresses Pathogenic Retinal Muller Cell Activation and Chronic Progression by Inhibiting the mTOR/NF-kappaB Signaling Pathway and Regulating Autophagy. Curr. Eye Res. 2024, 1–10. [Google Scholar] [CrossRef]
- Karamichos, D.; Guo, X.Q.; Hutcheon, A.E.; Zieske, J.D. Human corneal fibrosis: An in vitro model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1382–1388. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicholas, S.E.; Basu, S.K.; Mandal, N.; Karamichos, D. Amelioration of Fibrosis via S1P Inhibition Is Regulated by Inactivation of TGF-β and SPL Pathways in the Human Cornea. Int. J. Mol. Sci. 2024, 25, 6560. https://doi.org/10.3390/ijms25126560
Nicholas SE, Basu SK, Mandal N, Karamichos D. Amelioration of Fibrosis via S1P Inhibition Is Regulated by Inactivation of TGF-β and SPL Pathways in the Human Cornea. International Journal of Molecular Sciences. 2024; 25(12):6560. https://doi.org/10.3390/ijms25126560
Chicago/Turabian StyleNicholas, Sarah E., Sandip K. Basu, Nawajes Mandal, and Dimitrios Karamichos. 2024. "Amelioration of Fibrosis via S1P Inhibition Is Regulated by Inactivation of TGF-β and SPL Pathways in the Human Cornea" International Journal of Molecular Sciences 25, no. 12: 6560. https://doi.org/10.3390/ijms25126560
APA StyleNicholas, S. E., Basu, S. K., Mandal, N., & Karamichos, D. (2024). Amelioration of Fibrosis via S1P Inhibition Is Regulated by Inactivation of TGF-β and SPL Pathways in the Human Cornea. International Journal of Molecular Sciences, 25(12), 6560. https://doi.org/10.3390/ijms25126560