

Supplementation of a High-Fat Diet with Pentadecylresorcinol Increases the Representation of Akkermansia muciniphila in the Mouse Small and Large Intestines and May Protect against Complications Caused by Imbalanced Nutrition
 , ,
, ,
Abstract
:1. Introduction
2. Results
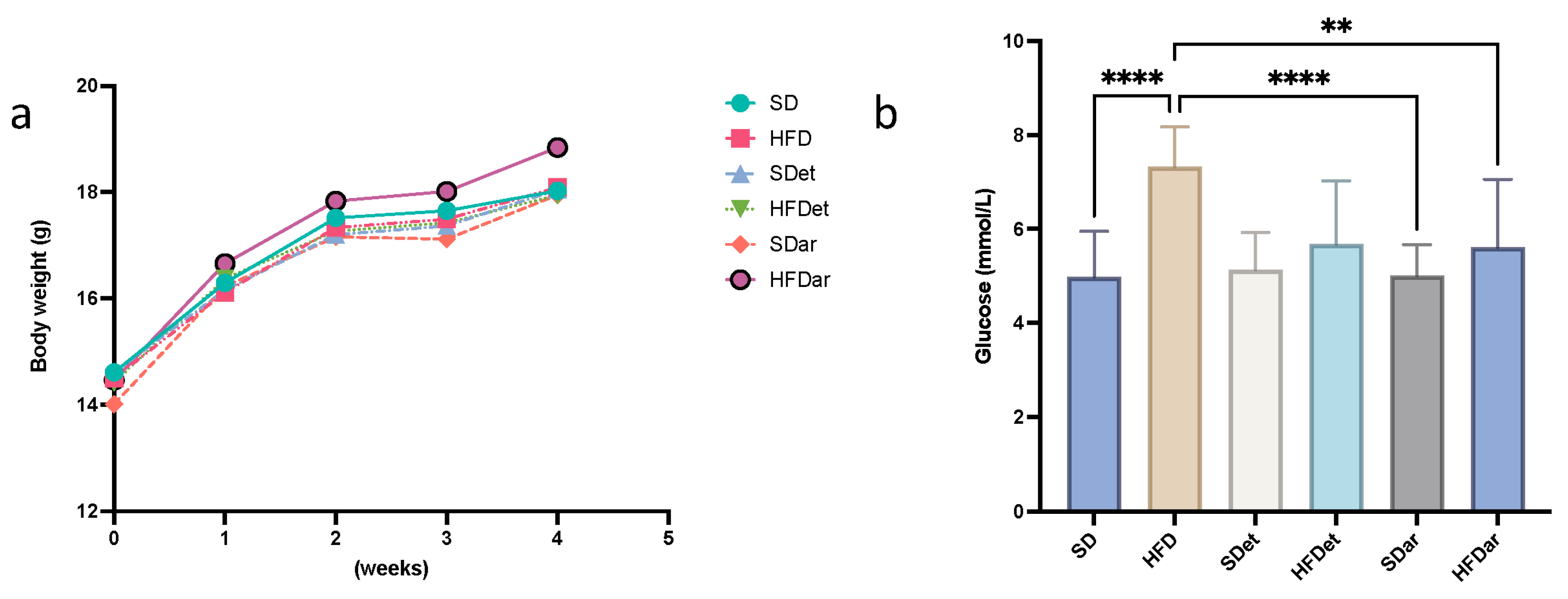
2.1. Body Weight (BW) and Food and Water Intake
2.2. Fasting Blood Glucose (FBG), Serum Triacylglyceride (TAG), and Cholesterol Levels
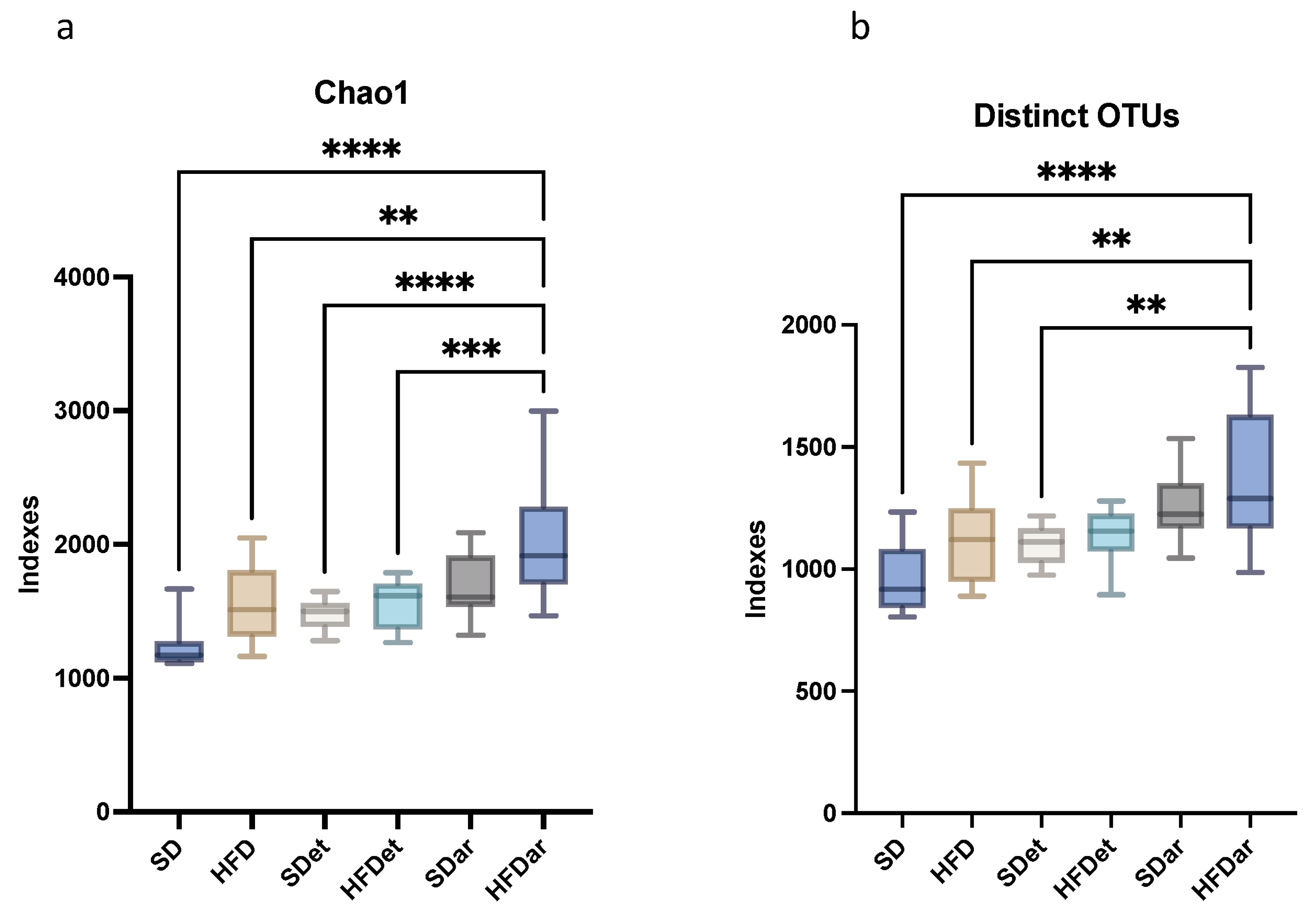
2.3. Composition of the Gut Microbiota in the Mouse Small and Large Intestines
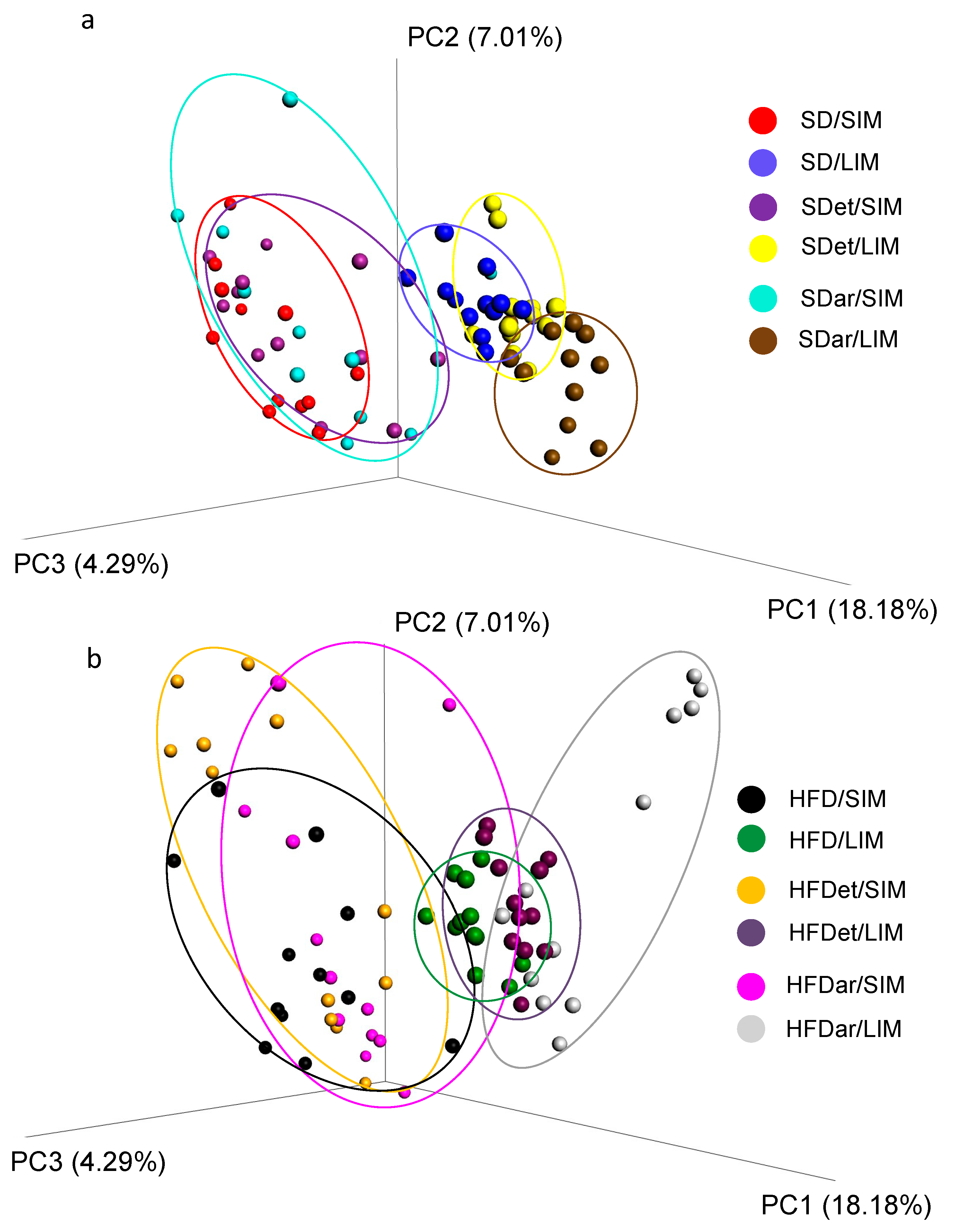
2.4. Composition of the Microbial Community in the Mouse Small and Large Intestines
2.5. Reconstruction of the Metabolic Activity of the Mouse Gut Microbiota
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Study Design
4.2. Body Weight (BW), Food and Water Intake, Fasting Blood Glucose (FBG), Serum Triglyceride (TG), and Cholesterol Levels Were Measured
4.3. Large Intestinal and Small Intestinal Microbiota Sampling for Metagenome Analysis
4.4. High-Throughput Sequencing Analysis and Reconstruction of Intestinal Microbiota Metabolic Activity
4.5. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martinez-Lomeli, J.; Deol, P.; Deans, J.R.; Jiang, T.; Ruegger, P.; Borneman, J.; Sladek, F.M. Impact of Various High Fat Diets on Gene Expression and the Microbiome across the Mouse Intestines. Sci. Rep. 2023, 13, 22758. [Google Scholar] [CrossRef] [PubMed]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells. 2021, 11, 3164. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.; Gholami, A.M.; Berry, D.; Desmarchelier, C.; Hahne, H.; Loh, G.; Mondot, S.; Lepage, P.; Rothballer, M.; Walker, A.; et al. High-Fat Diet Alters Gut Microbiota Physiology in Mice. ISME J. 2014, 8, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill-Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716–1724.e2. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Chi, M.M.; Scull, B.P.; Rigby, R.; Schwerbrock, N.M.J.; Magness, S.; Jobin, C.; Lund, P.K. High-Fat Diet: Bacteria Interactions Promote Intestinal Inflammation Which Precedes and Correlates with Obesity and Insulin Resistance in Mouse. PLoS ONE 2010, 5, e12191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Pan, Q.; Shen, F.; Cao, H.; Ding, W.; Chen, Y.; Fan, J. Total Fecal Microbiota Transplantation Alleviates High-Fat Diet-Induced Steatohepatitis in Mice via Beneficial Regulation of Gut Microbiota. Sci. Rep. 2017, 7, 1529. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Vogt, J.K.; Kristensen, M.; Hansen, L.B.S.; Ibrügger, S.; Mærkedahl, R.B.; Bahl, M.I.; Lind, M.V.; Nielsen, R.L.; Frøkiær, H.; et al. Whole Grain-Rich Diet Reduces Body Weight and Systemic Low-Grade Inflammation without Inducing Major Changes of the Gut Microbiome: A Randomised Cross-over Trial. Gut 2019, 68, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Malin, S.K.; Kullman, E.L.; Scelsi, A.R.; Haus, J.M.; Filion, J.; Pagadala, M.R.; Godin, J.-P.; Kochhar, S.; Ross, A.B.; Kirwan, J.P. A Whole-Grain Diet Reduces Peripheral Insulin Resistance and Improves Glucose Kinetics in Obese Adults: A Randomized-Controlled Trial. Metabolism 2018, 82, 111–117. [Google Scholar] [CrossRef]
- Guo, H.; Ding, J.; Liang, J.; Zhang, Y. Associations of Whole Grain and Refined Grain Consumption With Metabolic Syndrome. A Meta-Analysis of Observational Studies. Front. Nutr. 2021, 8, 695620. [Google Scholar] [CrossRef]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the Human Gut Microbiota by Phenolics and Phenolic Fiber-rich Foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1268–1298. [Google Scholar] [CrossRef]
- Prasadi V. P., N.; Joye, I.J. Dietary Fibre from Whole Grains and Their Benefits on Metabolic Health. Nutrients 2020, 12, 3045. [Google Scholar] [CrossRef]
- Pedrazzani, C.; Vanara, F.; Bhandari, D.R.; Bruni, R.; Spengler, B.; Blandino, M.; Righetti, L. 5-n-Alkylresorcinol Profiles in Different Cultivars of Einkorn, Emmer, Spelt, Common Wheat, and Tritordeum. J. Agric. Food Chem. 2021, 69, 14092–14102. [Google Scholar] [CrossRef] [PubMed]
- El-Shabasy, R.M.; Farag, M.A. Dissecting Dietary Alkylresorcinols: A Compile of Their Distribution, Biosynthesis, Extraction and Functional Properties. Crit. Rev. Biotechnol. 2023, 44, 581–617. [Google Scholar] [CrossRef]
- Ross, A.B. Alkylresorcinols. In Whole Grains and Their Bioactives; Wiley: Hoboken, NJ, USA, 2019; pp. 393–406. [Google Scholar]
- Liu, J.; Zhang, D.; Yang, Z.; Hao, Y.; Wang, Z.; Wang, J.; Wang, Z. Wheat Alkylresorcinols Modulate Glucose Homeostasis through Improving GLP-1 Secretion in High-Fat-Diet-Induced Obese Mice. J. Agric. Food Chem. 2023, 71, 16125–16136. [Google Scholar] [CrossRef] [PubMed]
- Reshma, A.; Tamilanban, T.; Chitra, V.; Subramaniyan, V.; Gupta, G.; Fuloria, N.K.; Sekar, M.; Fuloria, S.; Sahu, R.; Narayanan, J.; et al. Anti-Obesity Effects of Olivetol in Adult Zebrafish Model Induced by Short-Term High-Fat Diet. Sci. Rep. 2023, 13, 18449. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Yang, H.; Zhang, D.; Zhang, Y.; Wang, J.; Liu, J. 5-Heptadecylresorcinol Ameliorates Obesity-Associated Skeletal Muscle Mitochondrial Dysfunction through SIRT3-Mediated Mitophagy. J. Agric. Food Chem. 2023, 71, 16032–16042. [Google Scholar] [CrossRef] [PubMed]
- Zabolotneva, A.A.; Gaponov, A.M.; Roumiantsev, S.A.; Vasiliev, I.Y.; Grigoryeva, T.V.; Kit, O.I.; Zlatnik, E.Y.; Maksimov, A.Y.; Goncharova, A.S.; Novikova, I.A.; et al. Alkylresorcinols as New Modulators of the Metabolic Activity of the Gut Microbiota. Int. J. Mol. Sci. 2023, 24, 14206. [Google Scholar] [CrossRef]
- Kerem, G.; Yu, X.; Ismayi, A.; Teng, B.; Udduttula, A.; Liu, C.; Yu, Z.; Tohty, D.; Zhang, J.V.; Ren, P.-G. Small Intestinal Microbiota Composition Altered in Obesity-T2DM Mice with High Salt Fed. Sci. Rep. 2023, 13, 8256. [Google Scholar] [CrossRef]
- Volmer, J.G.; McRae, H.; Morrison, M. The Evolving Role of Methanogenic Archaea in Mammalian Microbiomes. Front. Microbiol. 2023, 14, 1268451. [Google Scholar] [CrossRef]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human Gut Microbiota in Obesity and after Gastric Bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef]
- Brumer, R.P.; Corrêa-Velloso, J.C.; Thomas, S.J.; Sandiford, O.A.; Thomas, A.P.; Bartlett, P.J. Short-term High-fat Diet Feeding of Mice Suppresses Catecholamine-stimulated Ca2+ Signalling in Hepatocytes and Intact Liver. J. Physiol. 2023, 601, 1383–1405. [Google Scholar] [CrossRef]
- Achterbergh, R.; Lammers, L.A.; Klümpen, H.-J.; Mathôt, R.A.A.; Romijn, J.A. Short-Term High-Fat Diet Alters Acetaminophen Metabolism in Healthy Individuals. Ther. Drug Monit. 2022, 44, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Nanizawa, E.; Otsuka, S.; Hatayama, N.; Tamaki, Y.; Hayashi, Y.; Ishikawa, T.; Hirai, S.; Naito, M. Short-Term High-Fat and High-Carbohydrate Diets Increase Susceptibility to Liver Injury by Inducing Hepatic Procoagulant and Proinflammatory Conditions with Different Balances. Nutrition 2022, 101, 111710. [Google Scholar] [CrossRef]
- Foley, K.P.; Zlitni, S.; Denou, E.; Duggan, B.M.; Chan, R.W.; Stearns, J.C.; Schertzer, J.D. Long Term but Not Short Term Exposure to Obesity Related Microbiota Promotes Host Insulin Resistance. Nat. Commun. 2018, 9, 4681. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Hao, Y.; Wang, Z.; Yang, Z.; Wang, Z.; Wang, J. 5-Heptadecylresorcinol Attenuates Oxidative Damage and Mitochondria-Mediated Apoptosis through Activation of the SIRT3/FOXO3a Signaling Pathway in Neurocytes. Food Funct. 2020, 11, 2535–2542. [Google Scholar] [CrossRef]
- Liu, J.; Hao, Y.; Wang, Z.; Ni, F.; Wang, Y.; Gong, L.; Sun, B.; Wang, J. Identification, Quantification, and Anti-Inflammatory Activity of 5-n-Alkylresorcinols from 21 Different Wheat Varieties. J. Agric. Food Chem. 2018, 66, 9241–9247. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Aboul-Enein, B.; Bernstein, J.; Marchlewicz, M. Dietary Alkylresorcinols and Cancer Prevention: A Systematic Review. Eur. Food Res. Technol. 2017, 243, 1693–1710. [Google Scholar] [CrossRef]
- Oishi, K.; Yamamoto, S.; Itoh, N.; Nakao, R.; Yasumoto, Y.; Tanaka, K.; Kikuchi, Y.; Fukudome, S.; Okita, K.; Takano-Ishikawa, Y. Wheat Alkylresorcinols Suppress High-Fat, High-Sucrose Diet-Induced Obesity and Glucose Intolerance by Increasing Insulin Sensitivity and Cholesterol Excretion in Male Mice. J. Nutr. 2015, 145, 199–206. [Google Scholar] [CrossRef]
- Basseri, R.J.; Basseri, B.; Pimentel, M.; Chong, K.; Youdim, A.; Low, K.; Hwang, L.; Soffer, E.; Chang, C.; Mathur, R. Intestinal Methane Production in Obese Individuals Is Associated with a Higher Body Mass Index. Gastroenterol. Hepatol. 2012, 8, 22–28. [Google Scholar]
- Bin Waqar, S.H.; Rehan, A. Methane and Constipation-Predominant Irritable Bowel Syndrome: Entwining Pillars of Emerging Neurogastroenterology. Cureus 2019, 11, e4764. [Google Scholar] [CrossRef] [PubMed]
- Zabolotneva, A.A.; Itov, A.B.; Grigorieva, T.V.; Vasiliev, I.Y.; Roumiantsev, S.A.; Gaponov, A.M.; Shestopalov, A.V. Reconstruction of Metabolic Activity of the Gut Microbiota in Children and Adults with Obesity and Its Relationship with the Representation of Alkylresorcinols in Stool Samples. J. Evol. Biochem. Physiol. 2023, 59, 1037–1056. [Google Scholar] [CrossRef]
- Panattoni, A.; Calvigioni, M.; Benvenuti, L.; D’Antongiovanni, V.; Pellegrini, C.; Di Salvo, C.; Mazzantini, D.; Celandroni, F.; Fornai, M.; Antonioli, L.; et al. The Administration of Enterococcus Faecium SF68 Counteracts Compositional Shifts in the Gut Microbiota of Diet-Induced Obese Mice. Front. Microbiol. 2022, 13, 1054097. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Kaicen, W.; Bian, X.; Yang, L.; Ding, S.; Li, Y.; Li, S.; Zhuge, A.; Li, L. Akkermansia Muciniphila Alleviates high-fat-diet-related Metabolic-associated Fatty Liver Disease by Modulating Gut Microbiota and Bile Acids. Microb. Biotechnol. 2023, 16, 1924–1939. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia Muciniphila: Paradigm for next-Generation Beneficial Microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Martinez-Guryn, K.; Hubert, N.; Frazier, K.; Urlass, S.; Musch, M.W.; Ojeda, P.; Pierre, J.F.; Miyoshi, J.; Sontag, T.J.; Cham, C.M.; et al. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe 2018, 23, 458–469.e5. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SD | HFD | SDet | HFDet | SDar | HFDar | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | StDev | Mean | StDev | Mean | StDev | Mean | StDev | Mean | StDev | Mean | StDev | |
| Jejunum | ||||||||||||
| Distinct OTUs | 846.00 | 219.18 | 1004.58 | 154.91 | 968.58 | 202.39 | 954.08 | 262.23 | 1076.09 | 237.01 | 1248.00 | 404.42 |
| Shannon Entropy | 6.16 | 1.63 | 6.67 | 0.49 | 6.44 | 0.99 | 6.11 | 1.24 | 6.36 | 1.19 | 6.65 | 0.96 |
| Berger–Parker Dominance | 0.17 | 0.20 | 0.09 | 0.04 | 0.14 | 0.09 | 0.16 | 0.13 | 0.15 | 0.13 | 0.13 | 0.06 |
| Chao1 Richness | 1209.52 | 287.42 | 1407.98 | 248.51 | 1342.13 | 227.88 | 1353.98 | 296.85 | 1526.96 | 382.74 | 1856.23 | 632.63 |
| Simpson Index | 0.92 | 0.14 | 0.97 | 0.01 | 0.95 | 0.04 | 0.94 | 0.08 | 0.94 | 0.07 | 0.96 | 0.03 |
| Inverse Simpson Index | 1.13 | 0.28 | 1.03 | 0.01 | 1.05 | 0.05 | 1.08 | 0.11 | 1.06 | 0.09 | 1.05 | 0.04 |
| Gini–Simpson Index | 0.08 | 0.14 | 0.03 | 0.01 | 0.05 | 0.04 | 0.06 | 0.08 | 0.06 | 0.07 | 0.04 | 0.03 |
| Colon | ||||||||||||
| Distinct OTUs * | 1109.33 | 318.61 | 1123.83 | 174.48 | 1099.58 | 77.38 | 1136.08 | 114.13 | 1248.50 | 145.17 | 1365.00 * | 262.00 |
| Shannon Entropy | 7.25 | 0.50 | 7.07 | 0.24 | 7.12 | 0.32 | 7.00 | 0.34 | 7.12 | 0.52 | 7.12 | 0.45 |
| Berger–Parker Dominance | 0.07 | 0.02 | 0.09 | 0.03 | 0.10 | 0.04 | 0.09 | 0.02 | 0.11 | 0.05 | 0.10 | 0.03 |
| Chao1 Richness * | 1503.54 | 473.34 | 1558.98 | 277.30 | 1476.15 | 115.74 | 1553.27 | 181.44 | 1673.63 | 253.43 | 2018.356 * | 433.85 |
| Simpson Index | 0.98 | 0.01 | 0.98 | 0.01 | 0.98 | 0.01 | 0.97 | 0.01 | 0.97 | 0.01 | 0.97 | 0.01 |
| Inverse Simpson Index | 1.02 | 0.01 | 1.02 | 0.01 | 1.02 | 0.01 | 1.03 | 0.01 | 1.03 | 0.02 | 1.03 | 0.01 |
| Gini–Simpson Index | 0.02 | 0.01 | 0.02 | 0.01 | 0.02 | 0.01 | 0.03 | 0.01 | 0.03 | 0.01 | 0.03 | 0.01 |
| SD vs. HFDar (SIM) | |||
|---|---|---|---|
| p Value | Mean Rank Diff. | q Value | |
| ubiquinol-7 biosynthesis (prokaryotic) | 0.000009 | 11.33 | 0.000523 |
| ubiquinol-9 biosynthesis (prokaryotic) | 0.000009 | 11.33 | 0.000523 |
| ubiquinol-10 biosynthesis (prokaryotic) | 0.000009 | 11.33 | 0.000523 |
| superpathway of L-phenylalanine biosynthesis | 0.000009 | −11.33 | 0.000523 |
| superpathway of L-tyrosine biosynthesis | 0.000009 | −11.33 | 0.000523 |
| ubiquinol-8 biosynthesis (prokaryotic) | 0.000009 | 11.33 | 0.000523 |
| superpathway of ubiquinol-8 biosynthesis (prokaryotic) | 0.000009 | 11.33 | 0.000523 |
| heme biosynthesis I (aerobic) | 0.000033 | 10.83 | 0.001717 |
| methanogenesis from acetate | 0.000144 | −10.17 | 0.00541 |
| heterolactic fermentation | 0.000144 | 10.17 | 0.00541 |
| superpathay of heme biosynthesis from glutamate | 0.000144 | 10.17 | 0.00541 |
| Bifidobacterium shunt | 0.000371 | 9.667 | 0.012028 |
| polyisoprenoid biosynthesis (E. coli) | 0.000496 | 9.5 | 0.012028 |
| peptidoglycan biosynthesis II (staphylococci) | 0.000496 | −9.5 | 0.012028 |
| superpathway of glucose and xylose degradation | 0.000496 | 9.5 | 0.012028 |
| pyrimidine deoxyribonucleotides de novo biosynthesis I | 0.000496 | 9.5 | 0.012028 |
| mevalonate pathway I | 0.000496 | 9.5 | 0.012028 |
| NAD salvage pathway II | 0.000656 | 9.333 | 0.012305 |
| superpathway of demethylmenaquinol-6 biosynthesis I | 0.000656 | −9.333 | 0.012305 |
| superpathway of demethylmenaquinol-9 biosynthesis | 0.000656 | −9.333 | 0.012305 |
| superpathway of geranylgeranyldiphosphate biosynthesis I (via mevalonate) | 0.000656 | 9.333 | 0.012305 |
| peptidoglycan biosynthesis IV (Enterococcus faecium) | 0.000656 | 9.333 | 0.012305 |
| superpathway of (Kdo)2-lipid A biosynthesis | 0.001115 | 9 | 0.019997 |
| L-1,2-propanediol degradation | 0.001433 | 8.833 | 0.022737 |
| pyrimidine deoxyribonucleotides de novo biosynthesis II | 0.001433 | 8.833 | 0.022737 |
| superpathway of pyrimidine deoxyribonucleotides de novo biosynthesis (E. coli) | 0.001433 | 8.833 | 0.022737 |
| superpathway of N-acetylneuraminate degradation | 0.002316 | 8.5 | 0.031863 |
| superpathway of menaquinol-9 biosynthesis | 0.002316 | −8.5 | 0.031863 |
| superpathway of menaquinol-6 biosynthesis I | 0.002316 | −8.5 | 0.031863 |
| superpathway of menaquinol-10 biosynthesis | 0.002316 | −8.5 | 0.031863 |
| L-lysine biosynthesis II | 0.002914 | 8.333 | 0.033402 |
| superpathway of heme biosynthesis from glycine | 0.002914 | 8.333 | 0.033402 |
| superpathway of pyrimidine ribonucleosides salvage | 0.002914 | 8.333 | 0.033402 |
| superpathway of guanosine nucleotides de novo biosynthesis I | 0.002914 | 8.333 | 0.033402 |
| peptidoglycan maturation (meso-diaminopimelate containing) | 0.002914 | 8.333 | 0.033402 |
| nitrate reduction VI (assimilatory) | 0.002914 | −8.333 | 0.033402 |
| reductive acetyl coenzyme A pathway | 0.003637 | −8.167 | 0.03573 |
| enterobacterial common antigen biosynthesis | 0.003637 | 8.167 | 0.03573 |
| gluconeogenesis I | 0.003637 | 8.167 | 0.03573 |
| superpathway of glycerol degradation to 1,3-propanediol | 0.003637 | 8.167 | 0.03573 |
| acetylene degradation | 0.003637 | 8.167 | 0.03573 |
| pyrimidine deoxyribonucleotide phosphorylation | 0.003637 | 8.167 | 0.03573 |
| fatty acid elongation—saturated | 0.004513 | 8 | 0.041385 |
| ppGpp biosynthesis | 0.004513 | 8 | 0.041385 |
| pyruvate fermentation to acetate and lactate II | 0.004513 | 8 | 0.041385 |
| pyruvate fermentation to propanoate I | 0.00556 | −7.833 | 0.048812 |
| superpathway of guanosine nucleotides de novo biosynthesis II | 0.00556 | 7.833 | 0.048812 |
| HFD vs. HFDar (SIM) | |||
|---|---|---|---|
| p Value | Mean Rank Diff. | q Value | |
| superpathway of ubiquinol-8 biosynthesis (prokaryotic) | 0.000033 | 10.83 | 0.004412 |
| ubiquinol-7 biosynthesis (prokaryotic) | 0.00005 | 10.67 | 0.004412 |
| ubiquinol-9 biosynthesis (prokaryotic) | 0.00005 | 10.67 | 0.004412 |
| ubiquinol-10 biosynthesis (prokaryotic) | 0.00005 | 10.67 | 0.004412 |
| ubiquinol-8 biosynthesis (prokaryotic) | 0.00005 | 10.67 | 0.004412 |
| heme biosynthesis I (aerobic) | 0.000274 | 9.833 | 0.016529 |
| hexitol fermentation to lactate, formate, ethanol and acetate | 0.000274 | 9.833 | 0.016529 |
| superpathway of menaquinol-8 biosynthesis II | 0.000371 | 9.667 | 0.016529 |
| 1,4-dihydroxy-6-naphthoate biosynthesis II | 0.000371 | 9.667 | 0.016529 |
| 1,4-dihydroxy-6-naphthoate biosynthesis I | 0.000371 | 9.667 | 0.016529 |
| superpathway of L-alanine biosynthesis | 0.000656 | −9.333 | 0.026551 |
| superpathway of sulfur oxidation (Acidianus ambivalens) | 0.000858 | 9.167 | 0.027282 |
| superpathay of heme biosynthesis from glutamate | 0.000858 | 9.167 | 0.027282 |
| L-1,2-propanediol degradation | 0.000858 | 9.167 | 0.027282 |
| HFD vs. HFDar (LIM) | |||
|---|---|---|---|
| p Value | Mean Rank Diff. | q Value | |
| glucose and glucose-1-phosphate degradation | <0.000001 | −12 | 0.000031 |
| 1,4-dihydroxy-2-naphthoate biosynthesis I | <0.000001 | −12 | 0.000031 |
| superpathway of menaquinol-8 biosynthesis I | <0.000001 | −12 | 0.000031 |
| superpathway of menaquinol-7 biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of demethylmenaquinol-8 biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of phylloquinol biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of menaquinol-11 biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of menaquinol-12 biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of menaquinol-13 biosynthesis | <0.000001 | −12 | 0.000031 |
| superpathway of L-alanine biosynthesis | <0.000001 | −12 | 0.000031 |
| sulfate reduction I (assimilatory) | 0.000001 | −11.83 | 0.000051 |
| superpathway of sulfate assimilation and cysteine biosynthesis | 0.000001 | −11.83 | 0.000051 |
| superpathway of L-methionine biosynthesis (by sulfhydrylation) | 0.000005 | −11.5 | 0.000165 |
| fucose degradation | 0.000014 | −11.17 | 0.000417 |
| superpathway of pyridoxal 5′-phosphate biosynthesis and salvage | 0.000022 | −11 | 0.000614 |
| superpathway of 2,3-butanediol biosynthesis | 0.000033 | 10.83 | 0.000813 |
| 1,4-dihydroxy-6-naphthoate biosynthesis I | 0.000033 | 10.83 | 0.000813 |
| TCA cycle VI (obligate autotrophs) | 0.00005 | −10.67 | 0.001083 |
| 1,4-dihydroxy-6-naphthoate biosynthesis II | 0.00005 | 10.67 | 0.001083 |
| preQ0 biosynthesis | 0.000072 | −10.5 | 0.001489 |
| adenosylcobalamin salvage from cobinamide I | 0.000144 | −10.17 | 0.002603 |
| L-histidine degradation I | 0.000144 | 10.17 | 0.002603 |
| pyridoxal 5′-phosphate biosynthesis I | 0.000144 | −10.17 | 0.002603 |
| superpathway of (Kdo)2-lipid A biosynthesis | 0.000273 | 9.833 | 0.004556 |
| superpathway of menaquinol-8 biosynthesis II | 0.000274 | 9.833 | 0.004556 |
| glucose degradation (oxidative) | 0.000337 | −9 | 0.005373 |
| superpathway of UDP-N-acetylglucosamine-derived O-antigen building blocks biosynthesis | 0.000496 | 9.5 | 0.007619 |
| L-lysine biosynthesis II | 0.000656 | 9.333 | 0.009726 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zabolotneva, A.A.; Vasiliev, I.Y.; Grigoryeva, T.; Gaponov, A.M.; Chekhonin, V.P.; Roumiantsev, S.A.; Shestopalov, A.V. Supplementation of a High-Fat Diet with Pentadecylresorcinol Increases the Representation of Akkermansia muciniphila in the Mouse Small and Large Intestines and May Protect against Complications Caused by Imbalanced Nutrition. Int. J. Mol. Sci. 2024, 25, 6611. https://doi.org/10.3390/ijms25126611
Zabolotneva AA, Vasiliev IY, Grigoryeva T, Gaponov AM, Chekhonin VP, Roumiantsev SA, Shestopalov AV. Supplementation of a High-Fat Diet with Pentadecylresorcinol Increases the Representation of Akkermansia muciniphila in the Mouse Small and Large Intestines and May Protect against Complications Caused by Imbalanced Nutrition. International Journal of Molecular Sciences. 2024; 25(12):6611. https://doi.org/10.3390/ijms25126611
Chicago/Turabian StyleZabolotneva, Anastasia A., Ilya Yu. Vasiliev, Tatiana Grigoryeva, Andrei M. Gaponov, Vladimir P. Chekhonin, Sergei A. Roumiantsev, and Aleksandr V. Shestopalov. 2024. "Supplementation of a High-Fat Diet with Pentadecylresorcinol Increases the Representation of Akkermansia muciniphila in the Mouse Small and Large Intestines and May Protect against Complications Caused by Imbalanced Nutrition" International Journal of Molecular Sciences 25, no. 12: 6611. https://doi.org/10.3390/ijms25126611