
The Purinergic P2X7 Receptor as a Target for Adjunctive Treatment for Drug-Refractory Epilepsy

 , ,
, , 
{kind=link}
Abstract
1. Introduction
2. The P2X7 Receptor
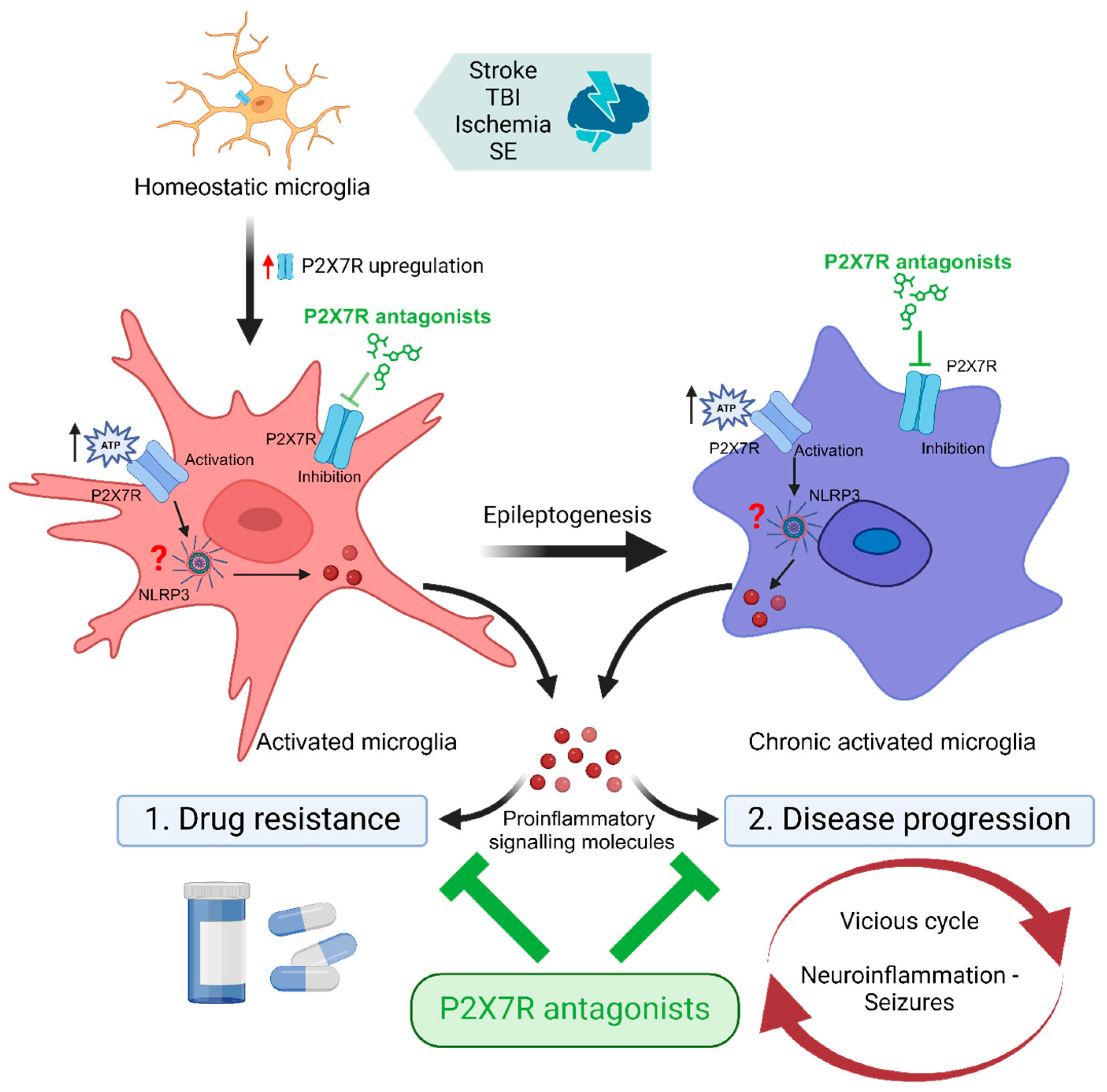
3. The P2X7R as a Treatment Target for Drug-Refractory Epilepsy
3.1. ATP Release during Seizures and Epilepsy
3.2. P2X7R Expression and Cell-Specific Localisation following Seizures and in Epilepsy
3.3. P2X7R Function during Seizures and Epilepsy
3.4. P2X7R as an Adjunctive Treatment Target for Drug-Refractory Epilepsy
4. Challenges, Considerations, and Future Perspectives
- (i)
- Foremost, we need a clearer picture of how P2X7R down-stream signalling affects seizures. Even though the P2X7R-dependent activation of inflammation seems to be the most plausible mechanism of how P2X7Rs contribute to seizures and drug-refractoriness [62,131], the P2X7R is involved in numerous other (patho)physiological processes. For instance, the large C-terminus of P2X7Rs can interact with more than 50 cytosolic proteins involved in different physiological and pathophysiological mechanisms. Moreover, the recent discovery of opposing functions of the P2X7R during seizures with microglia P2X7Rs contributing to seizures and neuronal P2X7Rs reducing seizures [62] further demonstrates the complex P2X7R signalling cascades during epilepsy. Therefore, the identification of the cell type-specific P2X7R functions and mechanism during seizures and epilepsy will not only allow us to design the best treatment strategy, but also help to predict potential side-effects.
- (ii)
- While the data have shown increased eATP concentrations during seizures/epilepsy, we still do not know how, when, and where ATP is released during seizures, which is critical to predict P2X7R activation. Interestingly, as mentioned before, P2X7Rs themselves can contribute to ATP release. In addition, several endogenous-positive allosteric modulators such as phosphoinositide, lysophosphatidylcholine, nicotinamide, and adenine dinucleotide may sensitise the P2X7R to lower eATP concentrations [36]. While ATP availability is an important factor to take into account when considering P2X7R activation during seizures, P2X7R activity may also be regulated via the expression of different splice variants [157], which exhibit modified functions, or its subcellular redistribution (e.g., trafficking, cell surface expression changes, as reported following i.p. pilocarpine-induced SE in rats [158]).
- (iii)
- It is increasingly recognised that effective treatments require the identification of companion biomarkers. In this line, recent research has shown that P2X7R radioli-gand uptake measured via positron emission tomography (PET) imaging increased in the brain and correlated with seizure severity during IAKA-induced SE and the SE-induced underlying pathology [159,160], representing a possible readout of seizure-induced neuropathology. In addition, the P2X7R protein levels have been reported to be increased in patients with TLE and several inflammatory markers have been shown to be altered in the blood of mice post-SE in a P2X7R-dependent manner [161]. Future studies should identify biomarkers not only to predict P2X7R activation, but also to identify patients most likely benefiting from P2X7R-based treatments (e.g., P2X7R activation in the microglia).
- (iv)
- While combination treatments have been shown to be effective in SE, future research should determine their potential in reducing seizures during drug-refractory epilepsy. Future studies should also determine whether P2X7R antagonists potentiate the effects of only a certain subclass of ASMs and whether there are undesired drug interactions between P2X7R antagonists and ASMs. P2X7R function/cell-specific localisation (e.g., microglia vs. neurons) may change during disease progression, which, in turn, may impact the treatment effectiveness.
- (v)
- Inflammation is a well-accepted pathomechanism in age-associated disorders including epilepsy [162]. Notably, P2X7Rs are increasingly recognised as a treatment target for an array of age-related diseases of the CNS (e.g., neurodegenerative diseases Alzheimer’s and Parkinson’s, cancers, arthritis), possibly contributing to the chronic low-level systemic inflammation observed in these diseases [64,163]. Future studies should, therefore, be designed to establish whether chronic P2X7R activation contributes to sustained brain inflammation during ageing and whether this contributes to the observed increase in epilepsy prevalence among the aged population [164].
- (vi)
- The human P2RX7 gene is highly polymorphic with more than 150 non-synonymous single-nucleotide polymorphisms (SNPs) reported either as loss- or gain-of-function variants (e.g., ion channel activity, pore function) and agonist-binding affinities [165], which should be taken into account when designing P2X7R-targeting drugs. Interestingly, the P2X7R rs208294 His155Tyr polymorphism has been associated with childhood febrile seizure susceptibility [166], suggesting that P2X7R polymorphisms have an impact on brain hyperexcitability. Interestingly, several P2X7R polymorphisms have been associated with common comorbidities associated with epilepsy (e.g., depression, anxiety) [25]. Future studies, should be designed to establish the impact of certain P2X7R SNPs on one’s predisposition to develop epilepsy, responsiveness to ASMs, and the presence of epilepsy-associated comorbidities.
- (vii)
- While P2X7Rs are believed to be mainly activated following tissue injury, acting as damage-sensing receptors, P2X7Rs have been ascribed numerous functions during normal physiology, e.g., the stimulation of glutamate and GABA release from astrocytes and neurons as well as the microglial release of neurotropic factors that may be inhibited by P2X7R blockade. P2X7Rs have a well-established role in the immune-inflammatory system. Would P2X7R antagonism increase the risk of infectious diseases? Importantly, P2X7Rs are expressed throughout the body (e.g., heart, gastrointestinal system [167]), thus effects of P2X7R antagonism may not be restricted to the CNS, but involve other organs. While the risk of peripheral side-effects should be minimised by using CNS-permeable P2X7R antagonists, future studies should carefully determine P2X7R safety profiles during treatments.
- (viii)
- Finally, the possible systemic side-effects caused by P2X7R antagonists and the opposing effects of the P2X7R in different cell types [62] suggest that a focal/cell type-specific delivery may provide a safer and more efficient treatment strategy. In this regard, CNS-directed gene therapy targeting the appropriate cell type(s) (e.g., overexpression of P2X7R siRNA in microglia) in target tissue(s) represents a powerful tool to achieve the long-term corrections of disorders following a single treatment [168]. Nanoparticles delivered to specific cell types or brain regions represent another potential approach. Nanostructures can be utilised as delivery agents by encapsulating drugs or attaching therapeutic drugs (e.g., siRNA) and delivering them to target tissues more precisely with a controlled release, including microglia [169,170]. Critically, epilepsy treatment offers the unique opportunity for local drug delivery directed into the seizure focus, thereby avoiding systemic drug delivery and maximising drug effects without the need for additional invasive surgery.
Funding
Acknowledgments
Conflicts of Interest
References
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshe, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Sarmast, S.T.; Abdullahi, A.M.; Jahan, N. Current Classification of Seizures and Epilepsies: Scope, Limitations and Recommendations for Future Action. Cureus 2020, 12, e10549. [Google Scholar] [CrossRef] [PubMed]
- Thijs, R.D.; Surges, R.; O‘Brien, T.J.; Sander, J.W. Epilepsy in adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Moshe, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef] [PubMed]
- Löscher, W.; Potschka, H.; Sisodiya, S.M.; Vezzani, A. Drug Resistance in Epilepsy: Clinical Impact, Potential Mechanisms, and New Innovative Treatment Options. Pharmacol. Rev. 2020, 72, 606–638. [Google Scholar] [CrossRef] [PubMed]
- Shlobin, N.A.; Sander, J.W. Learning from the comorbidities of epilepsy. Curr. Opin. Neurol. 2022, 35, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.S.; Lowenstein, D.H. Epilepsy. N. Engl. J. Med. 2003, 349, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Beamer, E.; Kuchukulla, M.; Boison, D.; Engel, T. ATP and adenosine-Two players in the control of seizures and epilepsy development. Prog. Neurobiol. 2021, 204, 102105. [Google Scholar] [CrossRef]
- Klein, P.; Dingledine, R.; Aronica, E.; Bernard, C.; Blumcke, I.; Boison, D.; Brodie, M.J.; Brooks-Kayal, A.R.; Engel, J., Jr.; Forcelli, P.A.; et al. Commonalities in epileptogenic processes from different acute brain insults: Do they translate? Epilepsia 2018, 59, 37–66. [Google Scholar] [CrossRef]
- Pitkanen, A.; Engel, J., Jr. Past and present definitions of epileptogenesis and its biomarkers. Neurotherapeutics 2014, 11, 231–241. [Google Scholar] [CrossRef]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef]
- Consales, A.; Casciato, S.; Asioli, S.; Barba, C.; Caulo, M.; Colicchio, G.; Cossu, M.; de Palma, L.; Morano, A.; Vatti, G.; et al. The surgical treatment of epilepsy. Neurol. Sci. 2021, 42, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Foutz, T.J.; Wong, M. Brain stimulation treatments in epilepsy: Basic mechanisms and clinical advances. Biomed. J. 2022, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.M.; Boison, D. The metabolic basis of epilepsy. Nat. Rev. Neurol. 2022, 18, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, S.; Trinka, E.; Zaccara, G.; Striano, P.; Russo, E.; Del Giovane, C.; Silvestrini, M.; Brigo, F. Third-Generation Antiseizure Medications for Adjunctive Treatment of Focal-Onset Seizures in Adults: A Systematic Review and Network Meta-analysis. Drugs 2022, 82, 199–218. [Google Scholar] [CrossRef]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Allen Hauser, W.; Mathern, G.; Moshe, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Hartz, A.M.S.; Bauer, B. Drug-Resistant Epilepsy: Multiple Hypotheses, Few Answers. Front. Neurol. 2017, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Bazhanova, E.D.; Kozlov, A.A.; Litovchenko, A.V. Mechanisms of Drug Resistance in the Pathogenesis of Epilepsy: Role of Neuroinflammation. A Literature Review. Brain Sci. 2021, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, G.; Depietri, G.; Michev, A.; Riva, A.; Foiadelli, T.; Savasta, S.; Bonuccelli, A.; Peroni, D.; Consolini, R.; Marseglia, G.L.; et al. Targeting Inflammatory Mediators in Epilepsy: A Systematic Review of Its Molecular Basis and Clinical Applications. Front. Neurol. 2022, 13, 741244. [Google Scholar] [CrossRef]
- Perucca, E.; Perucca, P.; White, H.S.; Wirrell, E.C. Drug resistance in epilepsy. Lancet Neurol. 2023, 22, 723–734. [Google Scholar] [CrossRef]
- Kanner, A.M.; Bicchi, M.M. Antiseizure Medications for Adults with Epilepsy: A Review. JAMA 2022, 327, 1269–1281. [Google Scholar] [CrossRef]
- Engel, T.; Smith, J.; Alves, M. Targeting Neuroinflammation via Purinergic P2 Receptors for Disease Modification in Drug-Refractory Epilepsy. J. Inflamm. Res. 2021, 14, 3367–3392. [Google Scholar] [CrossRef]
- Henshall, D.C.; Engel, T. P2X purinoceptors as a link between hyperexcitability and neuroinflammation in status epilepticus. Epilepsy Behav. 2015, 49, 8–12. [Google Scholar] [CrossRef]
- Beamer, E.; Fischer, W.; Engel, T. The ATP-Gated P2X7 Receptor As a Target for the Treatment of Drug-Resistant Epilepsy. Front. Neurosci. 2017, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Gil, B.; Smith, J.; Tang, Y.; Illes, P.; Engel, T. Beyond Seizure Control: Treating Comorbidities in Epilepsy via Targeting of the P2X7 Receptor. Int. J. Mol. Sci. 2022, 23, 2380. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, C.; Bravo-Tobar, I.D.; Duarte, Y.; Barrio, L.C.; Saez, J.C. Contribution of non-selective membrane channels and receptors in epilepsy. Pharmacol. Ther. 2022, 231, 107980. [Google Scholar] [CrossRef] [PubMed]
- Engel, T. The P2X7 Receptor as a Mechanistic Biomarker for Epilepsy. Int. J. Mol. Sci. 2023, 24, 5410. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.H.; Tang, Y.; Illes, P. The neuroinflammatory astrocytic P2X7 receptor: Alzheimer’s disease, ischemic brain injury, and epileptic state. Expert. Opin. Ther. Targets 2023, 27, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Burnstock, G. Biology of purinergic signalling: Its ancient evolutionary roots, its omnipresence and its multiple functional significance. Bioessays 2014, 36, 697–705. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic nerves. Pharmacol. Rev. 1972, 24, 509–581. [Google Scholar] [PubMed]
- Burnstock, G. Introduction to Purinergic Signalling in the Brain. Adv. Exp. Med. Biol. 2020, 1202, 1–12. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Sarti, A.C.; Coutinho-Silva, R. Purinergic signaling, DAMPs, and inflammation. Am. J. Physiol. Cell Physiol. 2020, 318, C832–C835. [Google Scholar] [CrossRef] [PubMed]
- Illes, P.; Xu, G.Y.; Tang, Y. Purinergic Signaling in the Central Nervous System in Health and Disease. Neurosci. Bull. 2020, 36, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef] [PubMed]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Cuesta, M.A.; Blanch-Ruiz, M.A.; Ortega-Luna, R.; Sanchez-Lopez, A.; Alvarez, A. Structural and Functional Basis for Understanding the Biological Significance of P2X7 Receptor. Int. J. Mol. Sci. 2020, 21, 8454. [Google Scholar] [CrossRef] [PubMed]
- De Salis, S.K.F.; Li, L.; Chen, Z.; Lam, K.W.; Skarratt, K.K.; Balle, T.; Fuller, S.J. Alternatively Spliced Isoforms of the P2X7 Receptor: Structure, Function and Disease Associations. Int. J. Mol. Sci. 2022, 23, 8174. [Google Scholar] [CrossRef]
- Nicke, A. Homotrimeric complexes are the dominant assembly state of native P2X7 subunits. Biochem. Biophys. Res. Commun. 2008, 377, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Dubyak, G.R. Go it alone no more--P2X7 joins the society of heteromeric ATP-gated receptor channels. Mol. Pharmacol. 2007, 72, 1402–1405. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Kennedy, C. P2X receptors in health and disease. Adv. Pharmacol. 2011, 61, 333–372. [Google Scholar] [CrossRef]
- Skaper, S.D.; Debetto, P.; Giusti, P. The P2X7 purinergic receptor: From physiology to neurological disorders. FASEB J. 2010, 24, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Na, H.Y.; Chong, K.H.; Kim, T.J. P2X7 receptor-dependent ATP-induced shedding of CD27 in mouse lymphocytes. Immunol. Lett. 2006, 102, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 2011, 63, 641–683. [Google Scholar] [CrossRef] [PubMed]
- Costa-Junior, H.M.; Sarmento Vieira, F.; Coutinho-Silva, R. C terminus of the P2X7 receptor: Treasure hunting. Purinergic Signal. 2011, 7, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, A.; Michalski, K.; Mikhelzon, P.; Kawate, T. The P2X7 receptor forms a dye-permeable pore independent of its intracellular domain but dependent on membrane lipid composition. Elife 2017, 6, 31186. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.E.; Yoshioka, C.; Mansoor, S.E. Full-Length P2X(7) Structures Reveal How Palmitoylation Prevents Channel Desensitization. Cell 2019, 179, 659–670.e613. [Google Scholar] [CrossRef] [PubMed]
- Kopp, R.; Krautloher, A.; Ramirez-Fernandez, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Cevoli, F.; Arnould, B.; Peralta, F.A.; Grutter, T. Untangling Macropore Formation and Current Facilitation in P2X7. Int. J. Mol. Sci. 2023, 24, 10896. [Google Scholar] [CrossRef]
- Allsopp, R.C.; Evans, R.J. Contribution of the Juxtatransmembrane Intracellular Regions to the Time Course and Permeation of ATP-gated P2X7 Receptor Ion Channels. J. Biol. Chem. 2015, 290, 14556–14566. [Google Scholar] [CrossRef]
- Falzoni, S.; Donvito, G.; Di Virgilio, F. Detecting adenosine triphosphate in the pericellular space. Interface Focus 2013, 3, 20120101. [Google Scholar] [CrossRef]
- Dona, F.; Conceicao, I.M.; Ulrich, H.; Ribeiro, E.B.; Freitas, T.A.; Nencioni, A.L.; da Silva Fernandes, M.J. Variations of ATP and its metabolites in the hippocampus of rats subjected to pilocarpine-induced temporal lobe epilepsy. Purinergic Signal. 2016, 12, 295–302. [Google Scholar] [CrossRef]
- Beamer, E.; Conte, G.; Engel, T. ATP release during seizures—A critical evaluation of the evidence. Brain Res. Bull. 2019, 151, 65–73. [Google Scholar] [CrossRef]
- Moro, N.; Ghavim, S.S.; Sutton, R.L. Massive efflux of adenosine triphosphate into the extracellular space immediately after experimental traumatic brain injury. Exp. Ther. Med. 2021, 21, 575. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.J.; Tome, A.R.; Cunha, R.A. ATP as a multi-target danger signal in the brain. Front. Neurosci. 2015, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Solle, M.; Labasi, J.; Perregaux, D.G.; Stam, E.; Petrushova, N.; Koller, B.H.; Griffiths, R.J.; Gabel, C.A. Altered cytokine production in mice lacking P2X(7) receptors. J. Biol. Chem. 2001, 276, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Sperlagh, B.; Kofalvi, A.; Deuchars, J.; Atkinson, L.; Milligan, C.J.; Buckley, N.J.; Vizi, E.S. Involvement of P2X7 receptors in the regulation of neurotransmitter release in the rat hippocampus. J. Neurochem. 2002, 81, 1196–1211. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.C.; Parfitt, G.M.; Polese, C.E.; Coutinho-Silva, R.; Morrone, F.B.; Barros, D.M. Pharmacological blockage and P2X7 deletion hinder aversive memories: Reversion in an enriched environment. Neuroscience 2014, 280, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Leeson, H.C.; Chan-Ling, T.; Lovelace, M.D.; Brownlie, J.C.; Gu, B.J.; Weible, M.W. P2X7 receptor signaling during adult hippocampal neurogenesis. Neural Regen. Res. 2019, 14, 1684–1694. [Google Scholar] [CrossRef] [PubMed]
- Mut-Arbona, P.; Huang, L.; Baranyi, M.; Tod, P.; Iring, A.; Calzaferri, F.; de Los Rios, C.; Sperlagh, B. Dual Role of the P2X7 Receptor in Dendritic Outgrowth during Physiological and Pathological Brain Development. J. Neurosci. 2023, 43, 1125–1142. [Google Scholar] [CrossRef]
- Conte, G.; Nguyen, N.T.; Alves, M.; de Diego-Garcia, L.; Kenny, A.; Nicke, A.; Henshall, D.C.; Jimenez-Mateos, E.M.; Engel, T. P2X7 Receptor-Dependent microRNA Expression Profile in the Brain Following Status Epilepticus in Mice. Front. Mol. Neurosci. 2020, 13, 127. [Google Scholar] [CrossRef]
- Smith, J.; Menendez Mendez, A.; Alves, M.; Parras, A.; Conte, G.; Bhattacharya, A.; Ceusters, M.; Nicke, A.; Henshall, D.C.; Jimenez-Mateos, E.M.; et al. The P2X7 receptor contributes to seizures and inflammation-driven long-lasting brain hyperexcitability following hypoxia in neonatal mice. Br. J. Pharmacol. 2023, 180, 1710–1729. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Gil, B.; Villegas-Salmeron, J.; Salari, V.; Martins-Ferreira, R.; Arribas Blazquez, M.; Menendez Mendez, A.; Da Rosa Gerbatin, R.; Smith, J.; de Diego-Garcia, L.; et al. Opposing effects of the purinergic P2X7 receptor on seizures in neurons and microglia in male mice. Brain Behav. Immun. 2024, 120, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Sperlagh, B.; Illes, P. P2X7 receptor: An emerging target in central nervous system diseases. Trends Pharmacol. Sci. 2014, 35, 537–547. [Google Scholar] [CrossRef]
- Andrejew, R.; Oliveira-Giacomelli, A.; Ribeiro, D.E.; Glaser, T.; Arnaud-Sampaio, V.F.; Lameu, C.; Ulrich, H. The P2X7 Receptor: Central Hub of Brain Diseases. Front. Mol. Neurosci. 2020, 13, 124. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Pacheco, A.; Diaz-Hernandez, M.; Arribas-Blazquez, M.; Sanz-Rodriguez, A.; Olivos-Ore, L.A.; Artalejo, A.R.; Alves, M.; Letavic, M.; Miras-Portugal, M.T.; Conroy, R.M.; et al. Transient P2X7 Receptor Antagonism Produces Lasting Reductions in Spontaneous Seizures and Gliosis in Experimental Temporal Lobe Epilepsy. J. Neurosci. 2016, 36, 5920–5932. [Google Scholar] [CrossRef] [PubMed]
- Mamad, O.; Heiland, M.; Lindner, A.U.; Hill, T.D.M.; Ronroy, R.M.; Rentrup, K.; Sanz-Rodriguez, A.; Langa, E.; Heller, J.P.; Moreno, O.; et al. Anti-seizure effects of JNJ-54175446 in the intra-amygdala kainic acid model of drug-resistant temporal lobe epilepsy in mice. Front. Pharmacol. 2023, 14, 1308478. [Google Scholar] [CrossRef]
- Illes, P.; Khan, T.M.; Rubini, P. Neuronal P2X7 Receptors Revisited: Do They Really Exist? J. Neurosci. 2017, 37, 7049–7062. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek-Hajek, K.; Zhang, J.; Kopp, R.; Grosche, A.; Rissiek, B.; Saul, A.; Bruzzone, S.; Engel, T.; Jooss, T.; Krautloher, A.; et al. Re-evaluation of neuronal P2X7 expression using novel mouse models and a P2X7-specific nanobody. Elife 2018, 7, 36217. [Google Scholar] [CrossRef]
- Miras-Portugal, M.T.; Ortega, F.; Gomez-Villafuertes, R.; Gualix, J.; Perez-Sen, R.; Delicado, E.G. P2X7 receptors in the central nervous system. Biochem. Pharmacol. 2021, 187, 114472. [Google Scholar] [CrossRef]
- Danquah, W.; Meyer-Schwesinger, C.; Rissiek, B.; Pinto, C.; Serracant-Prat, A.; Amadi, M.; Iacenda, D.; Knop, J.H.; Hammel, A.; Bergmann, P.; et al. Nanobodies that block gating of the P2X7 ion channel ameliorate inflammation. Sci. Transl. Med. 2016, 8, 366ra162. [Google Scholar] [CrossRef]
- Metzger, M.W.; Walser, S.M.; Dedic, N.; Aprile-Garcia, F.; Jakubcakova, V.; Adamczyk, M.; Webb, K.J.; Uhr, M.; Refojo, D.; Schmidt, M.V.; et al. Heterozygosity for the Mood Disorder-Associated Variant Gln460Arg Alters P2X7 Receptor Function and Sleep Quality. J. Neurosci. 2017, 37, 11688–11700. [Google Scholar] [CrossRef] [PubMed]
- Pelegrin, P. P2X7 receptor and the NLRP3 inflammasome: Partners in crime. Biochem. Pharmacol. 2021, 187, 114385. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.; Sills, G.J. Inflammation and epilepsy: The foundations for a new therapeutic approach in epilepsy? Epilepsy Curr. 2012, 12, 8–12. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, E.A.; Aronica, E.; Vezzani, A.; Ravizza, T. Review: Neuroinflammatory pathways as treatment targets and biomarker candidates in epilepsy: Emerging evidence from preclinical and clinical studies. Neuropathol. Appl. Neurobiol. 2018, 44, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Hanin, A.; Muscal, E.; Hirsch, L.J. Second-line immunotherapy in new onset refractory status epilepticus. Epilepsia 2024, 65, 1203–1223. [Google Scholar] [CrossRef] [PubMed]
- Monif, M.; Reid, C.A.; Powell, K.L.; Smart, M.L.; Williams, D.A. The P2X7 receptor drives microglial activation and proliferation: A trophic role for P2X7R pore. J. Neurosci. 2009, 29, 3781–3791. [Google Scholar] [CrossRef] [PubMed]
- Campagno, K.E.; Mitchell, C.H. The P2X(7) Receptor in Microglial Cells Modulates the Endolysosomal Axis, Autophagy, and Phagocytosis. Front. Cell Neurosci. 2021, 15, 645244. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Franke, H.; Krugel, U.; Muller, H.; Dinkel, K.; Lord, B.; Letavic, M.A.; Henshall, D.C.; Engel, T. Critical Evaluation of P2X7 Receptor Antagonists in Selected Seizure Models. PLoS ONE 2016, 11, e0156468. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Deng, X.J.; Xu, D. Microglia in epilepsy. Neurobiol. Dis. 2023, 185, 106249. [Google Scholar] [CrossRef]
- Pitkanen, A.; Lukasiuk, K. Mechanisms of epileptogenesis and potential treatment targets. Lancet Neurol. 2011, 10, 173–186. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Giacomelli, A.; Petiz, L.L.; Andrejew, R.; Turrini, N.; Silva, J.B.; Sack, U.; Ulrich, H. Role of P2X7 Receptors in Immune Responses During Neurodegeneration. Front. Cell Neurosci. 2021, 15, 662935. [Google Scholar] [CrossRef] [PubMed]
- Territo, P.R.; Zarrinmayeh, H. P2X(7) Receptors in Neurodegeneration: Potential Therapeutic Applications From Basic to Clinical Approaches. Front. Cell Neurosci. 2021, 15, 617036. [Google Scholar] [CrossRef]
- Illes, P. P2X7 Receptors Amplify CNS Damage in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 5996. [Google Scholar] [CrossRef]
- Cicko, S.; Kohler, T.C.; Ayata, C.K.; Muller, T.; Ehrat, N.; Meyer, A.; Hossfeld, M.; Zech, A.; Di Virgilio, F.; Idzko, M. Extracellular ATP is a danger signal activating P2X7 receptor in a LPS mediated inflammation (ARDS/ALI). Oncotarget 2018, 9, 30635–30648. [Google Scholar] [CrossRef]
- Dale, N.; Frenguelli, B.G. Release of adenosine and ATP during ischemia and epilepsy. Curr. Neuropharmacol. 2009, 7, 160–179. [Google Scholar] [CrossRef]
- Zimmermann, H. History of ectonucleotidases and their role in purinergic signaling. Biochem. Pharmacol. 2021, 187, 114322. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, Y.; Lalo, U.; Verkhratsky, A.; North, R.A. Vesicular release of ATP at central synapses. Pflug. Arch. 2006, 452, 589–597. [Google Scholar] [CrossRef]
- Pankratov, Y.; Lalo, U.; Verkhratsky, A.; North, R.A. Quantal release of ATP in mouse cortex. J. Gen. Physiol. 2007, 129, 257–265. [Google Scholar] [CrossRef]
- Jo, Y.H.; Schlichter, R. Synaptic corelease of ATP and GABA in cultured spinal neurons. Nat. Neurosci. 1999, 2, 241–245. [Google Scholar] [CrossRef]
- Hugel, S.; Schlichter, R. Presynaptic P2X receptors facilitate inhibitory GABAergic transmission between cultured rat spinal cord dorsal horn neurons. J. Neurosci. 2000, 20, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Haydon, P.G.; Yeung, E.S. Direct observation of calcium-independent intercellular ATP signaling in astrocytes. Anal. Chem. 2000, 72, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Echigo, N.; Juge, N.; Miyaji, T.; Otsuka, M.; Omote, H.; Yamamoto, A.; Moriyama, Y. Identification of a vesicular nucleotide transporter. Proc. Natl. Acad. Sci. USA 2008, 105, 5683–5686. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.; Cherny, V.V.; Morgan, D.; DeCoursey, T.E. Voltage-gated proton channels help regulate pHi in rat alveolar epithelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L398–L408. [Google Scholar] [CrossRef]
- Johnson, M.D.; Bao, H.F.; Helms, M.N.; Chen, X.J.; Tigue, Z.; Jain, L.; Dobbs, L.G.; Eaton, D.C. Functional ion channels in pulmonary alveolar type I cells support a role for type I cells in lung ion transport. Proc. Natl. Acad. Sci. USA 2006, 103, 4964–4969. [Google Scholar] [CrossRef]
- Bao, L.; Locovei, S.; Dahl, G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Pellegatti, P.; Falzoni, S.; Pinton, P.; Rizzuto, R.; Di Virgilio, F. A novel recombinant plasma membrane-targeted luciferase reveals a new pathway for ATP secretion. Mol. Biol. Cell 2005, 16, 3659–3665. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Kang, N.; Lovatt, D.; Torres, A.; Zhao, Z.; Lin, J.; Nedergaard, M. Connexin 43 hemichannels are permeable to ATP. J. Neurosci. 2008, 28, 4702–4711. [Google Scholar] [CrossRef]
- Wieraszko, A.; Seyfried, T.N. Increased amount of extracellular ATP in stimulated hippocampal slices of seizure prone mice. Neurosci. Lett. 1989, 106, 287–293. [Google Scholar] [CrossRef]
- Heinrich, A.; Ando, R.D.; Turi, G.; Rozsa, B.; Sperlagh, B. K+ depolarization evokes ATP, adenosine and glutamate release from glia in rat hippocampus: A microelectrode biosensor study. Br. J. Pharmacol. 2012, 167, 1003–1020. [Google Scholar] [CrossRef]
- Lopatar, J.; Dale, N.; Frenguelli, B.G. Pannexin-1-mediated ATP release from area CA3 drives mGlu5-dependent neuronal oscillations. Neuropharmacology 2015, 93, 219–228. [Google Scholar] [CrossRef]
- Dossi, E.; Blauwblomme, T.; Moulard, J.; Chever, O.; Vasile, F.; Guinard, E.; Le Bert, M.; Couillin, I.; Pallud, J.; Capelle, L.; et al. Pannexin-1 channels contribute to seizure generation in human epileptic brain tissue and in a mouse model of epilepsy. Sci. Transl. Med. 2018, 10, eaar3796. [Google Scholar] [CrossRef]
- Boison, D. Adenosinergic signaling in epilepsy. Neuropharmacology 2016, 104, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Dunwiddie, T.V. Adenosine and suppression of seizures. Adv. Neurol. 1999, 79, 1001–1010. [Google Scholar]
- Pagonopoulou, O.; Efthimiadou, A.; Asimakopoulos, B.; Nikolettos, N.K. Modulatory role of adenosine and its receptors in epilepsy: Possible therapeutic approaches. Neurosci. Res. 2006, 56, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Hasko, G.; Cronstein, B. Regulation of inflammation by adenosine. Front. Immunol. 2013, 4, 85. [Google Scholar] [CrossRef]
- Baltos, J.A.; Casillas-Espinosa, P.M.; Rollo, B.; Gregory, K.J.; White, P.J.; Christopoulos, A.; Kwan, P.; O’Brien, T.J.; May, L.T. The role of the adenosine system in epilepsy and its comorbidities. Br. J. Pharmacol. 2023, 1–5. [Google Scholar] [CrossRef]
- Guieu, R.; Deharo, J.C.; Maille, B.; Crotti, L.; Torresani, E.; Brignole, M.; Parati, G. Adenosine and the Cardiovascular System: The Good and the Bad. J. Clin. Med. 2020, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Vianna, E.P.; Ferreira, A.T.; Naffah-Mazzacoratti, M.G.; Sanabria, E.R.; Funke, M.; Cavalheiro, E.A.; Fernandes, M.J. Evidence that ATP participates in the pathophysiology of pilocarpine-induced temporal lobe epilepsy: Fluorimetric, immunohistochemical, and Western blot studies. Epilepsia 2002, 43 (Suppl. S5), 227–229. [Google Scholar] [CrossRef]
- Dona, F.; Ulrich, H.; Persike, D.S.; Conceicao, I.M.; Blini, J.P.; Cavalheiro, E.A.; Fernandes, M.J. Alteration of purinergic P2X4 and P2X7 receptor expression in rats with temporal-lobe epilepsy induced by pilocarpine. Epilepsy Res. 2009, 83, 157–167. [Google Scholar] [CrossRef]
- Engel, T.; Gomez-Villafuertes, R.; Tanaka, K.; Mesuret, G.; Sanz-Rodriguez, A.; Garcia-Huerta, P.; Miras-Portugal, M.T.; Henshall, D.C.; Diaz-Hernandez, M. Seizure suppression and neuroprotection by targeting the purinergic P2X7 receptor during status epilepticus in mice. FASEB J. 2012, 26, 1616–1628. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Pacheco, A.; Mesuret, G.; Sanz-Rodriguez, A.; Tanaka, K.; Mooney, C.; Conroy, R.; Miras-Portugal, M.T.; Diaz-Hernandez, M.; Henshall, D.C.; Engel, T. Increased neocortical expression of the P2X7 receptor after status epilepticus and anticonvulsant effect of P2X7 receptor antagonist A-438079. Epilepsia 2013, 54, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Hu, J.; Liu, X.; Deng, X. Increased expression of the P2X7 receptor in temporal lobe epilepsy: Animal models and clinical evidence. Mol. Med. Rep. 2019, 19, 5433–5439. [Google Scholar] [CrossRef]
- Morgan, J.; Alves, M.; Conte, G.; Menendez-Mendez, A.; de Diego-Garcia, L.; de Leo, G.; Beamer, E.; Smith, J.; Nicke, A.; Engel, T. Characterization of the Expression of the ATP-Gated P2X7 Receptor Following Status Epilepticus and during Epilepsy Using a P2X7-EGFP Reporter Mouse. Neurosci. Bull. 2020, 36, 1242–1258. [Google Scholar] [CrossRef]
- Ramirez-Fernandez, A.; Urbina-Trevino, L.; Conte, G.; Alves, M.; Rissiek, B.; Durner, A.; Scalbert, N.; Zhang, J.; Magnus, T.; Koch-Nolte, F.; et al. Deviant reporter expression and P2X4 passenger gene overexpression in the soluble EGFP BAC transgenic P2X7 reporter mouse model. Sci. Rep. 2020, 10, 19876. [Google Scholar] [CrossRef]
- Rozmer, K.; Gao, P.; Araujo, M.G.L.; Khan, M.T.; Liu, J.; Rong, W.; Tang, Y.; Franke, H.; Krugel, U.; Fernandes, M.J.S.; et al. Pilocarpine-Induced Status Epilepticus Increases the Sensitivity of P2X7 and P2Y1 Receptors to Nucleotides at Neural Progenitor Cells of the Juvenile Rodent Hippocampus. Cereb. Cortex 2017, 27, 3568–3585. [Google Scholar] [CrossRef] [PubMed]
- Conte, G.; Parras, A.; Alves, M.; Olla, I.; De Diego-Garcia, L.; Beamer, E.; Alalqam, R.; Ocampo, A.; Mendez, R.; Henshall, D.C.; et al. High concordance between hippocampal transcriptome of the mouse intra-amygdala kainic acid model and human temporal lobe epilepsy. Epilepsia 2020, 61, 2795–2810. [Google Scholar] [CrossRef] [PubMed]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.; Avoli, M. The pilocarpine model of temporal lobe epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef]
- Dhir, A. Pentylenetetrazol (PTZ) kindling model of epilepsy. Curr. Protoc. Neurosci. 2012, Chapter 9, Unit9 37. [Google Scholar] [CrossRef]
- Nieoczym, D.; Socala, K.; Wlaz, P. Evaluation of the Anticonvulsant Effect of Brilliant Blue G, a Selective P2X7 Receptor Antagonist, in the iv PTZ-, Maximal Electroshock-, and 6 Hz-Induced Seizure Tests in Mice. Neurochem. Res. 2017, 42, 3114–3124. [Google Scholar] [CrossRef]
- Huang, C.; Chi, X.S.; Li, R.; Hu, X.; Xu, H.X.; Li, J.M.; Zhou, D. Inhibition of P2X7 Receptor Ameliorates Nuclear Factor-Kappa B Mediated Neuroinflammation Induced by Status Epilepticus in Rat Hippocampus. J. Mol. Neurosci. 2017, 63, 173–184. [Google Scholar] [CrossRef]
- Dogan, E.; Aygun, H.; Arslan, G.; Rzayev, E.; Avci, B.; Ayyildiz, M.; Agar, E. The Role of NMDA Receptors in the Effect of Purinergic P2X7 Receptor on Spontaneous Seizure Activity in WAG/Rij Rats With Genetic Absence Epilepsy. Front. Neurosci. 2020, 14, 414. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kang, T.C. The P2X7 receptor-pannexin-1 complex decreases muscarinic acetylcholine receptor-mediated seizure susceptibility in mice. J. Clin. Investig. 2011, 121, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Welzel, L.; Schidlitzki, A.; Twele, F.; Anjum, M.; Loscher, W. A face-to-face comparison of the intra-amygdala and intrahippocampal kainate mouse models of mesial temporal lobe epilepsy and their utility for testing novel therapies. Epilepsia 2020, 61, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Amhaoul, H.; Ali, I.; Mola, M.; Van Eetveldt, A.; Szewczyk, K.; Missault, S.; Bielen, K.; Kumar-Singh, S.; Rech, J.; Lord, B.; et al. P2X7 receptor antagonism reduces the severity of spontaneous seizures in a chronic model of temporal lobe epilepsy. Neuropharmacology 2016, 105, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.; Koushal, P.; Reddy, B.V.; Deshmukh, R.; Kumar, P. Effect of GLT-1 modulator and P2X7 antagonists alone and in combination in the kindling model of epilepsy in rats. Epilepsy Behav. 2015, 48, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Amorim, R.P.; Araujo, M.G.L.; Valero, J.; Lopes-Cendes, I.; Pascoal, V.D.B.; Malva, J.O.; da Silva Fernandes, M.J. Silencing of P2X7R by RNA interference in the hippocampus can attenuate morphological and behavioral impact of pilocarpine-induced epilepsy. Purinergic Signal. 2017, 13, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.C.; Morse, R.P.; Scott, R.C.; Holmes, G.L.; Lenck-Santini, P.P. SCN1A mutations in Dravet syndrome: Impact of interneuron dysfunction on neural networks and cognitive outcome. Epilepsy Behav. 2012, 23, 177–186. [Google Scholar] [CrossRef]
- Kim, J.E.; Ryu, H.J.; Yeo, S.I.; Kang, T.C. P2X7 receptor regulates leukocyte infiltrations in rat frontoparietal cortex following status epilepticus. J. Neuroinflammation 2010, 7, 65. [Google Scholar] [CrossRef]
- Kim, J.E.; Ryu, H.J.; Kang, T.C. P2X7 receptor activation ameliorates CA3 neuronal damage via a tumor necrosis factor-alpha-mediated pathway in the rat hippocampus following status epilepticus. J. Neuroinflammation 2011, 8, 62. [Google Scholar] [CrossRef]
- Beamer, E.; Morgan, J.; Alves, M.; Menendez Mendez, A.; Morris, G.; Zimmer, B.; Conte, G.; de Diego-Garcia, L.; Alarcon-Vila, C.; Yiu Ng, N.K.; et al. Increased expression of the ATP-gated P2X7 receptor reduces responsiveness to anti-convulsants during status epilepticus in mice. Br. J. Pharmacol. 2022, 179, 2986–3006. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, B.D.S.; Kahoud, R.J.; McCarthy, C.B.; Howe, C.L. Inflammatory cytokine-induced changes in neural network activity measured by waveform analysis of high-content calcium imaging in murine cortical neurons. Sci. Rep. 2017, 7, 9037. [Google Scholar] [CrossRef] [PubMed]
- Noe, F.M.; Polascheck, N.; Frigerio, F.; Bankstahl, M.; Ravizza, T.; Marchini, S.; Beltrame, L.; Bandero, C.R.; Loscher, W.; Vezzani, A. Pharmacological blockade of IL-1beta/IL-1 receptor type 1 axis during epileptogenesis provides neuroprotection in two rat models of temporal lobe epilepsy. Neurobiol. Dis. 2013, 59, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Dyomina, A.V.; Zubareva, O.E.; Smolensky, I.V.; Vasilev, D.S.; Zakharova, M.V.; Kovalenko, A.A.; Schwarz, A.P.; Ischenko, A.M.; Zaitsev, A.V. Anakinra Reduces Epileptogenesis, Provides Neuroprotection, and Attenuates Behavioral Impairments in Rats in the Lithium-Pilocarpine Model of Epilepsy. Pharmaceuticals 2020, 13, 340. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, G.; Ishida, Y.; Kanou, K.; Suzuki, S.; Watanabe, Y.; Takamatsu, T.; Morichi, S.; Go, S.; Oana, S.; Yamazaki, T.; et al. Towards a Treatment for Neuroinflammation in Epilepsy: Interleukin-1 Receptor Antagonist, Anakinra, as a Potential Treatment in Intractable Epilepsy. Int. J. Mol. Sci. 2021, 22, 6282. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, S.; Koyama, R. Pro- and anti-epileptic roles of microglia. Neural Regen. Res. 2021, 16, 1369–1371. [Google Scholar] [CrossRef] [PubMed]
- Masuch, A.; Shieh, C.H.; van Rooijen, N.; van Calker, D.; Biber, K. Mechanism of microglia neuroprotection: Involvement of P2X7, TNFalpha, and valproic acid. Glia 2016, 64, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Strzelczyk, A.; Reif, P.S.; Schorlemmer, K.; Bauer, S.; Norwood, B.A.; Oertel, W.H.; Rosenow, F.; Strik, H.; Hamer, H.M. Minocycline as potent anticonvulsant in a patient with astrocytoma and drug resistant epilepsy. Seizure 2012, 21, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, E.M.; Means, R.E.; Michaud, M.; Madri, J.A.; Katz, S.G. Minocycline mitigates the effect of neonatal hypoxic insult on human brain organoids. Cell Death Dis. 2019, 10, 325. [Google Scholar] [CrossRef]
- Barros-Barbosa, A.R.; Fonseca, A.L.; Guerra-Gomes, S.; Ferreirinha, F.; Santos, A.; Rangel, R.; Lobo, M.G.; Correia-de-Sa, P.; Cordeiro, J.M. Up-regulation of P2X7 receptor-mediated inhibition of GABA uptake by nerve terminals of the human epileptic neocortex. Epilepsia 2016, 57, 99–110. [Google Scholar] [CrossRef]
- Barros-Barbosa, A.R.; Oliveira, A.; Lobo, M.G.; Cordeiro, J.M.; Correia-de-Sa, P. Under stressful conditions activation of the ionotropic P2X7 receptor differentially regulates GABA and glutamate release from nerve terminals of the rat cerebral cortex. Neurochem. Int. 2018, 112, 81–95. [Google Scholar] [CrossRef]
- Armstrong, J.N.; Brust, T.B.; Lewis, R.G.; MacVicar, B.A. Activation of presynaptic P2X7-like receptors depresses mossy fiber-CA3 synaptic transmission through p38 mitogen-activated protein kinase. J. Neurosci. 2002, 22, 5938–5945. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.F.; Tang, Y.; Illes, P. Astrocytic and Oligodendrocytic P2X7 Receptors Determine Neuronal Functions in the CNS. Front. Mol. Neurosci. 2021, 14, 641570. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, J.; Watters, O.; de Diego-Garcia, L.; Mendez, A.M.; Alves, M.; Dinkel, K.; Hamacher, M.; Prehn, J.H.M.; Henshall, D.C.; Engel, T. Functional expression of the ATP-gated P2X7 receptor in human iPSC-derived astrocytes. Purinergic Signal. 2023, 20, 303–309. [Google Scholar] [CrossRef]
- Vezzani, A.; Ravizza, T.; Bedner, P.; Aronica, E.; Steinhauser, C.; Boison, D. Astrocytes in the initiation and progression of epilepsy. Nat. Rev. Neurol. 2022, 18, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Knowles, J.K.; Xu, H.; Soane, C.; Batra, A.; Saucedo, T.; Frost, E.; Tam, L.T.; Fraga, D.; Ni, L.; Villar, K.; et al. Maladaptive myelination promotes generalized epilepsy progression. Nat. Neurosci. 2022, 25, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, Y.; Wang, J.; Dong, L.; Liu, S.; Li, S.; Wu, Q. Purinergic signaling: A gatekeeper of blood-brain barrier permeation. Front. Pharmacol. 2023, 14, 1112758. [Google Scholar] [CrossRef] [PubMed]
- Grinalds, M.S.; Yoder, C.; Krauss, Z.; Chen, A.M.; Rhoney, D.H. Scoping review of rational polytherapy in patients with drug-resistant epilepsy. Pharmacotherapy 2023, 43, 53–84. [Google Scholar] [CrossRef]
- Goldberg, E.M.; Coulter, D.A. Mechanisms of epileptogenesis: A convergence on neural circuit dysfunction. Nat. Rev. Neurosci. 2013, 14, 337–349. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Janjoppi, L.; Bonone, F.M.; Pansani, A.P.; da Silva, A.V.; Scorza, F.A.; Cavalheiro, E.A. Whole transcriptome analysis of the hippocampus: Toward a molecular portrait of epileptogenesis. BMC Genom. 2010, 11, 230. [Google Scholar] [CrossRef]
- Pitkanen, A.; Lukasiuk, K.; Dudek, F.E.; Staley, K.J. Epileptogenesis. Cold Spring Harb. Perspect. Med. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Mouri, G.; Jimenez-Mateos, E.; Engel, T.; Dunleavy, M.; Hatazaki, S.; Paucard, A.; Matsushima, S.; Taki, W.; Henshall, D.C. Unilateral hippocampal CA3-predominant damage and short latency epileptogenesis after intra-amygdala microinjection of kainic acid in mice. Brain Res. 2008, 1213, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, C.M.; Barron, J.C.; Brymer, K.J.; Barnes, J.R.; Nafar, F.; Parsons, M.P. The Effect of GLT-1 Upregulation on Extracellular Glutamate Dynamics. Front. Cell Neurosci. 2021, 15, 661412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Xu, J.; Sun, Q.; Yu, F.; Cheng, J.; Peng, B.; Liu, W.; Xiao, Z.; Yin, J.; et al. Inhibition of DPP4 enhances inhibitory synaptic transmission through activating the GLP-1/GLP-1R signaling pathway in a rat model of febrile seizures. Biochem. Pharmacol. 2018, 156, 78–85. [Google Scholar] [CrossRef]
- Jamali-Raeufy, N.; Barati, H.; Baluchnejadmojarad, T.; Roghani, M.; Goudarzi, M. Combination therapy with dipeptidyl peptidase-4 and P2X7 purinoceptor inhibitors gives rise to antiepileptic effects in rats. J. Chem. Neuroanat. 2020, 110, 101855. [Google Scholar] [CrossRef] [PubMed]
- Recourt, K.; de Boer, P.; van der Ark, P.; Benes, H.; van Gerven, J.M.A.; Ceusters, M.; van Nueten, L.; Drevets, W.C.; Bhatacharya, A.; Browning, M.; et al. Characterization of the central nervous system penetrant and selective purine P2X7 receptor antagonist JNJ-54175446 in patients with major depressive disorder. Transl. Psychiatry 2023, 13, 266. [Google Scholar] [CrossRef]
- Benzaquen, J.; Heeke, S.; Janho Dit Hreich, S.; Douguet, L.; Marquette, C.H.; Hofman, P.; Vouret-Craviari, V. Alternative splicing of P2RX7 pre-messenger RNA in health and diseases: Myth or reality? Biomed. J. 2019, 42, 141–154. [Google Scholar] [CrossRef]
- Lee, D.S.; Kim, J.E. Protein disulfide isomerase-mediated S-nitrosylation facilitates surface expression of P2X7 receptor following status epilepticus. J. Neuroinflammation 2021, 18, 14. [Google Scholar] [CrossRef]
- Morgan, J.; Moreno, O.; Alves, M.; Baz, Z.; Menendez Mendez, A.; Leister, H.; Melia, C.; Smith, J.; Visekruna, A.; Nicke, A.; et al. Increased uptake of the P2X7 receptor radiotracer (18) F-JNJ-64413739 in the brain and peripheral organs according to the severity of status epilepticus in male mice. Epilepsia 2023, 64, 511–523. [Google Scholar] [CrossRef]
- Fu, Z.; Lin, Q.; Xu, Z.; Fu, W.; Shi, D.; Cheng, Y.; Yang, T.; Liu, G.; Shi, H.; Cheng, D. Longitudinal Positron Emission Tomography Imaging with P2X7 Receptor-Specific Radioligand (18)F-FTTM in a Kainic Acid Rat Model of Temporal Lobe Epilepsy. ACS Chem. Neurosci. 2022, 13, 3512–3522. [Google Scholar] [CrossRef]
- Conte, G.; Menendez-Mendez, A.; Bauer, S.; El-Naggar, H.; Alves, M.; Nicke, A.; Delanty, N.; Rosenow, F.; Henshall, D.C.; Engel, T. Circulating P2X7 Receptor Signaling Components as Diagnostic Biomarkers for Temporal Lobe Epilepsy. Cells 2021, 10, 2444. [Google Scholar] [CrossRef]
- Hermann, B.; Seidenberg, M.; Sager, M.; Carlsson, C.; Gidal, B.; Sheth, R.; Rutecki, P.; Asthana, S. Growing old with epilepsy: The neglected issue of cognitive and brain health in aging and elder persons with chronic epilepsy. Epilepsia 2008, 49, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Sanz, J.M.; Falzoni, S.; Morieri, M.L.; Passaro, A.; Zuliani, G.; Di Virgilio, F. Association of Hypomorphic P2X7 Receptor Genotype With Age. Front. Mol. Neurosci. 2020, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Suller Marti, A.; Bellosta Diago, E.; Vinueza Buitron, P.; Velazquez Benito, A.; Santos Lasaosa, S.; Mauri Llerda, J.A. Epilepsy in elderly patients: Does age of onset make a difference? Neurología 2022, 37, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Fuller, S.J.; Stokes, L.; Skarratt, K.K.; Gu, B.J.; Wiley, J.S. Genetics of the P2X7 receptor and human disease. Purinergic Signal. 2009, 5, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Emsley, H.C.; Appleton, R.E.; Whitmore, C.L.; Jury, F.; Lamb, J.A.; Martin, J.E.; Ollier, W.E.; de la Morandiere, K.P.; Southern, K.W.; Allan, S.M. Variations in inflammation-related genes may be associated with childhood febrile seizure susceptibility. Seizure 2014, 23, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Nistri, A.; Khakh, B.S.; Giniatullin, R. ATP-gated P2X receptors in health and disease. Front. Cell Neurosci. 2014, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, D.M.; Schorge, S.; Walker, M.C.; Wykes, R.C. Gene therapy in epilepsy-is it time for clinical trials? Nat. Rev. Neurol. 2014, 10, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Papa, S.; Ferrari, R.; De Paola, M.; Rossi, F.; Mariani, A.; Caron, I.; Sammali, E.; Peviani, M.; Dell’Oro, V.; Colombo, C.; et al. Polymeric nanoparticle system to target activated microglia/macrophages in spinal cord injury. J. Control Release 2014, 174, 15–26. [Google Scholar] [CrossRef]
- Guo, S.; Cazarez-Marquez, F.; Jiao, H.; Foppen, E.; Korpel, N.L.; Grootemaat, A.E.; Liv, N.; Gao, Y.; van der Wel, N.; Zhou, B.; et al. Specific Silencing of Microglial Gene Expression in the Rat Brain by Nanoparticle-Based Small Interfering RNA Delivery. ACS Appl. Mater. Interfaces 2022, 14, 5066–5079. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakku Sivakumar, D.; Jain, K.; Alfehaid, N.; Wang, Y.; Teng, X.; Fischer, W.; Engel, T. The Purinergic P2X7 Receptor as a Target for Adjunctive Treatment for Drug-Refractory Epilepsy. Int. J. Mol. Sci. 2024, 25, 6894. https://doi.org/10.3390/ijms25136894
Thakku Sivakumar D, Jain K, Alfehaid N, Wang Y, Teng X, Fischer W, Engel T. The Purinergic P2X7 Receptor as a Target for Adjunctive Treatment for Drug-Refractory Epilepsy. International Journal of Molecular Sciences. 2024; 25(13):6894. https://doi.org/10.3390/ijms25136894
Chicago/Turabian StyleThakku Sivakumar, Divyeshz, Krishi Jain, Noura Alfehaid, Yitao Wang, Xinchen Teng, Wolfgang Fischer, and Tobias Engel. 2024. "The Purinergic P2X7 Receptor as a Target for Adjunctive Treatment for Drug-Refractory Epilepsy" International Journal of Molecular Sciences 25, no. 13: 6894. https://doi.org/10.3390/ijms25136894
APA StyleThakku Sivakumar, D., Jain, K., Alfehaid, N., Wang, Y., Teng, X., Fischer, W., & Engel, T. (2024). The Purinergic P2X7 Receptor as a Target for Adjunctive Treatment for Drug-Refractory Epilepsy. International Journal of Molecular Sciences, 25(13), 6894. https://doi.org/10.3390/ijms25136894