
The Construction of an Environmentally Friendly Super-Secreting Strain of Bacillus subtilis through Systematic Modulation of Its Secretory Pathway Using the CRISPR-Cas9 System

Abstract
1. Introduction
2. Results
2.1. The Effect of Single and Tandem Promoters on the Expression of Recombinant AmyQ α-Amylase in B. subtilis
2.2. Secretory Expression of AmyQ via the Non-Classical (Tat) Secretion Pathway
2.3. Maximizing amyQ Expression by Inserting Multiple amyQ Gene Copies into the B. subtilis Chromosome
2.4. Enhancing AmyQ Secretion in the BSQ6 Strain through Genome Modification of the Sec Pathway and Other Potential Post-Transcriptional Bottlenecks
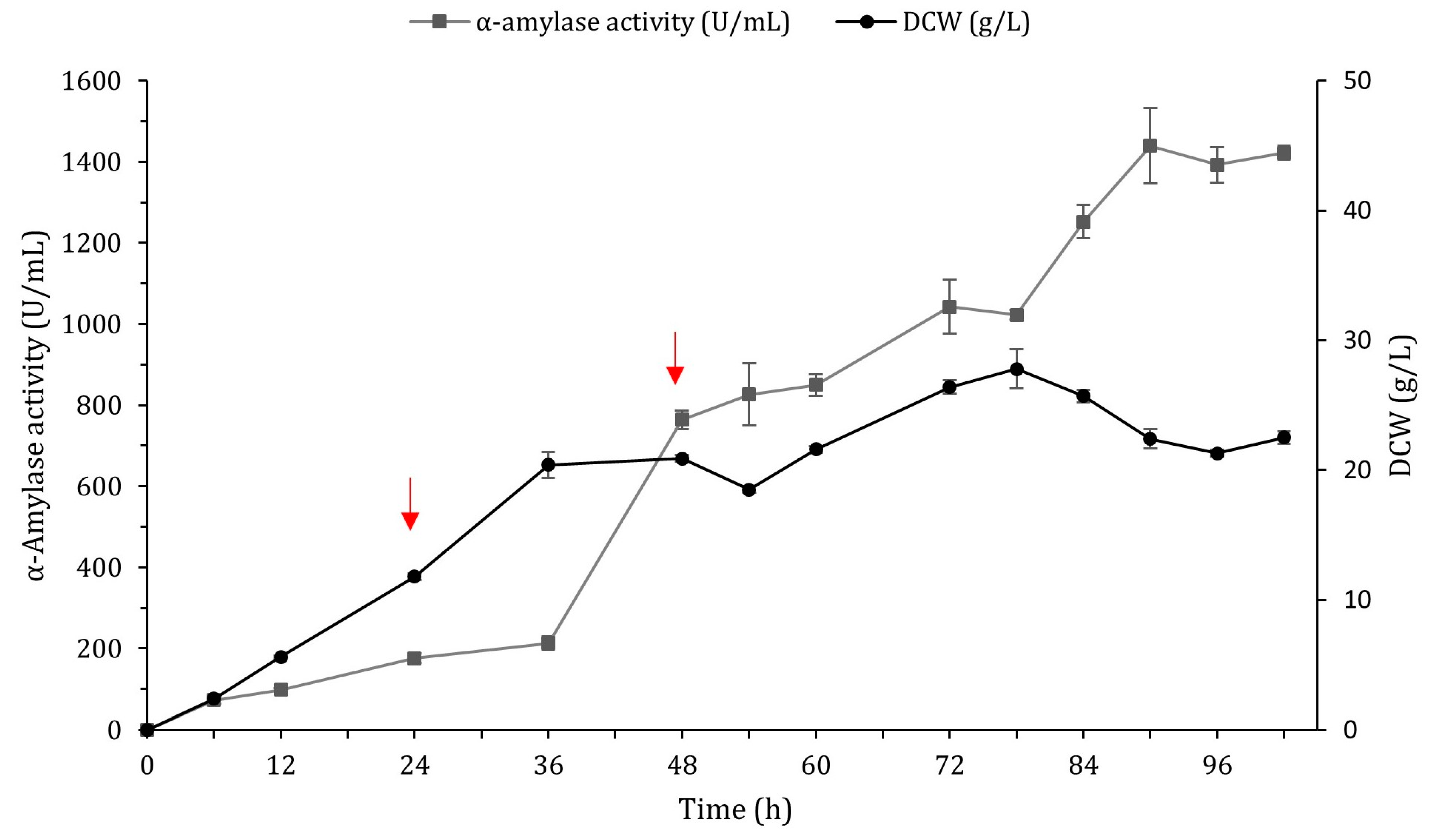
2.5. Scale-Up of α-Amylase Production in a 3 L Fermentor
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Culture Conditions
4.2. DNA Manipulations
4.3. Construction of the Integration Vectors for amyQ Overexpression and Sec Pathway Modulation
4.4. Cloning of sgRNA
4.5. Cloning of the Editing Templates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Characteristics | Reference |
|---|---|---|
| pBS2EXylRPxylA | Plasmid containing the xylose-inducible promoter/xylose repressor system | [70] |
| pJOE8999.1 | PmanP-cas9, pUC, pE194ts, kanr | [28] |
| pJOE2 | amyE gene knock-out plasmid derived from pJOE8999.1 | This work |
| pJOE3 | amyQ gene (PspoVG) knock-in plasmid derived from pJOE8999.1. Integration at the spoVG locus. | This work |
| pJOE4 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the amyE locus. | This work |
| pJOE5 | amyQ gene (PspoVG-PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the spoVG locus. | This work |
| pJOE6 | amyQ gene (SPywbN’) knock-in plasmid derived from pJOE8999.1. Integration at the ywbN locus. | This work |
| pJOE7 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the pksG locus. | This work |
| pJOE8 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the ppsE locus. | This work |
| pJOE9 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the cotB locus. | This work |
| pJOE10 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the ylbP locus. | This work |
| pJOE11 | amyQ gene (PamyQ-Pcry3A) knock-in plasmid derived from pJOE8999.1. Integration at the veg locus. | This work |
| pJOE12 | hag gene knock-out plasmid derived from pJOE8999.1 | This work |
| pJOE13 | pssA gene knock-out plasmid derived from pJOE8999.1 | This work |
| pJOE14 | yusX gene knock-out plasmid derived from pJOE8999.1 | This work |
| pJOE15 | hrcA gene knock-out plasmid derived from pJOE8999.1 | This work |
| pJOE16 | sipT gene knock-in plasmid derived from pJOE8999.1. Integration at the thrC locus. | This work |
| pJOE17 | rasP gene knock-in plasmid derived from pJOE8999.1. Integration at the spoVG locus. | This work |
| pJOE18 | sppA gene knock-in plasmid derived from pJOE8999.1. Integration at the yqeZ locus. | This work |
| pJOE19 | secYEG artificial operon knock-in plasmid derived from pJOE8999.1. Integration at the sigX locus. | This work |
| pJOE20 | gudB gene restoration plasmid derived from pJOE8999.1 | This work |
4.6. Plasmid Curing and Genome Edition Modifications
4.7. Quantification of α-Amylase Activity in Shake Flasks
4.8. SDS-PAGE Analysis
4.9. Dry Cell Weight
4.10. Three-Liter Fermentor Experiments
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- van Dijl, J.M.; Hecker, M. Bacillus subtilis: From soil bacterium to super-secreting cell factory. Microb. Cell Fact. 2013, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, X.; Lu, P. Bottlenecks in the expression and secretion of heterologous proteins in Bacillus subtilis. Res. Microb. 2004, 155, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wu, G. Bottleneck in secretion of α-amylase in Bacillus subtilis. Microb. Cell Fact. 2017, 16, 124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Ye, B.; Zhang, Q.; Cheng, D.; Zhou, C.; Cheng, S.; Yan, X. Construction of second generation protease-deficient hosts of Bacillus subtilis for secretion of foreign proteins. Biotechnol. Bioeng. 2019, 116, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Schumann, W. Production of recombinant proteins in Bacillus subtilis. Adv. Appl. Microbiol. 2007, 62, 137–189. [Google Scholar] [PubMed]
- Wang, Y.; Shi, Y.; Hu, L.; Du, G.; Chen, J.; Kang, Z. Synthetic and systems biotechnology engineering strong and stress-responsive promoters in Bacillus subtilis by interlocking sigma factor binding motifs. Synth. Syst. Biotechnol. 2019, 4, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, J.; Filluelo, O.; Zeigler, D.R.; Picart, P. Barriers to simultaneous multilocus integration in Bacillus subtilis tumble down: Development of a straightforward screening method for the colorimetric detection of one-step multiple gene insertion using the CRISPR-Cas9 system. Microb. Cell Fact. 2023, 22, 21. [Google Scholar] [CrossRef]
- Watzlawick, H.; Altenbuchner, J. Multiple integration of the gene ganA into the Bacillus subtilis chromosome for enhanced β-galactosidase production using the CRISPR/Cas9 system. AMB Express 2019, 9, 158. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, B.; Cheng, S.; Zhao, L.; Liu, Y.; Jiang, J.; Yan, X. Promoter engineering enables overproduction of foreign proteins from a single copy expression cassette in Bacillus subtilis. Microb. Cell Fact. 2019, 18, 111. [Google Scholar] [CrossRef]
- Chen, J.; Fu, G.; Gai, Y.; Zheng, P.; Zhang, D.; Wen, J. Combinatorial Sec pathway analysis for improved heterologous protein secretion in Bacillus subtilis: Identification of bottlenecks by systematic gene overexpression. Microb. Cell Fact. 2015, 14, 92. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Ye, R.; Wu, X.C.; Ng, S.C.; Wong, S.L. Enhanced secretory production of a single-chain antibody fragment from Bacillus subtilis by coproduction of molecular chaperones. J. Bacteriol. 1998, 180, 2830–2835. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Su, L.; Li, N.; Wu, J. Enhanced extracellular expression of Bacillus stearothermophilus α-amylase in Bacillus subtilis through signal peptide optimization, chaperone overexpression and α-amylase mutant selection. Microb. Cell Fact. 2019, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Kolkman, M.A.B.; Van Der Ploeg, R.; Bertels, M.; Van Dijk, M.; Van Der Laan, J.; Van Dijl, J.M.; Ferrari, E. The twin-arginine signal peptide of Bacillus subtilis YwbN can direct either Tat- or Sec-dependent secretion of different cargo proteins: Secretion of active subtilisin via the B. subtilis Tat pathway. Appl. Environ. Microbiol. 2008, 74, 7507–7513. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.H.; Cao, L.C.; Kong, W.; Wang, Z.J.; Liu, Y.H. Efficient secretion of the β-galactosidase Bgal1-3 via both Tat-dependent and Tat-independent pathways in Bacillus subtilis. J. Agric. Food Chem. 2016, 64, 5708–5716. [Google Scholar] [CrossRef] [PubMed]
- Mulder, K.C.L.; Bandola, J.; Schumann, W. Construction of an artificial secYEG operon allowing high level secretion of α-amylase. Protein Expr. Purif. 2013, 89, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Oswald, J.; Njenga, R.; Natriashvili, A.; Sarmah, P.; Koch, H.G. The Dynamic SecYEG Translocon. Front. Mol. Biosci. 2021, 8, 664241. [Google Scholar] [CrossRef]
- Vitikainen, M.; Pummi, T.; Airaksinen, U.; Wahlström, E.; Wu, H.; Sarvas, M.; Kontinen, V.P. Quantitation of the capacity of the secretion apparatus and requirement for PrsA in growth and secretion of α-amylase in Bacillus subtilis. J. Bacteriol. 2001, 183, 1881–1890. [Google Scholar] [CrossRef]
- Cai, D.; Wang, H.; He, P.; Zhu, C.; Wang, Q.; Wei, X.; Nomura, C.T.; Chen, S. A novel strategy to improve protein secretion via overexpression of the SppA signal peptide peptidase in Bacillus licheniformis. Microb. Cell Fact. 2017, 16, 70. [Google Scholar] [CrossRef]
- Neef, J.; Bongiorni, C.; Goosens, V.J.; Schmidt, B.; van Dijl, J.M. Intramembrane protease RasP boosts protein production in Bacillus. Microb. Cell Fact. 2017, 16, 57. [Google Scholar] [CrossRef]
- Cao, H.; Heel, A.J.; Ahmed, H.; Mols, M.; Kuipers, O.P. Cell surface engineering of Bacillus subtilis improves production yields of heterologously expressed α-amylases. Microb. Cell Fact. 2017, 16, 56. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Ganuza, A.; Antelo-Varela, M.; Mouritzen, J.C.; Bartel, J.; Becher, D.; Gjermansen, M.; Hallin, P.F.; Appel, K.F.; Kilstrup, M.; Rasmussen, M.D.; et al. Identification and optimization of PrsA in Bacillus subtilis for improved yield of amylase. Microb. Cell Fact. 2019, 18, 158. [Google Scholar] [CrossRef]
- Vojnovic, S.; Aleksic, I.; Ilic-Tomic, T.; Stevanovic, M.; Nikodinovic-Runic, J. Bacillus and Streptomyces spp. as hosts for production of industrially relevant enzymes. Appl. Microbiol. Biotechnol. 2024, 108, 185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Su, L.; Wu, J. Recent Advances in Recombinant Protein Production by Bacillus subtilis. Annu. Rev. Food Sci. Technol. 2020, 11, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.C.; Han, L.L.; Lu, Y.B.; Feng, H. Construction of a high-expression system in Bacillus through transcriptomic profiling and promoter engineering. Microorganisms 2020, 8, 1030. [Google Scholar] [CrossRef]
- Widner, B.; Thomas, M.; Sternberg, D.; Lammon, D.; Behr, R.; Sloma, A. Development of marker-free strains of Bacillus subtilis capable of secreting high levels of industrial enzymes. J. Ind. Microbiol. Biotechnol. 2000, 25, 204–212. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Y.; Liu, Y.; Li, J.; Du, G.; Lv, X.; Liu, L. A genetic toolkit for efficient production of secretory protein in Bacillus subtilis. Bioresour. Technol. 2022, 363, 127885. [Google Scholar] [CrossRef]
- Altenbuchner, J. Editing of the Bacillus subtilis genome by the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2016, 82, 5421–5427. [Google Scholar] [CrossRef]
- Gan, T.; Fang, J.; Wang, Y.; Liu, K.; Sang, Y.; Chen, H.; Lu, Y.; Zhu, L.; Chen, X. Promoter engineering for efficient production of sucrose phosphorylase in Bacillus subtilis and its application in enzymatic synthesis of 2-O-α-D-glucopyranosyl-L-ascorbic acid. Enzym. Microb. Technol. 2023, 169, 110267. [Google Scholar] [CrossRef]
- Guan, C.; Cui, W.; Cheng, J.; Liu, R.; Liu, Z.; Zhou, L.; Zhou, Z. Construction of a highly active secretory expression system via an engineered dual promoter and a highly efficient signal peptide in Bacillus subtilis. New Biotechnol. 2016, 33, 372–379. [Google Scholar] [CrossRef]
- Jun, J.S.; Jeong, H.E.; Hong, K.W. Exploring and engineering novel strong promoters for high-level protein expression in Bacillus subtilis DB104 through transcriptome analysis. Microorganisms 2023, 11, 2929. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.J.; Wang, Y.H.; Liu, L.; Bai, L.L.; Ban, R. Production enhancement of the extracellular lipase LipA in Bacillus subtilis: Effects of expression system and Sec pathway components. Protein Expr. Purif. 2018, 142, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Frain, K.M.; Robinson, C.; Maarten Van Dijl, J. Transport of folded proteins by the Tat System. Protein J. 2019, 38, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Jongbloed, J.D.H.; Grieger, U.; Antelmann, H.; Hecker, M.; Nijland, R.; Bron, S.; Van Dijl, J.M. Two minimal Tat translocases in Bacillus. Mol. Microbiol. 2004, 54, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.; Berks, B.C. The twin-arginine translocation (Tat) protein export pathway. Nat. Rev. Microbiol. 2012, 10, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zuo, Z.; Xu, Y.; Song, C.; Jiang, H.; Qiao, C.; Xu, P.; Zhou, Q.; Yang, C. Twin-arginine signal peptide of Bacillus subtilis YwbN can direct Tat-dependent secretion of methyl parathion hydrolase. J. Agric. Food Chem. 2014, 62, 2913–2918. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Yang, Y.; Liu, X.; Li, D.; Li, J.; Guo, X.; Zhou, Z. Secretory expression of a heterologous protein, Aiio-AIO6BS, in Bacillus subtilis via a non-classical secretion pathway. Biochem. Biophys. Res. Commun. 2016, 478, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ma, Y.; Zhao, Y.; Shen, W.; Chen, X. Systematic engineering of transport and transcription to boost alkaline α-amylase production in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2020, 104, 2973–2985. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, T.; Kadoya, R.; Endo, K.; Tohata, M.; Sawada, K.; Liu, S.; Ozawa, T.; Kodama, T.; Kakeshita, H.; Kageyama, Y.; et al. Enhanced recombinant protein productivity by genome reduction in Bacillus subtilis. DNA Res. 2008, 15, 73–81. [Google Scholar] [CrossRef]
- Zhu, B.; Stülke, J. SubtiWiki in 2018: From genes and proteins to functional network annotation of the model organism Bacillus subtilis. Nucleic Acids Res. 2018, 46, D743–D748. [Google Scholar] [CrossRef]
- Young, M. Gene amplification in Bacillus subtilis. Microbiology 1984, 130, 1613–1621. [Google Scholar] [CrossRef]
- Neef, J.; Bongiorni, C.; Schmidt, B.; Goosens, V.J.; van Dijl, J.M. Relative contributions of non-essential Sec pathway components and cell envelope-associated proteases to high-level enzyme secretion by Bacillus subtilis. Microb. Cell Fact. 2020, 19, 6298–6305. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R.; Sonenshein, A.L. Role and regulation of Bacillus subtilis glutamate dehydrogenase genes. J. Bacteriol. 1998, 180, 6298–6305. [Google Scholar] [CrossRef]
- Simonen, M.; Palva, I. Protein secretion in Bacillus species. Microbiol. Rev. 1993, 57, 109. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, J.; Geng, Y.; Li, Y.; Wang, Q.; Liang, Q.; Qi, Q. A strategy of gene overexpression based on tandem repetitive promoters in Escherichia coli. Microb. Cell Fact. 2012, 11, 19. [Google Scholar] [CrossRef]
- Yang, S.; Du, G.; Chen, J.; Kang, Z. Characterization and application of endogenous phase-dependent promoters in Bacillus subtilis. Appl. Microbiol. Biotechnol. 2017, 101, 4151–4161. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Pan, J.G.; Park, S.H.; Choi, S.K. Development of a stationary phase-specific autoinducible expression system in Bacillus subtilis. J. Biotechnol. 2010, 149, 16–20. [Google Scholar] [CrossRef]
- Mathy, N.; Bé, L.; Pellegrini, O.; Daou, R.; Wen, T.; Condon, C.N. 5′-to-3′ exoribonuclease activity in bacteria: Role of RNase J1 in rRNA maturation and 5′ stability of mRNA. Cell 2007, 129, 681–692. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; Wang, B.; Pan, L. Efficient production of extracellular pullulanase in Bacillus subtilis ATCC6051 using the host strain construction and promoter optimization expression system. Microb. Cell Fact. 2018, 17, 163. [Google Scholar] [CrossRef]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef]
- Tjalsma, H.; Bolhuis, A.; Jongbloed, J.D.H.; Bron, S.; van Dijl, J.M. Signal peptide-dependent protein transport in Bacillus subtilis: A genome-based survey of the secretome. Microbiol. Mol. Biol. Rev. 2000, 64, 515–547. [Google Scholar] [CrossRef]
- Yuan, G.; Wong, S.L. Isolation and characterization of Bacillus subtilis groE regulatory mutants: Evidence for orf39 in the dnaK operon as a repressor gene in regulating the expression of both groE and dnaK. J. Bacteriol. 1995, 177, 6462. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.P.; Van Der Ploeg, R.; Eijlander, R.T.; Nenninger, A.; Mendel, S.; Rozeboom, R.; Kuipers, O.P.; Van Dijl, J.M.; Robinson, C. The twin-arginine translocation (Tat) systems from Bacillus subtilis display a conserved mode of complex organization and similar substrate recognition requirements. FEBS J. 2009, 276, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Westers, L.; Westers, H.; Quax, W.J. Bacillus subtilis as cell factory for pharmaceutical proteins: A biotechnological approach to optimize the host organism. Biochim. Biophys. Acta–Mol. Cell Res. 2004, 1694, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.L.; Xu, Z.R.; Li, W.F.; Shuai, J.B.; Lu, P.; Hu, C.X. Protein secretion pathways in Bacillus subtilis: Implication for optimization of heterologous protein secretion. Biotechnol. Adv. 2007, 25, 1–12. [Google Scholar]
- Bolhuis, A.; Matzen, A.; Hyyryläinen, H.L.; Kontinen, V.P.; Meima, R.; Chapuis, J.; Venema, G.; Bron, S.; Freudl, R.; Van Dijl, J.M. Signal peptide peptidase- and ClpP-like proteins of Bacillus subtilis required for efficient translocation and processing of secretory proteins. J. Biol. Chem. 1999, 274, 24585–24592. [Google Scholar] [CrossRef]
- Yu, X.; Xu, J.; Liu, X.; Chu, X.; Wang, P.; Tian, J.; Wu, N.; Fan, Y. Identification of a highly efficient stationary phase promoter in Bacillus subtilis. Sci. Rep. 2015, 5, 18405. [Google Scholar] [CrossRef]
- Nielsen, A.K.; Rasmussen, M.D. Mutated Prokaryotic Cells with High Secretion-Levels. U.S. Patent No. 7,951,556, 31 May 2011. [Google Scholar]
- Buescher, J.M.; Liebermeister, W.; Jules, M.; Uhr, M.; Muntel, J.; Botella, E.; Hessling, B.; Kleijn, R.J.; Le Chat, L.; Lecointe, F.; et al. Global network reorganization during dynamic adaptations of Bacillus subtilis metabolism. Science 2012, 335, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, A.; Tian, R.; Li, J.; Liu, L.; Du, G. Developing rapid growing Bacillus subtilis for improved biochemical and recombinant protein production. Metab. Eng. Commun. 2020, 11, e00141. [Google Scholar] [CrossRef]
- Colletti, P.F.; Goyal, Y.; Varman, A.M.; Feng, X.; Wu, B.; Tang, Y.J. Evaluating factors that influence microbial synthesis yields by linear regression with numerical and ordinal variables. Biotechnol. Bioeng. 2011, 108, 893–901. [Google Scholar] [CrossRef]
- Gunka, K.; Stannek, L.; Care, R.A.; Commichau, F.M. Selection-driven accumulation of suppressor mutants in Bacillus subtilis: The apparent high mutation frequency of the cryptic gudB gene and the rapid clonal expansion of gudB+ suppressors are due to growth under selection. PLoS ONE 2013, 8, e00141. [Google Scholar] [CrossRef] [PubMed]
- Palva, I. Molecular cloning of α-amylase gene from Bacillus amyloliquefaciens and its expression in B. subtilis. Gene 1982, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Lulko, A.T.; Veening, J.W.; Buist, G.; Smits, W.K.; Blom, E.J.; Beekman, A.C.; Bron, S.; Kuipers, O.P. Production and secretion stress caused by overexpression of heterologous alpha-amylase leads to inhibition of sporulation and a prolonged motile phase in Bacillus subtilis. Appl. Environ. Microbiol. 2007, 73, 5354–5362. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.Y.; Chau, V.W.; Landas, J.A.; Pang, Y. Preparation of calcium competent Escherichia coli and heat-shock transformation. JEMI Methods 2017, 1, 22–25. [Google Scholar]
- Yasbin, R.E.; Wilson, G.A.; Young, F.E. Transformation and transfection in lysogenic strains of Bacillus subtilis: Evidence for selective induction of prophage in competent cells. J. Bacteriol. 1975, 121, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.W.; Sambrook, J. Molecular cloning: A laboratory manual. Cold Spring Harb. Lab. 2001, 1, 112. [Google Scholar]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef]
- Wu, G.; Drufva, E.; Wu, K. Fast genome editing in Bacillus subtilis. Eng. Life Sci. 2019, 19, 471–477. [Google Scholar] [CrossRef]
- Popp, P.F.; Dotzler, M.; Radeck, J.; Bartels, J.; Mascher, T. The Bacillus BioBrick Box 2.0: Expanding the genetic toolbox for the standardized work with Bacillus subtilis. Sci. Rep. 2017, 7, 15058. [Google Scholar] [CrossRef]

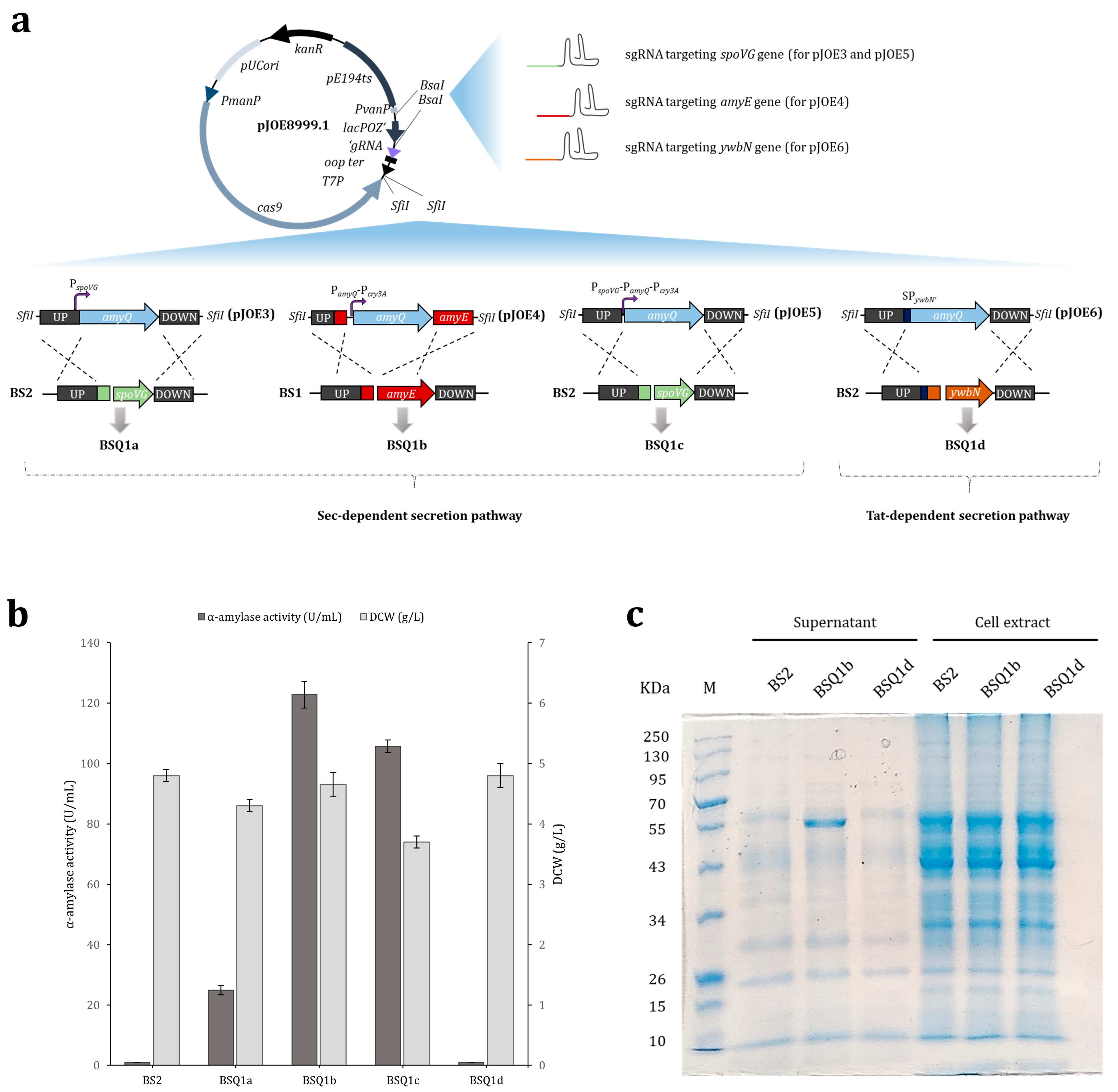
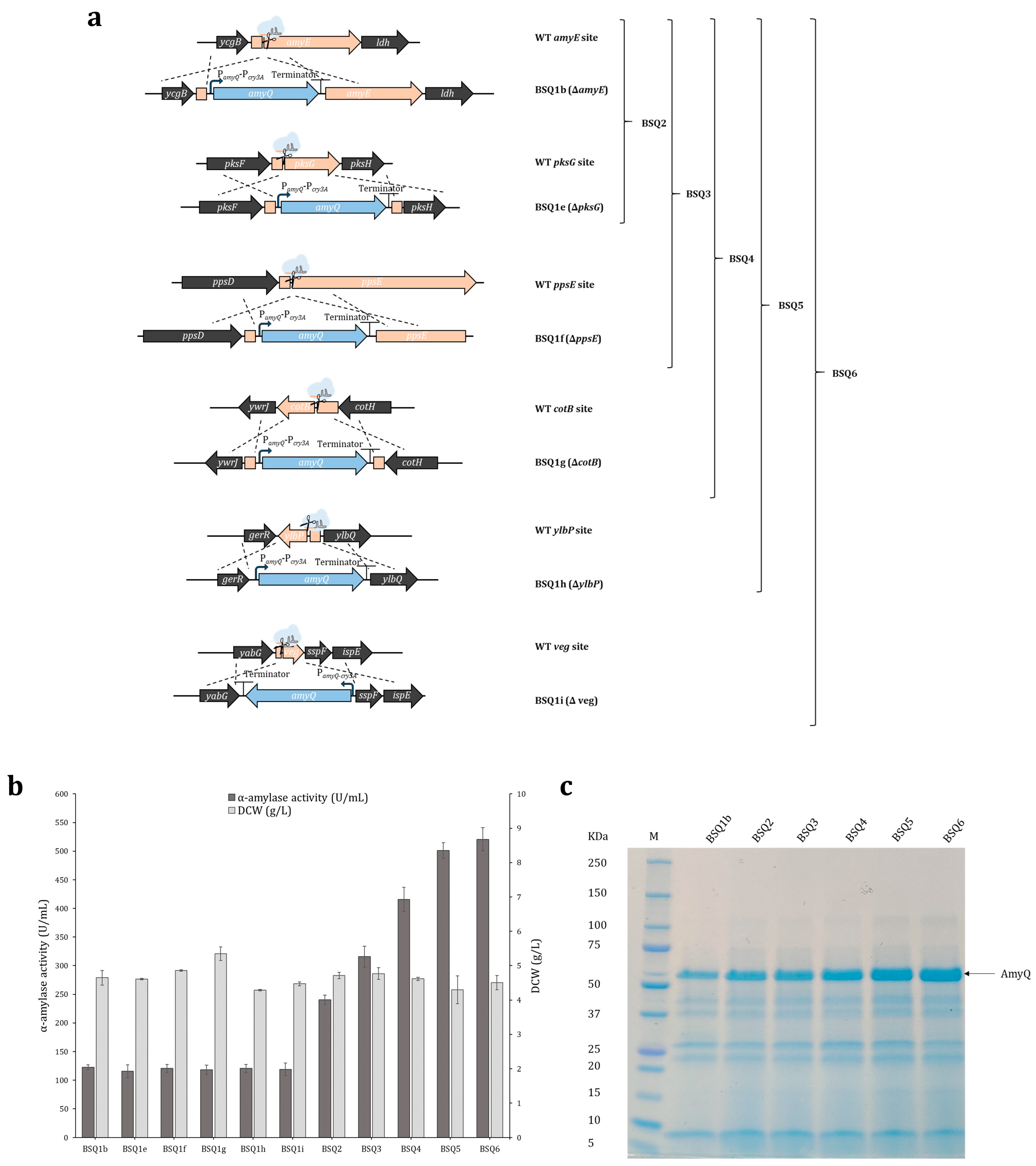
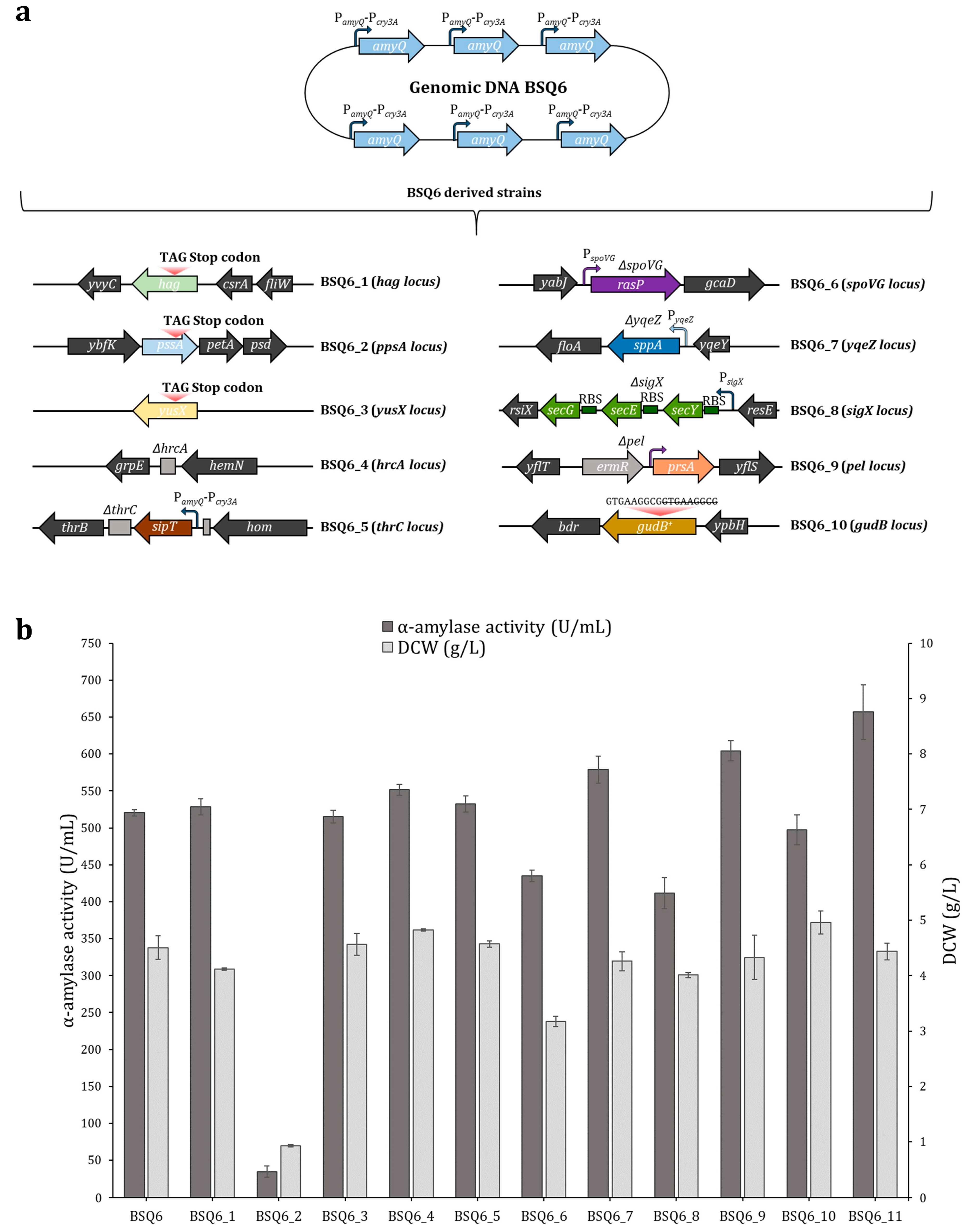
| Strain | Amylase Activity (U/mL) | Cumulative Increase a | DCW (g/L) | |
|---|---|---|---|---|
| Total Fold Change | p-Value b | |||
| BSQ1a | 24.9 ± 1.5 | 1 | 4.3 ± 0.1 | |
| BSQ1b | 122.8 ± 4.4 | 4.9 | <0.001 | 4.7 ± 0.2 |
| BSQ2 | 240.4 ± 8.0 | 9.7 | <0.001 | 4.7 ± 0.1 |
| BSQ3 | 315.7 ± 18.4 | 12.7 | 0.003 | 4.8 ± 0.2 |
| BSQ4 | 415.7 ± 21.1 | 16.7 | 0.003 | 4.6 ± 0.1 |
| BSQ5 | 501.2 ± 13.5 | 20.2 | 0.004 | 4.3 ± 0.4 |
| BSQ6 | 520.6 ± 20.2 | 20.9 | 0.3 | 4.5 ± 0.2 |
| BSQ6_7 | 579.0 ± 25.4 | 23.3 | 0.04 | 4.3 ± 0.3 |
| BSQ6_9 | 604.3 ± 13.5 | 24.3 | 0.004 c | 4.3 ± 0.2 |
| BSQ6_11 | 656.8 ± 24.3 | 26.4 | 0.03 | 4.4 ± 0.01 |
| BSQ6_11_F d | 1439.2 ± 92.7 | 57.9 | <0.001 | 22.4 ± 0.7 |
| Strain | Characteristics | Reference |
|---|---|---|
| E. coli DH5α | fhuA2 lac(del)U169 phoA glnV44 Φ80’ lacZ(del)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17 | Laboratory stock |
| E. coli NEB® turbo | F’ proA + B + lacIq ∆lacZM15/fhuA2 ∆(lac-proAB) glnV galK16 galE15 R(zgb-210::Tn10)TetS endA1 thi-1 ∆(hsdS-mcrB)5 | Laboratory stock |
| B. subtilis strains | ||
| BS0 | ΔnprE, ΔaprE, Δepr, Δmpr, ΔnprB, Δvpr, Δbpr, ΔsigF | BGSC |
| BS1 | BS0 derivative, ΔlytC | Laboratory stock |
| BS2 | BS1 derivative, ΔamyE | This work |
| BSQ1a | BS2 derivative, amyQ (PspoVG) knock-in mutant (ΔspoVG) | This work |
| BSQ1b | BS1 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (ΔamyE) | This work |
| BSQ1c | BS2 derivative, amyQ (PspoVG-PamyQ-Pcry3A) knock-in mutant (ΔspoVG) | This work |
| BSQ1d | BS2 derivative, amyQ (SPywbN’) knock-in mutant (ΔywbN) | This work |
| BSQ1e | BS2 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (ΔpksG) | This work |
| BSQ1f | BS2 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (ΔppsE) | This work |
| BSQ1g | BS2 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (ΔcotB) | This work |
| BSQ1h | BS2 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (ΔylbP) | This work |
| BSQ1i | BS2 derivative, amyQ (PamyQ-Pcry3A) knock-in mutant (Δveg) | This work |
| BSQ2 | BSQ1b derivative, amyQ double knock-in mutant (ΔamyE, ΔpksG) | This work |
| BSQ3 | BSQ2 derivative, amyQ triple knock-in mutant (ΔamyE, ΔpksG,ΔppsE) | This work |
| BSQ4 | BSQ3 derivative, amyQ quadruple knock-in mutant (ΔamyE, ΔpksG,ΔppsE, ΔcotB) | This work |
| BSQ5 | BSQ4 derivative, amyQ quintuple knock-in mutant (ΔamyE, ΔpksG, ΔppsE, ΔcotB, ΔylbP) | This work |
| BSQ6 | BSQ4 derivative, amyQ sextuple knock-in mutant (ΔamyE, ΔpksG, ΔppsE, ΔcotB, ΔylbP, Δveg) | This work |
| BSQ6_1 | BSQ6 derivative, Δhag | This work |
| BSQ6_2 | BSQ6 derivative, ΔpssA | This work |
| BSQ6_3 | BSQ6 derivative, ΔyusX | This work |
| BSQ6_4 | BSQ6 derivative, ΔhrcA | This work |
| BSQ6_5 | BSQ6 derivative, sipT knock-in mutant (ΔthrC) | This work |
| BSQ6_6 | BSQ6 derivative, rasP knock-in mutant (ΔspoVG) | This work |
| BSQ6_7 | BSQ6 derivative, sppA knock-in mutant (ΔyqeZ) | This work |
| BSQ6_8 | BSQ6 derivative, secYEG knock-in mutant (ΔsigX) | This work |
| BSQ6_9 | BSQ6 derivative, prsA knock-in mutant (Δpel; ermr) | This work |
| BSQ6_10 | BSQ6 derivative, restored gudB gene | This work |
| BSQ6_11 | BSQ6 derivative, sppA and prsA knock-in mutant (ΔyqeZ, Δpel, ermr) | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrando, J.; Miñana-Galbis, D.; Picart, P. The Construction of an Environmentally Friendly Super-Secreting Strain of Bacillus subtilis through Systematic Modulation of Its Secretory Pathway Using the CRISPR-Cas9 System. Int. J. Mol. Sci. 2024, 25, 6957. https://doi.org/10.3390/ijms25136957
Ferrando J, Miñana-Galbis D, Picart P. The Construction of an Environmentally Friendly Super-Secreting Strain of Bacillus subtilis through Systematic Modulation of Its Secretory Pathway Using the CRISPR-Cas9 System. International Journal of Molecular Sciences. 2024; 25(13):6957. https://doi.org/10.3390/ijms25136957
Chicago/Turabian StyleFerrando, Jordi, David Miñana-Galbis, and Pere Picart. 2024. "The Construction of an Environmentally Friendly Super-Secreting Strain of Bacillus subtilis through Systematic Modulation of Its Secretory Pathway Using the CRISPR-Cas9 System" International Journal of Molecular Sciences 25, no. 13: 6957. https://doi.org/10.3390/ijms25136957
APA StyleFerrando, J., Miñana-Galbis, D., & Picart, P. (2024). The Construction of an Environmentally Friendly Super-Secreting Strain of Bacillus subtilis through Systematic Modulation of Its Secretory Pathway Using the CRISPR-Cas9 System. International Journal of Molecular Sciences, 25(13), 6957. https://doi.org/10.3390/ijms25136957