

MbWRKY53, a M. baccata WRKY Transcription Factor, Contributes to Cold and Drought Stress Tolerance in Transgenic Arabidopsis thaliana

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Conserved Structure and Physicochemical Properties of the MbWRKY53 Protein
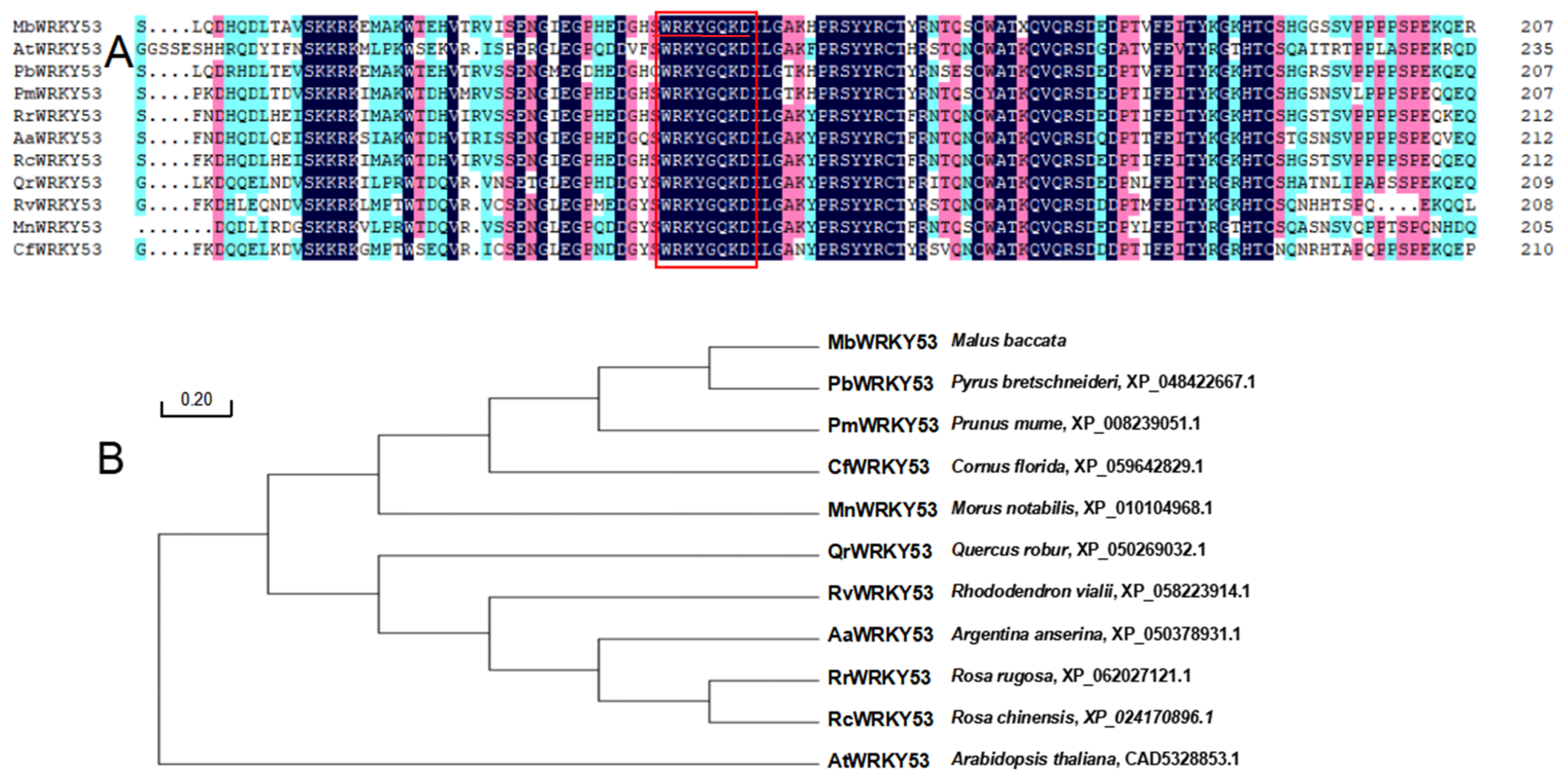
2.2. The Homology Analysis of the MbWRKY53 Protein
2.3. Structural Prediction of MbWRKY53 Protein
2.4. Subcellular Localization of MbWRKY53 Protein
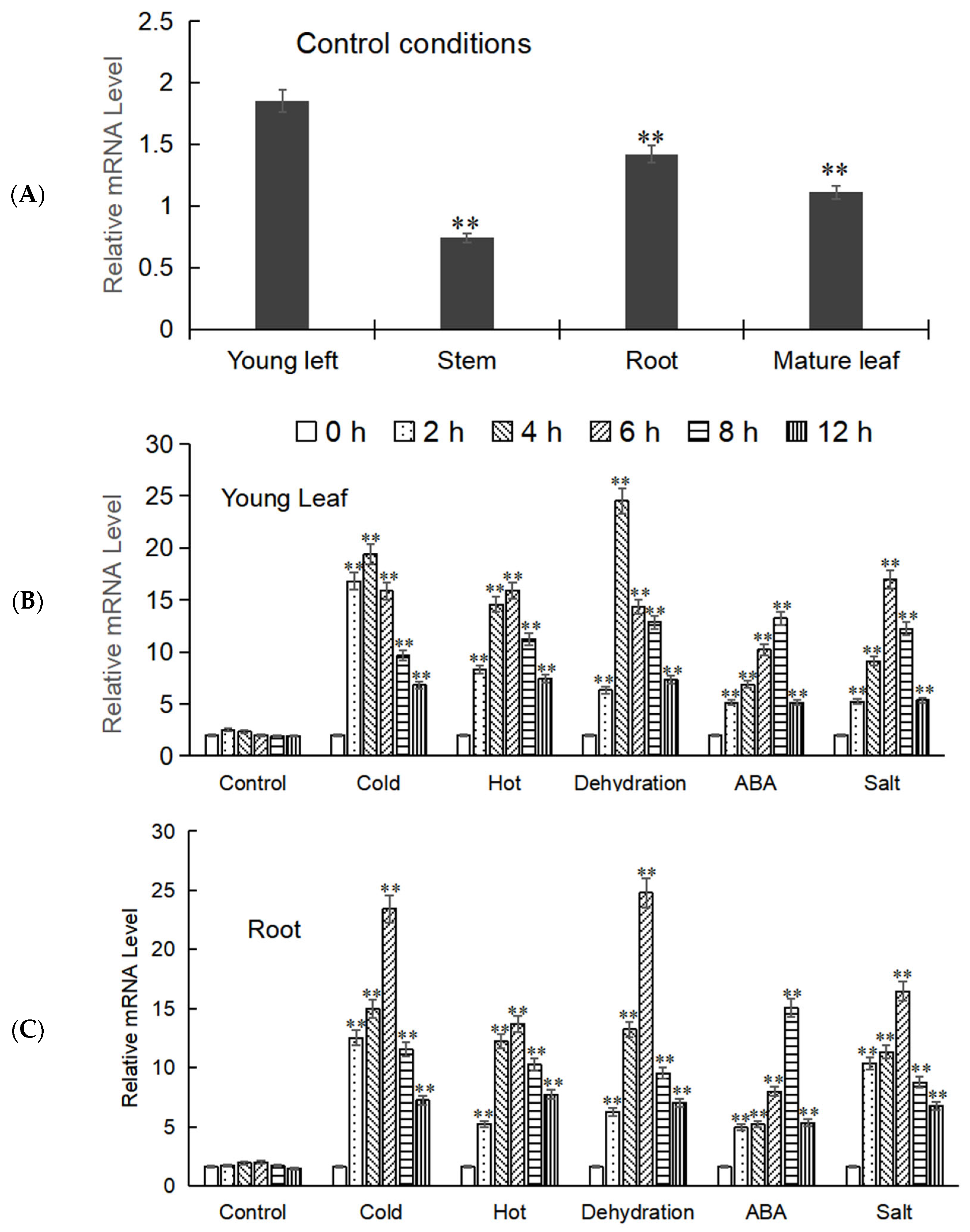
2.5. The Spatiotemporal Expression of MbWRKY53
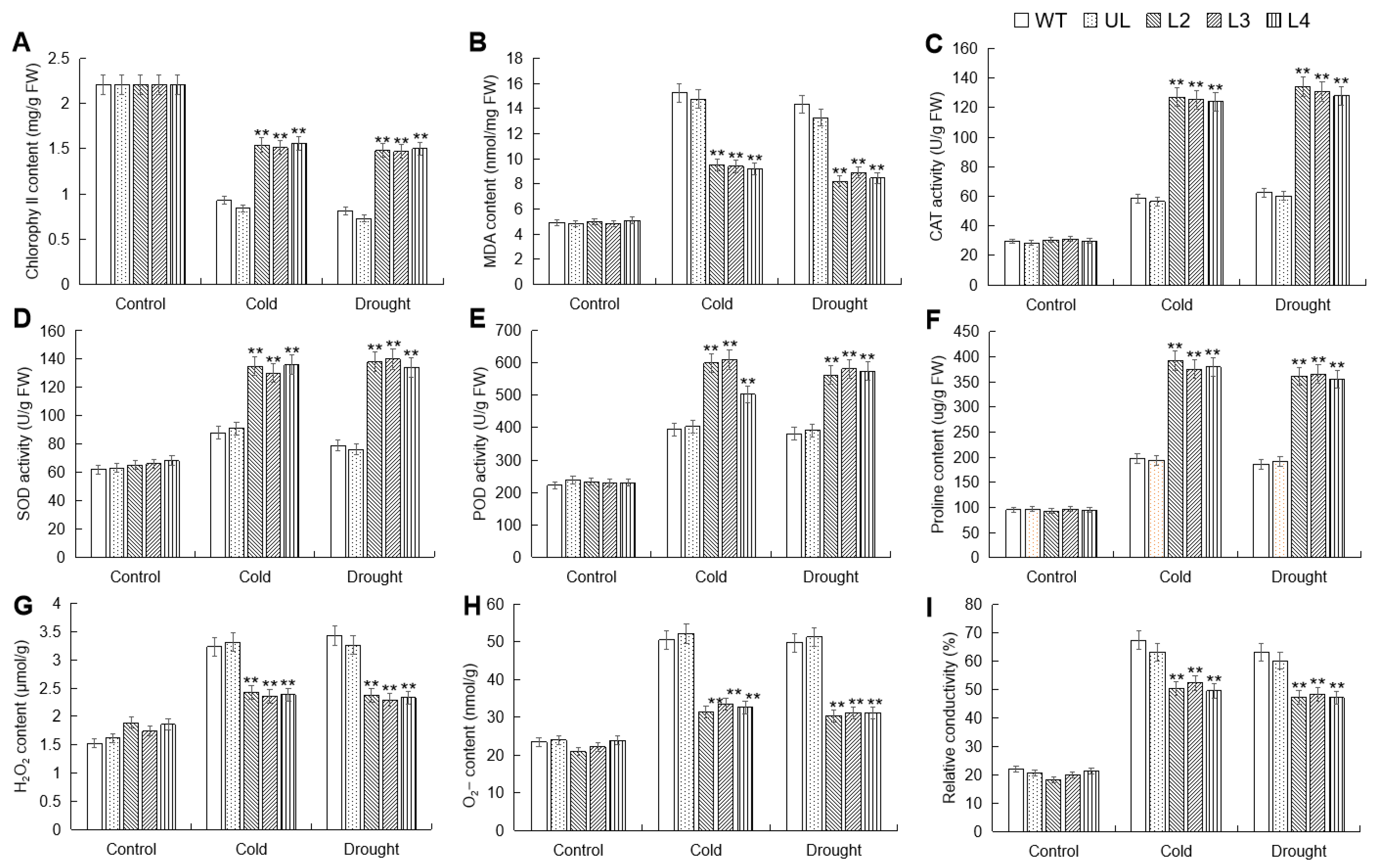
2.6. Analysis of Cold and Drought Tolerance of MbWRKY53-OE Arabidopsis
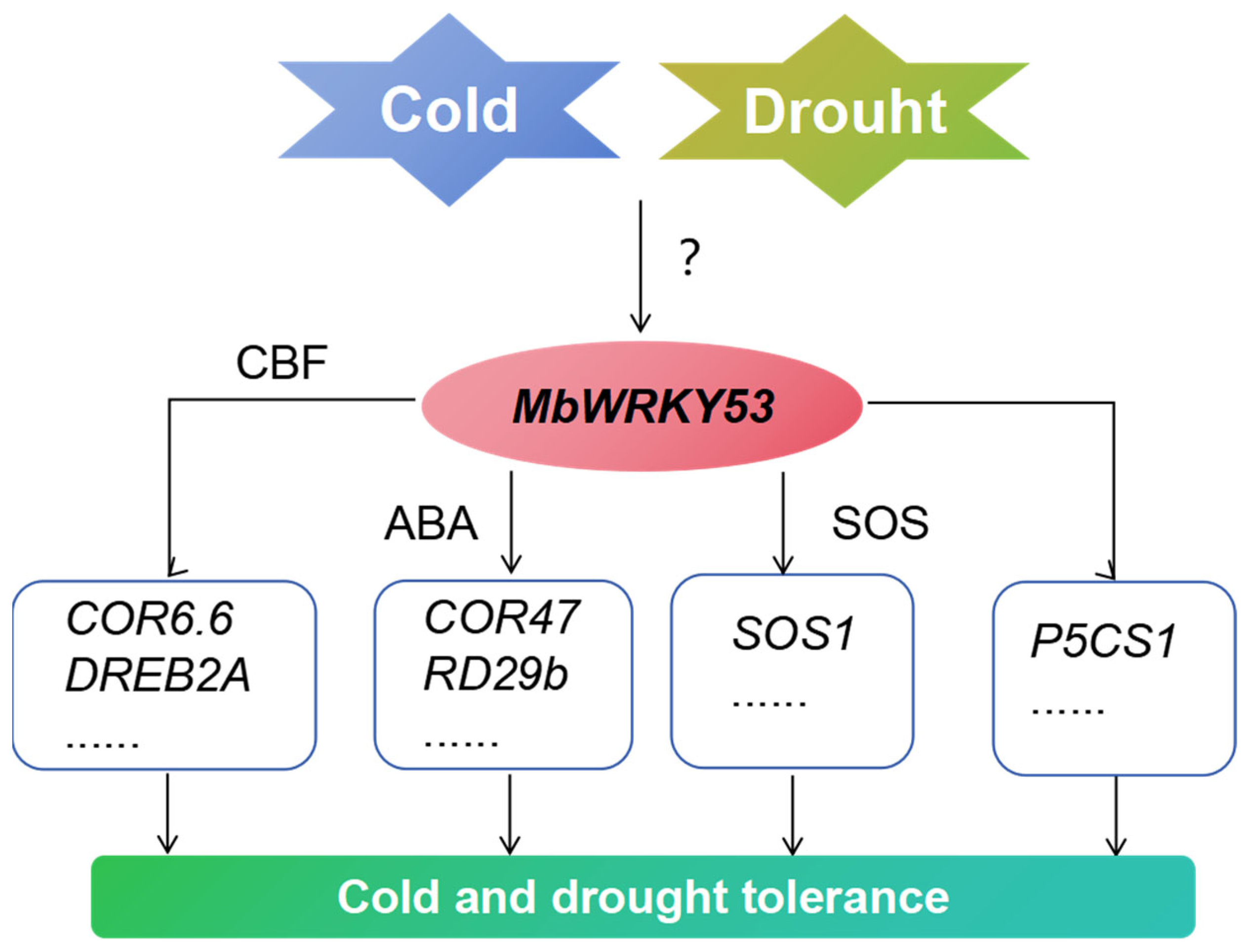
2.7. MbWRKY53 Induces the Expression of Cold and Drought-Resistant Genes in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Cultivation and Treatment of M. baccata
4.2. Sowing and Cultivation of Arabidopsis
4.3. Cloning and Sequencing of the MbWRKY53 Gene
4.4. Bioinformatic Analysis of the MbWRKY53 Gene
4.5. Subcellular Localization of MbWRKY53 Protein
4.6. Analysis of the Expression Level of the MbWRKY53 Gene
4.7. Obtaining Genetically Modified Arabidopsis
4.8. Determination of Relevant Physiological Indicators
4.9. Expression Analysis of Stress-Related Key Genes in Arabidopsis
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hridoy Ul Awall, R.; Tahjib-Ul-Arif, M.; Azim, M.A.; Tumpa, T.A.; Tipu, M.M.H.; Najnine, F.; Dawood, M.F.A.; Skalicky, M.; Brestič, M. Rice and food security: Climate change implications and the future prospects for nutritional security. Food Energy Secur. 2023, 12, e430. [Google Scholar]
- Chinnusamy, V.; Zhu, J.; Zhu, J. Salt stress signaling and mechanisms of plant salt tolerance. Genet. Eng. Princ. Methods 2006, 27, 141–177. [Google Scholar]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Biol. Res. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Du, M.; Zhou, Z.; Wang, S.; Li, T.; Han, J.; Xu, T.; Yang, G. An NAC transcription factor gene from Malus baccata, MbNAC29, increases cold and high salinity tolerance in Arabidopsis. In Vitro Cell. Dev-Plant. 2020, 56, 588–599. [Google Scholar] [CrossRef]
- Wang, Y.; He, W.; Wang, L.; Lan, Y.; Wu, M. TCP transcription factor identification in pecan (Carya illinoensis) and salt tolerance function analysis of CiTCP8. Sci. Hortic. 2024, 330, 113051. [Google Scholar] [CrossRef]
- Ishiguro, S.; Nakamura, K. Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 59 upstream regions of genes coding for sporamin and b-amylase from sweet potato. Mol. Gen. Genet. 1994, 244, 563–571. [Google Scholar] [CrossRef]
- Eulgem, T.; Somssich, I. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, R.; Jiang, S.; Kumar, N.; Venkatesh, P.N.; Ramachandran, S. A Comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant Cell Physiol. 2008, 49, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Jiang, Y.; Rahman, M.H.; Deyholos, M.K.; Kav, N.N. Identification and expression analysis of WRKY transcription factor genes in canola (Brassica napus L.) in response to fungal pathogens and hormone treatments. BMC Plant Biol. 2009, 9, 68. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.; Robatzek, S.; Kumar, S.; Nataraj, V.; Shivakumar, M.; Maheshwari, H.S.; Yadav, S.; Goswami, D. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, K.; Xu, Y.; Wang, L.; Liu, H.; Qin, Z.; Xiang, Y. The moso bamboo WRKY transcription factor, PheWRKY86, regulates drought tolerance in transgenic plants. Plant Physiol. Biochem. 2022, 170, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, H.; Yang, X.; Li, Q.; Ling, J.; Wang, H.; Gu, X.; Huang, S.; Jiang, W. CsWRKY46, a WRKY transcription factor from cucumber, confers cold resistance in transgenic-plant by regulating a set of cold-stress responsive genes in an ABA-dependent manner. Plant Physiol. Biochem. 2016, 108, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.; Kracher, B.; Somssich, I. Negative regulation of ABA signaling by WRKY33 is critical for Arabidopsis immunity towards Botrytis cinerea 2100. eLife 2015, 4, e07295. [Google Scholar]
- Han, D.; Wang, L.; Wang, Y.; Yang, W.; Gao, C.; Yu, Z.; Li, T.; Zhang, X.; Ma, L.; Xu, X.; et al. Overexpression of Malus xiaojinensis CS1 gene in tobacco affects plant development and increases iron stress tolerance. Sci. Hortic. 2013, 150, 65–72. [Google Scholar] [CrossRef]
- Liu, Q.; Zheng, L.; Wang, Y.; Zhou, Y.; Gao, F. AmDHN4, a winter accumulated SKn-type dehydrin from Ammopiptanthus mongolicus, and regulated by AmWRKY45, enhances the tolerance of Arabidopsis to low temperature and osmotic stress. Int. J. Biol. Macromol. 2024, 266, 131020. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Cur. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Song, H.; Guo, Z.; Duan, Z.; Zhang, J. WRKY transcription factors in Arachis hypogaea and its donors: From identification to function prediction. Plant Physiol. Biochem. 2023, 204, 108131. [Google Scholar] [CrossRef]
- Tang, W.; Wang, F.; Chu, H.; Li, M.; Zhang, J. WRKY transcription factors regulate phosphate uptake in plants. Plant Physiol. Biochem. 2023, 208, 05241. [Google Scholar] [CrossRef]
- Grzechowiak, M.; Ruszkowska, A.; Sliwiak, J.; Urbanowicz, A.; Jaskolski, M.; Ruszkowski, M. New aspects of DNA recognition by group II WRKY transcription factor revealed by structural and functional study of AtWRKY18 DNA binding domain. Int. J. Biol. Sci. 2022, 213, 589–601. [Google Scholar] [CrossRef]
- Li, A.; Chen, J.; Lin, Q.; Zhao, Y.; Duan, Y.; Wai, S.; Song, C.; Bi, J. Transcription Factor MdWRKY32 Participates in Starch-Sugar Metabolism by Binding to the MdBam5 Promoter in Apples During Postharvest Storage. J. Agric. Food Chem. 2021, 69, 14906–14914. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Song, T.; Wang, Y.; Zhang, M.; Li, N.; Yu, M.; Zhang, S.; Zhou, H.; Guo, S.; Bu, Y.; et al. The wheat WRKY transcription factor TaWRKY1-2D confers drought resistance in transgenic Arabidopsis and wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 2023, 226, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.; Macdonald, H.; Huttly, A.K.; Lazarus, C.M.; Hooley, R. Members of a new family of DNA-binding proteins bind to a conserved cis-element in the promoters of a-Amy2 genes. Plant Mol. Biol. 1995, 29, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Hou, Y.; Ding, H.; Zhou, Z.; Li, H.; Yang, G. Isolation and preliminary functional analysis of MbWRKY4 gene involved in salt tolerance in transgenic tobacco. Int. J. Agric. Biol. 2018, 20, 2045–2052. [Google Scholar]
- Meng, D.; Li, Y.; Bai, Y.; Li, M.; Cheng, L. Genome-wide identification and characterization of WRKY transcriptional factor family in apple and analysis of their responses to waterlogging and drought stress. Plant Physiol. Biochem. 2016, 103, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Wei, X.; Guan, W.; Nong, W.; Chen, R.; Tao, X.; Mao, L. ABA-responsive transcription factor ABF1-1 promotes JA biosynthesis to accelerate suberin polyphenolic formation in wounded kiwifruit (Actinidia chinensis). Postharvest Biol. Technol. 2022, 187, 111850. [Google Scholar] [CrossRef]
- Senuma, M.; Dobashi, S.; Bando, Y.; Ko, S.; Shiota, H. Overexpression of eelgrass Rare Cold Inducible 2 (RCI2) maintains chlorophyll content in Arabidopsis subjected to high salinity and dehydration. Plant Stress 2022, 6, 100116. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, A.; Sedlářová, V. Pavel Pospíšil, Malondialdehyde enhances PsbP protein release during heat stress in Arabidopsis. Plant Physiol. Biochem. 2023, 202, 107984. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Yasin, N.A.; Shah, A.A.; Khan, W.U.; Li, G.; Ahmad, A.; Ahmed, S.; Hussaan, M.; Rizwan, M.; Ali, S. Exogenous application of liquiritin alleviated salt stress and improved growth of Chinese kale plants. Sci. Hortic. 2022, 294, 110762. [Google Scholar] [CrossRef]
- Li, C.; Lu, X.; Liu, Y.; Xu, J.; Yu, W. Trehalose alleviates the inhibition of adventitious root formation caused by drought stress in cucumber through regulating ROS metabolism and activating trehalose and plant hormone biosynthesis. Plant Physiol. Biochem. 2023, 205, 108159. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.; Dubey, R. Oxidative Stress and Antioxidative Defense System in Plants Growing under Abiotic Stresses; CRC Press: Boca Raton, FL, USA, 2019; pp. 93–136. [Google Scholar]
- Srivastava, S.; Tripathi, R.; Dwivedi, U. Synthesis of phytochelatins and modulation of antioxidants in response to cadmium stress in Cuscuta reflexa—An angiospermic parasite. Plant Physiol. 2004, 161, 665–674. [Google Scholar] [CrossRef]
- Singh, K.; Foley, R.; Luis, O. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Wang, Y.; Zhang, L.; Ma, L.; Zhang, X.; Xu, X.; Han, Z. Isolation and functional characterization of MxCS1: A gene encoding a citrate synthase in Malus xiaojinensis. Biol. Plant. 2012, 56, 50–56. [Google Scholar] [CrossRef]
- Han, D.; Shi, Y.; Wang, B.; Liu, W.; Yu, Z.; Lv, B.; Yang, G. Isolation and preliminary functional analysis of MxCS2: A gene encoding a citrate synthase in Malus xiaojinensis. Plant Mol. Biol. Rep. 2015, 33, 133–142. [Google Scholar] [CrossRef]
- Kwon, T. Mitochondrial Porin Isoform AtVDAC1 Regulates the Competence of Arabidopsis thaliana to Agrobacterium-Mediated Genetic Transformation. Mol. Cells 2016, 39, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Yu, D.; Wang, X. VvWRKY22 transcription factor interacts with VvSnRK1.1/VvSnRK1.2 and regulates sugar accumulation in grape. Biochem. Biophys. Res. Commun. 2021, 554, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yang, L.; Liu, W.; Xu, X.; Ran, M.; Jin, Y.; Sun, X. MAP30 and luffin-α: Novel ribosome-inactivating proteins induce plant systemic resistance against plant viruses. Pestic. Biochem. Phys. 2023, 191, 105342. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, W.; Zhang, P.; Pan, Q.; Zhan, J.; Huang, W. Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries. Plant Sci. 2010, 179, 103–113. [Google Scholar] [CrossRef]
- Modesto, P.; Peletto, S.; Pisoni, G.; Cremonesi, P.; Castiglioni, B.; Colussi, S.; Caramelli, M.; Bronzo, V.; Moroni, P.; Acutis, P.L. Evaluation of internal reference genes for quantitative expression analysis by real-time reverse transcription-PCR in somatic cells from goat milk. J. Dairy Sci. 2013, 96, 7932–7944. [Google Scholar] [CrossRef]
- Karkute1, S.G.; Gujjar, R.S.; Rai1, A.; Akhtar, M.; Singh, M.; Singh, B. Genome wide expression analysis of WRKY genes in tomato (Solanum lycopersicum) under drought stress. Plant Gene 2018, 13, 8–17. [Google Scholar] [CrossRef]
- Han, D.; Yang, G.; Xu, K.; Shao, Q.; Yu, Z.; Wang, B.; Ge, Q.; Yu, Y. Overexpression of a Malus xiaojinensis Nas1 gene influences flower development and tolerance to iron stress in transgenic tobacco. Plant Mol. Biol. Rep. 2013, 31, 802–809. [Google Scholar] [CrossRef]
- Zheng, Y.; He, X.; Ying, Y.; Lu, J.; Gelvin, S.B.; Shou, H. Expression of the Arabidopsis thaliana Histone Gene AtHTA1 Enhances Rice Transformation Efficiency. Mol. Plant 2009, 2, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Wang, Y.; Zhang, Z.; Pu, Q.; Ding, H.; Han, J.; Fan, T.; Bai, X.; Yang, G. Isolation and functional analysis of MxCS3: A gene encoding a citrate synthase in Malus xiaojinensis, with functions in tolerance to iron stress and abnormal flower in transgenic Arabidopsis thaliana. Plant Growth Regul. 2017, 82, 479–489. [Google Scholar] [CrossRef]
- Karkute, S.G.; Gujjar, R.S.; Rai, A.; Akhtar, M.; Singh, M.; Singh, B. The citrus transcription factor CsMADS6 modulates carotenoid metabolism by directly regulating carotenogenic genes. Plant Physiol. 2018, 176, 2657–2676. [Google Scholar]
- Huang, Q.; Qian, X.; Jiang, T.; Zheng, X. Effect of eugenol fumigation treatment on chilling injury and CBF gene expression in eggplant fruit during cold storage. Food Chem. 2019, 292, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, Y.; Wan, H.; Tang, J.; Ni, Z. The sea-island cotton GbTCP4 transcription factor positively regulates drought and salt stress responses. Plant Sci. 2022, 322, 111329. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Luo, G.; Li, W.; Yao, A.; Liu, W.; Xie, L.; Han, M.; Li, X.; Han, D. Overexpression of a Malus baccata CBF transcription factor gene, MbCBF1, increases cold and salinity tolerance in Arabidopsis thaliana. Plant Physiol. Biochem. 2022, 192, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, F.; Garrido, I. Thallium induced increases on O2−, H2O2, NO and H2S production, and morpho-physiological alterations in Ditrichia viscosa plants. Free Radical Res. 2022, 189, 45–46. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, F.; Wang, Y.; Zhang, S.; Wang, F.; Ma, Y. The cytochrome P450 gene, MdCYP716B1, is involved in regulating plant growth and anthracnose resistance in apple. Plant Sci. 2023, 335, 111832. [Google Scholar] [CrossRef]
- Han, D.; Shi, Y.; Yu, Z.; Liu, W.; Lv, B.; Wang, B.; Yang, G. Isolation and functional analysis of MdCS1: A gene encoding a citrate synthase in Malus domestica (L.) Borkh. Plant Growth Regul. 2015, 75, 209–218. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Wang, T.; Liang, X.; Ye, Q.; Wang, Y.; Han, J.; Han, D. MbWRKY53, a M. baccata WRKY Transcription Factor, Contributes to Cold and Drought Stress Tolerance in Transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2024, 25, 7626. https://doi.org/10.3390/ijms25147626
Liu W, Wang T, Liang X, Ye Q, Wang Y, Han J, Han D. MbWRKY53, a M. baccata WRKY Transcription Factor, Contributes to Cold and Drought Stress Tolerance in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences. 2024; 25(14):7626. https://doi.org/10.3390/ijms25147626
Chicago/Turabian StyleLiu, Wanda, Tianhe Wang, Xiaoqi Liang, Qinglei Ye, Yu Wang, Jilong Han, and Deguo Han. 2024. "MbWRKY53, a M. baccata WRKY Transcription Factor, Contributes to Cold and Drought Stress Tolerance in Transgenic Arabidopsis thaliana" International Journal of Molecular Sciences 25, no. 14: 7626. https://doi.org/10.3390/ijms25147626
APA StyleLiu, W., Wang, T., Liang, X., Ye, Q., Wang, Y., Han, J., & Han, D. (2024). MbWRKY53, a M. baccata WRKY Transcription Factor, Contributes to Cold and Drought Stress Tolerance in Transgenic Arabidopsis thaliana. International Journal of Molecular Sciences, 25(14), 7626. https://doi.org/10.3390/ijms25147626