
Differential Expression of Circulating miRNAs and Carfilzomib-Related Cardiovascular Adverse Events in Patients with Multiple Myeloma

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Study Population
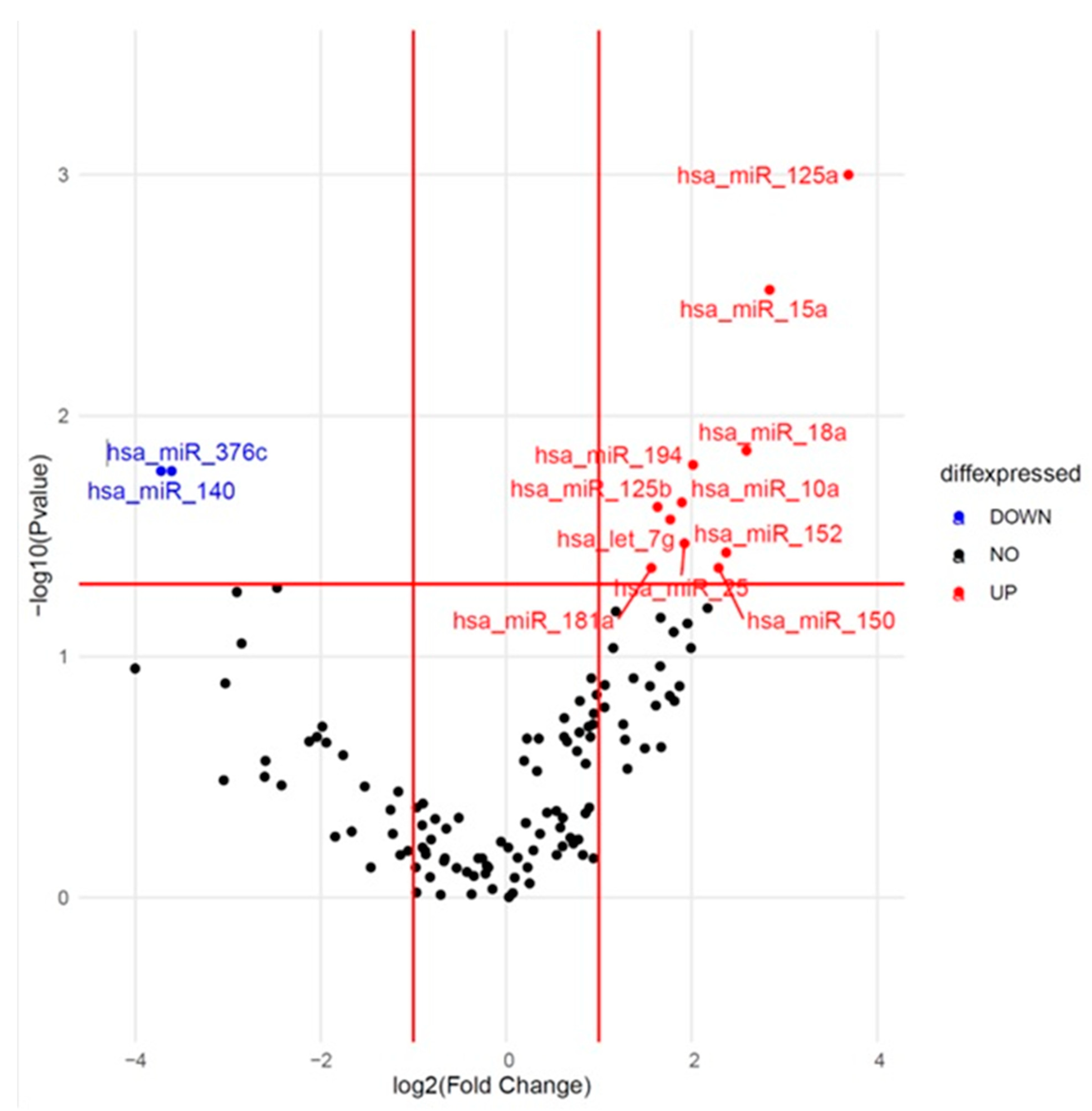
2.2. Identification of Differentially Expressed miRNAs at Baseline
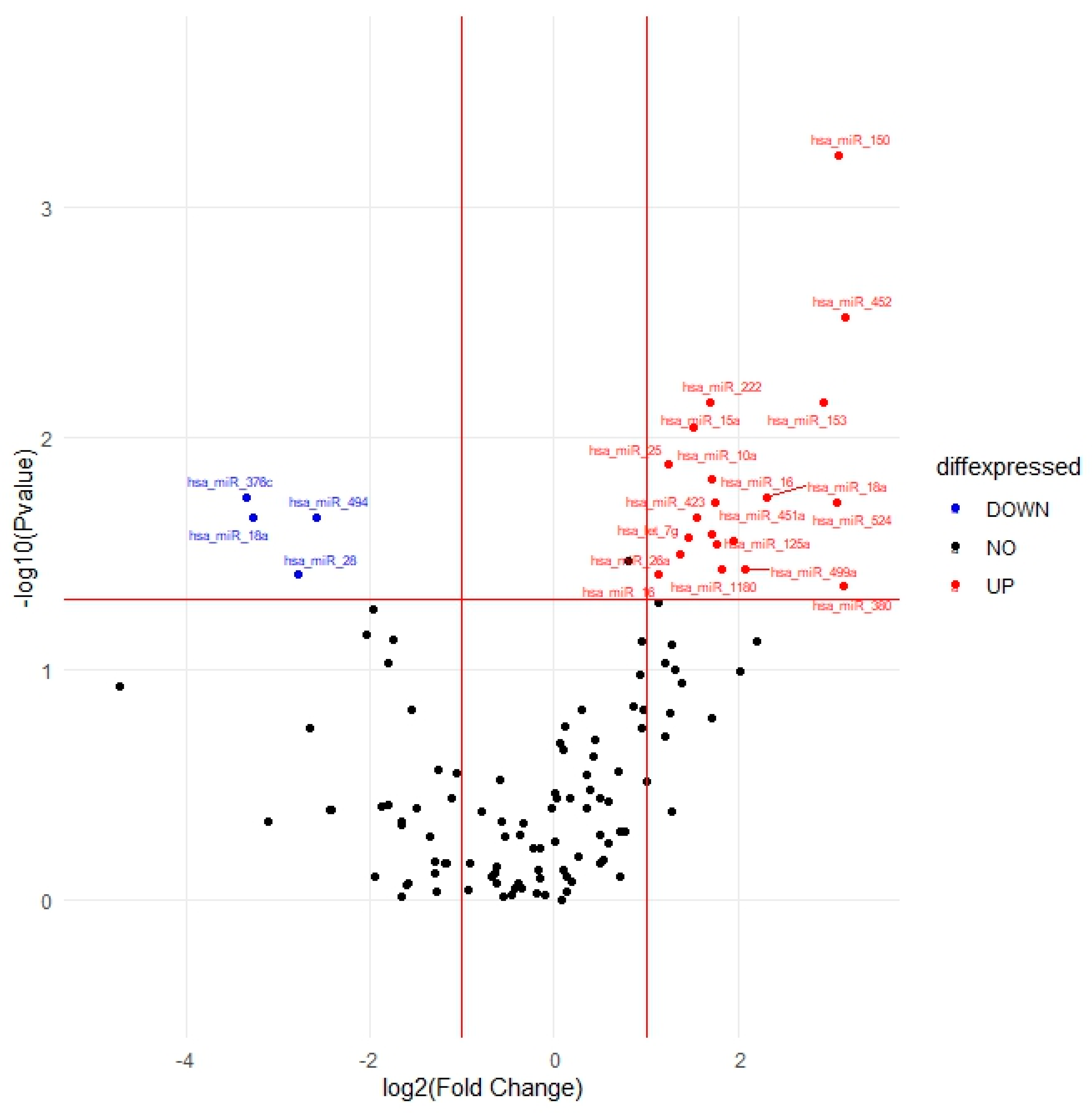
2.3. Identification of Differentially Expressed miRNAs Post-Treatment
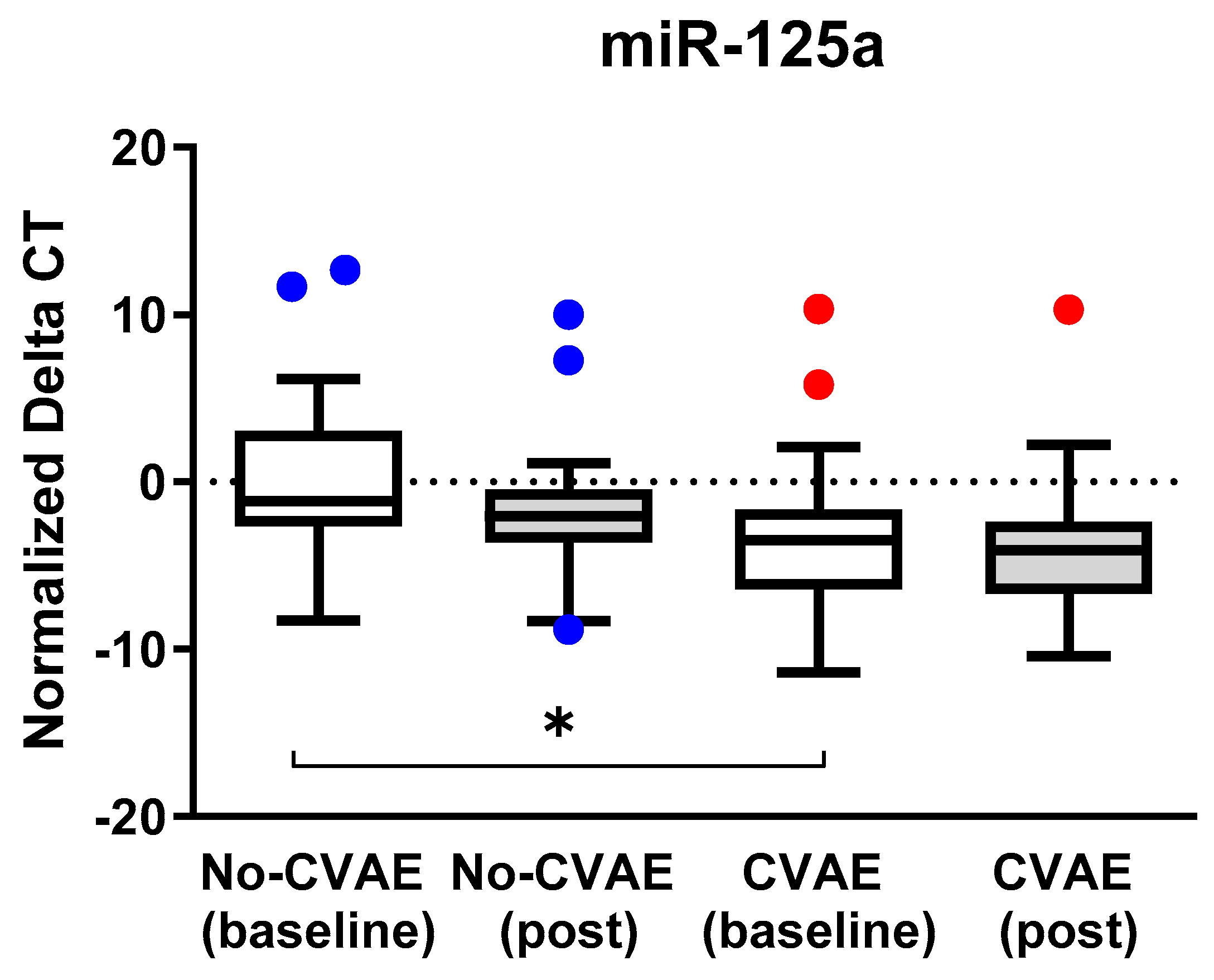
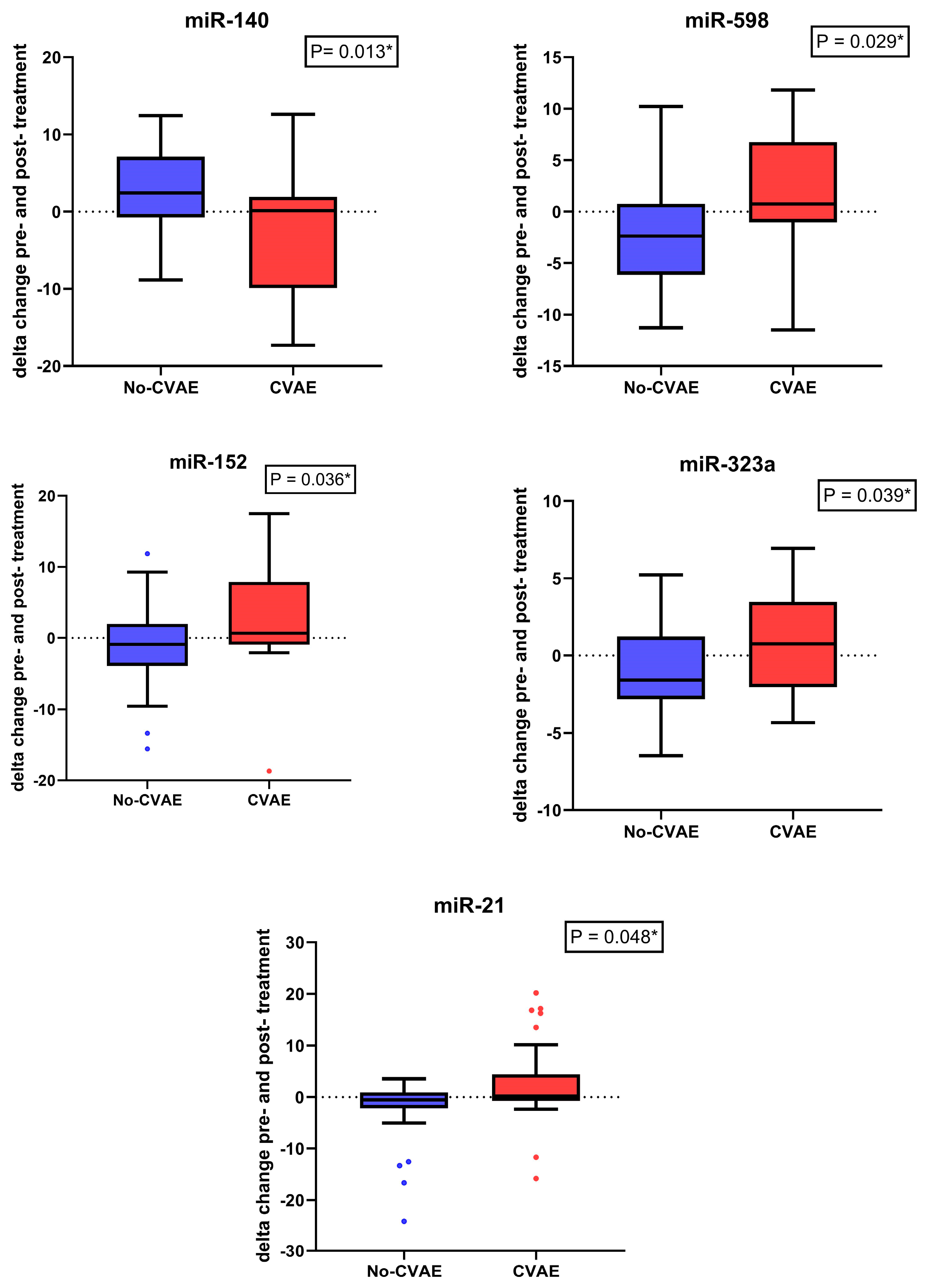
2.4. Change in the miRNA’s Expression between Baseline and Post-Treatment
2.5. Pathway Enrichment Analyses
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Circulating miRNA Isolation and Open Array Profiling
4.3. Statistical Analysis of Open Array Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, D.H.; Kumar, A.; Mohammed, T.; Peres, L.C.; Alsina, M.; Bachmeier, C.; Blue, B.J.; Brayer, J.; Chandrasekhar, S.; Grajales Cruz, A.; et al. Cardiac events after standard of care idecabtagene vicleucel for relapsed and refractory multiple myeloma. Blood Adv. 2023, 7, 4247–4257. [Google Scholar] [CrossRef]
- Fontes Oliveira, M.; Naaktgeboren, W.R.; Hua, A.; Ghosh, A.K.; Oakervee, H.; Hallam, S.; Manisty, C. Optimising cardiovascular care of patients with multiple myeloma. Heart 2021, 107, 1774–1782. [Google Scholar] [CrossRef]
- Kistler, K.D.; Kalman, J.; Sahni, G.; Murphy, B.; Werther, W.; Rajangam, K.; Chari, A. Incidence and Risk of Cardiac Events in Patients with Previously Treated Multiple Myeloma Versus Matched Patients without Multiple Myeloma: An Observational, Retrospective, Cohort Study. Clin Lymphoma Myeloma Leuk. 2017, 17, 89–96.e3. [Google Scholar] [CrossRef]
- Bringhen, S.; Milan, A.; Ferri, C.; Wäsch, R.; Gay, F.; Larocca, A.; Salvini, M.; Terpos, E.; Goldschmidt, H.; Cavo, M.; et al. Cardiovascular adverse events in modern myeloma therapy—Incidence and risks. A review from the European Myeloma Network (EMN) and Italian Society of Arterial Hypertension (SIIA). Haematologica 2018, 103, 1422–1432. [Google Scholar] [CrossRef]
- Chari, A.; Stewart, A.K.; Russell, S.D.; Moreau, P.; Herrmann, J.; Banchs, J.; Hajek, R.; Groarke, J.; Lyon, A.R.; Batty, G.N.; et al. Analysis of carfilzomib cardiovascular safety profile across relapsed and/or refractory multiple myeloma clinical trials. Blood Adv. 2018, 2, 1633–1644. [Google Scholar] [CrossRef]
- Waxman, A.J.; Clasen, S.; Hwang, W.-T.; Garfall, A.; Vogl, D.T.; Carver, J.; O’Quinn, R.; Cohen, A.D.; Stadtmauer, E.A.; Ky, B.; et al. Carfilzomib-Associated Cardiovascular Adverse Events. JAMA Oncol. 2018, 4, e174519. [Google Scholar] [CrossRef]
- Efentakis, P.; Kremastiotis, G.; Varela, A.; Nikolaou, P.E.; Papanagnou, E.D.; Davos, C.H.; Tsoumani, M.; Agrogiannis, G.; Konstantinidou, A.; Kastritis, E.; et al. Molecular mechanisms of carfilzomib-induced cardiotoxicity in mice and the emerging cardioprotective role of metformin. Blood 2019, 133, 710–723. [Google Scholar] [CrossRef]
- Gandolfi, S.; Laubach, J.P.; Hideshima, T.; Chauhan, D.; Anderson, K.C.; Richardson, P.G. The proteasome and proteasome inhibitors in multiple myeloma. Cancer Metastasis Rev. 2017, 36, 561–584. [Google Scholar] [CrossRef]
- Teicher, B.A.; Tomaszewski, J.E. Competitive Landscape report. Biochem. Pharmacol. 2015, 96, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Atrash, S.; Tullos, A.; Panozzo, S.; Bhutani, M.; Van Rhee, F.; Barlogie, B.; Usmani, S.Z. Cardiac complications in relapsed and refractory multiple myeloma patients treated with carfilzomib. Blood Cancer J. 2015, 5, e272. [Google Scholar] [CrossRef] [PubMed]
- Efentakis, P.; Varela, A.; Lamprou, S.; Papanagnou, E.; Chatzistefanou, M.; Christodoulou, A.; Davos, C.H.; Gavria-topoulou, M.; Trougakos, I.; Dimopoulos, M.A.; et al. Implications and hidden toxicity of cardiometabolic syndrome and early-stage heart failure in carfilzomib-induced cardiotoxicity. Br. J. Pharmacol. 2024, 181, 2964–2990. [Google Scholar] [CrossRef]
- Londina, E.; Lohera, P.; Telonis, A.G.; Quann, K.; Clark, P.; Jinga, Y.; Hatzimichael, E.; Kirino, Y.; Honda, S.; Lally, M.; et al. Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- And tissue-specific microRNAs. Proc. Natl. Acad. Sci. USA 2015, 112, E1106–E1115. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-J.; Yang, S.-Y.; Wang, D.-D.; Chen, X.; Shen, H.-Y.; Zhang, X.-H.; Zhong, S.-L.; Tang, J.-H.; Zhao, J.-H. The miR-30 family: Versatile players in breast cancer. Tumor Biol. 2017, 39, 101042831769220. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.K.; Makhoul, I.; Wei, J.; Klimberg, V. Circulating miRNA Profiles of Doxorubicin-induced Cardiotoxicity in Breast Cancer Patients. Ann. Clin Lab Sci. 2017, 47, 115–119. [Google Scholar] [PubMed]
- Qin, X.; Chang, F.; Wang, Z.; Jiang, W. Correlation of circulating pro-angiogenic miRNAs with cardiotoxicity induced by epirubicin/cyclophosphamide followed by docetaxel in patients with breast cancer. Cancer Biomarkers 2018, 23, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Boen, H.M.; Cherubin, M.; Franssen, C.; Gevaert, A.B.; Witvrouwen, I.; Bosman, M.; Guns, P.-J.; Heidbuchel, H.; Loeys, B.; Alaerts, M.; et al. Circulating MicroRNA as Biomarkers of Anthracycline-Induced Cardiotoxicity. JACC CardioOncol. 2024, 6, 183–199. [Google Scholar] [CrossRef]
- Tantawy, M.; Pamittan, F.G.; Singh, S.; Gong, Y. Epigenetic Changes Associated With Anthracycline-Induced Cardiotoxicity. Clin. Transl. Sci. 2020, 14, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Koturbash, I.; Tolleson, W.H.; Guo, L.; Yu, D.; Chen, S.; Hong, H.; Mattes, W.; Ning, B. microRNAs as pharmacogenomic biomarkers for drug efficacy and drug safety assessment. Biomark. Med. 2015, 9, 1153–1176. [Google Scholar] [CrossRef]
- Han, D.; Wang, Y.; Wang, Y.; Dai, X.; Zhou, T.; Chen, J.; Tao, B.; Zhang, J.; Cao, F. The Tumor-Suppressive Human Circular RNA CircITCH Sponges miR-330-5p to Ameliorate Doxorubicin-Induced Cardiotoxicity Through Upregulating SIRT6, Survivin, and SERCA2a. Circ. Res. 2020, 127, e108–e125. [Google Scholar] [CrossRef]
- Ananthan, K.; Lyon, A.R. The Role of Biomarkers in Cardio-Oncology. J. Cardiovasc. Transl. Res. 2020, 13, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Qi, Y.; Xu, L.; Tao, X.; Han, X.; Yin, L.; Peng, J. MicroRNA-140-5p aggravates doxorubicin-induced cardiotoxicity by promoting myocardial oxidative stress via targeting Nrf2 and Sirt2. Redox Biol. 2018, 15, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandão, B.B.; Kahn, C.R. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cao, Y.; Sun, M.; Feng, H. Expression, regulation, and function of exosome-derived miRNAs in cancer progression and therapy. FASEB J. 2021, 35, e21916. [Google Scholar] [CrossRef]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, C.; Gioffré, S.; Achilli, F.; Colombo, G.I.; D’Alessandra, Y. Role of microRNAs in doxorubicin-induced cardiotoxicity: An overview of preclinical models and cancer patients. Heart Fail. Rev. 2018, 23, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.G.; Vijay, V.; Lee, T.; Han, T.; Moland, C.L.; Phanavanh, B.; Herman, E.H.; Stine, K.; Fuscoe, J.C. MicroRNA-34a-5p as a promising early circulating preclinical biomarker of doxorubicin-induced chronic cardiotoxicity. J. Appl. Toxicol. 2022, 42, 1477–1490. [Google Scholar] [CrossRef]
- Cornell, R.F.; Ky, B.; Weiss, B.M.; Dahm, C.N.; Gupta, D.K.; Du, L.; Carver, J.R.; Cohen, A.D.; Engelhardt, B.G.; Garfall, A.L.; et al. Prospective study of cardiac events during proteasome inhibitor therapy for relapsed multiple myeloma. J. Clin. Oncol. 2019, 37, 1946–1955. [Google Scholar] [CrossRef]
- Aparicio-Puerta, E.; Hirsch, P.; Schmartz, G.P.; Kern, F.; Fehlmann, T.; Keller, A. miEAA 2023: Updates, new functional microRNA sets and improved enrichment visualizations. Nucleic Acids Res. 2023, 51, W319–W325. [Google Scholar] [CrossRef]
- Williams, A.L.; Khadka, V.S.; Anagaran, M.C.T.; Lee, K.; Avelar, A.; Deng, Y.; Shohet, R.V. miR-125 family regulates XIRP1 and FIH in response to myocardial infarction. Physiol. Genom. 2020, 52, 358–368. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, J.; Wang, L.; Pei, G.; Cheng, H.; Zhang, Q.; Wang, S.; He, C.; Fu, C.; Wei, Q. MiR-125 Family in Cardiovascular and Cerebrovascular Diseases. Front. Cell Dev. Biol. 2021, 9, 799049. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ha, T.; Zou, J.; Ren, D.; Liu, L.; Zhang, X.; Kalbfleisch, J.; Gao, X.; Williams, D.; Li, C. MicroRNA-125b protects against myocardial ischaemia/reperfusion injury via targeting p53-mediated apoptotic signalling and TRAF6. Cardiovasc. Res. 2014, 102, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Tang, W.; Lu, R.; Tao, Y.; Ren, T.; Gao, Y. Human Adipose-derived mesenchymal stem cells promote lymphocyte apoptosis and alleviate atherosclerosis via miR-125b-1-3p/BCL11B signal axis. Ann. Palliat. Med. 2021, 10, 2123–2133. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Li, L.; Dong, L.; Cheng, J.; Zhao, C.; Bao, C.; Wang, H. Circulating mRNA and microRNA profiling analysis in patients with ischemic stroke. Mol. Med. Rep. 2020, 22, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-M.; Lin, K.-Y.; Chen, Y.-Q. Diverse functions of miR-125 family in different cell contexts. J. Hematol. Oncol. 2013, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Niu, C. The correlation of long non-coding RNA intersectin 1-2 with disease risk, disease severity, inflammation, and prognosis of acute ischemic stroke. J. Clin. Lab. Anal. 2020, 34, e23053. [Google Scholar] [CrossRef]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Meese, E.; Saleh, M.A.; Keller, A.; Abdul-Khaliq, H.; Raedle-Hurst, T. Micro-RNA 150-5p predicts overt heart failure in patients with univentricular hearts. PLoS ONE 2019, 14, e0223606. [Google Scholar] [CrossRef] [PubMed]
- Scrutinio, D.; Conserva, F.; Passantino, A.; Iacoviello, M.; Lagioia, R.; Gesualdo, L. Circulating microRNA-150-5p as a novel biomarker for advanced heart failure: A genome-wide prospective study. J. Heart Lung Transplant. 2017, 36, 616–624. [Google Scholar] [CrossRef]
- Verma, A.; Bhattacharya, R.; Remadevi, I.; Li, K.; Pramanik, K.; Samant, G.V.; Horswill, M.; Chun, C.Z.; Zhao, B.; Wang, E.; et al. Endothelial cell-specific chemotaxis receptor (ecscr) promotes angioblast migration during vasculogenesis and enhances VEGF receptor sensitivity. Blood 2010, 115, 4614–4622. [Google Scholar] [CrossRef]
- Zhang, L.; Li, B.; Zhang, D.; Zhao, Y.; Yu, Q. lncRNA SNHG12 Inhibition Based on Microsystem Cell Imaging Technology Protects the Endothelium from LPS-Induced Inflammation by Inhibiting the Expression of miR-140-3p Target Gene fndc5. Contrast Media Mol. Imaging 2022, 2022, 1681864. [Google Scholar] [CrossRef]
- Zhang, H.; Ji, N.; Gong, X.; Ni, S.; Wang, Y. NEAT1/miR-140-3p/MAPK1 mediates the viability and survival of coronary endothelial cells and affects coronary atherosclerotic heart disease. Acta Biochim. Biophys. Sin. 2020, 52, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, J.-D.; Fang, X.-H.; Zhu, J.-N.; Yang, J.; Pan, R.; Yuan, S.-J.; Zeng, N.; Yang, Z.-Z.; Yang, H.; et al. Circular RNA circRNA_000203 aggravates cardiac hypertrophy via suppressing miR-26b-5p and miR-140-3p binding to Gata4. Cardiovasc. Res. 2020, 116, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Jiao, J.; Wang, J.; Li, Y.; Qin, D.; Li, P. Mitofusin 1 is negatively regulated by microRNA 140 in cardiomyocyte apoptosis. Mol. Cell. Biol. 2014, 34, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, J.; Li, Y.; Qin, D.; Li, P. Mitochondrial fission controls DNA fragmentation by regulating endonuclease G. Free Radic. Biol. Med. 2010, 49, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Ma, X.; Wang, J.; An, Z.; Xue, J.; Wang, Q. Evaluation of left ventricular dysfunction by three-dimensional speckle-tracking echocardiography and bioinformatics analysis of circulating exosomal miRNA in obese patients. BMC Cardiovasc. Disord. 2023, 23, 450. [Google Scholar] [CrossRef]
- Karakas, M.; Schulte, C.; Appelbaum, S.; Ojeda, F.; Lackner, K.J.; Münzel, T.; Schnabel, R.B.; Blankenberg, S.; Zeller, T. Circulating microRNAs strongly predict cardiovascular death in patients with coronary artery disease-results from the large AtheroGene study. Eur. Heart J. 2017, 38, 516–523. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Rehman, T.; Shabbir, M.A.; Inam-Ur-Raheem, M.; Manzoor, M.F.; Ahmad, N.; Liu, Z.-W.; Ahmad, M.H.; Siddeeg, A.; Abid, M.; Aadil, R.M. Cysteine and homocysteine as biomarker of various diseases. Food Sci. Nutr. 2020, 8, 4696–4707. [Google Scholar] [CrossRef]
- Ward, N.P.; Yoon, S.J.; Flynn, T.; Sherwood, A.M.; Olley, M.A.; Madej, J.; DeNicola, G.M. Mitochondrial respiratory function is preserved under cysteine starvation via glutathione catabolism in NSCLC. Nat. Commun. 2024, 15, 4244. [Google Scholar] [CrossRef]
- Liu, M.; Lv, J.; Pan, Z.; Wang, D.; Zhao, L.; Guo, X. Mitochondrial dysfunction in heart failure and its therapeutic implications. Front. Cardiovasc. Med. 2022, 9, 945142. [Google Scholar] [CrossRef] [PubMed]
- Westermann, D.; Lindner, D.; Kasner, M.; Zietsch, C.; Savvatis, K.; Escher, F.; von Schlippenbach, J.; Skurk, C.; Steendijk, P.; Riad, A.; et al. Cardiac inflammation contributes to changes in the extracellular matrix in patients with heart failure and normal ejection fraction. Circ. Heart. Fail. 2011, 4, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Paiva, B.; Anderson, K.C.; Durie, B.; Landgren, O.; Moreau, P.; Munshi, N.; Lonial, S.; Bladé, J.; Mateos, M.-V.; et al. International Myeloma Working Group consensus criteria for response and minimal residual disease assessment in multiple myeloma. Lancet Oncol. 2016, 17, e328–e346. [Google Scholar] [CrossRef] [PubMed]
- Cancer Institute, N. Common Terminology Criteria for Adverse Events (CTCAE) Version 4.0. 2009. Available online: http://www.meddramsso.com (accessed on 27 August 2020).
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T))method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: MultiExperiment Viewer. In Biomedical Informatics for Cancer Research; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | CVAEs (n = 31) | Non-CVAEs (n = 29) | p-Value |
|---|---|---|---|
| Age | 67.16 ± 8.81 | 64.66 ± 9.51 | 0.29 |
| Sex (Male) | 23 (74.19) | 22 (75.86) | 0.88 |
| Race | >0.99 | ||
| White | 27 (87.10) | 25 (86.21) | |
| Black | 4 (12.90) | 4(13.79) | |
| Smoker | 15 (48.39) | 11(37.93) | 0.41 |
| NYHA | 0.75 | ||
| Class 1 | 18 (58.06) | 18 (62.07) | |
| Class 2 | 13 (41.94) | 11 (37.93) | |
| Hypertension | 15 (48.39) | 8 (27.59) | 0.10 |
| CVD (family history) | 19 (61.29) | 15 (51.72) | 0.45 |
| Diabetes mellitus | 4 (12.90) | 4 (13.79) | >0.99 |
| Hypercholesterolemia | 8 (25.81) | 11 (37.93) | 0.31 |
| Thrombosis | 6 (19.35) | 4 (13.79) | 0.73 |
| Arrhythmia | 6(19.35) | 4 (13.79) | 0.73 |
| BNP above cutoff * | 18 (58.06) | 4 (13.79) | 0.0004 |
| Baseline LVEF | 60 (8) | 64 (10) | 0.17 |
| miRNA | Unadjusted p-Value | FC | Adjusted p-Value | OR | 95% CI |
|---|---|---|---|---|---|
| hsa-miR-125a-5p | 0.001 | 12.91 | 0.014 | 1.25 | 1.05–1.48 |
| hsa-miR-15a-5p | 0.003 | 7.16 | 0.026 | 1.22 | 1.02–1.45 |
| hsa-miR-18a-3p | 0.014 | 6.02 | 0.03 | 1.22 | 1.02–1.46 |
| hsa-miR-194-5p | 0.016 | 4.04 | 0.192 | 1.12 | 0.95–1.32 |
| hsa-miR-140-3p | 0.017 | 0.08 | 0.047 | 0.87 | 0.75–1.00 |
| hsa-miR-376c-3p | 0.017 | 0.08 | 0.074 | 0.89 | 0.79–1.01 |
| hsa-miR-10a-5p | 0.023 | 3.71 | 0.218 | 1.11 | 0.94–1.33 |
| hsa-miR-125b-5p | 0.024 | 3.10 | 0.188 | 1.16 | 0.93–1.44 |
| hsa-let-7g-5p | 0.027 | 3.41 | 0.07 | 1.20 | 0.99–1.46 |
| hsa-miR-25-3p | 0.034 | 3.79 | 0.156 | 1.14 | 0.95–1.36 |
| hsa-miR-152-3p | 0.037 | 5.18 | 0.031 | 1.16 | 1.01–1.34 |
| hsa-miR-150-5p | 0.043 | 4.89 | 0.362 | 1.08 | 0.91–1.28 |
| hsa-miR-181a-5p | 0.043 | 2.96 | 0.084 | 1.22 | 0.97–1.53 |
| Target Name | Unadjusted p-Value | FC | SE | Adjusted p-Value | OR | 95% CI |
|---|---|---|---|---|---|---|
| hsa-miR-150-5p | 0.0006 | 8.55 | 0.117 | 0.020 | 1.31 | 1.04–1.65 |
| hsa-miR-452-3p | 0.003 | 8.95 | 0.094 | 0.091 | 1.17 | 0.98–1.41 |
| hsa-miR-153-3p | 0.007 | 7.57 | 0.083 | 0.236 | 1.10 | 0.94–1.30 |
| hsa-miR-222-3p | 0.007 | 3.25 | 0.138 | 0.088 | 1.27 | 0.97–1.66 |
| hsa-miR-15a-5p | 0.009 | 2.87 | 0.083 | 0.117 | 1.14 | 0.97–1.34 |
| hsa-miR-25-3p | 0.013 | 2.37 | 0.115 | 0.124 | 1.19 | 0.95–1.50 |
| hsa-miR-10a-5p | 0.015 | 3.29 | 0.113 | 0.077 | 1.22 | 0.98–1.52 |
| hsa-miR-18a-3p | 0.018 | 5.00 | 0.092 | 0.056 | 1.19 | 1–1.43 |
| hsa-miR-376c-3p | 0.018 | 0.10 | 0.066 | 0.188 | 0.92 | 0.81–1.04 |
| hsa-miR-16-2-3p | 0.019 | 3.39 | 0.094 | 0.137 | 1.15 | 0.96–1.38 |
| hsa-miR-524-3p | 0.019 | 8.43 | 0.063 | 0.215 | 1.08 | 0.96–1.22 |
| hsa-miR-18a-5p | 0.022 | 0.10 | 0.065 | 0.024 | 0.86 | 0.76–0.98 |
| hsa-miR-423-3p | 0.022 | 2.95 | 0.075 | 0.344 | 0.93 | 0.80–1.08 |
| hsa-miR-494-3p | 0.022 | 0.17 | 0.099 | 0.018 | 0.79 | 0.65–0.96 |
| hsa-miR-451a | 0.026 | 3.28 | 0.08 | 0.251 | 1.10 | 0.94–1.28 |
| hsa-let-7g-5p | 0.027 | 2.75 | 0.104 | 0.179 | 1.15 | 0.94–1.41 |
| hsa-miR-125a-5p | 0.028 | 3.87 | 0.081 | 0.150 | 1.12 | 0.96–1.31 |
| hsa-miR-424-5p | 0.029 | 3.41 | 0.082 | 0.399 | 1.07 | 0.91–1.26 |
| hsa-miR-26a-5p | 0.032 | 2.59 | 0.095 | 0.132 | 1.16 | 0.96–1.39 |
| hsa-miR-1180-3p | 0.037 | 3.53 | 0.104 | 0.197 | 1.14 | 0.93–1.40 |
| hsa-miR-499a-5p | 0.037 | 4.22 | 0.102 | 0.075 | 1.20 | 0.98–1.47 |
| hsa-miR-16-5p | 0.039 | 2.20 | 0.094 | 0.137 | 1.15 | 0.96–1.38 |
| hsa-miR-28-3p | 0.039 | 0.15 | 0.079 | 0.161 | 0.89 | 0.77–1.05 |
| hsa-miR-380-3p | 0.044 | 8.83 | 0.064 | 0.183 | 1.09 | 0.96–1.23 |
| miRNA | Baseline | Post-Treatment | ||
|---|---|---|---|---|
| FC | Relative Expression | FC | Relative Expression | |
| hsa-miR-140-3p | 0.08 | Down | 2.0 | Up |
| hsa-miR-598-3p | 3.50 | Up | 0.26 | Down |
| hsa-miR-152-3p | 5.18 | Up | 0.16 | Down |
| hsa-miR-323a | 2.93 | Up | 0.89 | Down |
| hsa-miR-21-5p | 4.51 | Up | 0.12 | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tantawy, M.; Langaee, T.; Wang, D.; Rubinstein, S.M.; Cornell, R.F.; Lenihan, D.; Fradley, M.G.; Gong, Y. Differential Expression of Circulating miRNAs and Carfilzomib-Related Cardiovascular Adverse Events in Patients with Multiple Myeloma. Int. J. Mol. Sci. 2024, 25, 7795. https://doi.org/10.3390/ijms25147795
Tantawy M, Langaee T, Wang D, Rubinstein SM, Cornell RF, Lenihan D, Fradley MG, Gong Y. Differential Expression of Circulating miRNAs and Carfilzomib-Related Cardiovascular Adverse Events in Patients with Multiple Myeloma. International Journal of Molecular Sciences. 2024; 25(14):7795. https://doi.org/10.3390/ijms25147795
Chicago/Turabian StyleTantawy, Marwa, Taimour Langaee, Danxin Wang, Samuel M. Rubinstein, Robert F. Cornell, Daniel Lenihan, Michael G. Fradley, and Yan Gong. 2024. "Differential Expression of Circulating miRNAs and Carfilzomib-Related Cardiovascular Adverse Events in Patients with Multiple Myeloma" International Journal of Molecular Sciences 25, no. 14: 7795. https://doi.org/10.3390/ijms25147795
APA StyleTantawy, M., Langaee, T., Wang, D., Rubinstein, S. M., Cornell, R. F., Lenihan, D., Fradley, M. G., & Gong, Y. (2024). Differential Expression of Circulating miRNAs and Carfilzomib-Related Cardiovascular Adverse Events in Patients with Multiple Myeloma. International Journal of Molecular Sciences, 25(14), 7795. https://doi.org/10.3390/ijms25147795