Abstract
Bovicola ovis, commonly known as the sheep-biting louse, is an ectoparasite that adversely affects the sheep industry. Sheep louse infestation lowers the quality of products, including wool and leather, causing a loss of approximately AUD 123M per annum in Australia alone. The lack of a high-quality genome assembly for the sheep-biting louse, as well as any closely related livestock lice, has hindered the development of louse research and management control tools. In this study, we present the assembly of B. ovis with a genome size of ~123 Mbp based on a nanopore long-read sequencing library and Illumina RNA sequencing, complemented with a chromosome-level scaffolding using the Pore-C multiway chromatin contact dataset. Combining multiple alignment and gene prediction tools, a comprehensive annotation on the assembled B. ovis genome was conducted and recalled 11,810 genes as well as other genomic features including orf, ssr, rRNA and tRNA. A manual curation using alignment with the available closely related louse species, Pediculus humanus, increased the number of annotated genes to 16,024. Overall, this study reported critical genetic resources and biological insights for the advancement of sheep louse research and the development of sustainable control strategies in the sheep industry.
1. Introduction
In Australia, sheep flocks are farmed mainly for meat and wool, with Australian wool and meat production estimated at AUD 3.6B and 4.3B respectively. Bovicola ovis, (formerly Damalinia ovis), also commonly known as the sheep-biting louse, is an ectoparasite that lives and feeds on dead skin, secretions and bacteria normally found on sheep surface. Bovicola ovis is a 1.5 to 2 mm long pale-yellow insect that infests the skin of sheep at the neck, shoulder, sides and back [1]. Host-specific [2] lice infest livestock creating an adverse economic impact on agriculture. Bovicola ovis causes one of the top infestations that economically affect the Australian sheep industry. Infestation of B. ovis in sheep triggers the hosts’ pruritic responses, including rubbing, biting and scratching, causing a reduction in wool quality and yield by up to 1 kg per sheep in a single year [3,4,5]. In Australia alone, it is projected that 23.3% of sheep flocks will be afflicted by sheep lice infestations, which subsequently results in production losses of approximately AUD 123M per annum. Furthermore, farmers will incur additional costs associated with treating the affected sheep [6,7,8].
Sheep lice are primarily diagnosed by wool parting and observation; however, its effectiveness is subject to the inspector’s skills and experiences and is extremely time-consuming with a larger flock [9]. A contributing factor to the challenges associated with sheep lice accurate detection is the small size (2 mm) of the parasite making them difficult to identify. It has been reported that infestations of less than 400 sheep body lice often go unnoticed [10]. Currently, the only control method for sheep lice control is treating sheep with chemical lousicides. However, resistance, lack of specificity, improper use and continual reliance on chemicals make the long-term management of the parasite increasingly problematic for the Australian sheep industry [10]. The rapid emergence of resistance against many insecticides and the increased prevalence of treatment-resistant lice populations pose a challenge to the continued control of sheep lice. Additionally, the potential presence of chemical residues in wool scouring effluent and occupational exposure to chemicals has led to an urgent need for alternative methods for the sustainable control of sheep lice [11,12]. Some alternative control measures like vaccination and compounds like tea tree oil have been evaluated, but no effective solution is yet available against sheep lice [13].
There have been several attempts to improve the detection and diagnosis of sheep lice infestations. For example, an enzyme-linked immunosorbent assay (ELISA) with highly sensitive visual detection was developed and was used commercially as an “on-farm” diagnostic tool [14]. However, the B. ovis ELISA detection assay required a minimum 3-day processing time, costing AUD 134 per sample. Therefore the field application was eventually terminated due to limited utilisation by the sheep producers who preferred the less time-consuming and more cost-effective method of visual identification despite its limited sensitivity [6,14]. Since 2020, various molecular methods, including Polymerase Chain Reaction (PCR), looped-mediated isothermal amplification (LAMP) and quantitative PCR (qPCR), have been used for sheep louse detection [7,15]. A LAMP assay demonstrated a higher percentage of sensitivity and specificity (100% and 75%, respectively) than PCR (80% and 33%, respectively) when crude DNA was used for sheep louse detection, potentially because of the non-specific amplification of target genes posed by other species in the crude DNA extracted by direct boiling of the residues of cutters and combs that were used for shearing [7]. The sensitivity and specificity estimate of sheep lice detection using the PCR assay were improved to 100% when DNA was extracted with a commercial kit. However, using the crude DNA extract and LAMP assay was considered the preferred choice because this combination was less laborious and more time-efficient for field applications. However, this study utilised template DNAs from using 80 mg pooled samples and therefore may only be useful for managing lice infestations at the flock level [8]. It is likely that a sensitive and specific detection at the individual animal level is required for long-term and sustainable control detection of lice infestations.
Another study explored the capability of molecular tests to call out individual infestations [15]. This study reported improved detection power to confidently detect low levels of B. ovis DNA down to 5 × 10−8 and 5 × 10−6 ng/μL for qPCR and LAMP assays, respectively. Additionally, this approach allowed flexible wool collection times and shortened the molecular assay processing time to less than an hour for multiple samples. A field-appropriate fleece swabbing method was developed in the study to replace the original fleece dissolving method, which decreased the sample processing time by 2 h. Although the sensitivity was decreased slightly, when combined, fleece sampling by swabbing and the LAMP assay provided a high-throughput and time-efficient tool for B. ovis detection [15].
The requirement to find more sustainable, safer, pest-specific alternatives to enable improved control, or potential to eradicate, sheep lice remain an unresolved challenge that requires a new approach. One such novel approach for sheep lice control is the application of RNA interference (RNAi) technology. RNAi has demonstrated great promise in combating various pests and diseases including those that affect livestock [16]. RNAi uses short RNA molecules to suppress the expression of genes critical for pests/pathogens. The availability of genomic and transcriptomic information on sheep lice can provide researchers with the opportunity to potentially develop a sustainable species-specific biological control against sheep lice [17]. For example, malaria research accelerated with respect to both diagnostic and disease management when the first reference genome assemblies were published for the malaria parasite, Plasmodium falciparum, and its most important mosquito vector, Anopheles gambiae [18]. Due to the detailed genomic data provided by reference genomes, breakthroughs were achieved in understanding the pathology, epidemiology, drug resistance, vaccine development and disease control [19,20,21].
The aim of this study was the establishment of a reference genome for B. ovis to aid in the development of new biological-based strategies for the prevention and control of lice infestation of sheep. By effectively combining genome sequence data and chromatin conformation information, this research culminated in a meticulously curated and well-annotated B. ovis genome derived using mixed-life stages of B. ovis (nymph, female and male). Notably, this study imparts valuable insights for an array of subsequent inquiries, encompassing not only the fundamental biology of lice but also their epidemiology. Furthermore, this comprehensive genome dataset holds significant promise for pest diagnostic and treatment investigations, which hold substantial potential to enhance the well-being of livestock industries.
2. Results
In total, 50 µL of 92 ng/µL of high molecular weight whole genomic DNA was extracted from the pooled sheep lice sample. Ideally, the reference genome of an organism should be derived from a single cell line or individual in order to avoid technical issues with genetic polymorphisms. However, due to the lack of B. ovis cell lines and the requirement of high molecular weight DNA as the starting material for sequencing, a pool of sheep lice B. ovis was collected for DNA extraction. Approximately 372 Gb of raw sequencing signal data was generated, which resulted in 38.39 Gb of sequence (7.94 million reads). The average N50 of the sequenced reads was 16.4 kb. After filtering out reads that were below 500 bp or had a quality score < 12, as well as reads that mapped to either the prokaryotic or the ovine assemblies, 8.53 Gb bases (~2.23 million reads) were retained for the assembly of the B. ovis genome. GenomeScope 2.0 estimated a haploid genome size of 227.39 Mbp, heterozygosity of 0.62% and 1.62 error rate based on the K-mer distribution on the filtered reads (Figure 1A). After removing duplicated contigs and assembly polishing, the B. ovis genome comprised 788 contigs with a total size of 150,622,374 bp, with a raw read coverage of ×54.
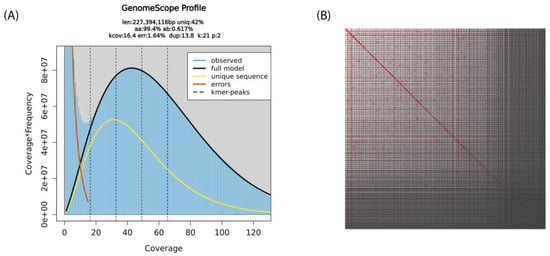
Figure 1.
Schematic representation of the B. ovis genome statistics: (A) Distribution of k-mer frequency in B. ovis sequencing data generated with Nanopore sequencing technology at k-mer size of 21. This GenomeScope profile describes the estimated length (len), percent of the genome that is unique (uniq), rate of homozygosity (aa), rate of heterozygosity (ab), mean k-mer coverage (kcov), error rate (err) and average rate of duplication (dup). (B) The B. ovis genome contig contact matrix using pore-C data. The colour intensity represents the Pore-C contact density in the plot.
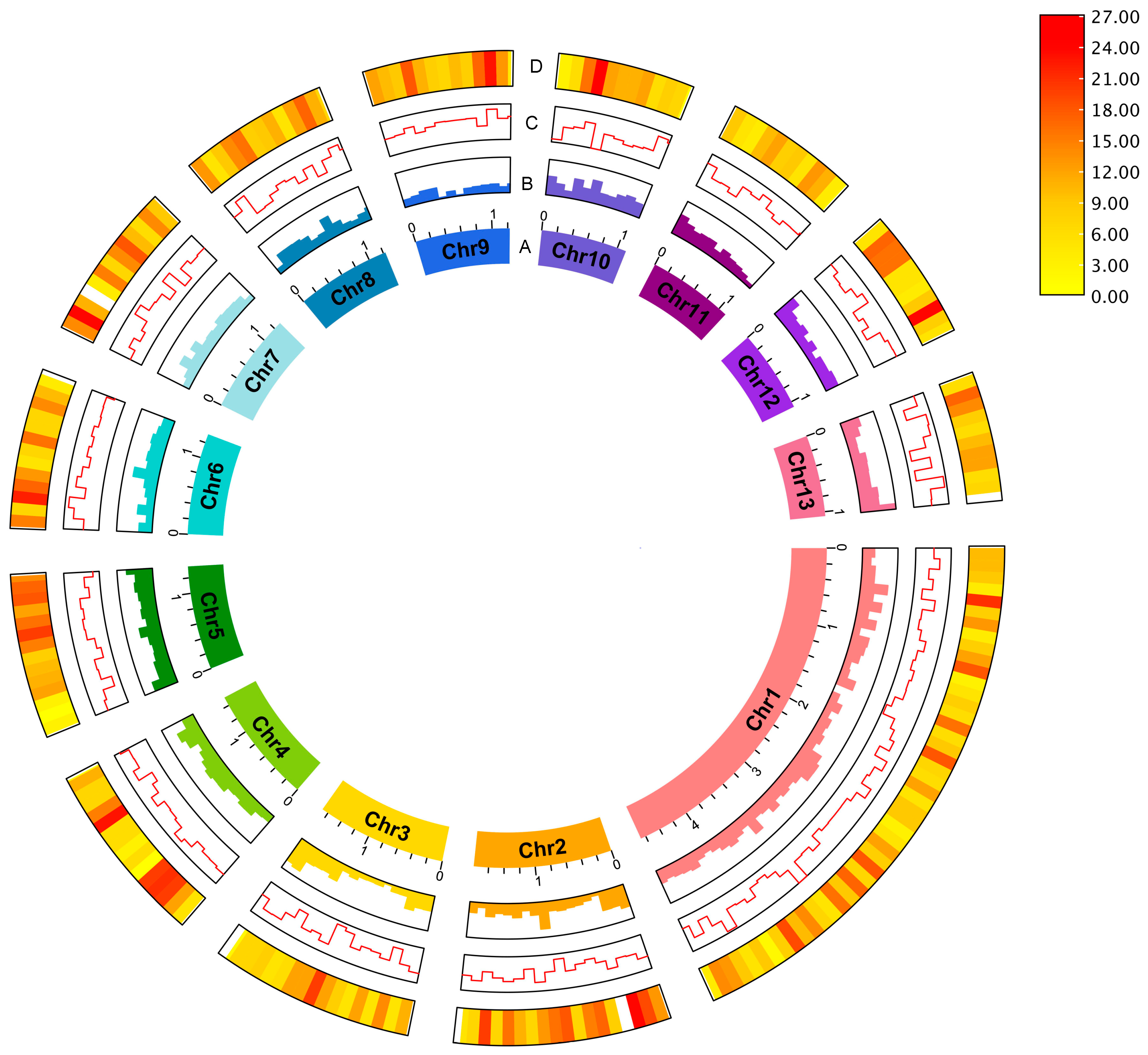
To improve the final assembly, scaffolding of the decontaminated draft assembly using the amplification-free Pore-C data (Figure 1B) and careful curation were performed. The final B. ovis genome comprises 320 scaffolds, with a total genome size of 123,066,105 bp, scaffold N50 of 624 Kb and BUSCO score 96.6% (Figure 2). The largest scaffold was 4.67 Mb in size while the smallest scaffold was 26 Kb. The final B. ovis genome assembly in this study was ~123 Mbp in size, which is consistent with other louse genomes, including P. humanus (~110 Mbp), Brueelia nebulosa (~114 Mbp), Ricinus arcuatus (~155 Mbp) and Columbicola columbae (~208 Mbp) (Table 1). The BUSCO completeness score remained at 96.6% after scaffolding; however, the number of scaffolds was reduced by approximately 60% from 788 to 320 scaffolds.
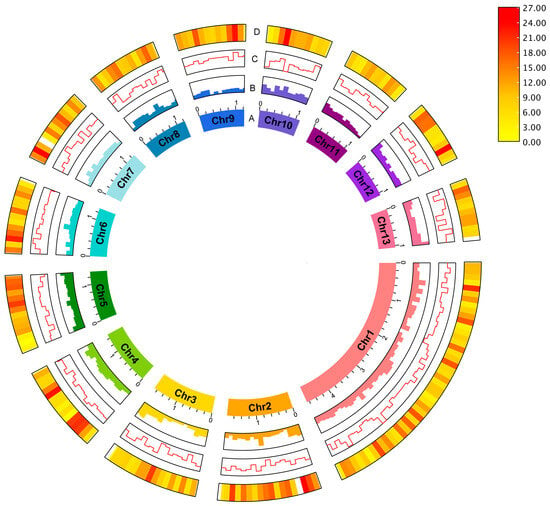
Figure 2.
The sheep-biting louse B. ovis genome. The circus plot shows the genomic features for 13 pseudo-chromosomes: (A) Pseudo-chromosome, (B) GC content, (C) GC skew and (D) gene density. Gene density was represented by a colour gradient ranging from red to yellow, with higher density indicated by red, and lower density indicated by yellow. The pseudo-chromosome size scale is shown in Mb scale.

Table 1.
Genome size and BUSCO score of several louse species.
The low-complexity genome regions and interspersed repeats were identified by homologous comparison using RepeatMasker by searching against the Repbase database of A. gambiae and Drosophila. Approximately 0.05% of interspersed repeats and 11.17% of simple repeats were identified. The repetitive content of B. ovis assembly generated in this study had a higher repetitive element content than P. humanus (~7%) [24] and C. columbae (~9%) [23], but a smaller number of repetitive elements than B. nebulosa (~15%) [22]. A masked consensus was created from the RepeatMasker results.
Evidence-based alignment using either the published P. humanus corporis (GCF_000006295.1), or general dataset including RefSeq [25] and SWISS-PROT [26] was performed to train the gene prediction tools and to provide guidance for manual structural curation. In general, alignments against the RefSeq database reported a higher number of matches compared to alignments against the P. humanus corporis (GCF_000006295.1), regardless of whether transcripts or polypeptides were used (Table 2). Using the P. humanus corporis data, the transcript-based alignment using blastn 2.12.0 [27] and BLAT v2.5 [28] returned 3801 and 241 matched, respectively, while the protein-based alignment using blastx [27] and DIAMOND [29] returned 7253 and 15,863 matches, respectively. In comparison, alignment against the RefSeq invertebrate database reported 110,523 matches with blastn [27], 331,625 matches with BLAT [28] and 6,043,377 matches with the DIAMOND alignment tool [29].
Table 2.
Number of alignments matches identified for Bovicola ovis transcripts or polypeptides against different databases using different search engines.
The reference-based structural gene prediction using AUGUSTUS 3.4.0 [30] based on the D. melanogaster predefined training set only identified 1612 genes, while it identified 20,977 genes based on the customised training set of P. humanus corporis (GCF_000006295.1) (Table 3). Using the alignment results generated by HISAT2 2.2.1 [31] and Tophat 2.1.1 [32], BRAKER2 2.1.5 pipeline [33] predicted 11,184 genes, with an average gene length of 4874 bp, from the 85,358 exons and 73,024 introns identified. The annotation was estimated to be 94.7% complete based on the arthropoda_odb10 dataset by BUSCO 5.2.2 [34]. Ab initio prediction with GeneMarkES 4.48 [35] predicted 11,313 genes from the unsupervised training set and achieved 93.8% completeness by BUSCO 5.2.2 [34] on the arthropoda_odb10 dataset. SNAP 11/29/2013 [36] found only 1438 genes based on the D. melanogaster pre-trained HMM dataset. Other genomic structures, including the open reading frames, simple sequence repeat (SSR) markers, ribosomal RNA (rRNA) and transfer RNA (tRNA) in the masked consensus were annotated (Table 4).
Table 3.
Number of predicted genes, mRNA and polypeptides from the B. ovis genome using different gene prediction tools.
Table 4.
Other genomic features identified within the B. ovis genome.
To obtain a comprehensive gene structure annotation, an official gene set for the B. ovis assembly was produced using EvidenceModeler 1.1.1 [37] by combining the differently weighted functional annotations from the sequence homologies and the structural annotation result generated by the gene prediction tools. PASA [38] was employed to weigh and refine the official predicted gene set using the cDNA transcripts of P. humanus corporis (GCF_000006295.1) (Table 5), which resulted in 11,810 genes, 11,810 mRNA and 11,444 proteins in total. The official gene set (n = 11,810) was functionally annotated using various tools, including the P. humanus corporis (GCF_000006295.1) protein sequences, RefSeq invertebrate [25] and SwissProt [26], using different functional annotation platforms (blastp [27], DIAMOND [29], InterProScan 5.53 [39], Pfam 1.6 [40] and SignalP 5.0 [41] (Table 6). The number of genes identified from the B. ovis assembly in the current study was comparable to that of P. humanus (10,993 genes) and B. nebulosa (10,587 genes) [22,24]. Manual gene curation using blastn transcript alignment against P. humanus (GCF_000006295.1) and RefSeq invertebrate database increased the number of annotated genes to 16,024.
Table 5.
The weight of each annotation tool in generating the consensus sequences using EvidenceModeler 1.1.1 [37].
Table 6.
Functional annotation for the B. ovis genome identified using different tools and against various databases.
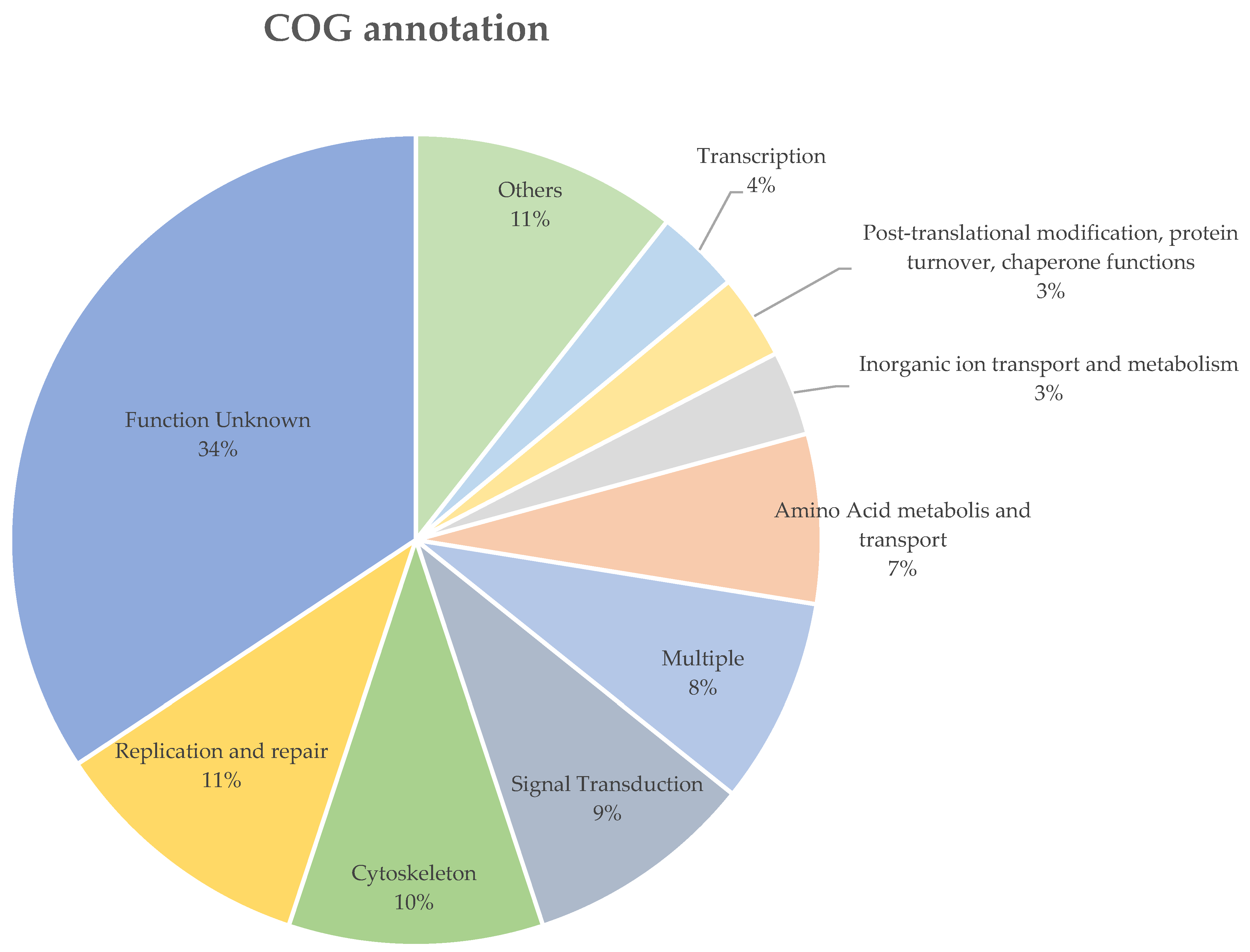
The B. ovis sequenced in this study and the P. humanus corporis (GCF_000006295.1) shared 99.1% and 98.4% of their orthogroups with each other, respectively. There were 541 genes (4.6%), which were grouped in 121 orthogroups, uniquely expressed in B. ovis. In total, only 195 of the 541 uniquely expressed genes in B. ovis were assigned to a COG functional group, of which 71 (36.4%) were labelled as “Function unknown” (Supplementary File S1). The most abundant COG group with known function was “Replication and repair” (n = 22), followed by and “Cytoskeleton” (n = 21), “Signal Transduction” (n = 19) (Figure 3). Additionally, 167 of the unique genes returned no blast results and 170 were labelled as either hypothetical, unnamed proteins or uncharacterized proteins (Supplementary File S2).
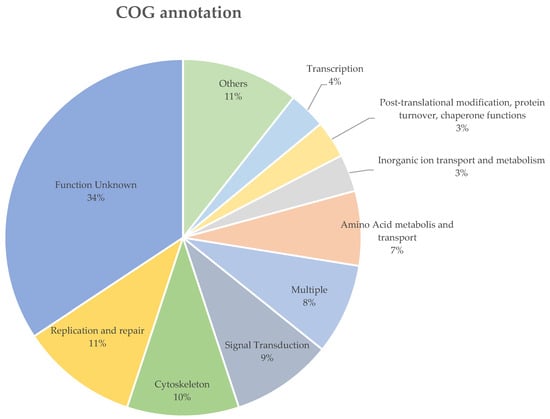
Figure 3.
The COG functional annotation of the uniquely expressed genes in B. ovis. COG groups with less than 4% were grouped as “Others”, the details of these COG groups are listed in Supplementary File S1.
3. Discussion
Here, we present the first genome assembly for the sheep louse (B. ovis). We incorporated innovative approaches, including third-generation sequencing and Pore-C analysis, to generate a high-quality reference assembly and annotation for B. ovis, a pernicious parasite affecting Australian sheep industries. The incorporation of third-generation sequencing enabled a highly continuous de novo genome assembly to be completed. The target organism in this study, B. ovis, previously had no assembled and annotated genome, nor was there any genome assembly from any organism within the same genus. The closest related published genome, Pediculus humanus, which was the human head louse, also belongs to the same taxonomical Order of Phthiraptera as B. ovis. The significance of this research work on the sheep lice genome lies in the fact that it will serve as a valuable reference for other animal lice genomes, as currently there is limited available information on them.
The high BUSCO completeness score and low number of contributing scaffolds achieved here demonstrated the advantages of Pore-C scaffolding for whole-genome assembly by capturing the proximal interactions, which were used to correct and merge the contigs. Therefore, the fragmentation of the assembled genome was minimized, and the chromosome-level assembly was achieved. An increasing number of studies [42,43,44] have incorporated three-dimensional chromatin conformation analysis, including Hi-C and Pore-C, in a whole-genome assembly study to improve the scale of “traditional” phased linear de novo assembly.
In this study, we reported a chromosome-scale genome assembly of the sheep-biting louse, B. ovis, using Nanopore sequencing data and Pore-C analysis. Pore-C captures a comparable number of chromosomal interactions to Hi-C with less sequencing data, demonstrating high efficiency [45,46,47]. Our chromatin conformation analysis showed 58.17% two-way interactions and 41.83% multi-way interactions, similar to human cells (47.56%) [47] and Arabidopsis (44%) [46]. Longer reads with more contacts provide richer multi-dimensional interaction data, enhancing the quality of de novo genome assembly. Although only one Pore-C library was generated due to limited tissue, the high consistency in Pore-C data from Arabidopsis [46], suggests the data here are of sufficient quality for assembly. This highlights Pore-C’s reliability and effectiveness, even with limited samples, justifying its use in complex genomic studies.
Our analysis revealed a relatively high similarity between B. ovis and P. humanus corporis (GCF_000006295.1) as 99.1% of the gene orthologs in B. ovis were also reported in P. humanus corporis (GCF_000006295.1). Further investigation of the unique genes in B. ovis reported several key genes that may be linked to pesticide resistance mechanisms in B. ovis. For instance, two cytochrome b-c1 complex subunit 8-like genes were identified, which are crucial for cellular respiration and energy production. These cytochromes might play significant roles in metabolic adaptations and resistance [48]. Several other genes that can have potential roles in resistance mechanisms have been identified: an ABC transporter (putative), known for mediating multidrug resistance [49] and acetylcholine receptor protein subunit alpha-L1 precurso(putative), potentially linked to neurotoxin resistance [50]; a dual specificity protein phosphatase CDC14 (putative) involved in cell cycle regulation [51]; and glucose dehydrogenase (putative) impacting energy metabolism [52]. Furthermore, identified genes like the ionotropic glutamate receptor, histone-lysine N-methyltransferase SETMAR, and Rho-associated protein kinase can also further our understanding of resistance to insecticides. Overall, these findings provide valuable insights into the molecular mechanisms of B. ovis resistance against control treatments, aiding the development of effective control strategies for the sheep industry.
In conclusion, our study described the assembly of the first B. ovis sheep lice genome. The assembly improvement with the amplification-free multiway chromatin conformation analysis and the comprehensive gene annotation in chromosome-level scaffolds. Since this is the first high-quality genome of ectoparasitic lice that affect livestock, the assembled genome and the genetic characteristics can be referred to as fundamental resources for lice science and inform the advancement of infection management and control, including accurate diagnostic methods and more effective prevention measures in livestock industries. For example, an accurate genome sequence could inform the development of sequence-specific treatments, such as those that utilise RNAi technology to target genes that are essential for sheep lice survival without adversely affecting host or other non-target organisms [53]. Another potential diagnostic approach using the B. ovis genome is adaptive sequencing for specific and accurate detection as demonstrated in a previous study [54]. With the advancement of technologies, a high-quality pest reference genome provides the basis for the future development of time-efficient and cost-effective control strategies against livestock pest diseases, which are crucial to both animal welfare and livestock production.
4. Materials and Methods
4.1. Sheep Louse Sampling
Wool samples from sheep suspected of louse infestation were generously provided by local Queensland sheep farms. A piece of black background cloth was laid out, and a warm lamp was set up above it. The sheep wool was illuminated using the warm lamp, causing the lice to fall onto the black cloth. Additionally, the wool was spread out on the black background cloth, and lice that were firmly attached to the wool were removed with tweezers. All collected samples were then placed into 1.5 mL Eppendorf tubes (Eppendorf, Hamburg, Germany) for the next steps.
4.2. Sheep Louse RNA Extraction
To extract the RNA, approximately 25 mg of sheep lice were added to TRIsure (Bioline, London, UK) prior to homogenisation using TissueLyser II (QIAGEN, Hilden, Germany). Chloroform was added for chemical cell lysis. After incubation and centrifugation, the aqueous phase of RNA was separated, and isopropyl alcohol was added for RNA precipitation. The precipitated RNA was purified with 70% ethanol and treated with DNaseI. The concentration and quality of the RNA were assessed with Nanodrop and agarose gel.
4.3. Sheep Louse High Molecular Weight Genomic DNA Extraction
Cryogenic homogenization was conducted by pulverising sheep lice with a mortar and pestle. The genomic DNA (gDNA) was extracted from approximately 25 mg of the ground sheep lice tissue using Puregene Tissue kit (QIAGEN, Hilden, Germany) by following the manufacturer’s instructions. Briefly, the ground tissue was treated with proteinase K and RNase A before protein precipitation. Isopropanol was added to the supernatant for gDNA precipitation. The precipitated gDNA was purified and stored at −20 °C. The quality and length of the extracted gDNA were observed with pulsed-field gel electrophoresis (Pippin Pulse, Sage Science, MA, USA) and the quantity of the gDNA was measured using QubitTM 4 fluorometer (Thermo Scientific, DE, USA).
4.4. Sheep Louse Chromatin Cross-Linked DNA Extraction
Another ~25 mg of cryo-ground sheep louse tissue was added to 1 mL of pre-chilled phosphate-buffered saline (PBS). Pore-C DNA extraction was conducted as described previously [42]. Briefly, the tissue pellet was resuspended with 1% formaldehyde solution and kept at room temperature for 10 min for cross-linking. The cross-linked sample pellet was treated with protease inhibitor cocktail-permeabilization solution for cell lysis. The suspended pellet was gently mixed with pre-chilled 1.5× digestion reaction buffer, followed by incubation at 65 °C with 1% sodium dodecyl sulphate (SDS), which was quenched by 10% ECOSURF EH-9 later, for chromatin denaturation. Restriction enzyme NlaIII was added to digest the chromatin. The restriction enzyme digestion was subsequently inactivated before the proximity ligation reaction (5% Tween-20, 0.5% SDS and proteinase K) was added to the mixture. Protein degradation reaction was added to the mixture for an incubation period of 18 h at 56 °C to reserve cross-links and degrade proteins. Pore-C DNA extraction DNA extraction was performed with pre-chilled phenol:chloroform:isoamyl alcohol 25:24:1. The pelleted cross-linked DNA was purified with 70% ethanol prior to precipitation.
4.5. Sheep Louse Transcriptomic Sequencing and Data Processing
Before preparing RNA-Seq libraries for sequencing, RNA samples were quantified using the Qubit™ RNA broad-range Assay Kit (Invitrogen, Carlsbad, CA, USA), and QC was performed using the Agilent RNA tapes (#5067-5576) on the TapeStation 4200 (Agilent # G2991AA) as per the manufacturer’s instructions. Total RNA (30 ng) input was used to prepare RNA-Seq libraries using Illumina Stranded Total RNA Prep with Ribo-Zero Plus (Illumina #20040529) as per the manufacturer’s instructions. The only alteration to the protocol was the reduction in the total reaction volume from XX µL to YY µL to accommodate samples with lower concentrations and/or volumes. According to the library preparation protocol and depending on the input RNA amount, 17 PCR cycles were performed to amplify and add indexes and primer sequences in preparation for sequencing. On completion of the library preparation protocol, each library was quantified, and QC was performed using the Quant-iT™ dsDNA HS Assay Kit (Invitrogen) and Agilent D1000 HS tapes (#5067-5582) on the TapeStation 4200 (Agilent # G2991AA) as per the manufacturer’s protocol.
RNA-Seq libraries were pooled at equimolar amounts of 2 nM per library to create a sequencing pool. The library pool was quantified in triplicates using the Qubit™ dsDNA HS Assay Kit (Invitrogen). Library QC is performed using the Agilent D1000 HS tapes (#5067-5582) on the TapeStation 4200 (Agilent # G2991AA) as per the manufacturer’s instructions. The library was prepared for sequencing on the NovaSeq6000 (Illumina) using NovaSeq6000 SP kit v1.5, 2 × 150 bp paired-end chemistry, in the Australian Centre for Ecogenomics according to the manufacturer’s instructions. The RNA reads were processed using Trimmomatic 0.4.0 [55] for a minimum length of 150 bp. FastQC 0.12.1 [56] was used to examine the quality of the reads before and after trimming.
4.6. Oxford Nanopore Long-Read Sequencing and Data Processing
Sequencing libraries were prepared for the extracted sheep louse gDNA and cross-linked DNA using the Ligation Sequencing Kit SQK-LSK-110 (Oxford Nanopore Technologies, Cambridge, UK) with modifications according to an in-house protocol [42]. The sequencing libraries were each loaded onto a PromethION flowcell (Oxford Nanopore Technologies, Cambridge, UK) for long-read sequencing with the MinKNOW software 22.05.10 on a PromethION sequencer (Oxford Nanopore Technologies, Cambridge, UK). The sequencing process was terminated when approximately 38 Gb of data from the gDNA and 5.5 Gb of data from the cross-linked DNA were generated. The modified base-calling from raw signal data was performed using Guppy 6.4.8 on a high-performance computing cluster.
Two forms of DNA contamination were considered: from the host and from the natural microbiota. The basecalled reads were mapped to the prokaryotic assemblies and Ovis aries (GCF_016772045.1) assembly downloaded from RefSeq with Minimap 2.25 [57]. The unmapped reads from the contamination filtering step were recruited for de novo assembly.
4.7. De Novo Assembly of Bovicola ovis Genome
To remove the adapters on the raw reads, Porechop 0.2.4 [58] was employed before the reads were filtered with NanoFilt 2.7.0 [59] for reads with quality greater than or equal to quality score 12. The remaining reads were mapped to the B. ovis mitochondrial minichromosomes (MH001201.1–MH0012012.1) [60] to separate reads belonging to the B. ovis mitochondrial genome and to the B. ovis chromosomes. The haploid genome size and heterozygosity were estimated using GenomeScope 2.0 [61]. The genome reads were filtered to remove those shorter than 500 bp and with a read quality below 12 using Nanofilt v2.8.0 [59] before being assembled with Flye 2.9.1 [62]. To improve the quality, Racon 1.4.3 [63] was implemented to polish the draft assemblies using the respective long-read sequences. Purge_Dups 1.2.6 [64] was used on the polished contigs to remove duplicated haplotypes. The completeness and quality of the complete genomes were evaluated using Samtools 1.10 [65], QUAST 5.0.2 [66] and BUSCO 5.4.5 [34] with the long reads sequenced from gDNA. The identity of the complete genomes was verified by BLAST [67].
4.8. Chromosome Assembly of B. ovis Using Pore-C Chromatin Interaction Mapping Analysis
The Pore-C data were incorporated for the scaffolding of the draft B. ovis genome assembled in this work using the Pore-C analysis pipeline developed by Oxford Nanopore Technologies (ONT) on snakemake workflow [68]. In short, a virtual in silico restriction map with NlaIII was first constructed with the draft assemblies of B. ovis using pore_c refgenome virtual digest tool. The Pore-C reads were mapped to the draft genome using BWA 0.7.17 [69] and the alignments were annotated with and assigned to the respective restriction segments based on the restriction map, thus recovering the multi-way contact. To produce the scaffolding, the long-range multi-way contact was decomposed to pairwise contact information, which were then submitted to SALSA 2.3 [70]. The resulting scaffolds with improved assembly continuity were visualised on Juicebox 2.3.0 [71] and curated accordingly. Scaffolds that were smaller than the reported size of the microchromosomes (0.5–1.5 μm) [72] and had yielded no BUSCO genes were removed from the draft assemblies.
4.9. Annotation
The structural and functional annotation of the draft assembly of B. ovis genome were conducted using the GenSAS 6.0 annotation pipeline [73]. The repetitive elements and non-coding regions were annotated using ab initio prediction with RepeatModeler 2.0.1 [74] and homologous comparison with RepeatMasker 4.1.1 [75]. The interspersed and low complexity repeats were masked with the ab initio database generated from RepeatMasker 4.1.1 [75], and the library of repeat sequences specific to Drosophila and Anopheles gambiae using NCBI/rmblast search engine in quick and sensitive mode. The draft assembly was masked according to the information generated from both RepeatModeler 2.0.1 [74] and RepeatMasker 4.1.1 [75] prior to feature prediction. Species-specific transcript alignment was performed by aligning the RNA-Seq data generated in this study to the draft genome using HISAT2 2.2.1 [31] and Tophat 2.1.1 [32]. Additionally, the species-related RNA and protein alignments were conducted by mapping the published RefSeq data of Pediculus humanus corporis (GCF_000006295.1) and invertebrates using pBLAT 2.5 [28], BLAST+ 2.12.0 [27] and DIAMOND 2.0.6 [29] alignment tools.
The ab initio structural annotations of the draft assembly were predicted by GeneMarkES 4.48 [35], and reference-based structural gene prediction was executed by AUGUSTUS 3.4.0 [30] based on the Drosophila melanogaster predefined training set and a customised training set of P. humanus corporis (GCF_000006295.1). Additionally, the gene prediction from the RNA alignment results from both HISAT2 [31] and Tophat [32] was performed with BRAKER2 2.1.5 [33], which incorporated Augustus [30] and GeneMark 4.48 [35] in its gene prediction pipeline.
EvidenceModeler 1.1.1 [37] was employed to generate a weighted consensus gene structures from the output generated from ab initio gene predictions and reference-based prediction from protein and transcript alignments (Table 5). OrthoFinder v2.5.5 [76] was used to identify the orthologues shared between the B. ovis and P. humanus corporis (GCF_000006295.1). Additionally, the unique genes expressed in B. ovis but not in P. humanus corporis were further investigated manually.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25147824/s1.
Author Contributions
Conceptualization, C.T.O., K.T.M., R.S. and E.M.R.; Methodology, C.T.O., K.T.M., A.S.C., Y.Y. and L.T.N.; Validation, C.T.O., R.S. and E.M.R.; Formal analysis, C.T.O.; Investigation, C.T.O., A.S.C., Y.Y. and E.M.R.; Resources, K.T.M. and R.S.; Data curation, C.T.O.; Writing—original draft, C.T.O.; Writing—review & editing, K.T.M., A.S.C., Y.Y., L.T.N., R.S., N.M., T.J.M. and E.M.R.; Visualization, C.T.O.; Supervision, K.T.M., N.M., T.J.M. and E.M.R.; Project administration, K.T.M.; Funding acquisition, K.T.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Advance Queensland Industry Research Fellowship with grant number CTS 25856/19 and UQ Research Support Package: Strategic Research Investment.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during the current study are available in the NCBI sequence read archive (SRA) database under BioProject PRJNA1121947 and BioSamples SAMN41769694.
Acknowledgments
This project was a part K.T.M.’s Advance Queensland Industry Research Fellowship by Queensland Government, Queensland, Australia. The authors would also like to thank Benjamin Jeal for his contribution during the initial stage of research.
Conflicts of Interest
The authors declare no conflict of interests.
References
- James, P. Biology of Sheep Lice (Bovicola ovis); Australian Wool Innovation Limited: Sydney, NSW, Australia, 2013. [Google Scholar]
- Wall, R.; Shearer, D. Lice (Phthiraptara). In Veterinary Entomology: Arthropod Ectoparasites of Veterinary Importance; Springer Science & Business Media: Dordrecht, The Netherland, 1997. [Google Scholar]
- James, P.J.; Moon, R.D. Pruritus and dermal response to insect antigens in sheep infested with Bovicola ovis. Int. J. Parasitol. 1998, 28, 419–427. [Google Scholar] [CrossRef]
- James, P.J.; Bartholomaeus, F.W.; Karlsson, L.J. Temporal relationship between infestation with lice (Bovicola ovis Schrank) and the development of pruritic behaviour and fleece derangement in sheep. Vet. Parasitol. 2007, 149, 251–257. [Google Scholar] [CrossRef] [PubMed]
- McLeod, R.S. Costs of major parasites to the Australian livestock industries. Int. J. Parasitol. 1995, 25, 1363–1367. [Google Scholar] [CrossRef]
- Popp, S.; Eppleston, J.; Watt, B.R.; Mansfield, S.; Bush, R.D. The prevalence of lice (Bovicola ovis) in sheep flocks on the central and southern Tablelands of New South Wales. Anim. Prod. Sci. 2012, 52, 659–664. [Google Scholar] [CrossRef]
- Wong, S.A.; Woodgate, R.G.; Pant, S.D.; Ghorashi, S.A. Rapid detection of Bovicola ovis using colourimetric loop-mediated isothermal amplification (LAMP): A potential tool for the detection of sheep lice infestation on farm. Parasitol. Res. 2020, 119, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Reeve, I.; Walkden-Brown, S. Benchmarking Australian Sheep Parasite Control: Cross-Sectional Survey Report; Contract No.: B.AHE.0069; Institute for Rural Futures and Animal Science: New South Wales, Australia, 2014. [Google Scholar]
- Morcombe, P.; Young, G.; Ball, M.; Dunlop, R. The detection of lice (Bovicola ovis) in mobs of sheep: A comparison of fleece parting, the lamp test and the table locks test. Aust. Vet. J. 1996, 73, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Joshua, E.; Junk, G.; Garry, L. Sheep Lice (Primefacts); Department of Primary Industries, New South Wales, Australia, 2010.
- Shaw, T. Agricultural chemicals in raw wool and the wool textile industry. Water Environ. J. 1994, 8, 287–290. [Google Scholar] [CrossRef]
- Ranford, S.; Swan, P.; van Koten, C. Chemical residue trends for Australian and New Zealand wool. Sci. Rep. 2022, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- James, P.J.; Callander, J.T. Dipping and jetting with tea tree (Melaleuca alternifolia) oil formulations control lice (Bovicola ovis) on sheep. Vet. Parasitol. 2012, 189, 338–343. [Google Scholar] [CrossRef]
- Wojtek, P.; Michalski, P.Y.; Shiell, B.; Levot, G. Development of a Lice Detection Test for “On-Farm” Use. In Proceedings of the “Flystrike and Lice IPM Control Strategies” Conference, Launceston, Tasmania, Australia, 25–27 June 2001. [Google Scholar]
- Tran, L.; Rawlin, G.T.; Beddoe, T. Development of molecular detection methods of Bovicola ovis from sheep fleece. Parasitol. Res. 2022, 121, 1597–1606. [Google Scholar] [CrossRef]
- Menezes, P.S.; Yan, Y.; Yang, Y.; Mitter, N.; Mahony, T.J.; Mody, K.T. RNAi-based biocontrol of pests to improve the productivity and welfare of livestock production. Appl. Biosci. 2022, 1, 229–243. [Google Scholar] [CrossRef]
- Akoniyon, O.P.; Adewumi, T.S.; Maharaj, L.; Oyegoke, O.O.; Roux, A.; Adeleke, M.A.; Maharaj, R.; Okpeku, M. Whole genome sequencing contributions and challenges in disease reduction focused on malaria. Biology 2022, 11, 587. [Google Scholar] [CrossRef]
- Neafsey, D.E.; Taylor, A.R.; MacInnis, B.L. Advances and opportunities in malaria population genomics. Nat. Rev. Genet. 2021, 22, 502–517. [Google Scholar] [CrossRef]
- Olivucci, G.; Iovino, E.; Innella, G.; Turchetti, D.; Pippucci, T.; Magini, P. Long read sequencing on its way to the routine diagnostics of genetic diseases. Front. Genet. 2024, 15, 1374860. [Google Scholar] [CrossRef]
- Oehler, J.B.; Wright, H.; Stark, Z.; Mallett, A.J.; Schmitz, U. The application of long-read sequencing in clinical settings. Hum. Genom. 2023, 17, 73. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Sweet, A.D.; Browne, D.R.; Hernandez, A.G.; Johnson, K.P.; Cameron, S.L. Draft genome assemblies of the avian louse Brueelia nebulosa and its associates using long-read sequencing from an individual specimen. G3 2023, 13, jkad030. [Google Scholar] [CrossRef]
- Baldwin-Brown, J.G.; Villa, S.M.; Vickrey, A.I.; Johnson, K.P.; Bush, S.E.; Clayton, D.H.; Shapiro, M.D. The assembled and annotated genome of the pigeon louse Columbicola columbae, a model ectoparasite. G3 2021, 11, jkab009. [Google Scholar] [CrossRef]
- Kirkness, E.F.; Haas, B.J.; Sun, W.; Braig, H.R.; Perotti, M.A.; Clark, J.M.; Lee, S.H.; Robertson, H.M.; Kennedy, R.C.; Elhaik, E.; et al. Genome sequences of the human body louse and its primary endosymbiont provide insights into the permanent parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2010, 107, 12168–12173. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Kent, W.J. BLAT-The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2014, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34, W435–W439. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Brůna, T.; Hoff, K.J.; Lomsadze, A.; Stanke, M.; Borodovsky, M. BRAKER2: Automatic eukaryotic genome annotation with GeneMark-EP+ and AUGUSTUS supported by a protein database. NAR Genom. Bioinform. 2021, 3, lqaa108. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Ter-Hovhannisyan, V.; Lomsadze, A.; Chernoff, Y.O.; Borodovsky, M. Gene prediction in novel fungal genomes using an ab initio algorithm with unsupervised training. Genome Res. 2008, 18, 1979–1990. [Google Scholar] [CrossRef]
- Korf, I. Gene finding in novel genomes. BMC Bioinform. 2004, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Salzberg, S.L.; Zhu, W.; Pertea, M.; Allen, J.E.; Orvis, J.; White, O.; Buell, C.R.; Wortman, J.R. Automated eukaryotic gene structure annotation using EVidenceModeler and the program to assemble spliced alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Delcher, A.L.; Mount, S.M.; Wortman, J.R.; Smith, R.K., Jr.; Hannick, L.I.; Maiti, R.; Ronning, C.M.; Rusch, D.B.; Town, C.D.; et al. Improving the Arabidopsis genome annotation using maximal transcript alignment assemblies. Nucleic Acids Res. 2003, 31, 5654–5666. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Jung, H.; Ma, J.; Andersen, S.; Ross, E. Long-read Pore-C shows the 3D structure of the cattle genome. Anim. Prod. Sci. 2023, 63, 972–982. [Google Scholar] [CrossRef]
- Gallardo-Escárate, C.; Valenzuela-Muñoz, V.; Nuñez-Acuña, G.; Valenzuela-Miranda, D.; Gonçalves, A.T.; Escobar-Sepulveda, H.; Liachko, I.; Nelson, B.; Roberts, S.; Warren, W. Chromosome-scale genome assembly of the sea louse Caligus rogercresseyi by SMRT sequencing and Hi-C analysis. Sci. Data 2021, 8, 60. [Google Scholar] [CrossRef]
- Ghurye, J.; Pop, M. Modern technologies and algorithms for scaffolding assembled genomes. PLoS Comput. Biol. 2019, 15, e1006994. [Google Scholar] [CrossRef]
- Wang, C.; Liu, C.; Roqueiro, D.; Grimm, D.; Schwab, R.; Becker, C.; Lanz, C.; Weigel, D. Genome-wide analysis of local chromatin packing in Arabidopsis thaliana. Genome Res. 2015, 25, 246–256. [Google Scholar] [CrossRef]
- Li, Z.; Long, Y.; Yu, Y.; Zhang, F.; Zhang, H.; Liu, Z.; Jia, J.; Mo, W.; Tian, S.Z.; Zheng, M.; et al. Pore-C simultaneously captures genome-wide multi-way chromatin interaction and associated DNA methylation status in Arabidopsis. Plant Biotechnol. J. 2022, 20, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- Ulahannan, N.; Pendleton, M.; Deshpande, A.; Schwenk, S.; Behr, J.M.; Dai, X.; Tyer, C.; Rughani, P.; Kudman, S.; Adney, E.; et al. Nanopore sequencing of DNA concatemers reveals higher-order features of chromatin structure. bioRxiv 2019. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, L.; Shulmeister, V.M.; Chi, Y.I.; Kim, K.K.; Hung, L.W.; Crofts, A.R.; Berry, E.A.; Kim, S.H. Electron transfer by domain movement in cytochrome bc1. Nature 1998, 392, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Dani, J.A. Neuronal nicotinic acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 2015, 124, 3–19. [Google Scholar] [CrossRef]
- Manzano-López, J.; Monje-Casas, F. The multiple roles of the Cdc14 phosphatase in cell cycle control. Int. J. Mol. Sci. 2020, 21, 709. [Google Scholar] [CrossRef] [PubMed]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The glutamate dehydrogenase pathway and its roles in cell and tissue biology in health and disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Meulenaere, K.D.; Cuypers, W.L.; Rosanas-Urgell, A.; Laukens, K.; Cuypers, B. Selective whole-genome sequencing of Plasmodium parasites directly from blood samples by Nanopore adaptive sampling. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrew, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 20 January 2023).
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef] [PubMed]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, H.; Liu, G.-H.; Wang, W.; James, P.; Colwell, D.D.; Tran, A.; Gong, S.; Cai, W.; Shao, R. Mitochondrial genome fragmentation unites the parasitic lice of eutherian mammals. Syst. Biol. 2018, 68, 430–440. [Google Scholar] [CrossRef]
- Ranallo-Benavidez, T.R.; Jaron, K.S.; Schatz, M.C. GenomeScope 2.0 and Smudgeplot for reference-free profiling of polyploid genomes. Nat. Commun. 2020, 11, 1432. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Vaser, R.; Sović, I.; Nagarajan, N.; Šikić, M. Fast and accurate de novo genome assembly from long uncorrected reads. Genome Res. 2017, 27, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; McCarthy, S.A.; Wood, J.; Howe, K.; Wang, Y.; Durbin, R. Identifying and removing haplotypic duplication in primary genome assemblies. Bioinformatics 2020, 36, 2896–2898. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Köster, J.; Rahmann, S. Snakemake—A scalable bioinformatics workflow engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Ghurye, J.; Pop, M.; Koren, S.; Bickhart, D.; Chin, C.-S. Scaffolding of long read assemblies using long range contact information. BMC Genom. 2017, 18, 527. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Turner, D.; Durand, N.C.; Thorvaldsdóttir, H.; Mesirov, J.P.; Aiden, E.L. Juicebox.js provides a cloud-based visualization system for Hi-C data. Cell Syst. 2018, 6, 256–258. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Ahmad, S.F.; Singchat, W.; Panthum, T. Why do some vertebrates have microchromosomes? Cells 2021, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Humann, J.L.; Lee, T.; Ficklin, S.; Main, D. Structural and functional annotation of eukaryotic genomes with GenSAS. Methods Mol. Biol. 2019, 1962, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.M.; Hubley, R.; Goubert, C.; Rosen, J.; Clark, A.G.; Feschotte, C.; Smit, A.F. RepeatModeler2 for automated genomic discovery of transposable element families. Proc. Natl. Acad. Sci. USA 2020, 117, 9451–9457. [Google Scholar] [CrossRef]
- Smit, A.; Hubley, R.; Green, P. RepeatMasker Open-4.0, 2013-2015. Available online: http://www.repeatmasker.org (accessed on 8 August 2023).
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).