Abstract
The metabolic network’s primary sources of free fatty acids (FFAs) are long- and medium-chain fatty acids of triglyceride origin and short-chain fatty acids produced by intestinal microorganisms through dietary fibre fermentation. Recent studies have demonstrated that FFAs not only serve as an energy source for the body’s metabolism but also participate in regulating arterial function. Excess FFAs have been shown to lead to endothelial dysfunction, vascular hypertrophy, and vessel wall stiffness, which are important triggers of arterial hypertension and atherosclerosis. Nevertheless, free fatty acid receptors (FFARs) are involved in the regulation of arterial functions, including the proliferation, differentiation, migration, apoptosis, inflammation, and angiogenesis of vascular endothelial cells (VECs) and vascular smooth muscle cells (VSMCs). They actively regulate hypertension, endothelial dysfunction, and atherosclerosis. The objective of this review is to examine the roles and heterogeneity of FFAs and FFARs in the regulation of arterial function, with a view to identifying the points of intersection between their actions and providing new insights into the prevention and treatment of diseases associated with arterial dysfunction, as well as the development of targeted drugs.
1. Introduction
Cardiovascular diseases (CVDs) are the primary cause of death globally [1,2]. A review of research data indicates that CVDs affect 523 million people globally, with 18.6 million deaths attributed to CVDs in 2019 [3]. CVDs are responsible for approximately 50% of all deaths in developed countries, with the incidence of this disease increasing annually in developing countries [4]. Atherosclerosis is the primary cause of CVD mortality, accounting for 84.9% of all CVD-related deaths [5]. Arterial stiffness and endothelial dysfunction represent the most prevalent forms of CVDs, predominantly resulting from dysfunction of VECs and lipid metabolism. The primary symptoms include vascular stenosis, vascular calcification, thickening of the vessel wall, formation of atheromatous plaques, and plaque rupture, which are the main causes of acute CVDs such as coronary heart disease, cerebral infarction, and cerebral thrombosis [6,7,8]. Serum FFAs have been identified as an independent risk factor for atherosclerosis and arterial dysfunction [9,10,11].
FFAs are physiological ligands for specific G protein-coupled receptors (GPCRs). They serve as an important energy source for human tissues and are involved in the regulation of cellular function, receptor signalling, gene expression, and whole-body energy homeostasis under physiological conditions [12]. The accumulation of saturated FFAs represents a significant contributing factor to lipotoxicity, which in turn results in the dysfunction and apoptosis of VECs [13]. FFAs, also known as non-esterified fatty acids (NEFAs), are independent risk factors for CVDs and are involved in the pathological process of CVDs [14]. Studies have revealed that an excess of fatty acids (FAs) can contribute to the generation of a remnant-rich dyslipidemia and the preconditioning of the arterial intima for lipoprotein deposition via alterations in the expression of matrix proteoglycans. Normalizing FA levels should be a primary objective in the treatment of atherogenic dyslipidemia associated with insulin resistance (IR) [15]. FFARs are receptors for FFAs, including GPR40 (FFAR1), GPR120 (FFAR4), GPR41 (FFAR2), and GPR43 (FFAR3). They are expressed throughout the body and regulate a multitude of biological processes. The activation of FFARs plays a pivotal role in regulating various physiological processes, such as insulin secretion from pancreatic β-cells, adipocyte differentiation, enteroendocrine cells, enteroinsulin secretion, regulation of food intake, adipose tissue biology, inflammation, and neuronal activity [16,17,18,19,20,21,22,23,24]. FFARs play a crucial role in the pathogenesis of various diseases, including diabetes [25,26,27,28], obesity, metabolic syndrome [29,30,31,32], asthma [33], arthritis [34], inflammatory bowel disease [35], and CVDs (arrhythmias, heart failure, hypertension, and myocardial infarction) [36,37,38,39]. A number of agonists of FFARs have been developed and tested in animal models and human trials [40,41,42,43]. Nevertheless, the relevance of FFARs in arterial function has not yet been comprehensively addressed. The objective of this paper is to conduct a comprehensive review of existing studies with the aim of exploring the regulatory role of FFAs and FFARs in arterial function.
2. FFAs and FFARs
2.1. The Classification and Production Pathways of FFAs
FAs are carboxylic acids with long saturated or unsaturated chains. The majority of naturally occurring FAs comprise a straight chain and 4–28 even carbon atoms [44]. FAs can be classified into three categories based on the length of the chain: short-chain fatty acids (SCFAs), medium-chain fatty acids (MCFAs), and long-chain fatty acids (LCFAs) [45] (see Table 1). It should be noted that SCFAs contain six or fewer carbon atoms, while MCFAs contain 6–12 carbon atoms, and LCFAs contain 12 or more carbon atoms [46]. The SCFAs include acetic acid (C2:0), propionic acid (C3:0), butyric acid (C4:0), valeric acid (C5:0), and hexanoic acid (C6:0). In contrast, the MCFAs comprise caprylic acid (C8:0), decanoic acid (C10:0), and lauric acid (C12:0), as well as other saturated fatty acids (SFAs), such as myristic acid (C14:0), palmitic acid (PA) (C16:0), stearic acid (SA) (C18:0), and all types of unsaturated FAs, collectively referred to as LCFAs [22]. FAs can be classified based on the presence of double bonds. SFAs lack double bonds, monounsaturated fatty acids (MUFAs) have only one double bond, and polyunsaturated fatty acids (PUFAs) contain two or more double bonds [47]. Carbon-carbon double-bonded unsaturated fatty acids (UFAs) comprise both cis and trans isomers, with the cis isomer being the predominant configuration of most natural UFAs [48]. FAs are primarily found in adipose tissue and cellular membranes and are components of phospholipids and glycolipids. FFAs are the form in which FAs leave the cell for translocation to other parts of the body for use, usually as NEFAs [49,50]. The geometric differences between various types of SFAs and UFAs play a crucial role in various biological processes and the formation of biological structures, such as cell membranes. It is typical for FFAs to be bound to transport proteins, such as albumin [44,51].

Table 1.
Classification and Production Pathways of FAs.
2.2. The Classification and Associated Signalling Pathways of FFARs
The available evidence indicates that FFARs include GPR40, GPR43, GPR41, and GPR120 [24]. These receptors belong to the retinoid-like family of GPCRs, and each contains seven transmembrane structural domains [45,65,66]. It has been demonstrated that FFARs bind to intracellular proteins, such as heterotrimeric G proteins and β-arrestin, to initiate intracellular signalling cascades [67,68,69] (see Figure 1). GPR40 and GPR120 were activated by LCFAs such as palmitate, oleate, and linoleate, whereas GPR43 and GPR41 were mainly activated by SCFAs (e.g., acetate, butyrate, and propionate) [21,70,71,72]. Currently, only the structure of GPR40 has been revealed by crystallographic methods [73]. In humans, the genes encoding the first three members of the FFAR family are clustered in tandem on the long arm of chromosome 19 (19q13.1) and precede the CD22 gene alignment [74]. The amino acid sequence similarity between GPR40 and GPR41 is 34%, between GPR40 and GPR43 is 41%, and between GPR43 and GPR41 is 52% [74]. The GPR120 gene, located on the long arm of chromosome 10 (10q23.33), exhibits a lower degree of similarity to GPR40 and is known to express both long (BC101175, GPR120L) and short (NM-181745, GPR120S) isoforms in humans [75]. GPR40 and GPR120 can be activated by MCFAs and LCFAs, whereas GPR41 and GPR43 are activated by SCFAs [76,77]. The half-maximal effective concentration (EC50) values for SFCAs are between 0.1 and 1.0 mM for GPR43 and GPR41, while those for LCFAs are 1.0 to 30 μM for GPR40 and GPR120 [78]. In recent times, there has been a great deal of interest in the role of FFARs in the regulation of the cardiovascular system, with a particular focus on the regulation of arterial function.
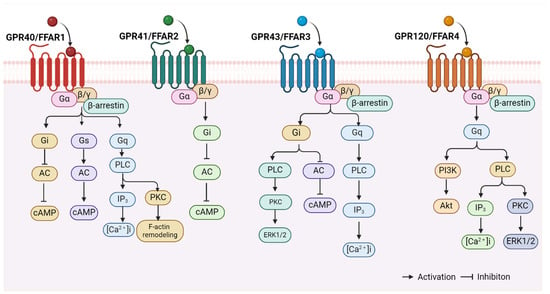
Figure 1.
FFAR-related signalling pathways. FFARs, including GPR40 (FFAR1), GPR43 (FFAR2), GPR41 (FFAR3), and GPR120 (FFAR4), bind to intracellular heterotrimeric G proteins and β-arrestin proteins to initiate intracellular signalling cascades. The figure is created with BioRender. Abbreviations: AC, adenylate cyclase; cAMP, cyclic adenosine monophosphate; PLC, phospholipase C; IP3, inositol triphosphate; [Ca2+]i, intracellular free Ca2+ concentration; PKC, protein kinase C; ERK1/2, extracellular regulatory protein kinase1/2; PI3K, phosphatidylinositol-3-hydroxykinase; Akt/PKB, protein kinase B.
3. Modulation of Arterial Function by FFAs
Dietary FAs serve a multitude of functions within the body. They provide energy, participate in cellular signalling cascades, and function as components of various molecules. Additionally, FAs play a crucial role in mediating inflammation, and maintaining the balance of FA intake is of paramount importance for the preservation of cellular function and tissue homeostasis [79]. A substantial body of research has indicated that diets with excessive amounts of SFAs, high ratios of pro-inflammatory omega-6 PUFAs may result in diet-induced obesity and various obesity-related comorbidities, including type 2 diabetes mellitus, CVDs, and musculoskeletal disorders [80,81,82]. An excessive intake of SFAs represents a significant contributing factor to the development of atherosclerosis. This is due to their ability to stimulate the production of low-density lipoprotein (LDL), which is regarded as the primary risk factor for atherosclerosis [83]. Research has demonstrated that LCFAs increase the risk of coronary heart disease more than SCFAs and MCFAs. Furthermore, excess FFAs exert a detrimental effect on the functionality of VECs, macrophages, and VSMCs in the vessel wall, which in turn impairs arterial function [84]. Consequently, elevated plasma FFA levels represent a significant contributing factor to arterial dysfunction. The regulation of several aspects of arterial function is mediated by FFAs, including endothelial dysfunction, angiogenesis, hypertension, and atherosclerosis.
3.1. Effects of FFAs on Endothelial Dysfunction
Excessive levels of UFAs, such as oleic acid (OA) and linoleic acid (LA), and SFAs, such as PA, have been demonstrated to induce endothelial dysfunction through a number of mechanisms. These include impaired insulin signalling, dysregulated nitric oxide (NO) synthesis, inflammation, activation of the renin-angiotensin system (RAS), and apoptosis. The primary causes of endothelial dysfunction induced by FFAs are inflammation, oxidative stress (OS), and IR [85,86,87]. NO is a vasodilatory factor, and the release of NO induced by changes in intracellular calcium levels plays an important role in the mechanosensitive response of VECs [88]. Free UFAs, such as OA and LA, have been demonstrated to reduce intracellular calcium mobilisation and influx in bovine aortic endothelial cell (BAEC) cultures. This impairment of NO production and release could potentially affect arterial diastole [89]. It has been demonstrated that SFAs can induce injury to VECs and increase the risk of CVDs by promoting atherosclerosis. In contrast, PUFAs, such as docosahexaenoic acid (DHA), are thought to inhibit the injury to VECs that is induced in the early stages of CVDs. Eicosapentaenoic acid (EPA) has been shown to prevent vascular endothelial dysfunction induced by SFAs [11]. Nevertheless, elevated levels of UFAs may also induce apoptosis of VECs through the activation of protein phosphatase 2Cb (PP2Cb) [90]. Ishida and colleagues [91] discovered that human umbilical-vein endothelial cells (HUVECs) stimulated by PA (100 μM) exhibited increased expression of intracellular adhesion molecules, cytokines, and inflammatory factors, including ICAM-1, MCP-1, IL-6, and PTX3. Furthermore, PA stimulates the expression of long-chain acyl-CoA synthetase and the cell cycle protein-dependent kinase inhibitor p21, as well as the phosphorylation of p65, which collectively promote inflammatory responses.
It has been demonstrated that excess FFAs represent a significant source of reactive oxygen species (ROS) in VECs, which can subsequently lead to OS events. These effects were observed at high glucose concentrations [92]. The production of ROS is primarily attributed to the activation of NADPH oxidase by PKC [93]. Additionally, PKC exerts a role in FFA-induced inflammation. Elevated plasma FFA levels have been linked to IKK/nuclear factor kappa B (NF-κB) inflammatory signalling, which can result in the activation of TNF-α, IL1-β, and IL-6, as well as increased MCP-1 plasma levels [94]. These components contribute to the development of chronic inflammation and may result in IR in VECs [16,95]. Another study has demonstrated that elevated FFA levels lead to OS, which impairs vasodilatory NO production and insulin-mediated vasodilation [96]. The administration of 5-aminoimidazole-4-carboxamide riboside has been demonstrated to protect VECs from oxidative damage induced by chronic palmitate overdose [97]. Moreover, physiological doses of oleic and palmitic acids can also protect VECs from OS. The pretreatment and co-treatment of cells with physiological concentrations of PA or OA in the low micromolar range effectively protect cell viability from oxidative damage [98] (see Figure 2). It can be seen from the above that high concentrations of FFAs in the vasculature have the effect of triggering arterial dysfunction. This is achieved by promoting the secretion of adhesion molecules, cytokines, and inflammatory factors; cholesterol transport; and by increasing the levels of OS in VECs. This, in turn, leads to the following consequences: endothelial oxidative damage and endothelial dysfunction. Conversely, physiological doses of FFAs afford VECs protection from OS.
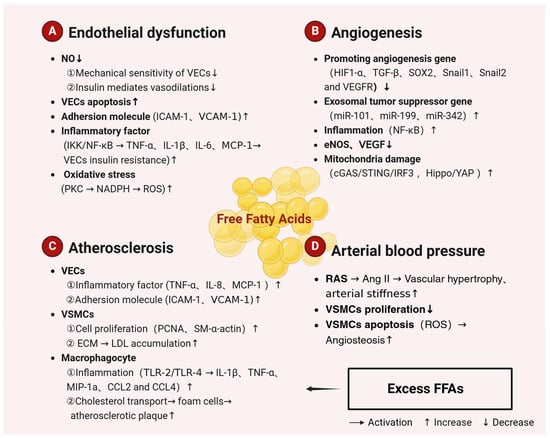
Figure 2.
Mechanisms related to the regulation of arterial function by FFAs. Excessive dietary intake of SFAs is a major cause of atherosclerosis. SFAs trigger endothelial dysfunction, reduce angiogenesis and atherosclerosis, and increase arterial blood pressure, leading to arterial dysfunction. Most UFAs, on the other hand, are protective against CVDs. The figure is created with BioRender. Abbreviations: NO, nitric oxide; VECs, vascular endothelial cells; VSMCs, vascular smooth muscle cells; ICAM-1, intercellular adhesion molecule-1; VCAM-1, vascular cell adhesion molecule-1; IKK, I-kappa-B kinase; NF-κB, nuclear factor kappa B; TNF-α, tumor necrosis factor-α; TNF-β, tumor necrosis factor β; IL, interleukin; MCP-1, monocyte chemoattractant protein-1; NADPH, nicotinamide adenine dinucleotide phosphate; ROS, reactive oxygen species; PCNA, proliferating cell nuclear antigen; ECM, extracellular matrix; TLR-2, Toll-like receptor-2; TLR-4, Toll-like receptor-4; MIP-1α, macrophage inflammatory protein-1α; CCL2, chemokine (C-C motif) ligand 2; CCL4, chemokine (C-C motif) ligand 4; HIF-1α, hypoxia-inducing factor 1α; SOX2, sex determination region Y box protein 2; VEGFR, vascular endothelial growth-factor receptor; cGAS, cyclic-bird adenylate synthetase; STING, stimulator of interferon genes; IRF3, interferon regulatory factor 3; YAP, Yes-associated protein; RAS, renin-angiotensin system; AngII, angiotensin II; SFAs, saturated fatty acids.
3.2. Effects of FFAs on Angiogenesis
Metabolic processes, including glycolysis, fatty acid oxidation, and glutamine metabolism, play distinctive and pivotal roles in angiogenesis [99]. Elevated free fatty acid levels have been demonstrated to cause metabolic stress, which has been shown to inhibit endothelial angiogenesis [100]. DHA, an omega-3 free fatty acid, has anti-angiogenic effects. Human breast cancer cell lines, including MDA-MB-231 and BT-474, were treated with 100 μM DHA for 24 h. In both normoxic and hypoxic conditions, DHA treatment resulted in a significant reduction in the expression of pro-angiogenic genes, including HIF1-α, TGF-β, SOX2, Snail1, Snail2, and VEGFR. Additionally, the expression levels of tumor suppressor microRNAs (miRs), including miR-101, miR-199, and miR-342, were increased, while the expression levels of oncomiRs, including miR-382 and miR-21, were decreased. DHA has the ability to modify the expression and miRs content of pro-angiogenic genes in breast cancer cells and their derived exosomes, thereby inhibiting angiogenesis [101].
In 2023, Wu et al. [102] conducted a study which found that treatment with caffeic acid phenethyl ester (CAPE) significantly inhibited the proliferation, migration, invasion, and angiogenesis of breast cancer MDA-MB-231 cells in the lipopolysaccharide (LPS)-stimulated inflammatory microenvironment, resulting in a decrease in mitochondrial membrane potential. Nevertheless, a study was conducted on the co-culturing of human adipocytes (SW872) cells and human primary myometrial smooth-muscle tumour cells. The study demonstrated that FFAs were transferred from adipocytes to smooth-muscle tumour cells. Additionally, adipocyte-smooth-muscle tumour cell interactions resulted in elevated levels of phosphorylated NF-κB. The phosphorylation of NF-κB plays a key role in the regulation of inflammation, the reorganization of metabolic pathways, and angiogenesis [103]. The Akt, eNOS, and ERK activation responses of 100 µM PA-pretreated HUVECs were markedly diminished in response to VEGF. PA inhibited VEGF-induced angiogenic cord formation in Matrigel [104]. Yuan et al. [100] reported that high concentrations of PA (0.3–0.5 mM) dysregulate the Hippo-YAP pathway, inhibit angiogenesis, and induce mitochondrial damage. Additionally, the researchers discovered that the cytoplasmic DNA sensor cGAS-STING-IRF3 signalling mechanism is activated. Higher levels of circulating OA (400 μM) have been shown to promote angiogenesis and tissue remodelling [105]. Nevertheless, elevated levels of oleic acid have also been associated with the induction of inflammatory and apoptotic processes. Joyal et al. [106] discovered that exposing human retinal pigment epithelial (RPE) ARPE-19 cells to monounsaturated OA at doses greater than 500 μM significantly increased autophagic flux and cell migration. Concurrently, OA treatment stimulated not only AMPK/mTOR/p70S6K signalling but also induced hyperphosphorylation of MAPK pathway mediators, including ERK, JNK, and p38 MAPK, as well as NF-κB activation. Kinase inhibition assays showed that the blockade of the PI3K/Akt, MAPK, and NF-κB pathways prevented the OA-upregulated VEGF transcription and its peptide release (see Figure 2). In conclusion, it appears that metabolic stress induced by elevated, excessive levels of UFAs, such as OA, and SFAs, such as PA, consistently has a negative effect on angiogenesis. Nevertheless, further research is required to fully elucidate the specific evidence and mechanisms involved.
3.3. Effects of FFAs on Atherosclerosis
Elevated plasma FFAs have been linked to a number of conditions, such as atherosclerosis, pancreatic β-cell dysfunction, metabolic syndrome, and IR [107,108,109]. In a study of a multiethnic cohort without baseline CVDs, the relationship between circulating omega-3 and omega-6 poly UFAs and arterial elasticity was examined. The study found that circulating omega-3 and omega-6 PUFAs were associated with large arterial elasticity (LAE) and small arterial elasticity (SAE) in participants of the Multi-Ethnic Study of Atherosclerosis (MESA) [110]. Another study demonstrated that a high-fat, high-cholesterol diet does not induce atherosclerosis in mice with an inactivated Δ6-fatty acid desaturase, which is deficient in omega-3 and omega-6 PUFAs synthesis [111]. FFAs have been shown to contribute to the development of atherosclerosis by affecting the function of VECs, macrophages, and VSMCs [112,113,114].
In VECs, elevated levels of FFAs promote the levels of inflammatory factors such as TNF-α, MCP-1, and IL-8 and increase the secretion of VCAM-1 and ICAM-1 [113]. In a study of VSMCs, Li et al. [115] observed that trilinolein and triolein increased the expression of PCNA and MCP-1 protein and mRNA in human umbilical-vein smooth-muscle cells (HUVSMCs), while concomitantly decreasing SM-α-actin expression. The results demonstrated that tristearin, trilinolein, and triolein significantly promote low-density lipoprotein (ox-LDL)-induced proliferation of HUVSMCs and inhibited atherogenesis. Additionally, it has been demonstrated that elevated levels of FFAs in VSMCs result in a reduction in the production of extracellular matrix, which in turn leads to an increase in LDL accumulation [9,84]. Furthermore, persistent excess of FFAs and glucose in plasma has been identified as a significant contributor to the development of metabolic inflammation and damage-associated molecular patterns (DAMPs) [116,117]. Excessive plasma FFAs induce the production of IL-1β, TNF-α, MIP-1α, CCL2, and CCL4 in macrophages via TRL-2 or TLR-4, thereby triggering arterial inflammation and inducing atherogenesis [118,119,120,121,122]. FFAs are involved in the transformation of macrophages into foam cells by affecting cholesterol transport [113] (see Figure 2). In conclusion, high levels of FFAs play a crucial role in atherosclerosis by regulating inflammation levels, adhesion molecules, cell proliferation, and the extracellular matrix in VECs, VSMCs, and macrophages.
3.4. Effects of FFAs on Arterial Blood Pressure
A number of prospective studies have indicated that elevated serum-FFA concentrations are an independent risk factor for the development of arterial hypertension [96]. The renin-angiotensin system (RAS) plays a crucial role in regulating arterial blood pressure. Angiotensin II (Ang II) has been identified as a potent vasoconstrictor [123]. Ang II has been demonstrated to suppress the oxidative pathway of FAs, which have been shown to activate the RAS [124]. The gene expression of enzymes involved in the breakdown of fats and the oxidation of FAs is elevated in mice lacking in angiotensin-converting enzyme (ACE−/−), which stimulates the generation of FFAs [125]. The release of FFAs induced by the activation of LPL was found to elevate blood pressure [96]. Furthermore, elevated levels of FFAs have been demonstrated to increase OS, resulting in a reduction in the production of vasodilatory NO and impaired insulin-mediated vasodilation. Furthermore, excess FFAs can also stimulate the proliferation of VSMCs, which in turn leads to vascular hypertrophy and stiffness of the vascular wall. Another study demonstrated that high concentrations of PA (100 μM) mediate apoptosis in VSMCs through the activation of the TLR4 pathway and the induction of ROS generation [126]. High-fat diet-caused vascular calcification was associated with PA-induced downregulation of SIRT6. The study found that overexpression of SIRT6 reduced palmitate-induced calcification and apoptosis in VSMCs. Primary VSMCs treated with high concentrations of PA showed decreased expression of SIRT6 and increased expression of BMP2 and RUNX2, resulting in apoptosis and vascular calcification [127] (see Figure 2). This evidence suggests that excess FFAs primarily stimulate the proliferation, migration, apoptosis, and vascular calcification of VSMCs. This leads to vascular hypertrophy and vessel-wall stiffness, which are associated with the development of arterial hypertension.
In conclusion, it is believed that a range of FFAs, such as PUFAs, at normal physiological concentrations, prevent the onset of impairments to arterial function, such as vascular endothelial dysfunction. Nevertheless, elevated levels of FFAs above those considered normal are the primary cause of induced arterial dysfunction. High concentrations of FFAs in the vasculature regulate inflammatory levels, adhesion molecule and cytokine secretion, cholesterol transport, cell proliferation and extracellular matrix formation in VECs, VSMCs, and macrophages. This results in endothelial dysfunction, vascular hypertrophy, and vascular wall stiffness, thereby promoting the development of arterial hypertension and atherosclerosis. Furthermore, it was demonstrated that metabolic stress induced by elevated serum concentrations of FFAs significantly inhibits angiogenesis. Nevertheless, research on the adverse effects of elevated concentrations of FFAs on arterial function has primarily focused on the relationship between FFAs and arterial function, with limited investigation into the intermediate mechanisms. For example, it is unclear whether FFAs exert deleterious effects on arteries through binding to FFARs. Further investigation of these mechanisms is recommended in subsequent studies.
4. Modulation of Arterial Function by FFARs
4.1. GPR40/FFAR1
GPR40 is primarily distributed in VECs, nerves, pancreatic β-cells, enteroendocrine cells, and immune cells [18,19,20,40,128,129,130]. It was demonstrated that GPR40, which is primarily activated by MCFAs and LCFAs, can couple with Gq proteins to initiate downstream PLC activation and promote IP3 or diacylglycerol-induced phosphorylation of PKC, resulting in elevated intracellular Ca2+ levels [131,132]. Consequently, in numerous in vitro studies, the extent of GPR40 activation was quantified based on the intracellular concentration of Ca2+. Additionally, it has been reported that GPR40 can be coupled to Gi and Gs proteins to either inhibit or promote cAMP production by activating AC [133,134,135]. GPR40 signalling is associated with the recruitment of β-arrestin-1 and 2 [136,137]. The coupling of GPR40-Gq activates the ERK1/2 signalling pathway [18,138], which in turn promotes cellular Ca2+ release and ERK1/2 phosphorylation [21].
In the vasculature, GPR40 is a low-affinity receptor for EETs. The epoxidation of EETs by arachidonic acid enhances the agonist activity of GPR40. Activation of GPR40 by 11,12-EET has been demonstrated to increase the expression of Cx43 and COX-2 in VECs, with the promotion of ERK phosphorylation being a key mechanism involved in this process [139]. In a study by Park et al. [140], it was discovered that GPR40 was involved in the secretion of IL-6 in ECs stimulated by PA (100 or 250 μM). Additionally, PA exerts a synergistic effect on LPS-induced IL-6 expression in VECs, which is mediated by MAPK and NF-κB-related signalling pathways. In HUVECs, AM1638, a GPR40 full agonist, enhanced nuclear factor erythroid 2-related factor 2 (NRF2) translocation to the nucleus and heme oxygenase-1 (HO-1) expression. This resulted in the blockade of PA-induced superoxide production and the enhancement of HUVECs viability [141].
Rosiglitazone (RGZ) has been demonstrated to promote the transcription of genes associated with human VECs through the activation of the GPR40/p38 MAPK-related pathway and PPARγ. The induction of PPARγ target genes by RGZ in primary human pulmonary artery endothelial cells was inhibited by knockdown of p38 MAPK or GPR40 [142]. The presence of excessive lipids in the bloodstream was found to impede retinal autophagy, which in turn led to the development of pathological angiogenesis in the Vldlr−/− RAP model. Additionally, the FAs derived from triglycerides and detected by GPR40 were found to restrict autophagy and oxidative metabolism in photoreceptors [143]. The study conducted by Joyal et al. [106] also discovered that in the retinas of Vldlr−/− mice with low fatty-acid uptake but high circulating lipid levels, GPR40 inhibits the expression of the glucose transporter Glut1. This inhibition results in impaired glucose entry into photoreceptors, leading to a shortage of both lipid and glucose fuels and a decrease in the levels of the Krebs cycle intermediate α-ketoglutarate (α-KG). Low levels of alpha-ketoglutarate promote the stabilization of HIF-1α and the secretion of vascular endothelial growth factor A (VEGFA) by starved Vldlr−/− photoreceptors, ultimately resulting in angiogenesis (see Figure 3). In summary, GPR40 primarily regulates inflammation, endothelial dysfunction, and angiogenesis in VECs. There are fewer studies on GPR40’s role in arterial lipid metabolism and its functional regulation of VSMCs.
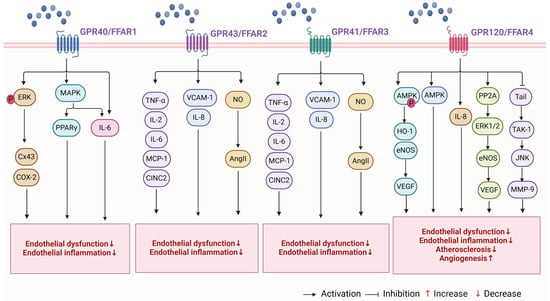
Figure 3.
Mechanisms involved in the regulation of arterial function by FFARs. FFARs bind to arterial FFARs and regulate arterial functions, including endothelial dysfunction, inflammation, angiogenesis, atherosclerosis, and arterial blood pressure. The figure is created with BioRender. Abbreviations: ERK, extracellular signal-regulated kinase; Cx43, gap connexin 43; COX-2, cyclooxygenase-2; MAPK, mitogen-activated protein kinase; PPARγ, peroxisome proliferator-activated receptor γ; AMPK, AMP-activated protein kinase; CREB, cAMP-response element binding protein; PGC1, peroxisome bioactivator receptor gamma coactivator 1; CINC2, cytokine-induced neutrophil chemoattractant 2; HO-1, heme oxygenase-1; PP2A, protein phosphatase 2A; eNOS, endothelial nitric oxide synthase; TAK-1, transforming growth factor-beta activating protein 1; MMP-9, matrix metalloproteinase 9; JNK, c-Jun amino-terminal kinase; P, phosphorylation.
4.2. GPR43/FFAR2
GPR43 is widely expressed in adipose tissue, VECs, liver, enteroendocrine cells, pancreatic islet β-cells, and immune tissues. It plays an important role in a variety of physiological functions, as evidenced by numerous studies [144,145,146,147,148,149]. GPR43 is primarily activated by 61 SCFAs such as acetate, propionate, and butyrate [150,151]. Studies that employ the [Ca2+]i assay to screen for biologically active compounds as ligands have demonstrated that GPR43 can be activated by acetate and other SCFAs, such as propionate and butyrate [152]. Activation of GPR43 by SCFAs inhibits cAMP production and induces the ERK cascade through downstream Gi/o and Gq proteins. In contrast, coupling with Gq proteins promotes elevated Ca2+ concentrations and induces MAPK cascades [152,153,154]. Additionally, research has demonstrated that β-arrestin-2 plays a role in GPR43-dependent signalling [155]. GPR43 also activates ERK1/2 phosphorylation, which in turn promotes MAPK activation and increases Ca2+ concentration [148]. This was demonstrated using a bioluminescence-resonance energy-transfer approach in HEK293 cells co-transfected with eYFP-tagged GPR43 and β-arrestin-2-Renilla luciferase. The results showed that the activation of GPR43 leads to the recruitment of its downstream β-arrestin-2 [155], which inhibits nuclear translocation and promotes the expression of the pro-inflammatory NF-κB [156].
At physiological concentrations (0.1–3 mM), SCFAs can affect leukocytes and endothelial cells by activating GPR41 and GPR43 and inhibiting histone deacetylase (HDAC). SCFAs modulate various leukocyte functions, including the production of inflammatory factors (e.g., TNF-α, IL-2, IL-6, and IL-10), eicosanoids, and chemokines (e.g., MCP-1 and CINC2) [157]. Activation of GPR41/43 is responsible for the effects of acetate on IL-6 and IL-8 production, as well as the effects of butyrate and propionate on IL-6 production. Moreover, inhibition of HDACs mediates the effects of butyrate and propionate on IL-8 production, VCAM-1 expression, and PBMC adhesion to an endothelial monolayer. These findings indicate that SCFAs may play a beneficial role in preventing vascular inflammation and relevant diseases. This is achieved by the activation of GPR41/43 and the inhibition of HDACs [158]. A further study discovered that SCFAs (5–10 mM, comprising acetic acid and butyrate) enhanced Ang II-induced endothelial dysfunction by augmenting endothelial NO bioavailability. This process was associated with butyrate-mediated activation of GPR41/43, whereas acetic acid was not linked to GPR41/43 [159]. Lower levels of the human metabolite-sensitive receptor GPR41/GPR43 have been associated with arterial stiffness, which reduces its response to blood pressure-lowering metabolites, such as butyrate [160] (see Figure 3). This suggests that GPR43 regulates arterial function by affecting the inflammatory process in VECs and modulating Ang II secretion in VSMCs. However, the precise mechanism by which GPR43 regulates arterial function and its role in atherosclerosis remain to be elucidated through further experimentation.
4.3. GPR41/FFAR3
GPR41 is expressed in a wide range of various tissues throughout the body, including pancreatic β-cells, enteroendocrine cells, the peripheral nervous system, immune cells, and the cardiovascular system. It can regulate whole-body energetic homeostasis through SCFA-induced signalling activation [39,161,162]. GPR41 is primarily activated by propionic, butyric, and valeric acids, which are produced by the fermentation of dietary fibre bacteria in the colon [152,163]. In comparison to GPR43, GPR41 is more readily activated by longer-chain SCFAs such as valerate and hexanoate [152]. GPR41 can couple with Gi/o to inhibit cAMP production and promote phosphorylation of ERK1/2 and activation of the βγ subunit [164,165]. GPR41 also appears to couple with Gq/11 proteins, inducing a phosphatidylinositol hydrolysis cascade reaction and stimulating intracellular Ca2+ signalling [39]. Studies utilising proximity linkage analysis, bimolecular fluorescence complementation (BiFC), and fluorescence resonance energy transfer (FRET) have demonstrated that the GPR43/GPR41 heterodimer employs a distinct mechanism of action in comparison to its parental isoform. The heterodimer displays enhanced intracellular Ca2+ signalling and augmented recruitment of β-arrestin-2 yet exhibits diminished capacity to suppress cAMP production [166].
GPR41 is expressed in several peripheral arteries, and its expression is increased following arterial injury. The inactivation of the butyrate receptor GPR41 has been demonstrated to exacerbate the development of neoplastic intimal hyperplasia following injury. Additionally, GPR41-gene deficiency is associated with delayed endothelial recovery in vivo [167]. The vasodilatory effect induced by propionate-activated GPR41 is mediated by Ca2+-dependent eNOS activation and endothelial NO production [168]. A scratch test revealed that treatment of HUVECs with 1-methylcyclopropane carboxylate, an agonist of GPR41, promoted cell migration and proliferation. However, no such effect was observed in VSMCs [169]. Another study found that GPR41 inhibits N-type Ca2+ channels via Gβγ signalling, thereby reducing catecholamine release from rat-sympathetic neurons innervating vascular smooth muscle [170]. Conversely, sodium/glucose cotransporter protein (SGLT)-2 inhibitors, such as dapagliflozin and empagliflozin, are antidiabetic/diuretic drugs that have been demonstrated to possess a multitude of beneficial cardiovascular effects. It is crucial to acknowledge that these drugs stimulate the production of ketone bodies, including β-hydroxybutyrate, in the heart and blood vessels [171,172].
A study on Olfr78 (an olfactory receptor expressed in the renal juxtaglomerular apparatus and activated by SCFAs) and GPR41 demonstrated that vascular GPR41 plays a significant role in regulating blood pressure and vascular tone, as well as in the development of hypertension [173]. In particular, Olfr78 and GPR41 were identified as being expressed in the VSMCs of small resistance vessels. Mice lacking Olfr78 and GPR41 developed hypertension following antibiotic treatment, which reduced the level of SCFAs produced by gut microbial fermentation [173]. Elevated levels of butyric acid (1.4–5.8 mmol/kg) in the colon have been demonstrated to lower arterial blood pressure through colonic vagal signalling and GPR41/43 receptor-related pathways [167]. In a study by Onyszkiewicz et al. [174], it was found that valproic acid (VA), at a physiological concentration of approximately 650 μM in the colonic content and 0.4 μM in the systemic blood, was able to produce vasodilatation and reduce blood pressure when supplemented in rats at a dose of 0.15–0.6 mmol/kg. Furthermore, it was demonstrated that colonic sources of valproic acid rapidly penetrated into tissues involved in the control of blood pressure. These findings are consistent with previous studies that have demonstrated the ability of propionate and other SCFAs to induce vasodilation in vitro, resulting in an acute hypotensive response and antihypertensive protection [175,176]. Nevertheless, the signalling mechanisms responsible for this antihypertensive effect of GPR41 remain to be elucidated. Another study found that a reduction in the availability of intestinal butyrate was linked to vascular remodelling of resistance arteries in hypertensive rats [177] (see Figure 3). In summary, while GPR41 mediates a range of signalling patterns, further research is required to gain a comprehensive understanding of the mechanisms underlying its antihypertensive and vasodilatory effects. Additionally, it is essential to examine the role of GPR41 in pathophysiological processes and ascertain whether receptor heterodimerisation is a prerequisite for its distinctive signalling or function.
4.4. GPR120/FFAR4
GPR120 is predominantly expressed in adipose tissue, enteroendocrine cells, liver, bone, lung, and immune cells [41,42,43]. Studies have demonstrated that GPR120 can be linked to Gq proteins. The activation of GPR120 by synthetic ligands and agonists, such as LCFAs, leads to an increase in intracellular Ca2+ levels without affecting cAMP concentrations [178,179]. FFAs promote glucagon-like peptide-1 (GLP-1) secretion by activating GPR120 in enteroendocrine STC-1 cells. Mice with oral ALA exhibit elevated levels of plasma insulin and GLP-1. To date, no study has demonstrated that GPR120 can be coupled to Gi or Gs proteins. Despite sharing only 10% homology in their amino-acid sequences, GPR120 and GPR40 exhibit similar ligand sensitivity and activity. Furthermore, GPR120 is more readily activated by PUFAs [180,181]. In contrast to GPR120S, which exerts its effects through the Gq/11 and β-arrestin-related pathways, GPR120L exerts its bioefficacy only through the activation of the β-arrestin-related pathway and does not cause an increase in intracellular Ca2+ concentration [182,183]. Additionally, GPR120L is expressed in a limited number of tissues in comparison to GPR120S, which is only detected in the colon. This is in contrast to GPR120S, which is expressed in multiple tissues [131].
With regard to arterial function, GPR120 has been demonstrated to play a protective role in the development of atherosclerosis [184]. In HUVECs and THP-1 monocytes treated with Ginsenoside Rb2, there was an increase in GPR120, AMPK phosphorylation, and HO-1 expression. Furthermore, siRNA silencing of GPR120 resulted in a reduction in Rb2-induced AMPK phosphorylation. Taken together, Rb2 has been demonstrated to reduce inflammation and endothelial stress, inhibit LPS-mediated apoptosis, and decrease the adhesion level of HUVECs through the GPR120/AMPK/HO-1-related pathway [185]. Hwang et al. [186] discovered that protectin DX (PDX) can reverse the production of ROS induced by hydrogen peroxide (H2O2), the Bax/Bcl-2 ratio, and the loss of mitochondrial membrane potential. This leads to an increase in cell viability and a decrease in the release of lactic dehydrogenase (LDH). Furthermore, PDX also enhances the expression and activity of antioxidant proteins such as catalase and superoxide dismutase 2 (SOD2). These proteins were inhibited by AMPK inhibitors and GPR120 antagonists. The evidence suggests that the PDX/AMPK axis protects VECs against OS induced by H2O2. Regarding vascular growth and migration, Zhang et al. [187] discovered that activating GPR120 in human breast cancer cells stimulated the secretion of vascular endothelial growth factor (VEGF) and IL-8, as well as cell migration and epithelial-mesenchymal transition. In HUVECs, 100 μM DHA inhibited VEGF-induced cell migration through the GPR120/PP2A/ERK1/2/eNOS signalling pathway [188].
In VSMCs, the EPA was observed to inhibit the activation of the TAK-1/JNK pathway and the expression of MMP-9. This effect was negated by the knockdown of the GPR120/AF-4 receptor. Additionally, the Trail/TAK-1/JNK/MMP-9 pathway has been demonstrated to promote the development of abdominal aortic aneurysm in OPG KO mice [189]. Another study found that EPA prevented the progression of intracranial aneurysms in rats [190]. Additionally, EPA (20–30 mg/kg) inhibits inflammatory responses induced by acute cerebral infarction by blocking NLRP3 inflammasome activation. Moreover, it attenuated apoptosis induced by acute cerebral infarction by inhibiting NLRP3 inflammatory vesicle activation via GPR40 and GPR120 [191]. In a study conducted by Nakamura et al. [192], the preventive effects of EPA on arterial calcification were confirmed. The study also found that EPA reduces NOX gene expression and activity via GPR120 in klotho mutant mice. In atherosclerosis-related studies, there was a negative correlation between AMPKα2 levels and UBC9 expression in human arteries. In VSMCs, the inactivation of AMPKα2 resulted in increased SUMOylation of the fatty-acid receptor GPR120, which blocked fish oil-induced internalisation and binding to β-arrestin. This suggests that AMPKα2 controls the anti-atherosclerotic effect of fish oil by regulating SUMOylation of GPR120 [193] (see Figure 3). In conclusion, GPR120 regulates the processes of inflammation, lipid metabolism, endothelial adhesion, and migration of VSMCs in VECs and plays a crucial role in angiogenic migration and protection against atherosclerosis and arterial calcification.
In summary, studies investigating the regulation of arterial function by FFARs are currently focused on in vitro cellular experiments. The majority of these studies investigate the beneficial effects of supplementation with SCFAs and UFAs on arterial function through the modulation of FFARs. Limited animal experiments have also indicated that the absence of FFARs is an important reason for the occurrence of arterial dysfunction. To date, studies have not been able to confirm that the deleterious effects of high concentrations of FFAs in the body on arteries are mediated through FFARs.
5. Conclusions
An excess of FFAs is a significant contributor to dyslipidaemia in the body. They affect the secretion of adhesion molecules, cytokines, and inflammatory factors in VECs, cholesterol transport, and promote OS levels. This leads to the development of endothelial inflammation, endothelial cell apoptosis, endothelial oxidative damage, and endothelial dysfunction. FFAs have been demonstrated to stimulate the proliferation, migration, and apoptosis of VSMCs, as well as the accumulation of lipids in VSMCs. This results in vascular hypertrophy and vessel-wall stiffness, which are associated with the development of arterial hypertension and atherosclerosis. These factors are significant contributors to arterial dysfunction. However, studies have indicated that FFARs may be implicated in arterial function-related processes, including the proliferation, differentiation, migration, apoptosis, inflammation, and angiogenesis of VECs and VSMCs. The FFARs play a multifaceted role in the pathogenesis of hypertension, endothelial dysfunction, and atherosclerosis. From the above, it can be concluded that the regulatory effects of excess FFAs on arterial function are predominantly negative, whereas FFARs have a mostly positive effect on arterial function. It is evident that this phenomenon is in opposition to the detrimental effects of elevated concentrations of FFAs on arteries. For example, high levels of PA in the organism adversely affect arterial function, whereas activation of FFARs by high levels of PA positively affects arteries. However, the current evidence does not yet confirm whether the deleterious effects of elevated concentrations of FFAs on arteries are related to their binding to FFARs.
Consequently, the overlapping signalling patterns between FFAs and FFARs, as well as the relationship between these patterns and arterial function, tissue expression patterns, and function, represent a significant challenge to the characterisation of their physiology and pharmacology. The role of FFAs and FFARs in cardiovascular homeostasis is regulatory and adjunctive rather than causal or absolutely essential. However, they play a significant role in regulating biological processes in response to changes in nutritional status and in linking dietary effects to arterial function or CVDs. The regulation of arterial function by FFARs is a relatively new field of study. Although it is known that FFARs can regulate arterial function, the specific direction and molecular mechanisms of this regulation by different receptors remain unclear. Therefore, in the future, this field should aim to validate and explain the specific mechanism by which FFAs regulate arterial function through the activation of FFARs. It is important to provide a scientific explanation of the role of FFARs in regulating arterial function at the molecular level, which will serve as a reliable basis for the development of drugs for arterial dysfunction.
Author Contributions
F.Y. and B.Z. conceived the idea of this review. F.Y. wrote and edited this manuscript and created Figures and Tables. L.J., P.S., D.J. and R.W. reviewed and revised the manuscript. D.J. and R.W. provided direction and guidance throughout the preparation of the manuscript. All authors critically reviewed and edited the content of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported in part by a grant from the National Natural Science Foundation of China (31971097, 32271226), the National Key R&D Program of China (2020YFA0803800), the Shanghai “Science and Technology Innovation Action Plan” Social Development Science and Technology Reach Project (22dz1204600), Shanghai Municipal Science and Technology Committee of Shanghai outstanding academic leaders plan (21XD1403200), Supported by research project of Shanghai University of Sport (2023STD023).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data was used for the research described in the article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AC | adenylate cyclase |
| Akt | serine/threonine kinase |
| AMPK | AMP-activated protein kinase |
| Ang II | angiotensin II |
| cAMP | cyclic adenosine monophosphate |
| [Ca2+]i | intracellular free Ca2+ concentration |
| CAPE | caffeic acid phenethyl ester |
| CCL2 | chemokine (C-C motif) ligand 2 |
| CCL4 | chemokine (C-C motif) ligand 4 |
| cGAS | cyclic bird adenylate synthetase |
| CINC2 | cytokine-induced neutrophil chemoattractant 2 |
| COX-2 | cyclooxygenase-2 |
| CREB | cAMP-response element binding protein |
| CVDs | cardiovascular diseases |
| Cx43 | gap connexin 43 |
| DAMPs | damage-associated molecular patterns |
| DHA | docosahexaenoic acid |
| ECM | extracellular matrix |
| eNOS | endothelial nitric oxide synthase |
| EPA | eicosapentaenoic acid |
| ERK | extracellular signal-regulated kinase |
| ERK1/2 | extracellular regulatory protein kinase1/2 |
| FFARs | free fatty acid receptors |
| FFAs | free fatty acids |
| GLP-1 | glucagon-like peptide-1 |
| GPCRs | G protein-coupled receptors |
| H2O2 | hydrogen peroxide |
| HDAC | histone deacetylase |
| HDAC | histone deacetylase |
| HIF-1α | hypoxia-inducing factor 1α |
| HO-1 | heme oxygenase-1 |
| HUVSMCs | human umbilical vein smooth muscle cells |
| ICAM-1 | intercellular adhesion molecule-1 |
| IKK | I-kappa-B kinase |
| IL | interleukin |
| IP3 | inositol triphosphate |
| IR | insulin resistance |
| IRF3 | interferon regulatory factor 3 |
| JNK | c-Jun amino-terminal kinase |
| KLF2 | Kruppel-like factor 2 |
| LA | linoleic acid |
| LAE | large arterial elasticity |
| LCFAs | long-chain fatty acids |
| LDH | lactic dehydrogenase |
| LPS | lipopolysaccharide |
| MAPK | mitogen-activated protein kinase |
| MCFAs | medium-chain fatty acids |
| MCP-1 | monocyte chemoattractant protein-1 |
| MIP-1α | macrophage inflammatory protein-1α |
| miRs | microRNAs |
| MMP-9 | matrix metalloproteinase 9 |
| MUFAs | monounsaturated fatty acids |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NEFAs | non-esterified fatty acids |
| NF-κB | nuclear factor kappa B |
| NO | nitric oxide |
| NRF2 | nuclear factor erythroid 2-related factor 2HO-1 |
| OA | oleic acid |
| OS | oxidative stress |
| ox-LDL | low-density lipoprotein |
| PA | palmitic acid |
| PCNA | proliferating cell nuclear antigen |
| PGC1 | peroxisome bioactivator receptor gamma coactivator 1 |
| PI3K | phosphatidylinositol-3-hydroxykinase |
| PKC | protein kinase C |
| PLC | phospholipase C |
| PP2Cb | protein phosphatase 2Cb |
| PPARγ | peroxisome proliferator-activated receptor γ |
| PUFAs | polyunsaturated fatty acids |
| RAS | renin-angiotensin system |
| RGZ | Rosiglitazone |
| ROS | reactive oxygen species |
| RPE | retinal pigment epithelial |
| SAE | small arterial elasticity |
| SCFAs | short-chain fatty acids |
| SFAs | saturated fatty acids |
| SOD2 | superoxide dismutase 2 |
| SOX2 | sex determination region Y box protein 2 |
| STING | stimulator of interferon genes |
| TAK-1 | transforming growth factor-beta activating protein 1 |
| TLR-2 | Toll-like receptor-2 |
| TLR-4 | Toll-like receptor-4 |
| TNF-α | tumor necrosis factor α |
| TNF-β | tumor necrosis factor β |
| UFAs | unsaturated fatty acids |
| VA | valproic acid |
| VCAM-1 | vascular cell adhesion molecule-1 |
| VECs | vascular endothelial cells |
| VEGF | vascular endothelial growth factor |
| VEGFA | vascular endothelial growth factor A |
| VEGFR | vascular endothelial growth factor receptor |
| VSMCs | vascular smooth muscle cell |
| YAP | Yes-associated protein |
| α-KG | α-ketoglutarate |
References
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of Dysfunction in the Aging Vasculature and Role in Age-Related Disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef] [PubMed]
- Vogel, B.; Acevedo, M.; Appelman, Y.; Bairey Merz, C.N.; Chieffo, A.; Figtree, G.A.; Guerrero, M.; Kunadian, V.; Lam, C.S.P.; Maas, A.; et al. The Lancet women and cardiovascular disease Commission: Reducing the global burden by 2030. Lancet 2021, 397, 2385–2438. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, E. Obesity and cardiovascular disease. Minerva Pediatr. 2015, 67, 25–32. [Google Scholar] [PubMed]
- Lloyd-Jones, D.; Adams, R.; Carnethon, M.; De Simone, G.; Ferguson, T.B.; Flegal, K.; Ford, E.; Furie, K.; Go, A.; Greenlund, K.; et al. Heart Disease and Stroke Statistics-2009 Update: A Report From the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation 2009, 119, e21–e181, Erratum in Circulation 2011, 124, E424. [Google Scholar]
- Back, M.; Yurdagul, A., Jr.; Tabas, I.; Oorni, K.; Kovanen, P.T. Inflammation and its resolution in atherosclerosis: Mediators and therapeutic opportunities. Nat. Rev. Cardiol. 2019, 16, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Goldsborough, E., 3rd; Osuji, N.; Blaha, M.J. Assessment of Cardiovascular Disease Risk: A 2022 Update. Endocrinol. Metab. Clin. N. Am. 2022, 51, 483–509. [Google Scholar] [CrossRef]
- Greenland, P.; Blaha, M.J.; Budoff, M.J.; Erbel, R.; Watson, K.E. Coronary Calcium Score and Cardiovascular Risk. J. Am. Coll. Cardiol. 2018, 72, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Pilz, S.; Marz, W. Free fatty acids as a cardiovascular risk factor. Clin. Chem. Lab. Med. 2008, 46, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Mallick, R.; Duttaroy, A.K. Modulation of endothelium function by fatty acids. Mol. Cell Biochem. 2022, 477, 15–38. [Google Scholar] [CrossRef]
- Yamagata, K. Fatty acids act on vascular endothelial cells and influence the development of cardiovascular disease. Prostaglandins Other Lipid Mediat. 2023, 165, 106704. [Google Scholar] [CrossRef] [PubMed]
- Haus, J.M.; Solomon, T.P.; Marchetti, C.M.; Edmison, J.M.; Gonzalez, F.; Kirwan, J.P. Free fatty acid-induced hepatic insulin resistance is attenuated following lifestyle intervention in obese individuals with impaired glucose tolerance. J. Clin. Endocrinol. Metab. 2010, 95, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Raja, A.A.; Dandare, A.; Khan, M.J.; Khan, M.J. Free Fatty Acid Overload Targets Mitochondria: Gene Expression Analysis of Palmitic Acid-Treated Endothelial Cells. Genes 2022, 13, 1704. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Curto, E.; Milligan, G. Metabolism meets immunity: The role of free fatty acid receptors in the immune system. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef]
- Rodriguez-Lee, M.; Bondjers, G.; Camejo, G. Fatty acid-induced atherogenic changes in extracellular matrix proteoglycans. Curr. Opin. Lipidol. 2007, 18, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.B.; Cacicedo, J.M.; Itani, S.; Yagihashi, N.; Saha, A.K.; Ye, J.M.; Chen, K.; Zou, M.; Carling, D.; Boden, G.; et al. Malonyl-CoA and AMP-activated protein kinase (AMPK): Possible links between insulin resistance in muscle and early endothelial cell damage in diabetes. Biochem. Soc. Trans. 2003, 31 Pt 1, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Kashihara, D.; Ichimura, A.; Kimura, I.; Tsujimoto, G.; Hirasawa, A. Role of free fatty acid receptors in the regulation of energy metabolism. Biochim. Biophys. Acta 2014, 1841, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H.; et al. Free fatty acids regulate insulin secretion from pancreatic beta cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.M.; Meier, K.E. Free Fatty Acid Receptors and Cancer: From Nutrition to Pharmacology. Handb. Exp. Pharmacol. 2017, 236, 233–251. [Google Scholar] [PubMed]
- Ghislain, J.; Poitout, V. Targeting lipid GPCRs to treat type 2 diabetes mellitus—progress and challenges. Nat. Rev. Endocrinol. 2021, 17, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Kimura, I.; Inoue, D.; Ichimura, A.; Hirasawa, A. Free fatty acid receptors and their role in regulation of energy metabolism. Rev. Physiol. Biochem. Pharmacol. 2013, 164, 77–116. [Google Scholar] [PubMed]
- Miyamoto, J.; Hasegawa, S.; Kasubuchi, M.; Ichimura, A.; Nakajima, A.; Kimura, I. Nutritional Signaling via Free Fatty Acid Receptors. Int. J. Mol. Sci. 2016, 17, 450. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Olefsky, J.M. G protein-coupled receptors as targets for anti-diabetic therapeutics. Nat. Rev. Drug Discov. 2016, 15, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Duah, M.; Zhang, K.; Liang, Y.; Ayarick, V.A.; Xu, K.; Pan, B. Immune regulation of poly unsaturated fatty acids and free fatty acid receptor 4. J. Nutr. Biochem. 2023, 112, 109222. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Villa, S.R.; Fuller, M.; Wicksteed, B.; Mackay, C.R.; Alquier, T.; Poitout, V.; Mancebo, H.; Mirmira, R.G.; Gilchrist, A.; et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 2015, 29, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Ahmed, K.; Gille, A.; Lu, S.; Grone, H.J.; Tunaru, S.; Offermanns, S. Loss of FFA2 and FFA3 increases insulin secretion and improves glucose tolerance in type 2 diabetes. Nat. Med. 2015, 21, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Sun, C.; Gu, W.; Yang, M.; Zhang, X.; Zhai, N.; Lu, Y.; Zhang, Z.; Shou, P.; Zhang, Z.; et al. Free fatty acid receptor 2, a candidate target for type 1 diabetes, induces cell apoptosis through ERK signaling. J. Mol. Endocrinol. 2014, 53, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Li, X.; Weiszmann, J.; Wang, P.; Baribault, H.; Chen, J.L.; Tian, H.; Li, Y. Activation of G protein-coupled receptor 43 in adipocytes leads to inhibition of lipolysis and suppression of plasma free fatty acids. Endocrinology 2008, 149, 4519–4526. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Admyre, T.; Goransson, M.; Marley, A.E.; Smith, D.M.; Oscarsson, J.; Bohlooly, Y.M. Improved glucose control and reduced body fat mass in free fatty acid receptor 2-deficient mice fed a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E211–E220. [Google Scholar] [CrossRef] [PubMed]
- McNelis, J.C.; Lee, Y.S.; Mayoral, R.; van der Kant, R.; Johnson, A.M.; Wollam, J.; Olefsky, J.M. GPR43 Potentiates beta-Cell Function in Obesity. Diabetes 2015, 64, 3203–3217. [Google Scholar] [CrossRef] [PubMed]
- Masui, R.; Sasaki, M.; Funaki, Y.; Ogasawara, N.; Mizuno, M.; Iida, A.; Izawa, S.; Kondo, Y.; Ito, Y.; Tamura, Y.; et al. G protein-coupled receptor 43 moderates gut inflammation through cytokine regulation from mononuclear cells. Inflamm. Bowel Dis. 2013, 19, 2848–2856. [Google Scholar] [CrossRef] [PubMed]
- Halnes, I.; Baines, K.J.; Berthon, B.S.; MacDonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Soluble Fibre Meal Challenge Reduces Airway Inflammation and Expression of GPR43 and GPR41 in Asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.T.; Macia, L.; Galvao, I.; Martins, F.S.; Canesso, M.C.; Amaral, F.A.; Garcia, C.C.; Maslowski, K.M.; De Leon, E.; Shim, D.; et al. A Role for Gut Microbiota and the Metabolite-Sensing Receptor GPR43 in a Murine Model of Gout. Arthritis Rheumatol. 2015, 67, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Meng, H.; Wang, X.; Chen, W.; Tian, X.; Meng, F. Low Expression of FFAR2 in Peripheral White Blood Cells May Be a Genetic Marker for Early Diagnosis of Acute Myocardial Infarction. Cardiol. Res. Pract. 2020, 2020, 3108124. [Google Scholar] [CrossRef]
- Natarajan, N.; Hori, D.; Flavahan, S.; Steppan, J.; Flavahan, N.A.; Berkowitz, D.E.; Pluznick, J.L. Microbial short chain fatty acid metabolites lower blood pressure via endothelial G protein-coupled receptor 41. Physiol. Genom. 2016, 48, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hua, Y.; Ren, J. Short-chain fatty acid propionate alleviates Akt2 knockout-induced myocardial contractile dysfunction. Exp. Diabetes Res. 2012, 2012, 851717. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Shimpukade, B.; Ulven, T.; Hudson, B.D. Complex Pharmacology of Free Fatty Acid Receptors. Chem. Rev. 2017, 117, 67–110. [Google Scholar] [CrossRef] [PubMed]
- Son, S.E.; Kim, N.J.; Im, D.S. Development of Free Fatty Acid Receptor 4 (FFA4/GPR120) Agonists in Health Science. Biomol. Ther. 2021, 29, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, M.; Bender, E.; Schamberger, J.; Eitner, F. Pharmacology of Free Fatty Acid Receptors and Their Allosteric Modulators. Int. J. Mol. Sci. 2021, 22, 1763. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Chavali, S.; Masuho, I.; Jahn, L.J.; Martemyanov, K.A.; Gloriam, D.E.; Babu, M.M. Pharmacogenomics of GPCR Drug Targets. Cell 2018, 172, 41–54.e19. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Mielenz, M. Invited review: Nutrient-sensing receptors for free fatty acids and hydroxycarboxylic acids in farm animals. Animal 2017, 11, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Gao, L.; Thakur, A.; Siu, P.M.; Lai, C.W.K. Role of free fatty acids in endothelial dysfunction. J. Biomed. Sci. 2017, 24, 50. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Gunn, P.J. The regulation of hepatic fatty acid synthesis and partitioning: The effect of nutritional state. Nat. Rev. Endocrinol. 2019, 15, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Wanders, A.J.; Zock, P.L.; Brouwer, I.A. Trans Fat Intake and Its Dietary Sources in General Populations Worldwide: A Systematic Review. Nutrients 2017, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo Rodriguez, J.L.; Dynarowicz-Latka, P.; Minones Conde, J. Structure of unsaturated fatty acids in 2D system. Colloids Surf. B Biointerfaces 2017, 158, 634–642. [Google Scholar] [CrossRef]
- Das, U.N. Can Post-Menopausal Osteoporosis Be Prevented by Essential Fatty Acids? J. Inflamm. Res. 2023, 16, 1553–1554. [Google Scholar] [CrossRef]
- Simard, J.R.; Zunszain, P.A.; Ha, C.E.; Yang, J.S.; Bhagavan, N.V.; Petitpas, I.; Curry, S.; Hamilton, J.A. Locating high-affinity fatty acid-binding sites on albumin by x-ray crystallography and NMR spectroscopy. Proc. Natl. Acad. Sci. USA 2005, 102, 17958–17963. [Google Scholar] [CrossRef] [PubMed]
- Ternes, D.; Tsenkova, M.; Pozdeev, V.I.; Meyers, M.; Koncina, E.; Atatri, S.; Schmitz, M.; Karta, J.; Schmoetten, M.; Heinken, A.; et al. The gut microbial metabolite formate exacerbates colorectal cancer progression. Nat. Metab. 2022, 4, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Formate: The Neglected Member of One-Carbon Metabolism. Annu. Rev. Nutr. 2016, 36, 369–388. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.E.; Faith, J.J.; Bain, J.; Muehlbauer, M.J.; Stevens, R.D.; Newgard, C.B.; Gordon, J.I. Dissecting the in vivo metabolic potential of two human gut acetogens. J. Biol. Chem. 2010, 285, 22082–22090. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Martin, J.C.; Campbell, G.; Mayer, C.D.; Flint, H.J. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium “Roseburia inulinivorans”. J. Bacteriol. 2006, 188, 4340–4349. [Google Scholar] [CrossRef]
- Duncan, S.H.; Barcenilla, A.; Stewart, C.S.; Pryde, S.E.; Flint, H.J. Acetate utilization and butyryl coenzyme A (CoA):acetate-CoA transferase in butyrate-producing bacteria from the human large intestine. Appl. Environ. Microbiol. 2002, 68, 5186–5190. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Araoka, R.; Kajihara, Y.; Ito, T.; Miyamoto, H.; Kodama, H. Valerate production by Megasphaera elsdenii isolated from pig feces. J. Biosci. Bioeng. 2018, 125, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Iino, T.; Mori, K.; Tanaka, K.; Suzuki, K.I.; Harayama, S. Oscillibacter valericigenes gen. nov., sp. nov., a valerate-producing anaerobic bacterium isolated from the alimentary canal of a Japanese corbicula clam. Int. J. Syst. Evol. Microbiol. 2007, 57 Pt 8, 1840–1845. [Google Scholar] [CrossRef]
- Augustin, K.; Khabbush, A.; Williams, S.; Eaton, S.; Orford, M.; Cross, J.H.; Heales, S.J.R.; Walker, M.C.; Williams, R.S.B. Mechanisms of action for the medium-chain triglyceride ketogenic diet in neurological and metabolic disorders. Lancet Neurol. 2018, 17, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Murzyn, A.; Krasowska, A.; Stefanowicz, P.; Dziadkowiec, D.; Lukaszewicz, M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. PLoS ONE 2010, 5, e12050. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Sekine, S.; Kojima, K.; Aoyama, T. The application of medium-chain fatty acids: Edible oil with a suppressing effect on body fat accumulation. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. S1), 320–323. [Google Scholar]
- Janssen, C.I.; Kiliaan, A.J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: The influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 2014, 53, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Ahn, S.; Shukla, A.K.; Lefkowitz, R.J. Molecular mechanism of beta-arrestin-biased agonism at seven-transmembrane receptors. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhou, Q.; Labroska, V.; Qin, S.; Darbalaei, S.; Wu, Y.; Yuliantie, E.; Xie, L.; Tao, H.; Cheng, J.; et al. G protein-coupled receptors: Structure- and function-based drug discovery. Signal Transduct. Target. Ther. 2021, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Nafria, J.; Nehme, R.; Edwards, P.C.; Tate, C.G. Cryo-EM structure of the serotonin 5-HT(1B) receptor coupled to heterotrimeric G(o). Nature 2018, 558, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Jean-Charles, P.Y.; Kaur, S.; Shenoy, S.K. G Protein-Coupled Receptor Signaling Through beta-Arrestin-Dependent Mechanisms. J. Cardiovasc. Pharmacol. 2017, 70, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.J.; Bock, A.; Zaccolo, M. G Protein-Coupled Receptor Signaling: New Insights Define Cellular Nanodomains. Annu. Rev. Pharmacol. Toxicol. 2023, 64, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Hirasawa, A.; Ichimura, A.; Kimura, I.; Tsujimoto, G. Free fatty acid receptors FFAR1 and GPR120 as novel therapeutic targets for metabolic disorders. J. Pharm. Sci. 2011, 100, 3594–3601. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hirasawa, A.; Hara, T.; Tsujimoto, G. Free fatty acid receptors act as nutrient sensors to regulate energy homeostasis. Prostaglandins Other Lipid Mediat. 2009, 89, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Vangaveti, V.; Shashidhar, V.; Jarrod, G.; Baune, B.T.; Kennedy, R.L. Free fatty acid receptors: Emerging targets for treatment of diabetes and its complications. Ther. Adv. Endocrinol. Metab. 2010, 1, 165–175. [Google Scholar] [PubMed]
- Li, Z.; Xu, X.; Huang, W.; Qian, H. Free Fatty Acid Receptor 1 (FFAR1) as an Emerging Therapeutic Target for Type 2 Diabetes Mellitus: Recent Progress and Prevailing Challenges. Med. Res. Rev. 2018, 38, 381–425. [Google Scholar] [CrossRef] [PubMed]
- Bartoszek, A.; Moo, E.V.; Binienda, A.; Fabisiak, A.; Krajewska, J.B.; Mosinska, P.; Niewinna, K.; Tarasiuk, A.; Martemyanov, K.; Salaga, M.; et al. Free Fatty Acid Receptors as new potential therapeutic target in inflammatory bowel diseases. Pharmacol. Res. 2020, 152, 104604. [Google Scholar] [CrossRef]
- Burns, R.N.; Moniri, N.H. Agonism with the omega-3 fatty acids alpha-linolenic acid and docosahexaenoic acid mediates phosphorylation of both the short and long isoforms of the human GPR120 receptor. Biochem. Biophys. Res. Commun. 2010, 396, 1030–1035. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Liou, A.P.; Paziuk, M.; Luevano, J.M., Jr.; Machineni, S.; Turnbaugh, P.J.; Kaplan, L.M. Conserved shifts in the gut microbiota due to gastric bypass reduce host weight and adiposity. Sci. Transl. Med. 2013, 5, 178ra41. [Google Scholar] [CrossRef] [PubMed]
- Dranse, H.J.; Kelly, M.E.; Hudson, B.D. Drugs or diet?—Developing novel therapeutic strategies targeting the free fatty acid family of GPCRs. Br. J. Pharmacol. 2013, 170, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Harasymowicz, N.S.; Dicks, A.; Wu, C.L.; Guilak, F. Physiologic and pathologic effects of dietary free fatty acids on cells of the joint. Ann. N. Y. Acad. Sci. 2019, 1440, 36–53. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Courties, A.; Sellam, J.; Berenbaum, F. Metabolic syndrome-associated osteoarthritis. Curr. Opin. Rheumatol. 2017, 29, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. JPEN J. Parenter. Enteral Nutr. 2015, 39 (Suppl. S1), S18–S32. [Google Scholar] [CrossRef] [PubMed]
- Briggs, M.A.; Petersen, K.S.; Kris-Etherton, P.M. Saturated Fatty Acids and Cardiovascular Disease: Replacements for Saturated Fat to Reduce Cardiovascular Risk. Healthcare 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Oram, J.F.; Bornfeldt, K.E. Direct effects of long-chain non-esterified fatty acids on vascular cells and their relevance to macrovascular complications of diabetes. Front. Biosci. 2004, 9, 1240–1253. [Google Scholar] [CrossRef] [PubMed]
- Iantorno, M.; Campia, U.; Di Daniele, N.; Nistico, S.; Forleo, G.B.; Cardillo, C.; Tesauro, M. Obesity, inflammation and endothelial dysfunction. J. Biol. Regul. Homeost. Agents 2014, 28, 169–176. [Google Scholar]
- Virdis, A. Endothelial Dysfunction in Obesity: Role of Inflammation. High. Blood Press. Cardiovasc. Prev. 2016, 23, 83–85. [Google Scholar] [CrossRef]
- Wang, Y.H.; Liu, Y.P.; Zhu, J.Q.; Zhou, G.H.; Zhang, F.; An, Q.; Yang, J.; Cho, K.W.; Jin, S.N.; Wen, J.F. Physcion prevents high-fat diet-induced endothelial dysfunction by inhibiting oxidative stress and endoplasmic reticulum stress pathways. Eur. J. Pharmacol. 2023, 943, 175554. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C.; Jo, H. Flow-dependent regulation of endothelial nitric oxide synthase: Role of protein kinases. Am. J. Physiol. Cell Physiol. 2003, 285, C499–C508. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, R.; Hirata, K.; Kawashima, S.; Yokoyama, M. Unsaturated free fatty acids inhibit Ca2+ mobilization and NO release in endothelial cells. Kobe J. Med. Sci. 2001, 47, 211–219. [Google Scholar]
- Hufnagel, B.; Dworak, M.; Soufi, M.; Mester, Z.; Zhu, Y.; Schaefer, J.R.; Klumpp, S.; Krieglstein, J. Unsaturated fatty acids isolated from human lipoproteins activate protein phosphatase type 2Cbeta and induce apoptosis in endothelial cells. Atherosclerosis 2005, 180, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Naoe, S.; Nakakuki, M.; Kawano, H.; Imada, K. Eicosapentaenoic Acid Prevents Saturated Fatty Acid-Induced Vascular Endothelial Dysfunction: Involvement of Long-Chain Acyl-CoA Synthetase. J. Atheroscler. Thromb. 2015, 22, 1172–1185. [Google Scholar] [CrossRef]
- Inoguchi, T.; Li, P.; Umeda, F.; Yu, H.Y.; Kakimoto, M.; Imamura, M.; Aoki, T.; Etoh, T.; Hashimoto, T.; Naruse, M.; et al. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C—dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 2000, 49, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Cury-Boaventura, M.F.; Curi, R. Regulation of reactive oxygen species (ROS) production by C18 fatty acids in Jurkat and Raji cells. Clin. Sci. 2005, 108, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139–143. [Google Scholar] [CrossRef]
- Savage, D.B.; Petersen, K.F.; Shulman, G.I. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol. Rev. 2007, 87, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Fagot-Campagna, A.; Balkau, B.; Simon, D.; Warnet, J.M.; Claude, J.R.; Ducimetiere, P.; Eschwege, E. High free fatty acid concentration: An independent risk factor for hypertension in the Paris Prospective Study. Int. J. Epidemiol. 1998, 27, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Samsonov, M.V.; Podkuychenko, N.V.; Khapchaev, A.Y.; Efremov, E.E.; Yanushevskaya, E.V.; Vlasik, T.N.; Lankin, V.Z.; Stafeev, I.S.; Skulachev, M.V.; Shestakova, M.V.; et al. AICAR Protects Vascular Endothelial Cells from Oxidative Injury Induced by the Long-Term Palmitate Excess. Int. J. Mol. Sci. 2021, 23, 211. [Google Scholar] [CrossRef] [PubMed]
- Palomino, O.M.; Giordani, V.; Chowen, J.; Alfonso, S.F.; Goya, L. Physiological Doses of Oleic and Palmitic Acids Protect Human Endothelial Cells from Oxidative Stress. Molecules 2022, 27, 5217. [Google Scholar] [CrossRef]
- Corvera, S.; Solivan-Rivera, J.; Yang Loureiro, Z. Angiogenesis in adipose tissue and obesity. Angiogenesis 2022, 25, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Mao, Y.; Luo, W.; Wu, W.; Xu, H.; Wang, X.L.; Shen, Y.H. Palmitic acid dysregulates the Hippo-YAP pathway and inhibits angiogenesis by inducing mitochondrial damage and activating the cytosolic DNA sensor cGAS-STING-IRF3 signaling mechanism. J. Biol. Chem. 2017, 292, 15002–15015. [Google Scholar] [CrossRef] [PubMed]
- Aslan, C.; Maralbashi, S.; Kahroba, H.; Asadi, M.; Soltani-Zangbar, M.S.; Javadian, M.; Shanehbandi, D.; Baradaran, B.; Darabi, M.; Kazemi, T. Docosahexaenoic acid (DHA) inhibits pro-angiogenic effects of breast cancer cells via down-regulating cellular and exosomal expression of angiogenic genes and microRNAs. Life Sci. 2020, 258, 118094. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, J.; Hao, S.; Guo, Y.; Li, Z.; Liu, Z.; Xuan, H. Caffeic acid phenethyl ester inhibits MDA-MB-231 cell proliferation in inflammatory microenvironment by suppressing glycolysis and lipid metabolism. Biomed. Pharmacother. 2023, 168, 115766. [Google Scholar] [CrossRef] [PubMed]
- Afrin, S.; El Sabah, M.; Manzoor, A.; Miyashita-Ishiwata, M.; Reschke, L.; Borahay, M.A. Adipocyte coculture induces a pro-inflammatory, fibrotic, angiogenic, and proliferative microenvironment in uterine leiomyoma cells. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166564. [Google Scholar] [CrossRef] [PubMed]
- Mehra, V.C.; Jackson, E.; Zhang, X.M.; Jiang, X.C.; Dobrucki, L.W.; Yu, J.; Bernatchez, P.; Sinusas, A.J.; Shulman, G.I.; Sessa, W.C.; et al. Ceramide-activated phosphatase mediates fatty acid-induced endothelial VEGF resistance and impaired angiogenesis. Am. J. Pathol. 2014, 184, 1562–1576. [Google Scholar] [CrossRef] [PubMed]
- Yenuganti, V.R.; Koczan, D.; Vanselow, J. Genome wide effects of oleic acid on cultured bovine granulosa cells: Evidence for the activation of pathways favoring folliculo-luteal transition. BMC Genom. 2021, 22, 486. [Google Scholar] [CrossRef] [PubMed]
- Joyal, J.S.; Sun, Y.; Gantner, M.L.; Shao, Z.; Evans, L.P.; Saba, N.; Fredrick, T.; Burnim, S.; Kim, J.S.; Patel, G.; et al. Retinal lipid and glucose metabolism dictates angiogenesis through the lipid sensor Ffar1. Nat. Med. 2016, 22, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Manabe, I.; Oishi-Tanaka, Y.; Ohsugi, M.; Kono, N.; Ogata, F.; Yagi, N.; Ohto, U.; Kimoto, M.; Miyake, K.; et al. Saturated fatty acid and TLR signaling link beta cell dysfunction and islet inflammation. Cell Metab. 2012, 15, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Rebelos, E.; Mari, A.; Honka, M.J.; Pekkarinen, L.; Latva-Rasku, A.; Laurila, S.; Rajander, J.; Salminen, P.; Iida, H.; Ferrannini, E.; et al. Renal cortical glucose uptake is decreased in insulin resistance and correlates inversely with serum free-fatty acids. J. Clin. Endocrinol. Metab. 2023, 109, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.R.; Wilson, P.W. Free fatty acids and atherosclerosis—guilty or innocent? J. Clin. Endocrinol. Metab. 2006, 91, 2506–2508. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.K.; Guan, W.; Nomura, S.; Weir, N.L.; Karger, A.B.; Duprez, D.; Tsai, M.Y. Associations of plasma omega-3 and omega-6 pufa levels with arterial elasticity: The multi-ethnic study of atherosclerosis. Eur. J. Clin. Nutr. 2022, 76, 1770–1775. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, W.; Binczek, E.; Schmidt-Soltau, I.; Brodesser, S.; Wegner, I. High fat/high cholesterol diet does not provoke atherosclerosis in the omega3-and omega6-polyunsaturated fatty acid synthesis-inactivated Delta6-fatty acid desaturase-deficient mouse. Mol. Metab. 2021, 54, 101335. [Google Scholar] [CrossRef] [PubMed]
- Son, N.H.; Basu, D.; Samovski, D.; Pietka, T.A.; Peche, V.S.; Willecke, F.; Fang, X.; Yu, S.Q.; Scerbo, D.; Chang, H.R.; et al. Endothelial cell CD36 optimizes tissue fatty acid uptake. J. Clin. Investig. 2018, 128, 4329–4342. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, E.E.; Gaubatz, J.W.; Burns, A.R.; Pownall, H.J. Sustained elevations in NEFA induce cyclooxygenase-2 activity and potentiate THP-1 macrophage foam cell formation. Atherosclerosis 2007, 192, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.I.; Lee, Y.H.; Chen, P.H.; Lin, Y.C.; Chou, M.H.; Kao, Y.H. Free Fatty Acids Induce Autophagy and LOX-1 Upregulation in Cultured Aortic Vascular Smooth Muscle Cells. J. Cell Biochem. 2017, 118, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pan, Y.; Luo, K.; Luo, T.; Fan, Y.; Deng, Z. Effects of Different Simple Triglycerides on Cell Fatty Acid Compositions, Proliferation-Related Protein, and Gene Expressions Induced by Oxidized-LDL in HUVSMCs. J. Food Sci. 2017, 82, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L. Complexity of danger: The diverse nature of damage-associated molecular patterns. J. Biol. Chem. 2014, 289, 35237–35245. [Google Scholar] [CrossRef] [PubMed]
- Lyons, C.L.; Kennedy, E.B.; Roche, H.M. Metabolic Inflammation-Differential Modulation by Dietary Constituents. Nutrients 2016, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef]
- Lee, J.Y.; Zhao, L.; Youn, H.S.; Weatherill, A.R.; Tapping, R.; Feng, L.; Lee, W.H.; Fitzgerald, K.A.; Hwang, D.H. Saturated fatty acid activates but polyunsaturated fatty acid inhibits Toll-like receptor 2 dimerized with Toll-like receptor 6 or 1. J. Biol. Chem. 2004, 279, 16971–16979. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Akhter, N.; Al-Roub, A.; Kochumon, S.; Wilson, A.; Thomas, R.; Ali, S.; Tuomilehto, J.; Sindhu, S. MIP-1alpha Induction by Palmitate in the Human Monocytic Cells Implicates TLR4 Signaling Mechanism. Cell Physiol. Biochem. 2019, 52, 212–224. [Google Scholar] [PubMed]
- Kochumon, S.; Wilson, A.; Chandy, B.; Shenouda, S.; Tuomilehto, J.; Sindhu, S.; Ahmad, R. Palmitate Activates CCL4 Expression in Human Monocytic Cells via TLR4/MyD88 Dependent Activation of NF-kappaB/MAPK/ PI3K Signaling Systems. Cell Physiol. Biochem. 2018, 46, 953–964. [Google Scholar] [CrossRef]
- Yang, W.; Cao, M.; Mao, X.; Wei, X.; Li, X.; Chen, G.; Zhang, J.; Wang, Z.; Shi, J.; Huang, H.; et al. Alternate-day fasting protects the livers of mice against high-fat diet-induced inflammation associated with the suppression of Toll-like receptor 4/nuclear factor kappaB signaling. Nutr. Res. 2016, 36, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Luscher, T.F. Endothelial dysfunction: The role and impact of the renin-angiotensin system. Heart 2000, 84 (Suppl. S1), i20–i22; discussion i50. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pellieux, C.; Montessuit, C.; Papageorgiou, I.; Lerch, R. Angiotensin II downregulates the fatty acid oxidation pathway in adult rat cardiomyocytes via release of tumour necrosis factor-alpha. Cardiovasc. Res. 2009, 82, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, S.; Kobayashi, T.; Taguchi, K.; Matsumoto, T.; Kamata, K. Losartan improves aortic endothelium-dependent relaxation via proline-rich tyrosine kinase 2/Src/Akt pathway in type 2 diabetic Goto-Kakizaki rats. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2383–H2394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, G.; Zhang, Y.; Liu, J.; Liu, X.; Li, W.; Lv, Y.; Wei, S.; Liu, J.; Quan, J. Palmitate induces VSMC apoptosis via toll like receptor (TLR)4/ROS/p53 pathway. Atherosclerosis 2017, 263, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wu, Y.; Jiang, C.; Wang, Q.; Geng, X.; Chen, L.; Zhou, S.; Wang, X.; Han, M.; Du, D.; et al. Saturated fatty acid promotes calcification via suppressing SIRT6 expression in vascular smooth muscle cells. J. Hypertens. 2023, 41, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, A.; Hara, T.; Ichimura, A.; Tsujimoto, G. Free Fatty Acid Receptors and Their Physiological Role in Metabolic Regulation. Yakugaku Zasshi 2011, 131, 1683–1689. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, J.W.; Roh, E.; Choi, K.M.; Yoo, H.J.; Hwang, H.J.; Baik, S.H. GPR40 Agonism Modulates Inflammatory Reactions in Vascular Endothelial Cells. Diabetes Metab. J. 2022, 46, 506–511. [Google Scholar] [CrossRef]
- Rady, B.; Liu, J.; Huang, H.; Bakaj, I.; Qi, J.; Lee, S.P.; Martin, T.; Norquay, L.; Player, M.; Pocai, A. A FFAR1 full agonist restores islet function in models of impaired glucose-stimulated insulin secretion and diabetic non-human primates. Front. Endocrinol. 2022, 13, 1061688. [Google Scholar] [CrossRef] [PubMed]
- Galindo, M.M.; Voigt, N.; Stein, J.; van Lengerich, J.; Raguse, J.D.; Hofmann, T.; Meyerhof, W.; Behrens, M. G protein-coupled receptors in human fat taste perception. Chem. Senses 2012, 37, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.; Shachar, S.; Sekler, I.; Hershfinkel, M.; Walker, M.D. Role of GPR40 in fatty acid action on the beta cell line INS-1E. Biochem. Biophys. Res. Commun. 2005, 335, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Schroder, R.; Schmidt, J.; Blattermann, S.; Peters, L.; Janssen, N.; Grundmann, M.; Seemann, W.; Kaufel, D.; Merten, N.; Drewke, C.; et al. Applying label-free dynamic mass redistribution technology to frame signaling of G protein-coupled receptors noninvasively in living cells. Nat. Protoc. 2011, 6, 1748–1760. [Google Scholar] [CrossRef]
- Feng, D.D.; Luo, Z.; Roh, S.G.; Hernandez, M.; Tawadros, N.; Keating, D.J.; Chen, C. Reduction in voltage-gated K+ currents in primary cultured rat pancreatic beta-cells by linoleic acids. Endocrinology 2006, 147, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.E.; Pedersen, M.H.; Dmytriyeva, O.; Nellemose, E.; Arora, T.; Engelstoft, M.S.; Asher, W.B.; Javitch, J.A.; Schwartz, T.W.; Trauelsen, M. Free fatty acid receptor 1 stimulates cAMP production and gut hormone secretion through Gq-mediated activation of adenylate cyclase 2. Mol. Metab. 2023, 74, 101757. [Google Scholar] [CrossRef]
- Hauge, M.; Vestmar, M.A.; Husted, A.S.; Ekberg, J.P.; Wright, M.J.; Di Salvo, J.; Weinglass, A.B.; Engelstoft, M.S.; Madsen, A.N.; Luckmann, M.; et al. GPR40 (FFAR1)—Combined Gs and Gq signaling in vitro is associated with robust incretin secretagogue action ex vivo and in vivo. Mol. Metab. 2015, 4, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.D.; Bertrand, G.; Vivot, K.; Carpentier, E.; Tremblay, C.; Ghislain, J.; Bouvier, M.; Poitout, V. beta-Arrestin Recruitment and Biased Agonism at Free Fatty Acid Receptor 1. J. Biol. Chem. 2015, 290, 21131–21140. [Google Scholar] [CrossRef] [PubMed]
- Karmokar, P.F.; Moniri, N.H. Free-fatty acid receptor-1 (FFA1/GPR40) promotes papillary RCC proliferation and tumor growth via Src/PI3K/AKT/NF-kappaB but suppresses migration by inhibition of EGFR, ERK1/2, STAT3 and EMT. Cancer Cell Int. 2023, 23, 126. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Herrnreiter, A.; Pfister, S.L.; Gauthier, K.M.; Falck, B.A.; Falck, J.R.; Campbell, W.B. GPR40 is a low-affinity epoxyeicosatrienoic acid receptor in vascular cells. J. Biol. Chem. 2018, 293, 10675–10691. [Google Scholar] [CrossRef]
- Lu, Z.; Li, Y.; Jin, J.; Zhang, X.; Hannun, Y.A.; Huang, Y. GPR40/FFA1 and neutral sphingomyelinase are involved in palmitate-boosted inflammatory response of microvascular endothelial cells to LPS. Atherosclerosis 2015, 240, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Kim, J.W.; Yun, S.; Park, M.J.; Song, E.; Jang, S.; Jang, A.; Choi, K.M.; Baik, S.H.; Yoo, H.J. AM1638, a GPR40-Full Agonist, Inhibited Palmitate- Induced ROS Production and Endoplasmic Reticulum Stress, Enhancing HUVEC Viability in an NRF2-Dependent Manner. Endocrinol. Metab. 2023, 38, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Awad, K.S.; Elinoff, J.M.; Dougherty, E.J.; Ferreyra, G.A.; Wang, J.Y.; Cai, R.; Sun, J.; Ptasinska, A.; Danner, R.L. G Protein-coupled Receptor 40 (GPR40) and Peroxisome Proliferator-activated Receptor gamma (PPARgamma): AN INTEGRATED TWO-RECEPTOR SIGNALING PATHWAY. J. Biol. Chem. 2015, 290, 19544–19557. [Google Scholar] [CrossRef] [PubMed]
- Heckel, E.; Cagnone, G.; Agnihotri, T.; Cakir, B.; Das, A.; Kim, J.S.; Kim, N.; Lavoie, G.; Situ, A.; Pundir, S.; et al. Triglyceride-derived fatty acids reduce autophagy in a model of retinal angiomatous proliferation. JCI Insight 2022, 7, e154174. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Li, A.; Wang, H.; Ma, J.; Hou, J. Positive Regulation of Acetate in Adipocyte Differentiation and Lipid Deposition in Obese Mice. Nutrients 2023, 15, 3736. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Zhang, X.; Liu, W.; Wei, H.; Liang, W.; Zhou, Y.; Ding, Y.; Ji, F.; Ho-Kwan Cheung, A.; Wong, N.; et al. Bifidobacterium pseudolongum-generated acetate suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma. J. Hepatol. 2023, 79, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Teyani, R.; Moniri, N.H. Gut feelings in the islets: The role of the gut microbiome and the FFA2 and FFA3 receptors for short chain fatty acids on beta-cell function and metabolic regulation. Br. J. Pharmacol. 2023, 180, 3113–3129. [Google Scholar] [CrossRef] [PubMed]
- Prado, C.; Espinoza, A.; Martinez-Hernandez, J.E.; Petrosino, J.; Riquelme, E.; Martin, A.J.M.; Pacheco, R. GPR43 stimulation on TCRalphabeta(+) intraepithelial colonic lymphocytes inhibits the recruitment of encephalitogenic T-cells into the central nervous system and attenuates the development of autoimmunity. J. Neuroinflamm. 2023, 20, 135. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA Receptor GPR43 and Energy Metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef]
- Voltolini, C.; Battersby, S.; Etherington, S.L.; Petraglia, F.; Norman, J.E.; Jabbour, H.N. A novel antiinflammatory role for the short-chain fatty acids in human labor. Endocrinology 2012, 153, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.D.; Tikhonova, I.G.; Pandey, S.K.; Ulven, T.; Milligan, G. Extracellular ionic locks determine variation in constitutive activity and ligand potency between species orthologs of the free fatty acid receptors FFA2 and FFA3. J. Biol. Chem. 2012, 287, 41195–41209. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.P.; Karunakar, P.; Taraphder, S.; Yadav, H. Free Fatty Acid Receptors 2 and 3 as Microbial Metabolite Sensors to Shape Host Health: Pharmacophysiological View. Biomedicines 2020, 8, 154. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Tsujikawa, T.; Fujiyama, Y. Role of dietary fiber and short-chain fatty acids in the colon. Curr. Pharm. Des. 2003, 9, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Smith, N.J.; Jenkins, L.; Brown, A.J.; Milligan, G. Conserved polar residues in transmembrane domains V, VI, and VII of free fatty acid receptor 2 and free fatty acid receptor 3 are required for the binding and function of short chain fatty acids. J. Biol. Chem. 2008, 283, 32913–32924. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.D.; Due-Hansen, M.E.; Christiansen, E.; Hansen, A.M.; Mackenzie, A.E.; Murdoch, H.; Pandey, S.K.; Ward, R.J.; Marquez, R.; Tikhonova, I.G.; et al. Defining the molecular basis for the first potent and selective orthosteric agonists of the FFA2 free fatty acid receptor. J. Biol. Chem. 2013, 288, 17296–17312. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.U.; In, H.J.; Kwon, M.S.; Park, B.O.; Jo, M.; Kim, M.O.; Cho, S.; Lee, S.; Lee, H.J.; Kwak, Y.S.; et al. beta-Arrestin 2 mediates G protein-coupled receptor 43 signals to nuclear factor-kappaB. Biol. Pharm. Bull. 2013, 36, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; van Esch, B.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor alpha-Stimulated Endothelial Cells via Activation of GPR41/43 and Inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar]
- Robles-Vera, I.; Toral, M.; de la Visitacion, N.; Aguilera-Sanchez, N.; Redondo, J.M.; Duarte, J. Protective Effects of Short-Chain Fatty Acids on Endothelial Dysfunction Induced by Angiotensin II. Front. Physiol. 2020, 11, 277. [Google Scholar] [CrossRef] [PubMed]
- Dinakis, E.; Nakai, M.; Gill, P.A.; Yiallourou, S.; Sata, Y.; Muir, J.; Carrington, M.; Head, G.A.; Kaye, D.M.; Marques, F.Z. The Gut Microbiota and Their Metabolites in Human Arterial Stiffness. Heart Lung Circ. 2021, 30, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Tsujimoto, G.; Kimura, I. Regulation of Energy Homeostasis by GPR41. Front. Endocrinol. 2014, 5, 81. [Google Scholar] [CrossRef]
- Lin, Y.; Lv, Y.; Mao, Z.; Chen, X.; Chen, Y.; Zhu, B.; Yu, Y.; Ding, Z.; Zhou, F. Polysaccharides from Tetrastigma Hemsleyanum Diels et Gilg ameliorated inflammatory bowel disease by rebuilding the intestinal mucosal barrier and inhibiting inflammation through the SCFA-GPR41/43 signaling pathway. Int. J. Biol. Macromol. 2023, 250, 126167. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, M.; Zhu, Y.; Qin, L. The Interplay of Dietary Fibers and Intestinal Microbiota Affects Type 2 Diabetes by Generating Short-Chain Fatty Acids. Foods 2023, 12, 1023. [Google Scholar] [CrossRef] [PubMed]
- Bolognini, D.; Tobin, A.B.; Milligan, G.; Moss, C.E. The Pharmacology and Function of Receptors for Short-Chain Fatty Acids. Mol. Pharmacol. 2016, 89, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Vogler, O.; Barcelo, J.M.; Ribas, C.; Escriba, P.V. Membrane interactions of G proteins and other related proteins. Biochim. Biophys. Acta 2008, 1778, 1640–1652. [Google Scholar] [CrossRef] [PubMed]
- Ang, Z.; Xiong, D.; Wu, M.; Ding, J.L. FFAR2-FFAR3 receptor heteromerization modulates short-chain fatty acid sensing. FASEB J. 2018, 32, 289–303. [Google Scholar] [CrossRef]
- Onyszkiewicz, M.; Gawrys-Kopczynska, M.; Konopelski, P.; Aleksandrowicz, M.; Sawicka, A.; Kozniewska, E.; Samborowska, E.; Ufnal, M. Butyric acid, a gut bacteria metabolite, lowers arterial blood pressure via colon-vagus nerve signaling and GPR41/43 receptors. Pflugers Arch. 2019, 471, 1441–1453. [Google Scholar] [CrossRef] [PubMed]
- Isenovic, E.R.; Soskic, S.; Dungen, H.D.; Dobutovic, B.; Elvis, T.; Simone, I.; Marche, P. Regulation of Endothelial Nitric Oxide Synthase in Pathophysiological Conditions. Cardiovasc. Hematol. Disord. Drug Targets 2011, 11, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Nooromid, M.; Chen, E.B.; Xiong, L.; Shapiro, K.; Jiang, Q.; Demsas, F.; Eskandari, M.; Priyadarshini, M.; Chang, E.B.; Layden, B.T.; et al. Microbe-Derived Butyrate and Its Receptor, Free Fatty Acid Receptor 3, But Not Free Fatty Acid Receptor 2, Mitigate Neointimal Hyperplasia Susceptibility After Arterial Injury. J. Am. Heart Assoc. 2020, 9, e016235. [Google Scholar] [CrossRef]
- Colina, C.; Puhl, H.L., 3rd; Ikeda, S.R. Selective tracking of FFAR3-expressing neurons supports receptor coupling to N-type calcium channels in mouse sympathetic neurons. Sci. Rep. 2018, 8, 17379. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Kuwahara, K. Sodium-Glucose Cotransporter-2 inhibitors are potential therapeutic agents for treatment of non-diabetic heart failure patients. J. Cardiol. 2020, 76, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lymperopoulos, A.; Borges, J.I.; Cora, N.; Sizova, A. Sympatholytic Mechanisms for the Beneficial Cardiovascular Effects of SGLT2 Inhibitors: A Research Hypothesis for Dapagliflozin’s Effects in the Adrenal Gland. Int. J. Mol. Sci. 2021, 22, 7684. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef] [PubMed]
- Onyszkiewicz, M.; Gawrys-Kopczynska, M.; Salagaj, M.; Aleksandrowicz, M.; Sawicka, A.; Kozniewska, E.; Samborowska, E.; Ufnal, M. Valeric acid lowers arterial blood pressure in rats. Eur. J. Pharmacol. 2020, 877, 173086. [Google Scholar] [CrossRef] [PubMed]
- Whelton, S.P.; Hyre, A.D.; Pedersen, B.; Yi, Y.; Whelton, P.K.; He, J. Effect of dietary fiber intake on blood pressure: A meta-analysis of randomized, controlled clinical trials. J. Hypertens. 2005, 23, 475–481. [Google Scholar] [CrossRef]
- Holmes, E.; Loo, R.L.; Stamler, J.; Bictash, M.; Yap, I.K.; Chan, Q.; Ebbels, T.; De Iorio, M.; Brown, I.J.; Veselkov, K.A.; et al. Human metabolic phenotype diversity and its association with diet and blood pressure. Nature 2008, 453, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Dardi, P.; Dos Santos-Eichler, R.A.; de Oliveira, S.; Vinolo, M.A.R.; Camara, N.O.S.; Rossoni, L.V. Reduced intestinal butyrate availability is associated with the vascular remodeling in resistance arteries of hypertensive rats. Front. Physiol. 2022, 13, 998362. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.D.; Shimpukade, B.; Mackenzie, A.E.; Butcher, A.J.; Pediani, J.D.; Christiansen, E.; Heathcote, H.; Tobin, A.B.; Ulven, T.; Milligan, G. The pharmacology of TUG-891, a potent and selective agonist of the free fatty acid receptor 4 (FFA4/GPR120), demonstrates both potential opportunity and possible challenges to therapeutic agonism. Mol. Pharmacol. 2013, 84, 710–725. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Hirasawa, A.; Sun, Q.; Sadakane, K.; Itsubo, C.; Iga, T.; Adachi, T.; Koshimizu, T.A.; Hashimoto, T.; Asakawa, Y.; et al. Novel selective ligands for free fatty acid receptors GPR120 and GPR40. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 380, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hasegawa, S.; Kasubuchi, M.; Kimura, I. Free fatty acid receptors as therapeutic targets for the treatment of diabetes. Front. Pharmacol. 2014, 5, 236. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.J.; Brown, A.J.; Holliday, N.D. Differential signaling by splice variants of the human free fatty acid receptor GPR120. Mol. Pharmacol. 2012, 81, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Park, B.O.; Kim, S.H.; Kong, G.Y.; Kim, D.H.; Kwon, M.S.; Lee, S.U.; Kim, M.O.; Cho, S.; Lee, S.; Lee, H.J.; et al. Selective novel inverse agonists for human GPR43 augment GLP-1 secretion. Eur. J. Pharmacol. 2016, 771, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kiepura, A.; Stachyra, K.; Olszanecki, R. Anti-Atherosclerotic Potential of Free Fatty Acid Receptor 4 (FFAR4). Biomedicines 2021, 9, 467. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.L.; Abd El-Aty, A.M.; Jeong, J.H.; Jung, T.W. Ginsenoside Rb2 Ameliorates LPS-Induced Inflammation and ER Stress in HUVECs and THP-1 Cells via the AMPK-Mediated Pathway. Am. J. Chin. Med. 2020, 48, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Jung, T.W.; Kim, J.W.; Kim, J.A.; Lee, Y.B.; Hong, S.H.; Roh, E.; Choi, K.M.; Baik, S.H.; Yoo, H.J. Protectin DX prevents H2O2-mediated oxidative stress in vascular endothelial cells via an AMPK-dependent mechanism. Cell Signal 2019, 53, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Qiu, S. Activation of GPR120 promotes the metastasis of breast cancer through the PI3K/Akt/NF-kappaB signaling pathway. Anticancer Drugs 2019, 30, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.Y.; Lii, C.K.; Ye, S.Y.; Li, C.C.; Lu, C.Y.; Lin, A.H.; Liu, K.L.; Chen, H.W. Docosahexaenoic acid inhibits vascular endothelial growth factor (VEGF)-induced cell migration via the GPR120/PP2A/ERK1/2/eNOS signaling pathway in human umbilical vein endothelial cells. J. Agric. Food Chem. 2014, 62, 4152–4158. [Google Scholar] [CrossRef] [PubMed]
- Kamata, R.; Bumdelger, B.; Kokubo, H.; Fujii, M.; Yoshimura, K.; Ishida, T.; Ishida, M.; Yoshizumi, M. EPA Prevents the Development of Abdominal Aortic Aneurysms through Gpr-120/Ffar-4. PLoS ONE 2016, 11, e0165132. [Google Scholar] [CrossRef] [PubMed]
- Abekura, Y.; Ono, I.; Kawashima, A.; Takizawa, K.; Koseki, H.; Miyata, H.; Shimizu, K.; Oka, M.; Kushamae, M.; Miyamoto, S.; et al. Eicosapentaenoic acid prevents the progression of intracranial aneurysms in rats. J. Neuroinflamm. 2020, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Tang, C.; Li, H.; Lei, J.; Zhu, L.; Kou, L.; Li, H.; Luo, S.; Li, C.; Chen, W.; et al. Eicosapentaenoic acid prevents inflammation induced by acute cerebral infarction through inhibition of NLRP3 inflammasome activation. Life Sci. 2020, 242, 117133. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Miura, D.; Saito, Y.; Yunoki, K.; Koyama, Y.; Satoh, M.; Kondo, M.; Osawa, K.; Hatipoglu, O.F.; Miyoshi, T.; et al. Eicosapentaenoic acid prevents arterial calcification in klotho mutant mice. PLoS ONE 2017, 12, e0181009. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.H.; Liu, H.W.; Tian, X.X.; Li, J.; Ding, Y.; Li, Y.; Mei, Z.; Zou, M.H.; Han, Y.L. AMPKalpha2 controls the anti-atherosclerotic effects of fish oils by modulating the SUMOylation of GPR120. Nat. Commun. 2022, 13, 7721. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).