
Flavonoids-Enriched Vegetal Extract Prevents the Activation of NFκB Downstream Mechanisms in a Bowel Disease In Vitro Model

 , , , , ,
, , , , ,  ,
,  and
and
Abstract

1. Introduction
2. Results and Discussion
2.1. Evaluation of Digested ACF Blend Extract’s Effects on Both Enterocyte Viability and Paracellular Permeability in the IBD Intestinal Barrier Model
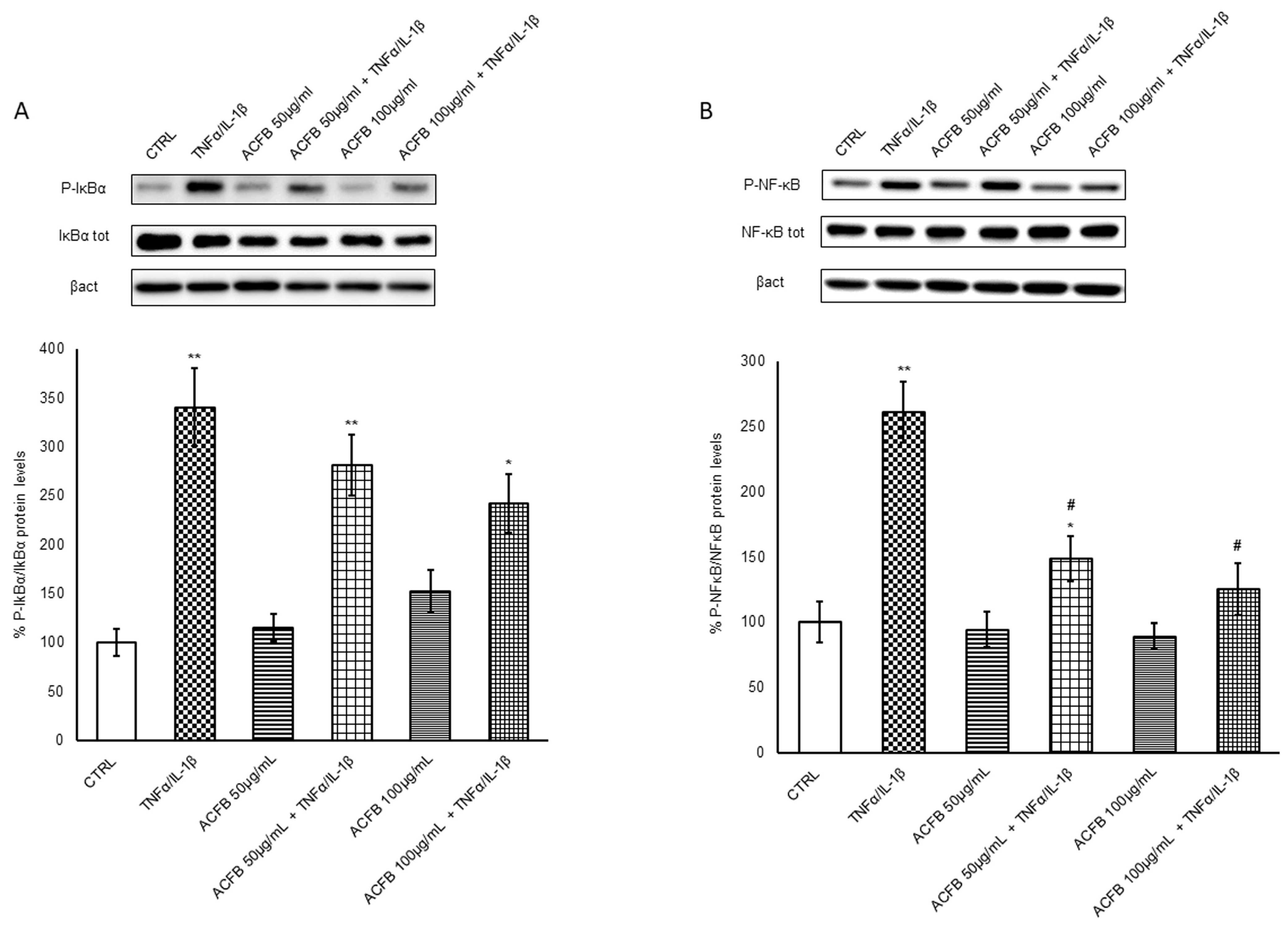
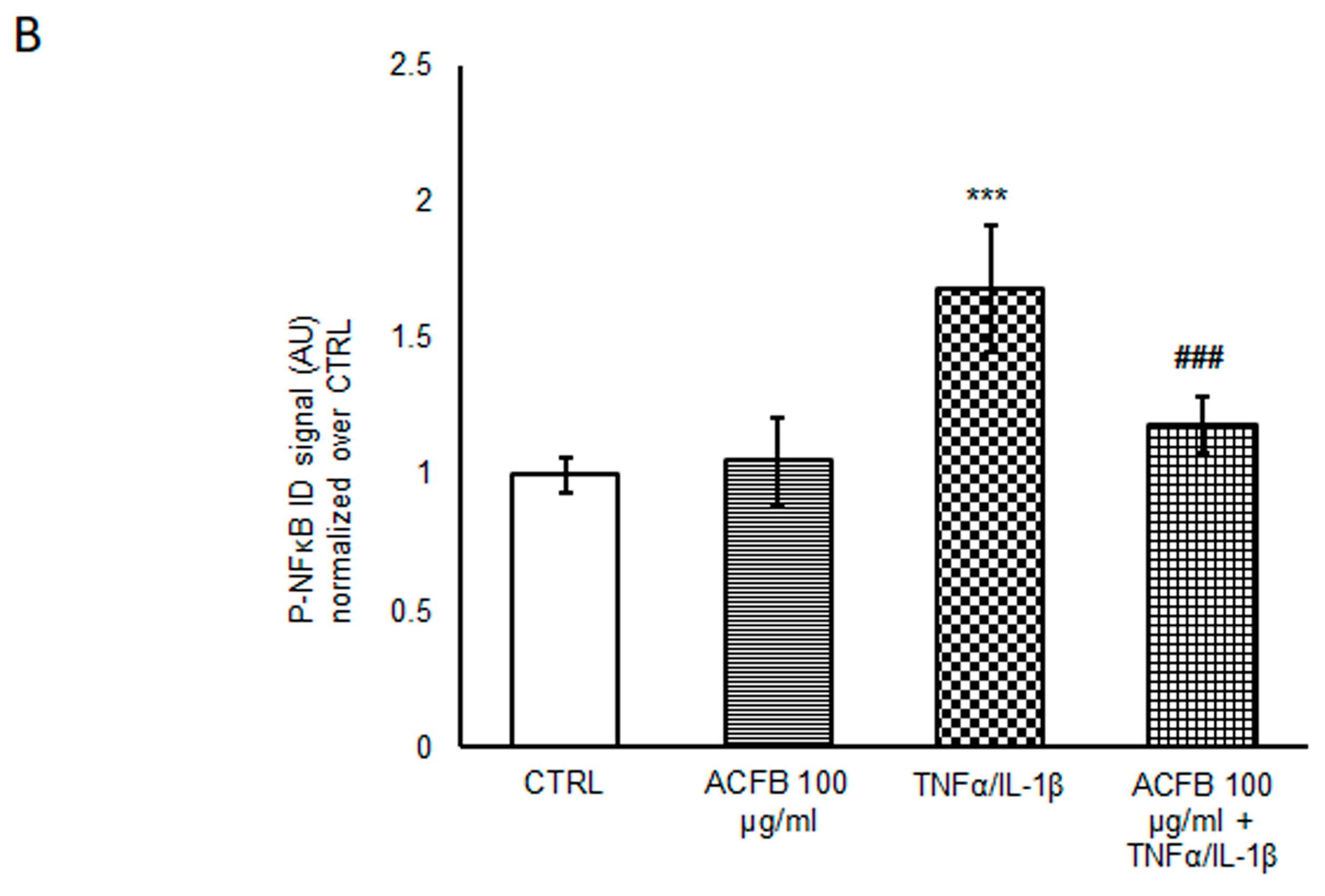
2.2. Effects of ACFB Digested Extract on NF-κB p65 Activation
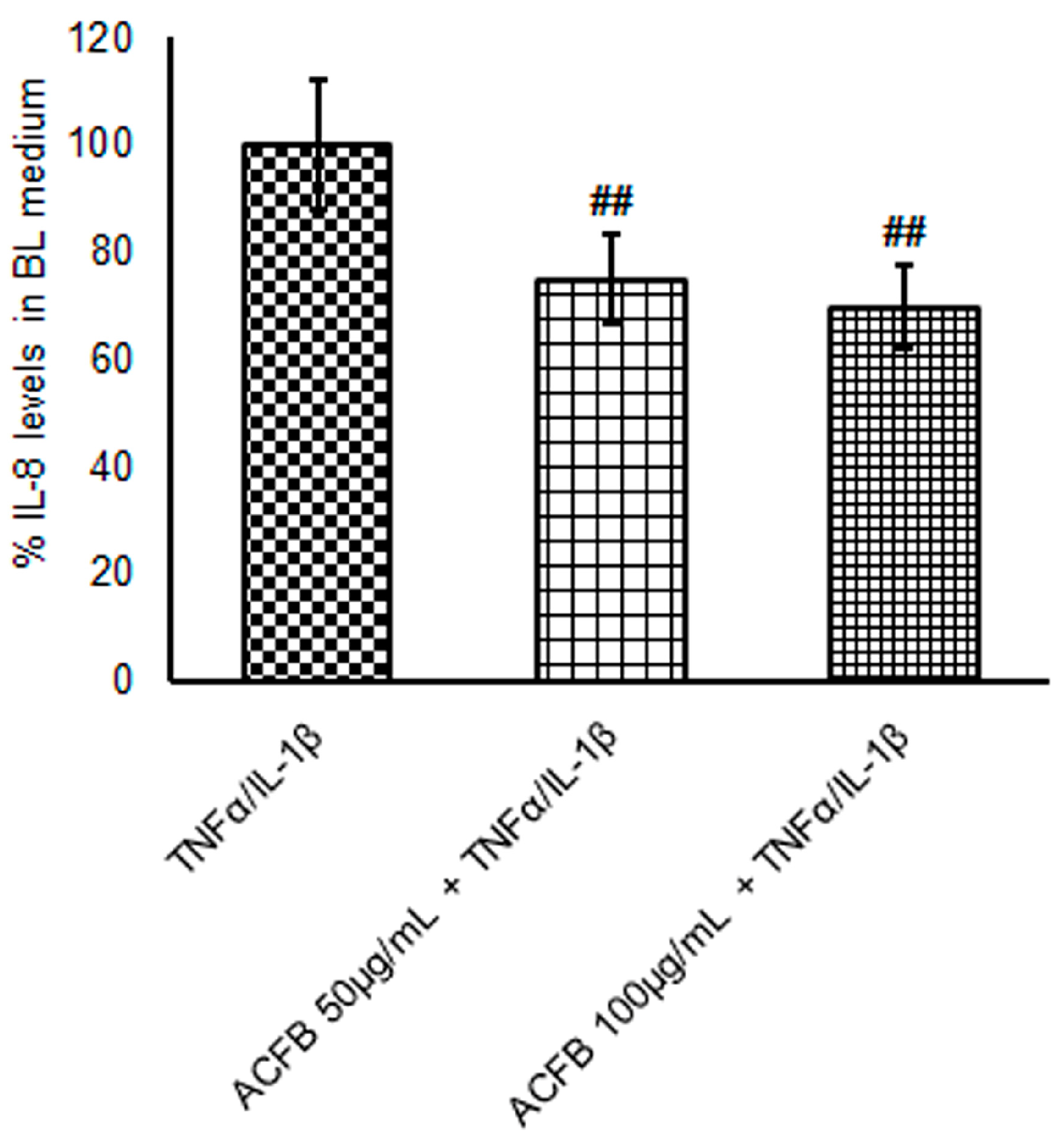
2.3. ACFB-Digested Extract Prevents IL-8 Secretion Highly Induced by TNF-α/IL-1β Exposure
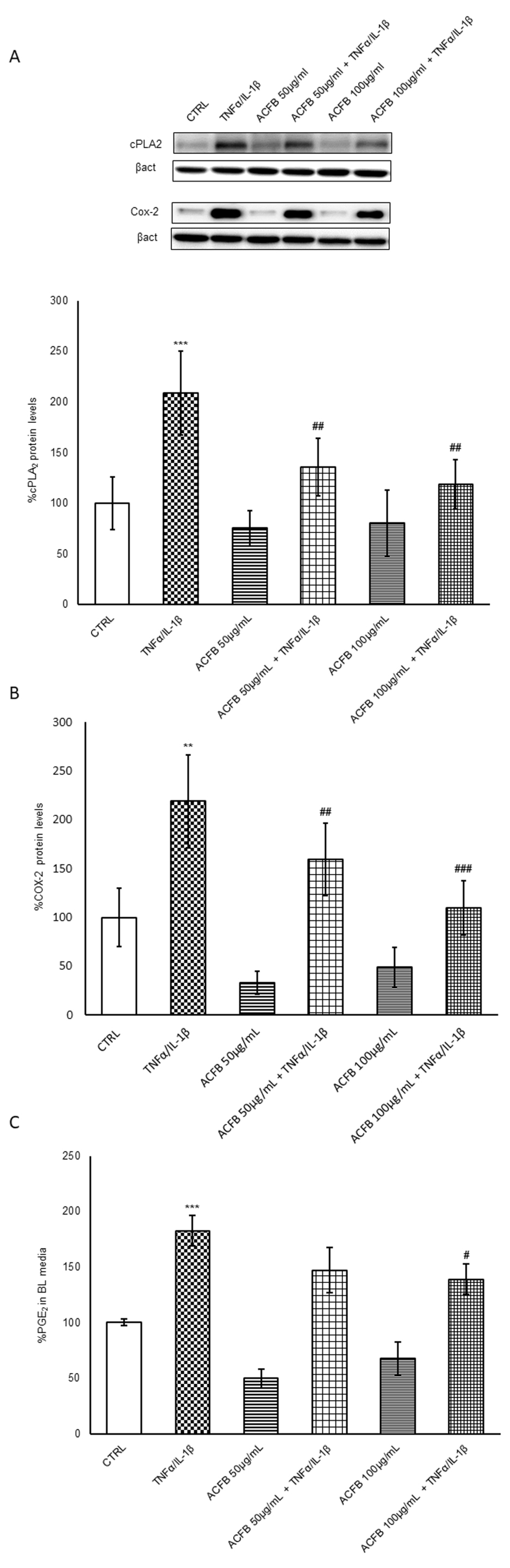
2.4. ACFB-Digested Extract Prevents the Cox-2/PGE2 Pathway Induced by TNF-α/IL-1β Exposure
3. Materials and Methods
3.1. Vegetal-Extract Selection for ACFB Formulation
3.2. UHPLC-DAD-HRMS/MS Profiles of ACFB
3.3. INFOGEST Methods for Digestion In Vitro Simulation
3.4. Cell Cultures
3.5. ACFB Treatment and Cell Viability Evaluation
3.6. Inflammatory Treatment and Evaluation of Intestinal Barrier Integrity
3.7. Electrophoresis and Immunoblotting
3.8. IL-8 ELISA Assay
3.9. Immunofluorescence Assay
3.10. PGE2 ELISA Assay
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.Y.; Liu, H.; Wang, M.C. Complementary and alternative medicine mention and recommendations in inflammatory bowel disease guidelines: Systematic review and assessment using AGREE II. BMC Complement. Med. Ther. 2023, 23, 230. [Google Scholar] [CrossRef] [PubMed]
- Villanacci, V.; Del Sordo, R.; Parigi, T.L.; Leoncini, G.; Bassotti, G. Inflammatory Bowel Diseases: Does One Histological Score Fit All? Diagnostics 2023, 13, 2112. [Google Scholar] [CrossRef] [PubMed]
- Pariente, B.; Cosnes, J.; Danese, S.; Sandborn, W.J.; Lewin, M.; Fletcher, J.G.; Chowers, Y.; D’Haens, G.; Feagan, B.G.; Hibi, T.; et al. Development of the Crohn’s disease digestive damage score, the Lémann score. Inflamm. Bowel Dis. 2011, 17, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Dhawan, P.; Srivastava, A.S.; Singh, A.B. Inflammatory bowel disease: Therapeutic limitations and prospective of the stem cell therapy. World J. Stem Cells 2020, 12, 1050–1066. [Google Scholar] [CrossRef] [PubMed]
- Imbrizi, M.; Magro, F.; Coy, C.S.R. Pharmacological Therapy in Inflammatory Bowel Diseases: A Narrative Review of the Past 90 Years. Pharmaceuticals 2023, 16, 1272. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. What Is Inflammatory Bowel Disease (IBD)?|IBD. 2022. Available online: https://www.cdc.gov/ibd/what-is-IBD.htm (accessed on 25 January 2024).
- Lin, S.C.; Cheifetz, A.S. The use of complementary and alternative medicine in patients with inflammatory bowel disease. Gastroenterol. Hepatol. 2018, 14, 415–425. [Google Scholar]
- Liu, F.; Li, D.; Wang, X.; Cui, Y.; Li, X. Polyphenols intervention is an effective strategy to ameliorate inflammatory bowel disease: A systematic review and metaanalysis. Int. J. Food Sci. Nutr. 2021, 72, 14–25. [Google Scholar] [CrossRef]
- Tursi, A.; Brandimarte, G.; Papa, A.; Giglio, A.; Elisei, W.; Giorgetti, G.M.; Forti, G.; Morini, S.; Hassan, C.; Pistoia, M.A.; et al. Treatment of relapsing mild-to-moderate ulcerative colitis with the probiotic VSL#3 as adjunctive to a standard pharmaceutical treatment: A double-blind, randomized, placebo-controlled study. Am. J. Gastroenterol. 2010, 105, 2218–2227. [Google Scholar] [CrossRef]
- Becker, C.; Neurath, M.F.; Wirtz, S. The Intestinal Microbiota in Inflammatory Bowel Disease. ILAR J. 2015, 56, 192–204. [Google Scholar] [CrossRef]
- Guerreiro, C.S.; Calado, Â.; Sousa, J.; Fonseca, J.E. Diet, Microbiota, and Gut PermeabilityThe Unknown Triad in Rheumatoid arthrites. Front. Med. 2018, 14, 349. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases-Promises, Perspectives, and Pitfalls. Oxid. Med. Cell Longev. 2016, 2016, 9346470. [Google Scholar] [CrossRef] [PubMed]
- Salaritabar, A.; Darvishi, B.; Hadjiakhoondi, F.; Manayi, A.; Sureda, A.; Nabavi, S.F.; Fitzpatrick, L.R.; Nabavi, S.M.; Bishayee, A. Therapeutic potential of flavonoids in inflammatory bowel disease: A comprehensive review. World J. Gastroenterol. 2017, 23, 5097–5114. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, S.; Del Bo’, C.; Marino, M.; Gargari, G.; Cherubini, A.; Andrés-Lacueva, C.; Hidalgo-Liberona, N.; Peron, G.; González-Dominguez, R.; Kroon, P.; et al. Polyphenols and Intestinal Permeability: Rationale and Future Perspectives. J. Agric. Food Chem. 2020, 68, 1816–1829. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Almeida, L.; Barbosa, R.M.; Laranjinha, J. Luteolin suppresses the JAK/STAT pathway in a cellular model of intestinal inflammation. Food Funct. 2017, 8, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Ghasemi-Dehnoo, M.; Lorigooini, Z.; Amini-Khoei, H.; Sabzevary-Ghahfarokhi, M.; Rafieian-Kopaei, M. Quinic acid ameliorates ulcerative colitis in rats, through the inhibition of two TLR4-NF-κB and NF-κB-INOS-NO signaling pathways. Immun. Inflamm. Dis. 2023, 11, e926. [Google Scholar] [CrossRef]
- Song, H.Y.; Sik, K.W.; Kim, J.M.; Bak, D.H.; Moo Han, J.; Lim, S.T.; Byun, E.B. A hydroxyethyl derivative of chrysin exhibits anti-inflammatory activity in dendritic cells and protective effects against dextran sodium salt-induced colitis in mice. Int. Immunopharmacol. 2019, 77, 105958. [Google Scholar] [CrossRef]
- Ai, X.Y.; Qin, Y.; Liu, H.J.; Cui, Z.H.; Li, M.; Yang, J.H.; Zhong, W.L.; Liu, Y.R.; Chen, S.; Sun, T.; et al. Apigenin inhibits colonic inflammation and tumorigenesis by suppressing STAT3-NF-κB signaling. Oncotarget 2017, 8, 100216–100226. [Google Scholar] [CrossRef]
- Frigerio, J.; Tedesco, E.; Benetti, F.; Insolia, V.; Nicotra, G.; Mezzasalma, V.; Pagliari, S.; Labra, M.; Campone, L. Anticholesterolemic Activity of Three Vegetal Extracts (Artichoke, Caigua, and Fenugreek) and Their Unique Blend. Front. Pharmacol. 2021, 12, 726199. [Google Scholar] [CrossRef] [PubMed]
- Pagliari, S.; Forcella, M.; Lonati, E.; Sacco, G.; Romaniello, F.; Rovellini, P.; Fusi, P.; Palestini, P.; Campone, L.; Labra, M.; et al. Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum verum J. Presl) Bark Extract after In Vitro Digestion Simulation. Foods 2023, 12, 452. [Google Scholar] [CrossRef]
- Luettig, J.; Rosenthal, R.; Barmeyer, C.; Schulzke, J.D. Claudin-2 as a mediator of leaky gut barrier during intestinal inflammation. Tissue Barriers 2015, 3, e977176. [Google Scholar] [CrossRef]
- Stolfi, C.; Maresca, C.; Monteleone, G.; Laudisi, F. Implication of Intestinal Barrier Dysfunction in Gut Dysbiosis and Diseases. Biomedicines 2022, 10, 289. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, J.Y. Cinnamon Subcritical Water Extract Attenuates Intestinal Inflammation and Enhances Intestinal Tight Junction in a Caco-2 and RAW264.7 Co-Culture Model. Food Funct. 2019, 10, 4350–4360. [Google Scholar] [CrossRef]
- Lu, Q.; Xie, Y.; Luo, J.; Gong, Q.; Li, C. Natural flavones from edible and medicinal plants exhibit enormous potential to treat ulcerative colitis. Front. Pharmacol. 2023, 14, 1168990. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Chlorogenic Acid (CGA) Isomers Alleviate Interleukin 8 (IL-8) Production in Caco-2 Cells by Decreasing Phosphorylation of p38 and Increasing Cell Integrity. Int. J. Mol. Sci. 2018, 19, 3873. [Google Scholar] [CrossRef]
- Wan, P.; Xie, M.; Chen, G.; Dai, Z.; Hu, B.; Zeng, X.; Sun, Y. Anti-inflammatory effects of dicaffeoylquinic acids from Ilex kudingcha on lipopolysaccharide-treated RAW264.7 macrophages and potential mechanisms. Food Chem. Toxicol. 2019, 126, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; Hu, S.; Spekhorst, L.M.; Zhernakova, D.V.; Vila, A.V.; Li, Y.; Voskuil, M.D.; Van Berkel, L.A.; Folly, B.B.; Charrout, M.; et al. The Effect of Phenotype and Genotype on the Plasma Proteome in Patients with Inflammatory Bowel Disease. J. Crohn’s Colitis 2022, 16, 414–429. [Google Scholar] [CrossRef]
- Scoville, E.A.; Allaman, M.M.; Adams, D.W.; Motley, A.K.; Peyton, S.C.; Ferguson, S.L.; Horst, S.N.; Williams, C.S.; Beaulieu, D.B.; Schwartz, D.A.; et al. Serum Polyunsaturated Fatty Acids Correlate with Serum Cytokines and Clinical Disease Activity in Crohn’s Disease. Sci. Rep. 2019, 9, 2882. [Google Scholar] [CrossRef]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Ilies, M.A. The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. Int. J. Mol. Sci. 2023, 24, 1353. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L. Eicosanoids in the Gastrointestinal Tract. Br. J. Pharmacol. 2019, 176, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Kikut, J.; Mokrzycka, M.; Drozd, A.; Grzybowska-Chlebowczyk, U.; Ziętek, M.; Szczuko, M. Involvement of Proinflammatory Arachidonic Acid (ARA) Derivatives in Crohn’s Disease (CD) and Ulcerative Colitis (UC). J. Clin. Med. 2022, 11, 1861. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.K.; Qian, Y.; Leonard, S.S.; Sbarra, D.C.; Shi, X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly in-hibit prostaglandin-E2 formation in RAW 264.7 cells. J. Nutr. 2006, 136, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Min, S.Y.; Yu, H.W.; Kim, K.; Kim, S.; Lee, H.J.; Kim, J.H.; Park, Y.J. Effects of Apigenin on RBL-2H3, RAW264.7, and HaCaT Cells: Anti-Allergic, Anti-Inflammatory, and Skin-Protective Activities. Int. J. Mol. Sci. 2020, 21, 4620. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Static in Vitro Digestion Method Suitable for Food—An International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Lonati, E.; Sala, G.; Corbetta, P.; Pagliari, S.; Cazzaniga, E.; Botto, L.; Rovellini, P.; Bruni, I.; Palestini, P.; Bulbarelli, A. Digested Cinnamon (Cinnamomum verum J. Presl) Bark Extract Modulates Claudin-2 Gene Expression and Protein Levels under TNFα/IL-1β Inflammatory Stimulus. Int. J. Mol. Sci. 2023, 24, 9201. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Vadillo, C.; Nau, F.; Dubiard, C.G.; Cheynier, V.; Meudec, E.; Sanz-Buenhombre, M.; Guadarrama, A.; Tóth, T.; Csavajda, É.; Hingyi, H.; et al. In Vitro Digestion of Dairy and Egg Products Enriched with Grape Extracts: Effect of the Food Matrix on Polyphenol Bioaccessibility and Antioxidant Activity. Food Res. Int. 2016, 88, 284–292. [Google Scholar] [CrossRef]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar] [CrossRef]
- Cimarosti, H.; Ashikaga, E.; Jaafari, N.; Dearden, L.; Rubin, P.; Wilkinson, K.A.; Henley, J.M. Enhanced SUMOylation and SENP-1 Protein Levels Following Oxygen and Glucose Deprivation in Neurones. J. Cereb. Blood Flow Metab. 2012, 32, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chigurupati, S.; Arumugam, T.V.; Jo, D.G.; Li, H.; Chan, S.L. Notch Activation Enhances the Microglia-Mediated Inflammatory Response Associated With Focal Cerebral Ischemia. Stroke 2011, 42, 2589–2594. [Google Scholar] [CrossRef] [PubMed]
- Farré, R.; Fiorani, M.; Abdu Rahiman, S.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | [M-H]- | Formula | Δppm | MS/MS | Compounds * |
|---|---|---|---|---|---|---|
| 1 | 2.26 | 353.0881 | C16H17O9 | 2.3 | 191.0551 | Mono-caffeoylquinic acid isomer |
| 2 | 3.27 | 353.0881 | C16H17O9 | 2.3 | 191.0551 | Mono-caffeoylquinic acid isomer |
| 3 | 3.39 | 353.0881 | C16H17O9 | 2.3 | 191.0551 | Mono-caffeoylquinic acid isomer |
| 4 | 4.68 | 593.1505 | C27H29O15 | −0.2 | 503.1175/473.1074/383.0759/353.0652 | Apigenin-di-C-hexoside isomer |
| 5 | 4.86 | 593.1505 | C27H29O15 | −0.2 | 503.1175/473.1074/383.0759/353.0652 | Apigenin-di-C-hexoside isomer |
| 6 | 5.34 | 563.1412 | C26H27O14 | 2.0 | 503.1175/473.1074/443.0973/383.0759/353.0652 | Apigenin C-hexoside-C-pentoside isomer |
| 7 | 5.57 | 515.1169 | C25H23O12 | −4.1 | 353.0880/191.0550/179.0332 | Di-caffeoylquinic acid isomer |
| 8 | 5.91 | 563.1412 | C26H27O14 | 2.0 | 503.1175/473.1074/443.0973/383.0759/353.0652 | Apigenin C-hexoside-C-pentoside isomer |
| 9 | 6.00 | 447.0933 | C21H19O11 | 1.3 | 357.0606/327.0497 | orientin |
| 10 | 6.79 | 431.0986 | C21H19O10 | 1.9 | 341.0652/311.0551/283.0601 | Apigenin-6-C-glucoside |
| 11 | 7.70 | 447.0930 | C21H19O11 | 0.7 | 285.0389 | cynaroside |
| 12 | 7.81 | 461.0720 | C21H17O12 | 0.0 | 285.0389 | Luteolin-7-O-glucuronide |
| 13 | 8.61 | 515.1169 | C25H23O12 | −4.1 | 353.0880/191.0550/179.0332 | Di-caffeoylquinic acid isomer |
| 14 | 8.98 | 431.0986 | C21H19O10 | 1.9 | 269.0424 | Apigenin-7-O-glucoside |
| 15 | 9.17 | 445.0778 | C21H17O11 | 1.6 | 269.0424 | Apigenin-7-O-glucoronide |
| 16 | 10.56 | 399.1081 | C21H19O8 | 0.3 | 325.0703/295.0604/267.0655 | Chrysin-6-C-fucopyranoside |
| 17 | 11.02 | 415.1035 | C21H19O9 | 1.4 | 325.0703/295.0604/267.0655 | Chrysin derivate |
| 18 | 12.17 | 399.1981 | C21H19O8 | 0.3 | 325.0703/295.0604/267.0655 | Chrysin-6-C-fucopyranoside |
| CTRL | ACFB 50 µg/mL | ACFB 100 µg/mL | TNFα/IL-1β | ACFB 50 µg/mL + TNFα/IL-1β | ACFB 100 µg/mL + TNFα/IL-1β | |
|---|---|---|---|---|---|---|
| pg/mL | 7.31 ± 3.21 | 4.80 ± 3.21 | 11.77 ± 3.81 | 303.69 ± 21.11 | 271.70 ± 11.72 | 256.36 ± 10.36 |
| vs CTRL | *** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corbetta, P.; Lonati, E.; Pagliari, S.; Mauri, M.; Cazzaniga, E.; Botto, L.; Campone, L.; Palestini, P.; Bulbarelli, A. Flavonoids-Enriched Vegetal Extract Prevents the Activation of NFκB Downstream Mechanisms in a Bowel Disease In Vitro Model. Int. J. Mol. Sci. 2024, 25, 7869. https://doi.org/10.3390/ijms25147869
Corbetta P, Lonati E, Pagliari S, Mauri M, Cazzaniga E, Botto L, Campone L, Palestini P, Bulbarelli A. Flavonoids-Enriched Vegetal Extract Prevents the Activation of NFκB Downstream Mechanisms in a Bowel Disease In Vitro Model. International Journal of Molecular Sciences. 2024; 25(14):7869. https://doi.org/10.3390/ijms25147869
Chicago/Turabian StyleCorbetta, Paolo, Elena Lonati, Stefania Pagliari, Mario Mauri, Emanuela Cazzaniga, Laura Botto, Luca Campone, Paola Palestini, and Alessandra Bulbarelli. 2024. "Flavonoids-Enriched Vegetal Extract Prevents the Activation of NFκB Downstream Mechanisms in a Bowel Disease In Vitro Model" International Journal of Molecular Sciences 25, no. 14: 7869. https://doi.org/10.3390/ijms25147869
APA StyleCorbetta, P., Lonati, E., Pagliari, S., Mauri, M., Cazzaniga, E., Botto, L., Campone, L., Palestini, P., & Bulbarelli, A. (2024). Flavonoids-Enriched Vegetal Extract Prevents the Activation of NFκB Downstream Mechanisms in a Bowel Disease In Vitro Model. International Journal of Molecular Sciences, 25(14), 7869. https://doi.org/10.3390/ijms25147869